Inhibitory Effect of N,N-Didesmethylgrossularine-1 on Inflammatory Cytokine Production in Lipopolysaccharide-Stimulated RAW 264.7 Cells

 ,
, 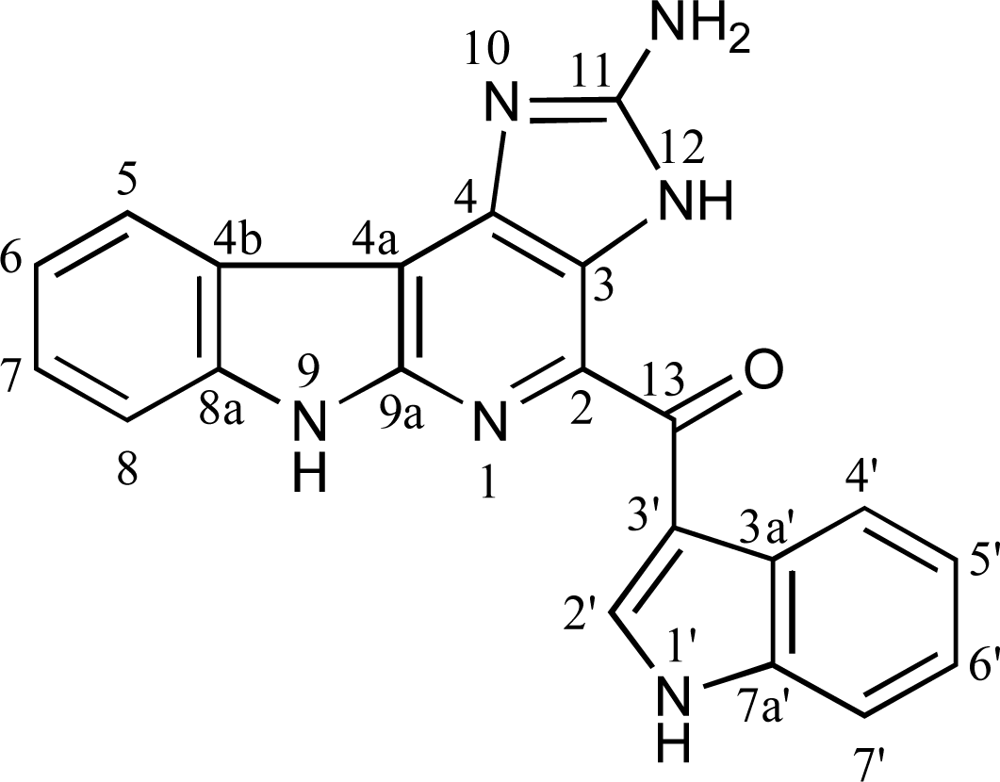{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Confirmation of DDMG-1
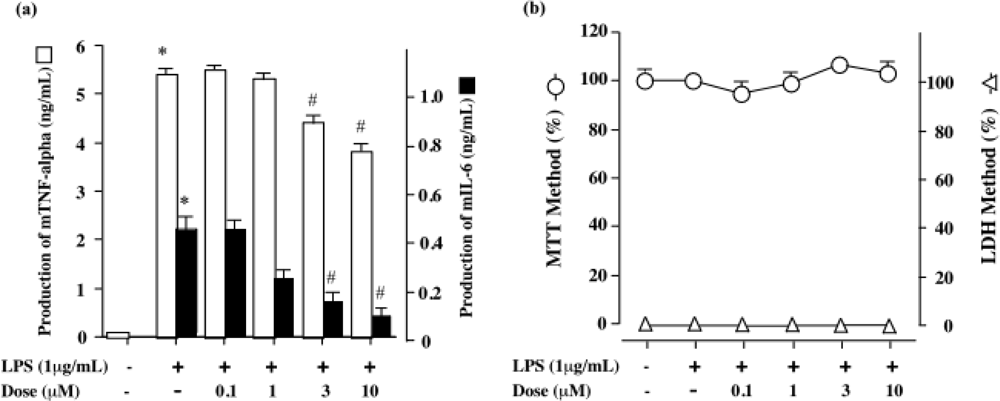
2.2. DDMG-1 inhibited LPS-induced mTNF-α and mIL-6 production in RAW 264.7 cells
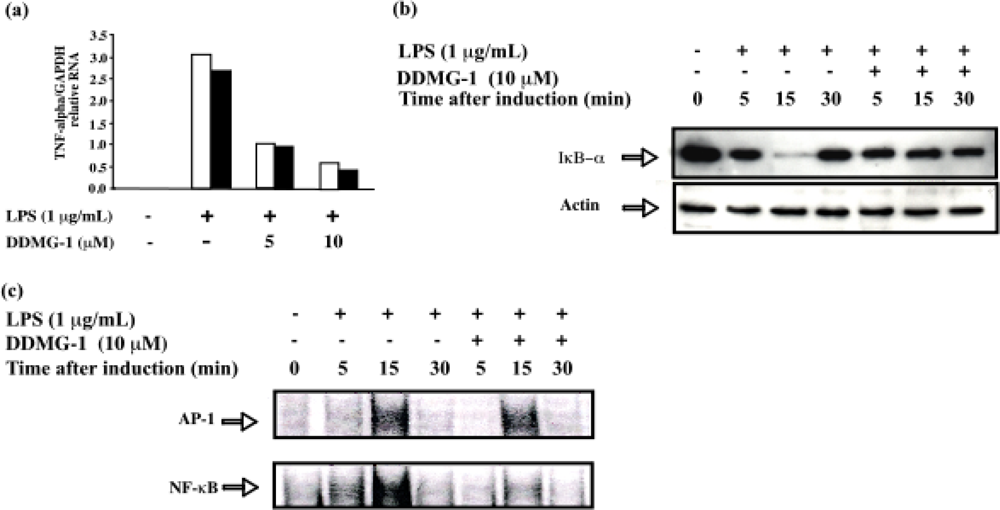
2.3. DDMG-1 down-regulated TNF-a mRNA expression at the transcriptional level
2.4. DDMG-1 inhibited IκB-α degradation in LPS-stimulated RAW 264.7 cells
2.5. DDMG-1 inhibited LPS-stimulated binding of NF-κB to DNA
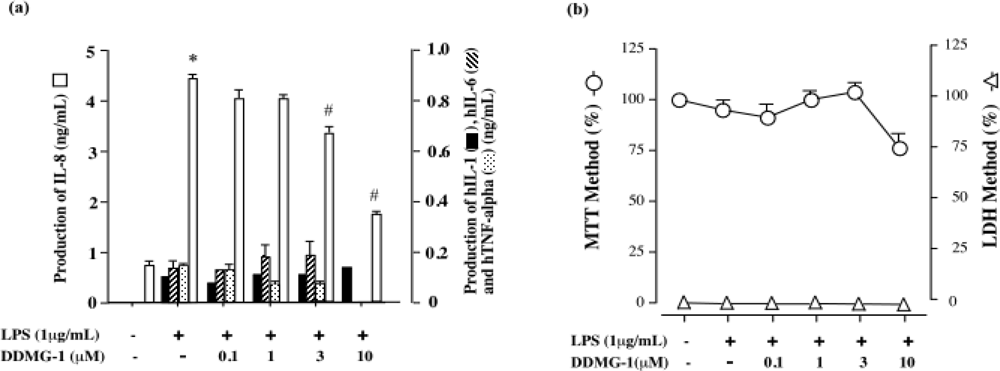
2.6. DDMG-1 inhibited LPS-induced hTNF-α hIL-1b, hIL-6, and IL-8 production in CD14+-THP-1 cells
3. Experimental Section
3.1. General
3.2. Isolation of DDMG-1
3.3. Materials
3.4. Cell culture
3.5. Murine TNF-α (mTNF-α), murine IL-6 (mIL-6), human TNF-α (hTNF-α), human IL-β (hIL-β), human IL-6 (hIL-6), and human IL-8 (hIL-8) quantification
3.6. Measurement of TNF-α mRNA expression
3.7. Western blot analysis
3.8. Cell viability assay and determination of cytotoxicity
3.9. Preparation of nuclear extracts and EMSA
3.10. Statistical analysis
4. Conclusions
Acknowledgments
References
- Ross, JA; Auger, MJ; Burke, B; Lewis, CE. The biology of the macrophage. In The Macrophage, 2nd ed; Burke, B, Lewis, CE, Eds.; Oxford Medical Publications: Oxford, UK, 2002; pp. 1–72. [Google Scholar]
- Poltorak, A; He, X; Smirnova, I; Liu, MY; van Huffel, C; De, X; Bridwell, D; Alejos, E; Silva, M; Galanos, C; Freudenberg, M; Ricciardi-Castagnoli, P; Layton, B; Beatler, B. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar]
- Mendes, SDS; Candi, A; Vansteenbrugge, M; Pignon, M-R; Bult, H; Boudjeltia, KZB; Munaut, C; Taes, M. Microarray analysis of the effects of NF-κB or p13K pathway inhibitors on the LPS-induced gene expression profile in RAW264.7 cells Synaergistic effects of rapamycin on LPS-induced MMP9-overexpression. Cell Signaling 2009, 21, 1109–1122. [Google Scholar]
- Hirohata, S; Yanagida, T; Itoh, K; Nakamura, H; Yoshino, S; Tomita, T. Accelerate generation of CD14+ monocyte-lineage cells from the bone marrow of theumatoid arthritis patients. Arthritis Rheum 1996, 39, 836–843. [Google Scholar]
- Abas, SA; Hossain, MB; Helm, D; Schmits, FJ; Laney, M; Cabusly, R; Schatzman, RC. Alkaloids from the Tunicate Polycarpa aurata from Chuuk Atoll. J Org Chem 1996, 61, 2709–2712. [Google Scholar]
- Miyake, FY; Yakushijin, K; Horne, DA. Biomimetic synthesis of grossularines-1. Angew Chem Int Ed 2005, 44, 3280–3282. [Google Scholar]
- Moquin-Pattey, C; Guyot, M. Grossularine-1 and grossularine-2, cytotoxic α-carbolines from the tunicate: Dendrodoa grossularia. Tetrahedron 1989, 45, 3445–3450. [Google Scholar]
- Helbecque, N; Moquin, C; Bernier, JL; Morel, E; Guyot, M; Henichart, JP. Grossularine-1 and grossularine-2, α-carbolines from Dendrodoa grossularia, as possible intercalative agents. Cancer Biochem Biophys 1987, 9, 271–279. [Google Scholar]
- Wang, W-F; Oda, T; Fujita, A; Mangindaan, REP; Nakazawa, T; Ukai, K; Kobayashi, H; Namikoshi, M. Three new sulfur-containing alkaloids, polycarpaurines A, B, and C, from an Indonesian ascidian. Polycarpa aurata Tetrahedron 2007, 63, 409–412. [Google Scholar]
- Keifer, AJ; Guttridge, CD; Ashburner, PB; Baldwin, SA. Inhibition of NF-kappaB activity by thalidomide through suppression of IkappaB kinase activity. J Biol Chem 2001, 276, 22382–22387. [Google Scholar]
- Nagahira, A; Nagahira, K; Murafuji, H; Abe, K; Magota, K; Matsui, M; Oikawa, S. Identification of a novel inhibitor of LPS-Induced TNF-α production with antiproliferative activity in monocyte/macrophages. Biochem Biophys Res Commun 2001, 281, 1030–1036. [Google Scholar]
- Bresnihan, B; Youssef, P; Burke, B; Lewin, GE. Macrophages in rheumatoid arthritis. In The Macrkophage, 2nd ed; Burke, B, Lewis, CE, Eds.; Oxford Medical Publications: Oxford, UK, 2002; pp. 391–433. [Google Scholar]
- Coussens, LM; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar]
- Siebenlist, U; Brown, K; Claudio, E. Control of lymphocyte development by nuclear factor-kappaB. Nat Rev Immunol 2005, 5, 435–445. [Google Scholar]
- Oda, T; Ueda, A; Shimizu, N; Handa, H; Kasahara, T. Suppression of monocyte chemoattractant protein 1, but not IL-8, by alprazolam: Effect of alprazolam on c-Re/p65 and c-Rel/p50 bindin g to the monocyte chemoattractant protein 1 promoter region. J Immunol 2002, 169, 3329–3335. [Google Scholar]
- Shimizu, S; Nomoto, M; Naito, S; Yamamoto, T; Momose, K. Stimulation of nitric oxide synthase during oxidative endothelial cell injury. Biochem Pharmacol 1998, 55, 77–83. [Google Scholar]
- Carmichael, J; DeGraff, WG; Gazdar, AF; Gazdar, AF; Minna, JD; Mitchell, JB. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of radiosensitivity. Cancer Res 1987, 47, 939–942. [Google Scholar]
- Rojanasakul, Y; Ye, J; Chen, F; Wang, L; Cheng, N; Castranova, V; Vallyathan, V; Shi, X. Dependence of NF-kappaB activation and free radical generation on silica-induced TNF-alpha production in macrophages. Mol Cell Biochem 1999, 200, 119–125. [Google Scholar]




© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Oda, T.; Lee, J.-S.; Sato, Y.; Kabe, Y.; Sakamoto, S.; Handa, H.; Mangindaan, R.E.P.; Namikoshi, M. Inhibitory Effect of N,N-Didesmethylgrossularine-1 on Inflammatory Cytokine Production in Lipopolysaccharide-Stimulated RAW 264.7 Cells. Mar. Drugs 2009, 7, 589-599. https://doi.org/10.3390/md7040589
Oda T, Lee J-S, Sato Y, Kabe Y, Sakamoto S, Handa H, Mangindaan REP, Namikoshi M. Inhibitory Effect of N,N-Didesmethylgrossularine-1 on Inflammatory Cytokine Production in Lipopolysaccharide-Stimulated RAW 264.7 Cells. Marine Drugs. 2009; 7(4):589-599. https://doi.org/10.3390/md7040589
Chicago/Turabian StyleOda, Taiko, Jong-Soo Lee, Yuta Sato, Yasuaki Kabe, Satoshi Sakamoto, Hiroshi Handa, Remy E. P. Mangindaan, and Michio Namikoshi. 2009. "Inhibitory Effect of N,N-Didesmethylgrossularine-1 on Inflammatory Cytokine Production in Lipopolysaccharide-Stimulated RAW 264.7 Cells" Marine Drugs 7, no. 4: 589-599. https://doi.org/10.3390/md7040589
APA StyleOda, T., Lee, J.-S., Sato, Y., Kabe, Y., Sakamoto, S., Handa, H., Mangindaan, R. E. P., & Namikoshi, M. (2009). Inhibitory Effect of N,N-Didesmethylgrossularine-1 on Inflammatory Cytokine Production in Lipopolysaccharide-Stimulated RAW 264.7 Cells. Marine Drugs, 7(4), 589-599. https://doi.org/10.3390/md7040589