1. Introduction
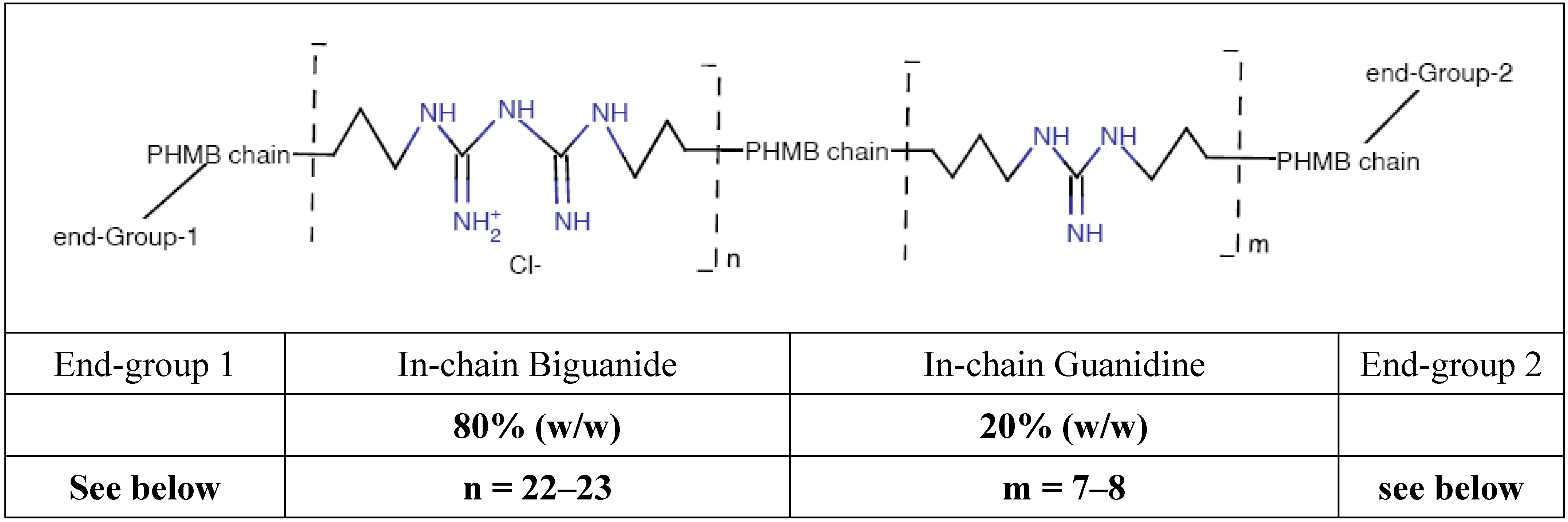
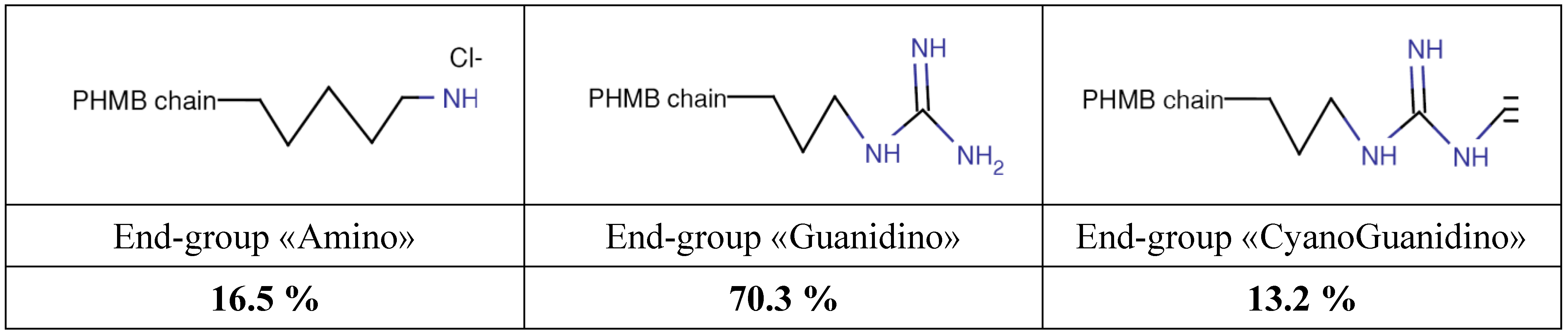
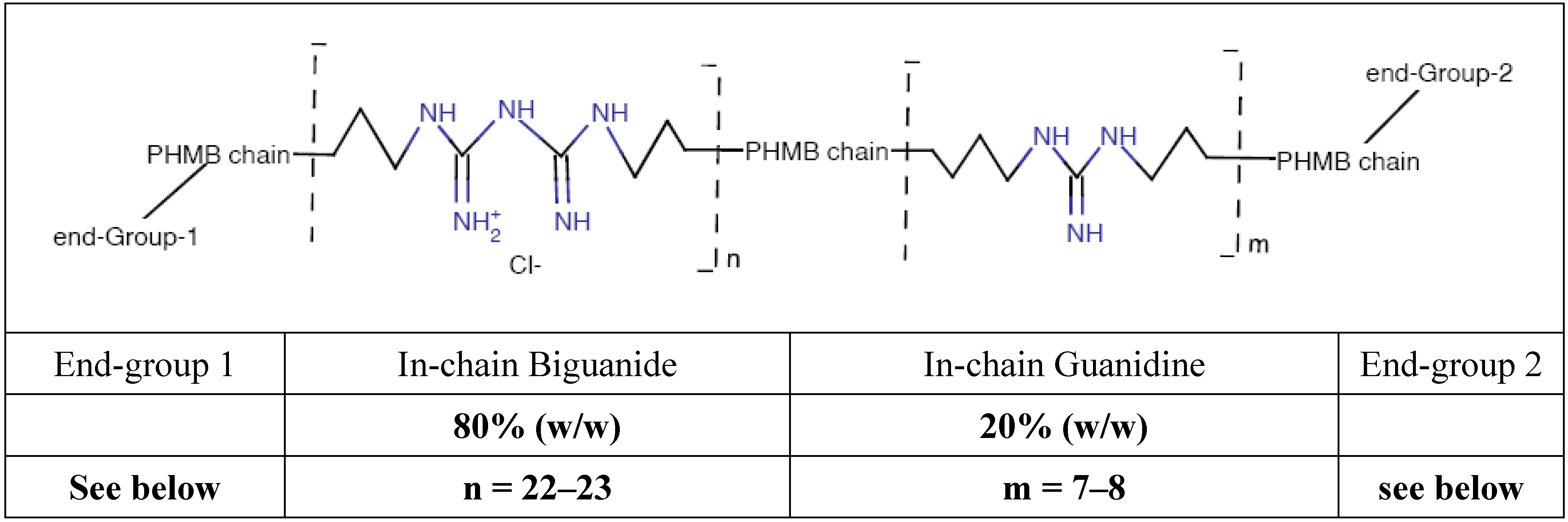
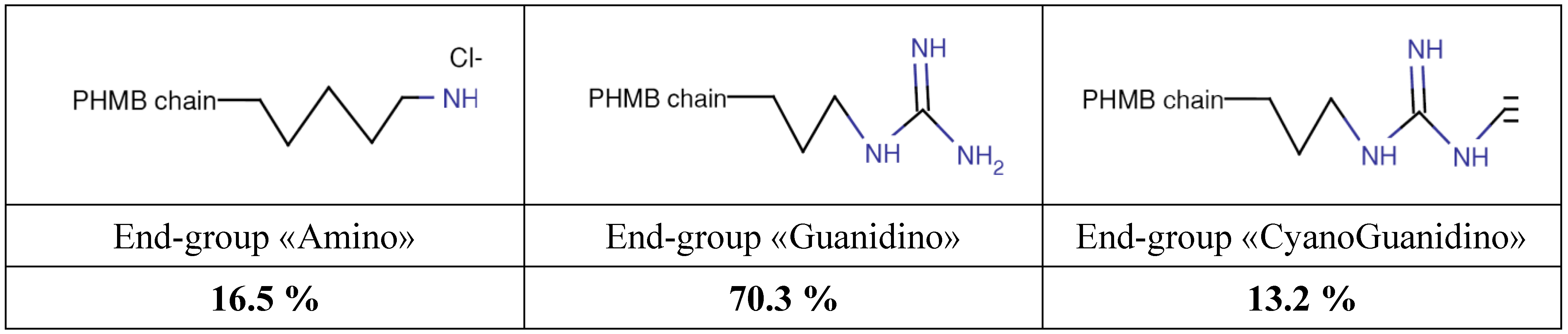
Poly (HexaMethylene Biguanide) hydrochloride (PHMB) (CAS No. 32289-58-0, previously 91403-50-8) belongs to the chemical family of antiseptic biguanides. It has been used since the early nineteen-fifties and has been found to be particularly effective as a hard surface disinfectant, water treatment agent (algaecide and sanitizer, in swimming pools, SPAs and water tanks), preservative agent in several products due to its bacteriostatic or bactericidal properties in a large number of industrial processes and water systems, and as a personal care products preservative. PHMB is effective against pathogens such as bacteria, amoebae and yeast [
1,
2,
3,
4]. Additionally there is an increasing number of reports showing that PHMB has anti-HIV activity [
5,
6,
7]. More interestingly it is active over a wide range of pH (at least from pH 3 to 10), and is temperature (no decomposition until 200–220 °C) and light (including UV) stable. PHMB does not generate chlorinated by-products (CBPs) or trihalomethanes (THMs) found in chlorine-based treatments. For general water treatment, the recommended dose is 20 to 40 mg/L of a 20% PHMB solution (4 to 8 mg/L of active substance) (US-EPA, 2004) [
8]. Compared to chlorine based products, PHMB is not irritating to the eyes or skin, at the recommended doses used in swimming pools.and even in cases of accidental exposure where doses reached 50 and 70 mg/L (10 and 14 mg/L of active substance, respectively) no toxicity was observed [
8].
According to the Biocidal Products Directive (Directive 98/8/EC, also known as BPD), PHMB has been classified as a suspected carcinogen in rodents, based upon three studies conducted during the review of the BPD dossier [
8,
9,
10,
11,
12]. This assessment led to a classification of PHMB as a Carc cat. 2 + H351 (Suspected of causing cancer), category 3; R40 (Limited evidence of carcinogenicity) under the Directive 67/548/EC of the European Parliament and of the Council of 16 February 1998 [
13].
PHMB is not genotoxic [
8,
10,
11,
12], so the present classification raises questions as to possible mechanisms for the rodent carcinogenesis of PHMB. The present experiments are therefore designed to confirm the nongenotoxicity of PHMB and assess its epigenetic effects [
14,
15,
16,
17,
18] including possible nongenotoxic DNA base modifications [
16,
17,
18,
19,
20], mitogenic cytokine production and integrity of the gap junction intercellular communication [
21,
22] using 3 cell types: (1) Caco-2 cells (from a human colon adenocarcinoma) with a non-functional
p53 gene (∆p53: mut p53), (2) N2-A (Neuro-2A cells, mouse neural cells), the brain being a possible target organ in rodents and (3) HepG2 cells (human hepatocellular carcinoma) with functional
p53 gene [
23,
24].
4. Discussion
The existing cytotoxicity data were obtained from studies using either skin painting or administration of PHMB through the diet. The studies mainly focused on PHMB effects on the liver, but also examined the gut and brain since these are the target organs [
8]. For these reasons the 3 cell types, Caco-2 cells (from a human colon adenocarcinoma) with a
p53 non-functional gene (∆p53: mut p53) and Neuro-2A (from a mouse neuroblastoma cells), and HepG2 cells (from a human hepatocellular carcinoma) with functional
p53 genes were selected.
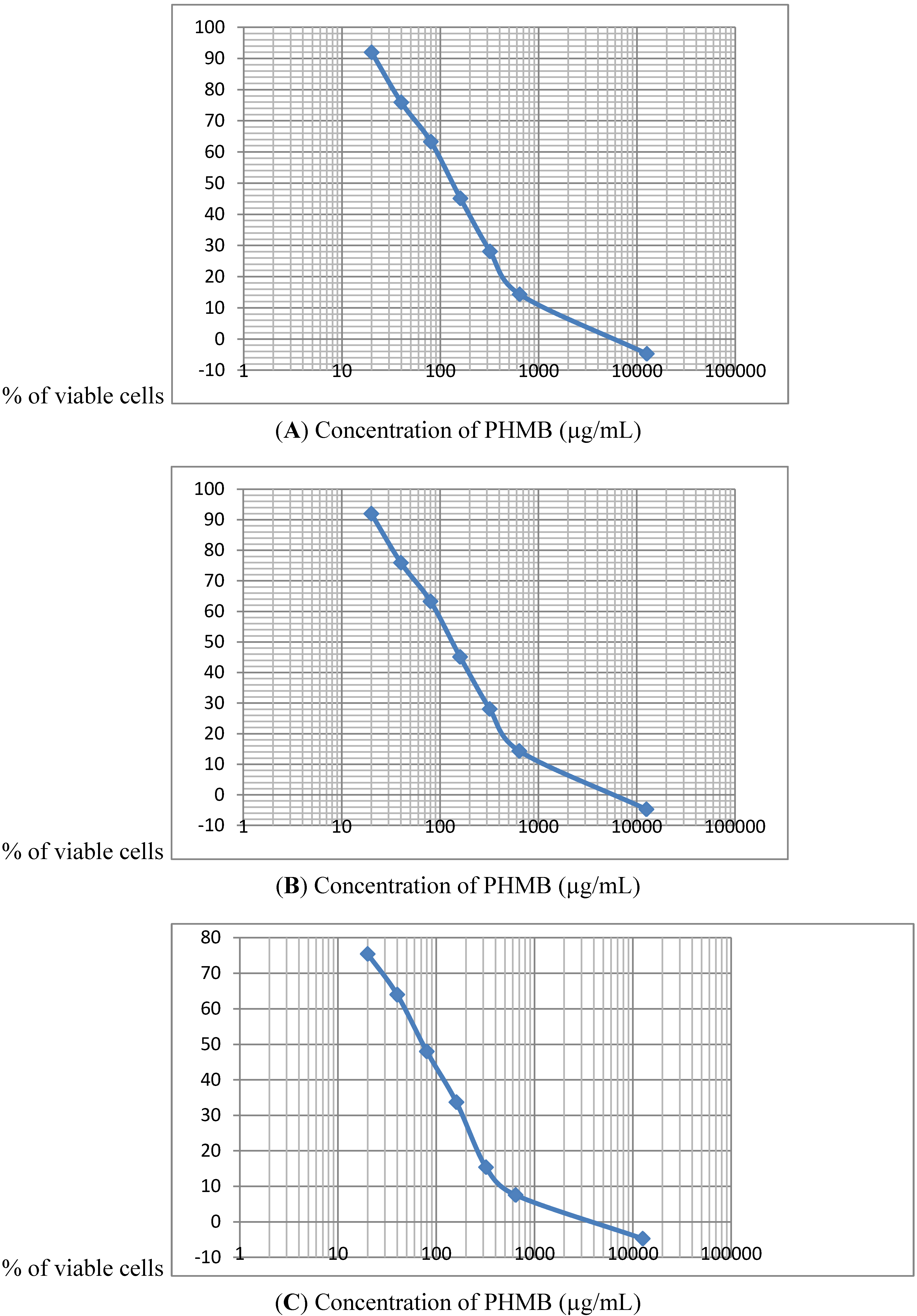
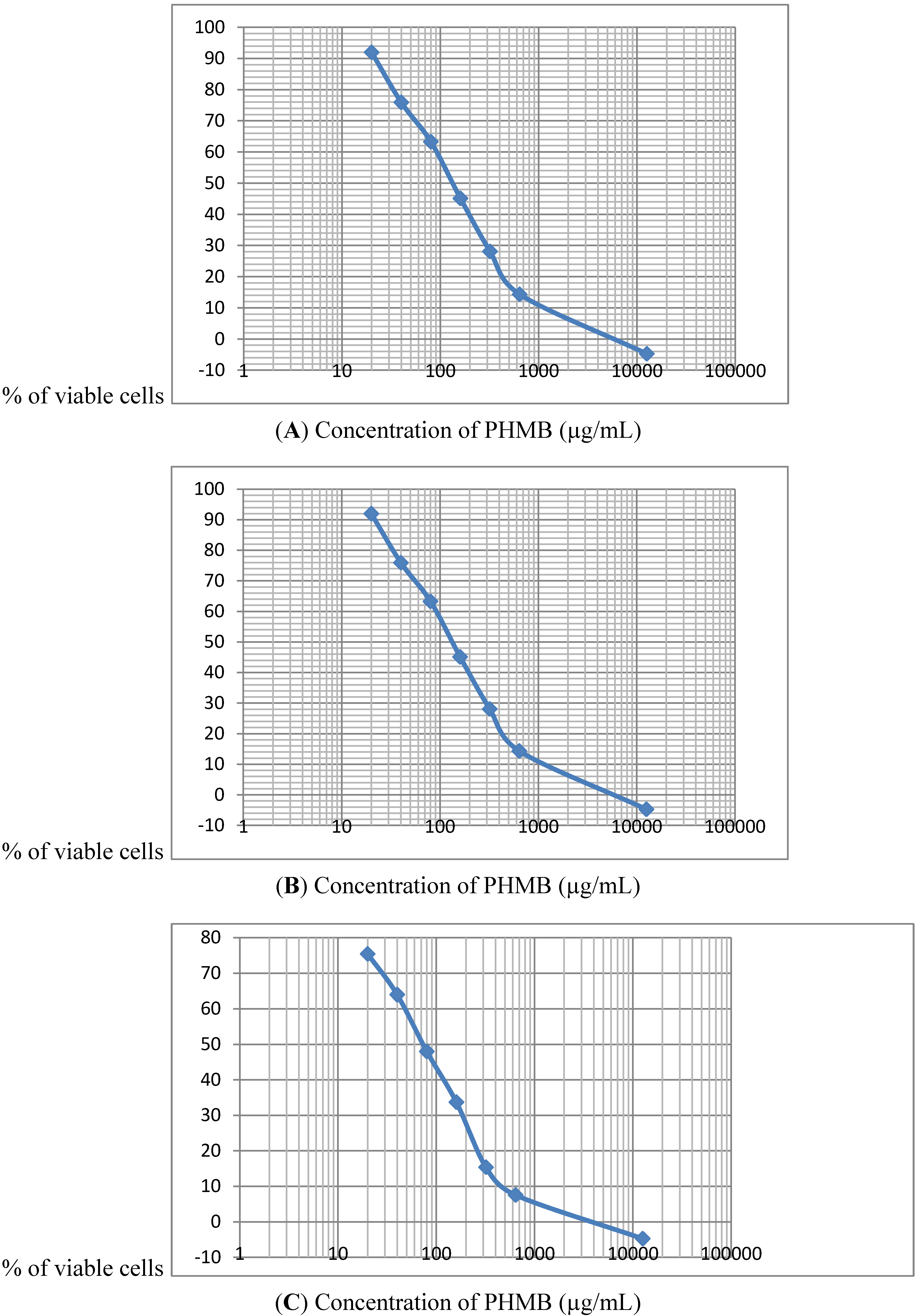
During the determination of the PHMB IC 50% and evaluation of cell viability, two aspects of the effects on mammalian cells were noted. Firstly cytotoxic effects were observed rapidly less than 3 after the beginning or treatment with the human liver cells (HepG2) being the most sensitive. Secondly, when PHMB concentrations were very low, an hormesis-like effect was found, with about a 20% increase in cell proliferation, as reported by Creppy [
35], however the studies reported here focused on the cytotoxic effects of PHMB which occur at much higher concentrations.
As measured by the MTT test, HepG2 cells are the most sensitive to the cytotoxic effects of PHMB among the three cell lines tested. This is in line with the fact that liver is regarded as the main target organ for PHMB in vivo [
8] (US-EPA, 2004). Interestingly the neutral red test showed that the cell lines had a similar sensitivity to PHMB at much lower concentrations than with the MTT assay, thus indicating that the cellular target is the membrane. This is the reason why all of the cells blow up when the respective cytotoxic concentrations for PHMB are reached in the culture medium. It is interesting to note that PHMB has a similar effect on bacteria [
2,
36]. Because of the structural similarities between antimicrobial peptides (AMPs) and PHMB, the latter can be incorporated into bacterial cell membranes and kill bacteria in a manner similar to AMPs [
2]. This mechanism of action is quick and means that bacteria are unlikely to develop resistance to PHMB [
36]. AMPs, which are found in all kingdoms, function primarily by permeabilizing the bacterial membrane. AMPs have several advantages over existing antibiotics including their rapid bactericidal activity, and PHMB bears very similar properties. The little data available for mammalian cells are similar to the present finding [
1,
3].
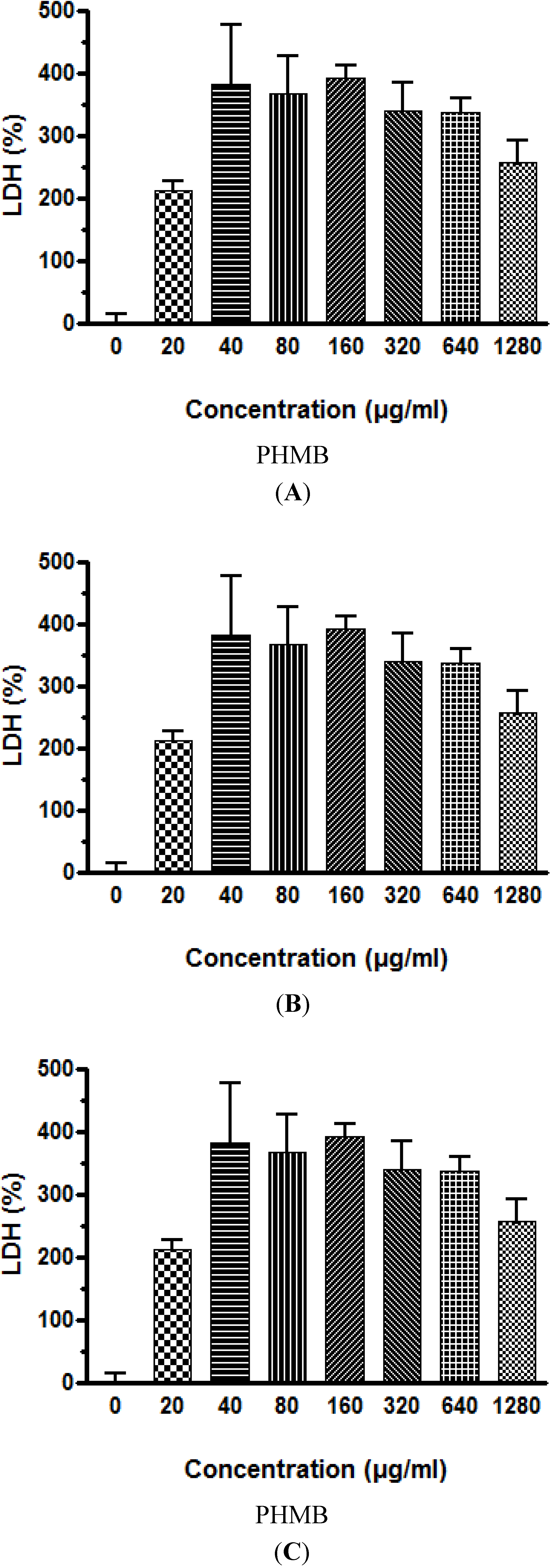
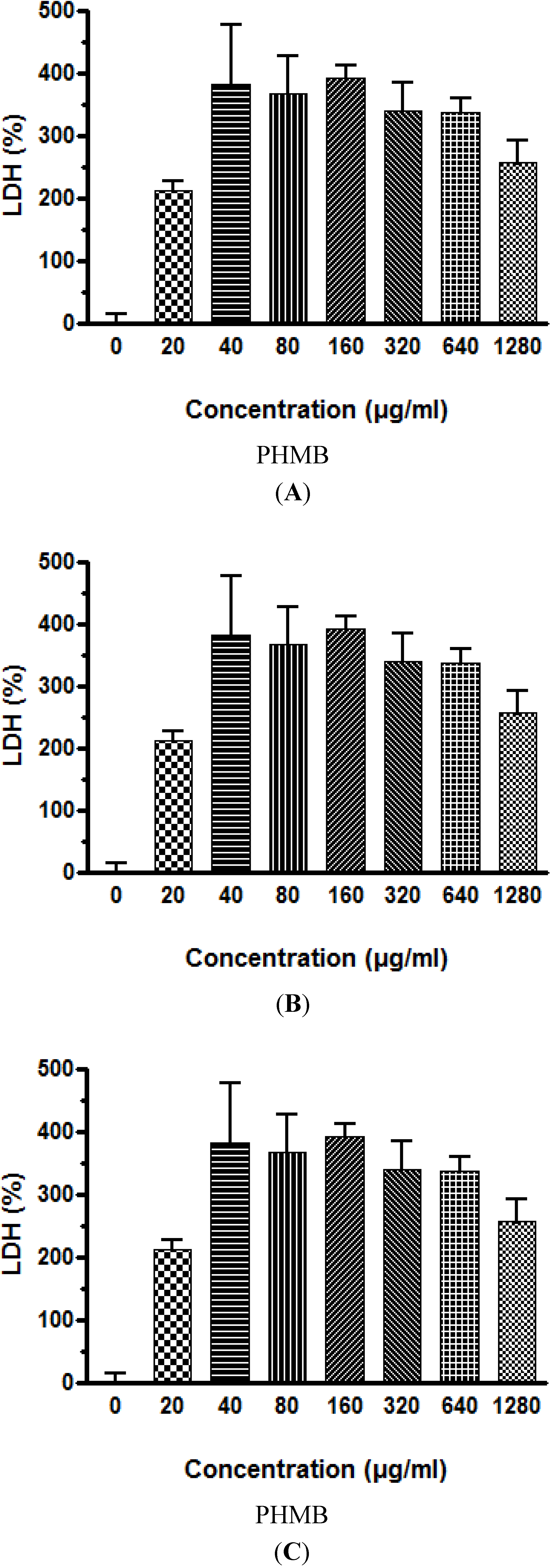
Similar to viability results using the neutral red test, cytotoxicity testing using LDH leakage into the culture medium also showed that PHMB is cytotoxic to the three cell lines studied, however, the HepG2 human liver cell line is again the most sensitive to the effects of PHMB. The cytotoxicity targeting the cell membrane occurs rapidly with a statistically significant increase in LDH leakage observed after 3 h exposure to PHMB at concentrations of 20 μg/mL, and maximum leakage at 40 to 80 μg/mL. At these levels of LDH leakage, 50% of HepG2 cells were killed whereas the LC 50% for the Caco-2 and Neuro-2A cells was 160 μg of PHMB/mL.
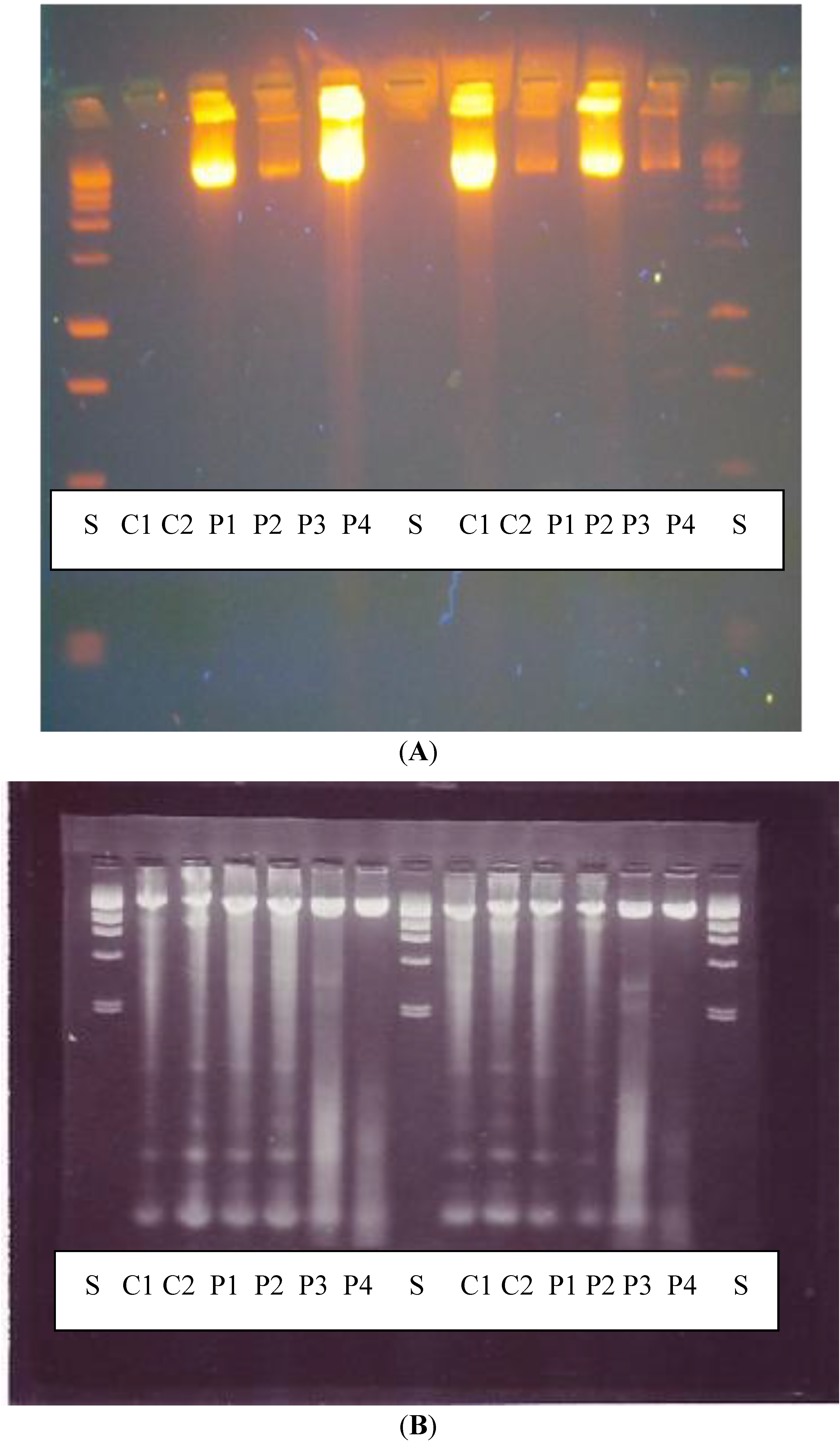
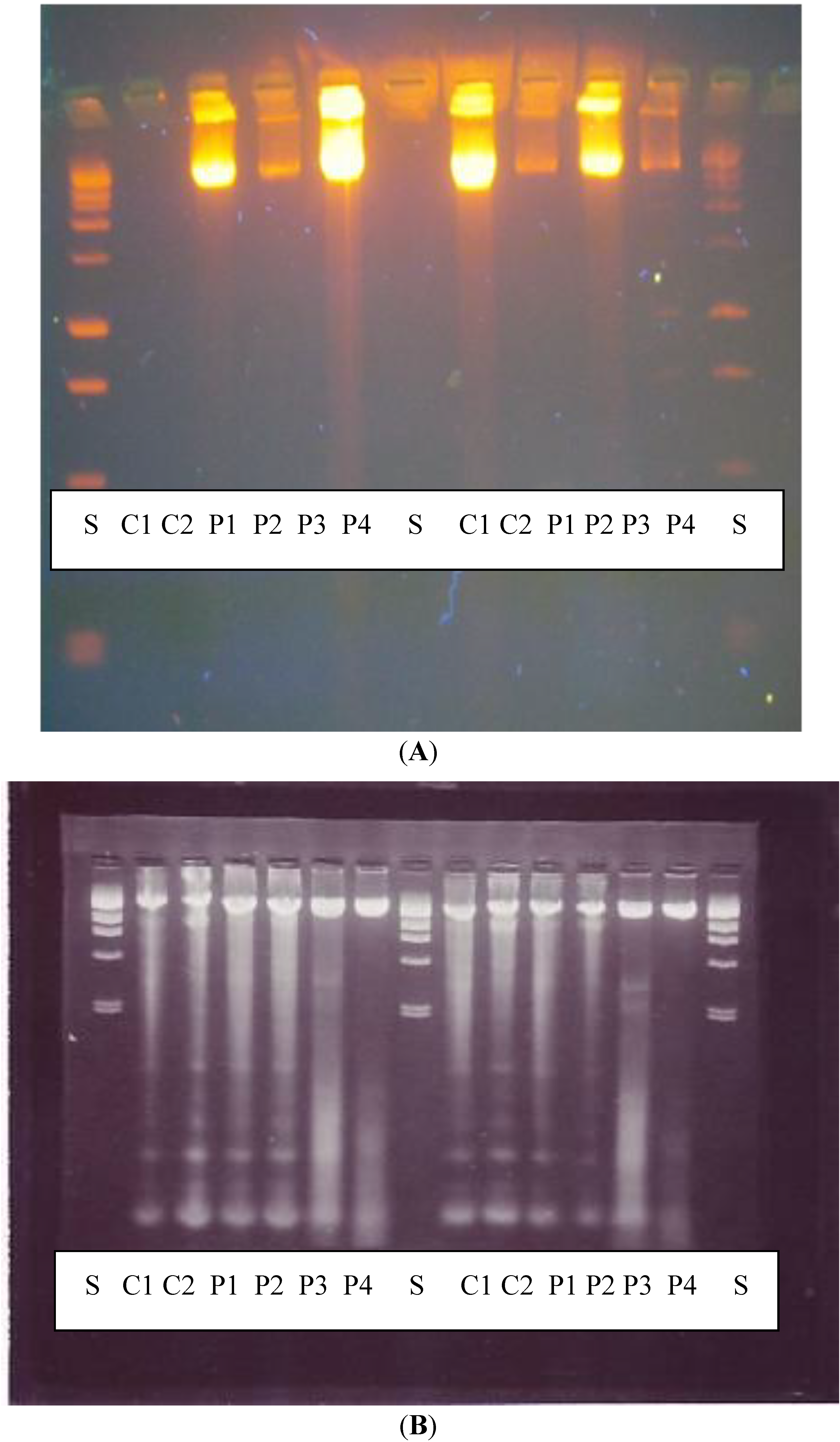
DNA fragmentation examined by agarose gel electrophoresis showed that there was no significant increase in DNA fragments following exposure to PHMB as would be expected if the cells were undergoing apoptosis, and results of the comet test, confirm that PHMB is not genotoxic (clastogenic).
Oxidative stress, a common cause of epigenetic changes to DNA [
33] was evaluated by quantifying the levels of MDA and 8-OHdG in PHMB treated cells. PHMB did not induce an increase in MDA concentration and DNA bases oxidation was not significantly increased (approximately 10%) compared to the negative control confirming the very low rate of oxidative stress. DNA methylation and histone modifications are two common epigenetic mechanisms that cells use to regulate replication [
16,
17,
18,
20,
29,
31,
32,
33,
37,
38] therefore, determination of the DNA methylation rate is necessary when dealing with a compound with possible epigenetic effects. In the case of PHMB, DNA cytosine methylation rate increased slightly (15–20%) but was not concentration dependent, occurring mainly in intestinal and hepatic cells and not in neuronal cells. The DNA methylation rate is generally higher in brain cells than in intestinal cells and in hepatocytes [
20,
32,
33,
38].
In addition to the DNA methylation rate, cytokine activity also has an influence on cell division and together, these parameters are determinant for overall cellular proliferation. Therefore, cytokine and transcription factor production (interleukin 1 alpha, TNF alpha and nuclear factor kappa B) were evaluated in the cells types studied. HepG2 cells, originating from liver, showed a higher production of cytokines (2–3 fold) compared to the control cells at 80 μg of PHMB, which is the IC 50% for this cell type in viability test, while it represents only half of the IC 50% value for Caco-2 and Neuro-2A cell types (
Figure 1 and
Table 13). The increase of these cytokines, although low, was significant in HepG2 cells, and there was a weak but significant increase in TNF alpha and in NF-κB in Caco-2 cells, and in Neuro-2A cells, respectively. The mitogenic effect of these cytokines could be enhanced if there was an interruption in the gap junction intercellular communication allowing cells to escape from contact inhibition [
16,
29]. However, there is apparently no inhibition of the GJIC by PHMB at concentrations in the range of the IC 50% values in any of the cell lines studies.
The mechanism by which PHMB kills mammalian cells is still a matter of investigation. It does appear that the membrane is the target resulting in cell lysis and cell death, just as in bacteria [
3,
39]. Röhner
et al. reported that lactate dehydrogenase activity showed a significant increase in the culture medium of human chondrocytes after a short incubation time with PHMB [
39], confirming the data in our studies and indicating that necrosis might occur in tissues followed by inflammation. However it is possible that PHMB exposed cells may undergo an apoptotic cell death, therefore, we also investigated several mechanisms leading to apoptosis and cell death.
There was very weak or no activation of caspase-3 in the any of the cells lines treated with PHMB indicating that the cells are not undergoing apoptosis. Choy
et al. very recently reported on the human corneal epithelial cells exposure to (PHMB) (0.00015%) (MPS-A), PHMB (0.0005%) (MPS-B) and PHMB (0.0001%) (MPS-C) diluted to 10–40%, for 1, 5 and 10 min [
4]. Cell viability and membrane integrity were assessed by MTT test flow cytometry following staining with AnnexinV-FITC/7-AAD. Their data showed that after 10 minutes exposure, almost 40 per cent of cells in MPS-A but less than five per cent in MPS-B or MPS-C, were in late necrotic stage. No apoptosis was observed. After 12 h of exposure, cell activity was significantly reduced in a concentration dependent manner for MPS-A treated cells only (
p > 0.05).
Although caspase-3 was not activated it is possible that other caspases such as caspase-1 which induces cell death by pyroptosis, were activated. Pyroptosis is a caspase-1-dependent programmed cell death, which features rapid plasma membrane rupture, DNA fragmentation, and release of proinflammatory intracellular contents [
40,
41]. This caspase 1-dependent cell death, is inherently inflammatory, is triggered by various pathological stimuli, such as stroke, heart attack or cancer, and is crucial for controlling microbial infections [
42]. Caspase-1 plays a key role in host defense through its dual functions in inducing a pro-inflammatory cell death and in promoting the secretion of pro-inflammatory cytokines, and a new study highlighted the specific importance of pyroptosis in resistance to intracellular pathogens [
41,
42,
43]. However, since our data do not show no indication of DNA cleavage, and since pyroptosis is mainly related to an inflammatory host response, it does not appear that caspase 1-dependent cell death occurred in the PHMB treated cell lines we investigated.
Additionally effectors in programmed cell death, such as p53, p21, bax, and bcl-2 did not show any increase in gene expression or at the protein level, therefore, it does not appear that PHMB induced cell death is not the result of apoptotsis, but rather is cytotoxic at the cell membrane level resulting in necrotic cell death.
At concentrations of 1 to 100–200 μg/mL, the effects of PHMB are weak only occurring at cytotoxic concentrations and are not concentration-dependent. Our proposed mechanism of action for PHMB induced cell death is as follows: at very low concentrations, 0.1 to 0.5–1 μg/mL, PHMB binds to the cell membrane facilitating uptake of nutrients and ion channel trafficking, resulting in increased cell viability and division (hormesis effect). However, when PHMB is administered at higher concentrations (>20–50 μg in culture medium) which correspond to 20–100 and up to 500 mg/L in vivo, the increased binding of PHMB to the membrane is cytotoxic resulting in lysis of the membrane with leakage of enzymes and cytokines and eventually cell death by necrosis.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}