
Jellyfish Collagen in the Mediterranean Spotlight: Transforming Challenges into Opportunities

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Jellyfish Collagen
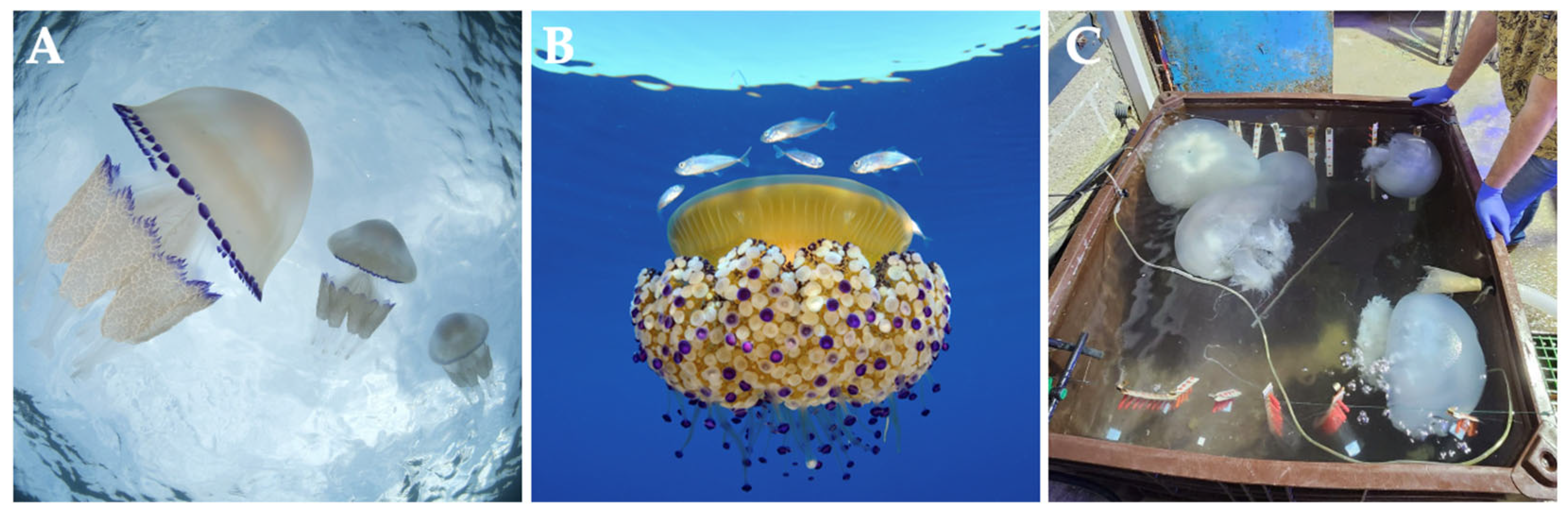
3. Jellyfish Rich in Collagen That Inhabit the Mediterranean Sea
3.1. Order Rhizostomeae
3.2. Jellyfish Species
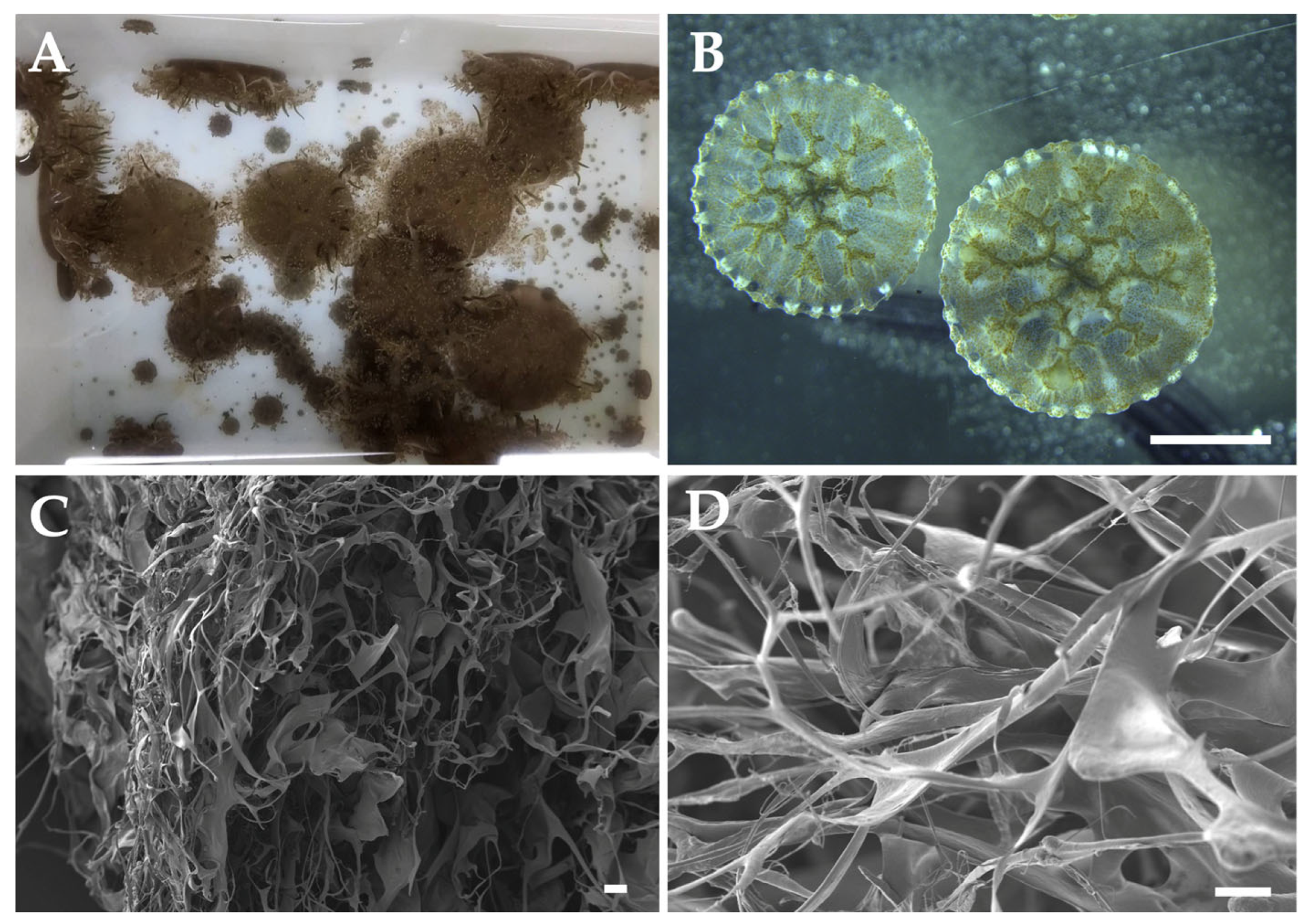
4. Jellyfish Collagen Extraction Methods
4.1. Preparation and Pre-Treatment
4.2. Extraction Process
4.2.1. Acid-Based
4.2.2. Pepsin
4.2.3. Ultrasonic Assisted

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collagen Extraction Method | Advantages | Disadvantages | Jellyfish Species | References | |
|---|---|---|---|---|---|
| Acid soluble collagen |  | No damage to collagen structure Cost-efficient Simple process | Long extraction time Low yield Darked protein coloration | Rhizostoma pulmo, Cotylorhiza tuberculata, Cassiopea andromeda, Catostylus tagi | [22,66,79,83,84] |
| Pepsin soluble collagen |  | Increased yield Non-flamable Non-toxic | High cost Difficult to scale up | Rhizostoma pulmo, Cassiopea andromeda, Catostylus tagi | [79,83,84,85] |
| Ultrasonic assisted |  | No damage to collagen structure Better purity Increased yield Reduced extraction time | Limited industrial scalability Complex steps | Rhizostoma pulmo | [65,66,79] |
4.2.4. Green Extraction Methods
4.3. Recovery, Purification, and Stabilization
5. Application of Jellyfish Collagen
5.1. Nutraceutical
5.2. Cosmeceuticals
5.3. Biomaterials
| Jellyfish Species | Collagen Extraction Method | Biomaterial Type | Additive | Cross-Linker | Biological Evaluation | Application | References | |
|---|---|---|---|---|---|---|---|---|
| Rhizostoma pulmo |  | PSC | Scaffolds | Keratin/nano-spherical hydroxyapatite from eggshells | EDC/NHS and uncross linked | Human periodontal ligament fibroblast | Bone tissue | [85] |
| Rhizostoma pulmo |  | Jellagen® * | Scaffolds | - | EDC and uncross linked | Wistar rats | Wound healing | [109] |
| Rhizostoma pulmo |  | Jellagen® * | Scaffolds | - | EDC | Ovarian cancer cells | Cell culture | [110] |
| Rhizostoma pulmo |  | Jellagen® * | Scaffolds | - | EDC/NHS | Wistar rats | Bone tissue | [111] |
| Rhizostoma pulmo |  | Jellagen® * | Collagen solution | - | - | Human induced pluripotent stem cells (UKBi005-A) | Cell culture | [112] |
| Rhizostoma pulmo |  | Jellagen® * | Scaffolds | - | EDC | Bovine chondroprogenitor cell | Cartilage tissue | [106] |
| Rhizostoma pulmo |  | Jellagen® * | Collagen solution and scaffolds | - | EDC | L929 fibroblasts and MC373-E1 pre-osteoblasts | Bone tissue | [117] |
| Rhizostoma pulmo |  | Jellagen® * | Hydrogel | - | Genipin | Chondrocytes | Cell culture | [113] |
| Rhizostoma pulmo |  | Jellagen® * | Hydrogel | Chitosan/ fucoidan | - | Chondrocyte-like cells (ATDC5) | Cartilage tissue | [118] |
| Rhizostoma pulmo |  | Jellagen® * | Collagen solution | - | - | Fibroblast and pre-osteoblasts | Cell culture | [119] |
| Rhizostoma pulmo |  | ASC | Scaffolds | Chitosan | - | Rat embryonic liver cells | Liver tissue | [77] |
| Rhizostoma pulmo |  | ASC | Hydrogel | - | HRP/H2O2 | Nasal Chondrocytes and MC3T3-E1 pre-osteoblastic | Cartilage tissue | [120] |
| Rhizostoma pulmo |  | Jellagen® * | Hydrogel | - | EDC/NHS; EDC/sNHS; PEG; Genipin | Immortalized human mesenchymal stem cells | Regenerative medicine | [121] |
| Rhizostoma pulmo |  | Jellagen® * | Collagen solution | - | - | Primary fibroblasts, HT-1080 human fibrosarcoma line and Y201 mesenchymal stem cells | Cell culture | [27] |
| Rhizostoma pulmo |  | ASC, PSC | Collagen solution | - | - | Human fibroblasts and MG-63 osteosarcoma cells | Cell culture | [22] |
| Cassiopea andromeda |  | ASC, PSC | Scaffolds | TPU | - | Human monocytes and porcine dermal fibroblasts | Regenerative medicine | [84] |
| Cassiopea andromeda |  | ASC, PSC | Hydrogel | TPU | - | Human monocytes and porcine dermal fibroblasts | Wound healing | [122] |
| Cassiopea andromeda |  | - | Scaffolds | - | - | Human fibroblasts | Skin tissue | [114] |
| Catostylus tagi |  | - | Hydrogel | - | EDC | - | Drug delivery | [116] |
| Catostylus tagi |  | ASC, PSC | Microparticles | - | - | - | Drug delivery | [83] |
6. Exploitation of the Resource
6.1. Active Fishing
6.2. By-Catch
6.3. Aquaculture
| Jellyfish Species | Strobilation Type | T (°C) | Salinity | Tank | Foods Regimen | References | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Polyp | Ephyra | Juvenile | Adult | Polyp | Ephyra | Juvenile | Adult | ||||||
| Cassiopea spp. * |  | Monodisc | 18–29 | 36–37, tolerant | WAK | WAK, MGT | MRT | MRT | R, AN | R, AN | R, AN | AN, WP | [45,48,138,139,145,148,150] |
| Catostylus spp. |  | Oligodisc | 19–28 | 32–37, tolerant | - | MGT | K, P-K | K, P-K, MRT | MG, R, AN | MM, R, AN | AN | AN, WP, FC | [48,139,151,152] |
| Cotylorhiza tuberculata * |  | Monodisc | 22–28 | 36–38, tolerant | WAK | WAK, MGT | P-K, MRT, RST | CT | R | R | AN | AN | [48,138,139,145] |
| Phyllorhiza punctata * |  | Monodisc | 13–25 | 36–37 | WAK | WAK | P-K, MRT, RST | K, P-K, CT | R, AN | R | AN | AN | [42,45,138,139,145,146] |
| Rhizostoma luteum |  | Monodisc | 17–26 | 34–38 | - | MGT | K | K | R | AN | AN | AN | [44,146] |
| Rhizostoma pulmo |  | Oligodisc | 13–28 | 36–37 | WAK | WAK | K, P-K | MRT | R, AN | R, AN | AN, WP | AN, WP | [45,48,138,139,145] |
| Rhopilema nomadica |  | Oligodisc a | 21–29 | 39–40 | - | K | K | K | AN | AN | AN | AN | [58,60] |
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | Extracellular matrix |
| SFE | Supercritical Fluid Extraction |
| DES | Deep Eutectic Solvents |
| NADESs | Natural Deep Eutectic Solvents |
| NIS | Non-indigenous species |
References
- Richardson, A.J.; Bakun, A.; Hays, G.C.; Gibbons, M.J. The Jellyfish Joyride: Causes, Consequences and Management Responses to a More Gelatinous Future. Trends Ecol. Evol. 2009, 24, 312–322. [Google Scholar] [CrossRef]
- Morandini, A.C. Impacts of Jellyfish: Gelatinous Problems or Opportunities? Arq. Ciênc. Mar 2022, 55, 123–140. [Google Scholar] [CrossRef]
- Kogovšek, T.; Bogunović, B.; Malej, A. Recurrence of Bloom-Forming Scyphomedusae: Wavelet Analysis of a 200-Year Time Series. Hydrobiologia 2010, 645, 81–96. [Google Scholar] [CrossRef]
- Brotz, L.; Cheung, W.W.L.; Kleisner, K.; Pakhomov, E.; Pauly, D. Increasing Jellyfish Populations: Trends in Large Marine Ecosystems. Hydrobiologia 2012, 690, 3–20. [Google Scholar] [CrossRef]
- Purcell, J.E.; Uye, S.I.; Lo, W.T. Anthropogenic Causes of Jellyfish Blooms and Their Direct Consequences for Humans: A Review. Mar. Ecol. Prog. Ser. 2007, 350, 153–174. [Google Scholar] [CrossRef]
- Condon, R.H.; Duarte, C.M.; Pitt, K.A.; Robinson, K.L.; Lucas, C.H.; Sutherland, K.R.; Mianzan, H.W.; Bogeberg, M.; Purcell, J.E.; Decker, M.B.; et al. Recurrent Jellyfish Blooms Are a Consequence of Global Oscillations. Proc. Natl. Acad. Sci. USA 2013, 110, 1000–1005. [Google Scholar] [CrossRef]
- Dror, H.; Angel, D. Rising Seawater Temperatures Affect the Fitness of Rhopilema Nomadica Polyps and Podocysts and the Expansion of This Medusa into the Western Mediterranean. Mar. Ecol. Prog. Ser. 2024, 728, 123–143. [Google Scholar] [CrossRef]
- Treible, L.M.; Condon, R.H. Temperature-Driven Asexual Reproduction and Strobilation in Three Scyphozoan Jellyfish Polyps. J. Exp. Mar. Biol. Ecol. 2019, 520, 151204. [Google Scholar] [CrossRef]
- Sperone, E.; Cipriano, G.; Chimienti, G.; Petrocelli, A.; Cillari, T.; Allegra, A.; Berto, D.; Bosch-Belmar, M.; Falautano, M.; Maggio, T.; et al. Snapshot of the Distribution and Biology of Alien Jellyfish Cassiopea Andromeda (Forsskål, 1775) in a Mediterranean Touristic Harbour. Biology 2022, 11, 319. [Google Scholar] [CrossRef]
- Duarte, C.M.; Pitt, K.A.; Lucas, C.H.; Purcell, J.E.; Uye, S.I.; Robinson, K.; Brotz, L.; Decker, M.B.; Sutherland, K.R.; Malej, A.; et al. Is Global Ocean Sprawl a Cause of Jellyfish Blooms? Front. Ecol. Environ. 2013, 11, 91–97. [Google Scholar] [CrossRef]
- Graham, W.M.; Gelcich, S.; Robinson, K.L.; Duarte, C.M.; Brotz, L.; Purcell, J.E.; Madin, L.P.; Mianzan, H.; Sutherland, K.R.; Uye, S.I.; et al. Linking Human Well-Being and Jellyfish: Ecosystem Services, Impacts, and Societal Responses. Front. Ecol. Environ. 2014, 12, 515–523. [Google Scholar] [CrossRef]
- Merquiol, L.; Romano, G.; Ianora, A.; D’Ambra, I. Biotechnological Applications of Scyphomedusae. Mar. Drugs 2019, 17, 604. [Google Scholar] [CrossRef]
- Bosch-Belmar, M.; Milisenda, G.; Basso, L.; Doyle, T.K.; Leone, A.; Piraino, S. Jellyfish Impacts on Marine Aquaculture and Fisheries. Rev. Fish. Sci. Aquac. 2021, 29, 242–259. [Google Scholar] [CrossRef]
- Canepa, A.; Fuentes, V.; Sabatés, A.; Piraino, S.; Boero, F.; Gili, J.-M. Pelagia Noctiluca in the Mediterranean Sea. In Jellyfish Blooms; Springer: Dordrecht, The Netherlands, 2014; pp. 237–266. ISBN 9789400770157. [Google Scholar]
- Nunes, P.A.L.D.; Loureiro, M.L.; Piñol, L.; Sastre, S.; Voltaire, L.; Canepa, A. Analyzing Beach Recreationists’ Preferences for the Reduction of Jellyfish Blooms: Economic Results from a Stated-Choice Experiment in Catalonia, Spain. PLoS ONE 2015, 10, e0126681. [Google Scholar] [CrossRef]
- Milisenda, G.; Rosa, S.; Fuentes, V.L.; Boero, F.; Guglielmo, L.; Purcell, J.E.; Piraino, S. Jellyfish as Prey: Frequency of Predation and Selective Foraging of Boops Boops (Vertebrata, Actinopterygii) on the Mauve Stinger Pelagia Noctiluca (Cnidaria, Scyphozoa). PLoS ONE 2014, 9, e94600. [Google Scholar] [CrossRef]
- Heaslip, S.G.; Iverson, S.J.; Bowen, W.D.; James, M.C. Jellyfish Support High Energy Intake of Leatherback Sea Turtles (Dermochelys Coriacea): Video Evidence from Animal-Borne Cameras. PLoS ONE 2012, 7, e33259. [Google Scholar] [CrossRef]
- Fernández-Alías, A.; Marcos, C.; Quispe, J.I.; Sabah, S.; Pérez-Ruzafa, A. Population Dynamics and Growth in Three Scyphozoan Jellyfishes, and Their Relationship with Environmental Conditions in a Coastal Lagoon. Estuar. Coast. Shelf Sci. 2020, 243, 106901. [Google Scholar] [CrossRef]
- Coppola, D.; Oliviero, M.; Vitale, G.A.; Lauritano, C.; D’Ambra, I.; Iannace, S.; de Pascale, D. Marine Collagen from Alternative and Sustainable Sources: Extraction, Processing and Applications. Mar. Drugs 2020, 18, 214. [Google Scholar] [CrossRef]
- Almeida, M.; Silva, T.; Solstad, R.G.; Lillebø, A.I.; Calado, R.; Vieira, H. How Significant Are Marine Invertebrate Collagens? Exploring Trends in Research and Innovation. Mar. Drugs 2025, 23, 2. [Google Scholar] [CrossRef]
- Chiarelli, P.G.; Suh, J.H.; Pegg, R.B.; Chen, J.; Mis Solval, K. The Emergence of Jellyfish Collagen: A Comprehensive Review on Research Progress, Industrial Applications, and Future Opportunities. Trends Food Sci. Technol. 2023, 141, 104206. [Google Scholar] [CrossRef]
- Addad, S.; Exposito, J.Y.; Faye, C.; Ricard-Blum, S.; Lethias, C. Isolation, Characterization and Biological Evaluation of Jellyfish Collagen for Use in Biomedical Applications. Mar. Drugs 2011, 9, 967–983. [Google Scholar] [CrossRef]
- Wang, H. A Review of the Effects of Collagen Treatment in Clinical Studies. Polymers 2021, 13, 3868. [Google Scholar] [CrossRef]
- Lin, K.; Zhang, D.; Macedo, M.H.; Cui, W.; Sarmento, B.; Shen, G. Advanced Collagen-Based Biomaterials for Regenerative Biomedicine. Adv. Funct. Mater. 2019, 29, 1804943. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—Structure, Function, and Biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef]
- Smith, I.P.; Domingos, M.; Richardson, S.M.; Bella, J. Characterization of the Biophysical Properties and Cell Adhesion Interactions of Marine Invertebrate Collagen from Rhizostoma Pulmo. Mar. Drugs 2023, 21, 59. [Google Scholar] [CrossRef]
- Privalov, P.L. Stability of Proteins: Proteins Which Do Not Present a Single Cooperative System. In Advances in Protein Chemistry; Anfinsen, C.B., Edsall, J.T., Richards, F.M., Eds.; Academic Press: Cambridge, MA, USA, 1982; Volume 35, pp. 1–104. [Google Scholar]
- Exposito, J.-Y.; Cluzel, C.; Garrone, R.; Lethias, C. Evolution of Collagens. Anat. Rec. 2002, 268, 302–316. [Google Scholar] [CrossRef]
- Frank, U.; Rinkevich, B. Scyphozoan Jellyfish’s Mesoglea Supports Attachment, Spreading and Migration of Anthozoans’ Cells in Vitro. Cell Biol. Int. 1999, 23, 307–311. [Google Scholar] [CrossRef]
- Mills, C.E. Density Is Altered in Hydromedusae and Ctenophores in Response to Changes in Salinity. Biol. Bull. 1984, 166, 206–215. [Google Scholar] [CrossRef]
- Morandini, A.C. Morphology of Rhizostomeae Jellyfishes: What Is Known and What We Advanced since the 1970s. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 2024; Volume 98, pp. 61–97. ISBN 978-0-443-29646-8. [Google Scholar]
- Acuña, J.L.; López-Urrutia, Á.; Colin, S. Faking Giants: The Evolution of High Prey Clearance Rates in Jellyfishes. Science 2011, 333, 1627–1629. [Google Scholar] [CrossRef]
- Schmid, V. The Transformational Potential of Striated Muscle in Hydromedusae. Dev. Biol. 1976, 49, 508–517. [Google Scholar] [CrossRef]
- Sinigaglia, C.; Peron, S.; Eichelbrenner, J.; Chevalier, S.; Steger, J.; Barreau, C.; Houliston, E.; Leclère, L. Pattern Regulation in a Regenerating Jellyfish. eLife 2020, 9, e54868. [Google Scholar] [CrossRef]
- Morishige, H.; Sugahara, T.; Nishimoto, S.; Muranaka, A.; Ohno, F.; Shiraishi, R.; Doi, M. Immunostimulatory Effects of Collagen from Jellyfish in Vivo. Cytotechnology 2011, 63, 481–492. [Google Scholar] [CrossRef]
- Brotz, L.; Angel, D.L.; D’Ambra, I.; Enrique-Navarro, A.; Lauritano, C.; Thibault, D.; Prieto, L. Rhizostomes as a Resource: The Expanding Exploitation of Jellyfish by Humans. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 2024; Volume 98, pp. 511–547. ISBN 978-0-443-29646-8. [Google Scholar]
- Straehler-Pohl, I.; Widmer, C.L.; Morandini, A.C. Characterizations of Juvenile Stages of Some Semaeostome Scyphozoa (Cnidaria), with Recognition of a New Family (Phacellophoridae). Zootaxa 2011, 2741, 1–37. [Google Scholar] [CrossRef]
- Ballesteros, A.; Östman, C.; Santín, A.; Marambio, M.; Narda, M.; Gili, J.-M. Cnidome and Morphological Features of Pelagia Noctiluca (Cnidaria: Scyphozoa) throughout the Different Life Cycle Stages. Front. Mar. Sci. 2021, 8, 1059. [Google Scholar] [CrossRef]
- Straehler-Pohl, I.; Jarms, G. Identification Key for Young Ephyrae: A First Step for Early Detection of Jellyfish Blooms. Hydrobiologia 2010, 645, 3–21. [Google Scholar] [CrossRef]
- Schiariti, A.; Holst, S.; Tiseo, G.R.; Miyake, H.; Morandini, A.C. Life Cycles and Reproduction of Rhizostomeae. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 2024; Volume 98, pp. 193–254. ISBN 978-0-443-29646-8. [Google Scholar]
- Helm, R.R. Evolution and Development of Scyphozoan Jellyfish. Biol. Rev. 2018, 93, 1228–1250. [Google Scholar] [CrossRef]
- Fuentes, V.; Straehler-Pohl, I.; Atienza, D.; Franco, I.; Tilves, U.; Gentile, M.; Acevedo, M.; Olariaga, A.; Gili, J.M. Life Cycle of the Jellyfish Rhizostoma Pulmo (Scyphozoa: Rhizostomeae) and Its Distribution, Seasonality and Inter-Annual Variability along the Catalan Coast and the Mar Menor (Spain, NW Mediterranean). Mar. Biol. 2011, 158, 2247–2266. [Google Scholar] [CrossRef]
- Kienberger, K.; Riera-Buch, M.; Schönemann, A.M.; Bartsch, V.; Halbauer, R.; Prieto, L. First Description of the Life Cycle of the Jellyfish Rhizostoma Luteum (Scyphozoa: Rhizostomeae). PLoS ONE 2018, 13, e0202093. [Google Scholar] [CrossRef]
- Schiariti, A.; Morandini, A.C.; Jarms, G.; Von Glehn Paes, R.; Franke, S.; Mianzan, H. Asexual Reproduction Strategies and Blooming Potential in Scyphozoa. Mar. Ecol. Prog. Ser. 2014, 510, 241–253. [Google Scholar] [CrossRef]
- Heins, A.; Glatzel, T.; Holst, S. Revised Descriptions of the Nematocysts and the Asexual Reproduction Modes of the Scyphozoan Jellyfish Cassiopea Andromeda (Forskål, 1775). Zoomorphology 2015, 134, 351–366. [Google Scholar] [CrossRef]
- Straehler-Pohl, I.; Jarms, G. Back to the Roots, Part 2—Rhopaliophora (Scyphozoa, Cubozoa and Staurozoa) Reborn Based on Early Life Cycle Data. Plankton Benthos Res. 2022, 17, 105–126. [Google Scholar] [CrossRef]
- Straehler-Pohl, I.; Jarms, G. Back to the Roots, Part 1—Early Life Cycle Data of Rhopaliophora (Scyphozoa, Cubozoa and Staurozoa). Plankton Benthos Res. 2022, 17, 1–33. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under Siege: Spatial Overlap between Marine Biodiversity, Cumulative Threats and Marine Reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Mariottini, G.L.; Pane, L. Mediterranean Jellyfish Venoms: A Review on Scyphomedusae. Mar. Drugs 2010, 8, 1122–1152. [Google Scholar] [CrossRef]
- Marambio, M.; Canepa, A.; Lòpez, L.; Gauci, A.A.; Gueroun, S.K.M.; Zampardi, S.; Boero, F.; Yahia, O.K.-D.; Yahia, M.N.D.; Fuentes, V.; et al. Unfolding Jellyfish Bloom Dynamics along the Mediterranean Basin by Transnational Citizen Science Initiatives. Diversity 2021, 13, 274. [Google Scholar] [CrossRef]
- Leoni, V.; Bonnet, D.; Ramírez-Romero, E.; Molinero, J.C. Biogeography and Phenology of the Jellyfish Rhizostoma Pulmo (Cnidaria: Scyphozoa) in Southern European Seas. Glob. Ecol. Biogeogr. 2021, 30, 622–639. [Google Scholar] [CrossRef]
- Marambio, M.; Ballesteros, A.; López-Castillo, L.; Fuentes, V.; Gili, J.M. Guía de Identificación de Medusas y Otros Organismos Gelatinosos; CSIC—Instituto de Ciencias del Mar (ICM): Barcelona, Spain, 2021. [Google Scholar] [CrossRef]
- Purcell, J.E.; Atienza, D.; Fuentes, V.; Olariaga, A.; Tilves, U.; Colahan, C.; Gili, J.-M. Temperature Effects on Asexual Reproduction Rates of Scyphozoan Species from the Northwest Mediterranean Sea. Hydrobiologia 2012, 690, 169–180. [Google Scholar] [CrossRef]
- Kikinger, R. Cotylorhiza Tuberculata (Cnidaria: Scyphozoa)—Life History of a Stationary Population. Mar. Ecol. 1992, 13, 333–362. [Google Scholar] [CrossRef]
- İşinibilir, M.; Yüksel, E.; Dalyan, C. First Record of Cotylorhiza Tuberculata (Macri, 1778) from the Sea of Marmara. Aquat. Sci. Eng. 2020, 36, 38–41. [Google Scholar] [CrossRef]
- Aouititen, M.; Ravibhanu, A.; Ang, S.C.; Mouanda, D.C.M.; Luan, X. New Records of Two Jellyfish Species Rhizostoma Luteum (Quoy and Gaimard 1827) and Cotylorhiza Tuberculata (Macri 1778) in the Moroccan Northwest Mediterranean Coast. Discov. Life 2024, 54, 5. [Google Scholar] [CrossRef]
- Lotan, A.; Ben-Hillel, R.; Loya, Y. Life Cycle of Rhopilema nomadica: A New Immigrant Scyphomedusan in the Mediterranean. Mar. Biol. 1992, 112, 237–242. [Google Scholar] [CrossRef]
- Galil, B.S. A Sea under Siege—Alien Species in the Mediterranean. Biol. Invasions 2020, 2, 177–186. [Google Scholar] [CrossRef]
- Kuplik, Z.; Kerem, D.; Angel, D.L. Respiration Rates, Metabolic Demands and Feeding of Ephyrae and Young Medusae of the Rhizostome Rhopilema Nomadica. Diversity 2021, 13, 320. [Google Scholar] [CrossRef]
- You, K.; Ma, C.; Gao, H.; Li, F.; Zhang, M.; Qiu, Y.; Wang, B. Research on the Jellyfish (Rhopilema Esculentum Kishinouye) and Associated Aquaculture Techniques in China: Current Status. Aquac. Int. 2007, 15, 479–488. [Google Scholar] [CrossRef]
- Kienberger, K.; Prieto, L. The Jellyfish Rhizostoma Luteum (Quoy & Gaimard, 1827): Not Such a Rare Species after All. Mar. Biodivers. 2018, 48, 1455–1462. [Google Scholar] [CrossRef]
- Fernández-Alías, A.; Quispe-Becerra, J.I.; Conde-Caño, M.R.; Marcos, C.; Pérez-Ruzafa, A. Mediterranean Biogeography, Colonization, Expansion, Phenology, and Life Cycle of the Invasive Jellyfish Phyllorhiza Punctata von Lendenfeld, 1884. Estuar. Coast. Shelf Sci. 2024, 299, 108699. [Google Scholar] [CrossRef]
- Calejo, M.T.; Morais, Z.B.; Fernandes, A.I. Isolation and Biochemical Characterisation of a Novel Collagen from Catostylus Tagi. J. Biomater. Sci. Polym. Ed. 2009, 20, 2073–2087. [Google Scholar] [CrossRef]
- James, S.; Tilvi, S.; Khandeparker, R.; Sreepada, R.A.; Thakur, N.; Gauthankar, M. Jellyfish Rhizostoma Pulmo Collected off Goa Coast (India) as a Rich Source of Tryptophan Containing Collagen and Its Enhanced Antioxidant Potential. J. Food Sci. Technol. 2023, 60, 2825–2834. [Google Scholar] [CrossRef]
- Khong, N.M.H.; Yusoff, F.M.; Jamilah, B.; Basri, M.; Maznah, I.; Chan, K.W.; Armania, N.; Nishikawa, J. Improved Collagen Extraction from Jellyfish (Acromitus Hardenbergi) with Increased Physical-Induced Solubilization Processes. Food Chem. 2018, 251, 41–50. [Google Scholar] [CrossRef]
- Rastian, Z.; Pütz, S.; Wang, Y.; Kumar, S.; Fleissner, F.; Weidner, T.; Parekh, S.H. Type I Collagen from Jellyfish Catostylus Mosaicus for Biomaterial Applications. ACS Biomater. Sci. Eng. 2018, 4, 2115–2125. [Google Scholar] [CrossRef]
- Barzkar, N.; Sukhikh, S.; Zhikhreva, A.; Cheliubeeva, E.; Kapitunova, A.; Malkov, D.; Babich, O.; Kulikova, Y. Aurelia Aurita Jellyfish Collagen: Recovery Properties. Foods Raw Mater. 2024, 13, 296–305. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, R.; Huang, L.; Song, Y.; Regenstein, J.M. Characterisation of Acid-Soluble and Pepsin-Solubilised Collagen from Jellyfish (Cyanea Nozakii Kishinouye). Food Chem. 2014, 150, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Gorbatenko, K.M.; Nikolayev, A.V.; Figurkin, A.L.; Il’inskii, E.N. Quantitative Composition, Distribution, and Feeding of Large Jellyfish (Scyphozoa et Hydrozoa) on the West Kamchatka Shelf in Summer. Russ. J. Mar. Biol. 2009, 35, 579–592. [Google Scholar] [CrossRef]
- Balikci, E.; Baran, E.T.; Tahmasebifar, A.; Yilmaz, B. Characterization of Collagen from Jellyfish Aurelia Aurita and Investigation of Biomaterials Potentials. Appl. Biochem. Biotechnol. 2024, 196, 6200–6221. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Shao, Z.; Li, C.; Yu, L.; Raja, M.A.; Liu, C. Isolation, Characterization and Evaluation of Collagen from Jellyfish Rhopilema Esculentum Kishinouye for Use in Hemostatic Applications. PLoS ONE 2017, 12, e0169731. [Google Scholar] [CrossRef]
- Nagai, T.; Worawattanamateekul, W.; Suzuki, N.; Nakamura, T.; Ito, T.; Fujiki, K.; Nakao, M.; Yano, T. Isolation and Characterization of Collagen from Rhizostomous Jellyfish (Rhopilema Asamushi). Food Chem. 2000, 70, 205–208. [Google Scholar] [CrossRef]
- Shaik, M.I.; Rahman, S.H.A.; Yusri, A.S.; Ismail-Fitry, M.R.; Kumar, N.S.S.; Sarbon, N.M. A Review on the Processing Technique, Physicochemical, and Bioactive Properties of Marine Collagen. J. Food Sci. 2024, 89, 5205–5229. [Google Scholar] [CrossRef]
- Sudirman, S.; Chen, C.-Y.; Chen, C.-K.; Felim, J.; Kuo, H.-P.; Kong, Z.-L. Fermented Jellyfish (Rhopilema Esculentum) Collagen Enhances Antioxidant Activity and Cartilage Protection on Surgically Induced Osteoarthritis in Obese Rats. Front. Pharmacol. 2023, 14, 1117893. [Google Scholar] [CrossRef]
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of Collagen and Collagen Hydrolysate from Jellyfish (Rhopilema Esculentum) on Mice Skin Photoaging Induced by UV Irradiation. J. Food Sci. 2009, 74, H183–H188. [Google Scholar] [CrossRef]
- Morelli, S.; D’Amora, U.; Piscioneri, A.; Oliviero, M.; Scialla, S.; Coppola, A.; De Pascale, D.; Crocetta, F.; De Santo, M.P.; Davoli, M.; et al. Methacrylated Chitosan/Jellyfish Collagen Membranes as Cell Instructive Platforms for Liver Tissue Engineering. Int. J. Biol. Macromol. 2024, 281, 136313. [Google Scholar] [CrossRef] [PubMed]
- Fassini, D.; Wilkie, I.C.; Pozzolini, M.; Ferrario, C.; Sugni, M.; Rocha, M.S.; Giovine, M.; Bonasoro, F.; Silva, T.H.; Reis, R.L. Diverse and Productive Source of Biopolymer Inspiration: Marine Collagens. Biomacromolecules 2021, 22, 1815–1834. [Google Scholar] [CrossRef] [PubMed]
- Farooq, S.; Ahmad, M.I.; Zheng, S.; Ali, U.; Li, Y.; Shixiu, C.; Zhang, H. A Review on Marine Collagen: Sources, Extraction Methods, Colloids Properties, and Food Applications. Collagen Leather 2024, 6, 11. [Google Scholar] [CrossRef]
- Song, E.; Yeon Kim, S.; Chun, T.; Byun, H.-J.; Lee, Y.M. Collagen Scaffolds Derived from a Marine Source and Their Biocompatibility. Biomaterials 2006, 27, 2951–2961. [Google Scholar] [CrossRef]
- Prajaputra, V.; Isnaini, N.; Maryam, S.; Ernawati, E.; Deliana, F.; Haridhi, H.A.; Fadli, N.; Karina, S.; Agustina, S.; Nurfadillah, N.; et al. Exploring Marine Collagen: Sustainable Sourcing, Extraction Methods, and Cosmetic Applications. S. Afr. J. Chem. Eng. 2024, 47, 197–211. [Google Scholar] [CrossRef]
- Lu, W.-C.; Chiu, C.-S.; Chan, Y.-J.; Mulio, A.T.; Li, P.-H. Characterization and Biological Properties of Marine By-Product Collagen through Ultrasound-Assisted Extraction. Aquac. Rep. 2023, 29, 101514. [Google Scholar] [CrossRef]
- Calejo, M.T.; Almeida, A.J.; Fernandes, A.I. Exploring a New Jellyfish Collagen in the Production of Microparticles for Protein Delivery. J. Microencapsul. 2012, 29, 520–531. [Google Scholar] [CrossRef]
- León-Campos, M.I.; Claudio-Rizo, J.A.; Rodriguez-Fuentes, N.; Cabrera-Munguía, D.A.; Becerra-Rodriguez, J.J.; Herrera-Guerrero, A.; Soriano-Corral, F. Biocompatible Interpenetrating Polymeric Networks in Hydrogel State Comprised from Jellyfish Collagen and Polyurethane. J. Polym. Res. 2021, 28, 291. [Google Scholar] [CrossRef]
- Arslan, Y.E.; Sezgin Arslan, T.; Derkus, B.; Emregul, E.; Emregul, K.C. Fabrication of Human Hair Keratin/Jellyfish Collagen/Eggshell-Derived Hydroxyapatite Osteoinductive Biocomposite Scaffolds for Bone Tissue Engineering: From Waste to Regenerative Medicine Products. Colloids Surf. B Biointerfaces 2017, 154, 160–170. [Google Scholar] [CrossRef]
- Barros, A.A.; Aroso, I.M.; Silva, T.H.; Mano, J.F.; Duarte, A.R.C.; Reis, R.L. Water and Carbon Dioxide: Green Solvents for the Extraction of Collagen/Gelatin from Marine Sponges. ACS Sustain. Chem. Eng. 2015, 3, 254–260. [Google Scholar] [CrossRef]
- Silva, J.C.; Barros, A.A.; Aroso, I.M.; Fassini, D.; Silva, T.H.; Reis, R.L.; Duarte, A.R.C. Extraction of Collagen/Gelatin from the Marine Demosponge Chondrosia Reniformis (Nardo, 1847) Using Water Acidified with Carbon Dioxide—Process Optimization. Ind. Eng. Chem. Res. 2016, 55, 6922–6930. [Google Scholar] [CrossRef]
- Hansen, B.B.; Spittle, S.; Chen, B.; Poe, D.; Zhang, Y.; Klein, J.M.; Horton, A.; Adhikari, L.; Zelovich, T.; Doherty, B.W.; et al. Deep Eutectic Solvents: A Review of Fundamentals and Applications. Chem. Rev. 2021, 121, 1232–1285. [Google Scholar] [CrossRef]
- Bisht, M.; Martins, M.; Dias, A.C.R.V.; Ventura, S.P.M.; Coutinho, J.A.P. Uncovering the Potential of Aqueous Solutions of Deep Eutectic Solvents on the Extraction and Purification of Collagen Type I from Atlantic Codfish (Gadus morhua). Green Chem. 2021, 23, 8940–8948. [Google Scholar] [CrossRef]
- Batista, M.P.; Fernández, N.; Gaspar, F.B.; Bronze, M.d.R.; Duarte, A.R.C. Extraction of Biocompatible Collagen From Blue Shark Skins Through the Conventional Extraction Process Intensification Using Natural Deep Eutectic Solvents. Front. Chem. 2022, 10, 937036. [Google Scholar] [CrossRef]
- Raposo, A.; Alasqah, I.; Alfheeaid, H.; Alsharari, Z.; Alturki, H.; Raheem, D. Jellyfish as Food: A Narrative Review. Foods 2022, 11, 2773. [Google Scholar] [CrossRef] [PubMed]
- De Domenico, S.; De Rinaldis, G.; Paulmery, M.; Piraino, S.; Leone, A. Barrel Jellyfish (Rhizostoma Pulmo) as Source of Antioxidant Peptides. Mar. Drugs 2019, 17, 134. [Google Scholar] [CrossRef]
- Leone, A.; Lecci, R.; Durante, M.; Meli, F.; Piraino, S. The Bright Side of Gelatinous Blooms: Nutraceutical Value and Antioxidant Properties of Three Mediterranean Jellyfish (Scyphozoa). Mar. Drugs 2015, 13, 4654–4681. [Google Scholar] [CrossRef]
- Prieto, L.; Enrique-Navarro, A.; Li Volsi, R.; Ortega, M. The Large Jellyfish Rhizostoma Luteum as Sustainable a Resource for Antioxidant Properties, Nutraceutical Value and Biomedical Applications. Mar. Drugs 2018, 16, 396. [Google Scholar] [CrossRef]
- Alkayali, A. Hydrolyzed Jellyfish Collagen Types I, II, and V and Use Thereof. U.S. Patent US10287339B2, 14 May 2019. [Google Scholar]
- Omori, M.; Nakano, E. Jellyfish Fisheries in Southeast Asia. In Jellyfish Blooms: Ecological and Societal Importance; Purcell, J.E., Graham, W.M., Dumont, H.J., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 19–26. ISBN 978-94-010-3835-5. [Google Scholar]
- Brotz, L.; Schiariti, A.; López-Martínez, J.; Álvarez-Tello, J.; Peggy Hsieh, Y.-H.; Jones, R.P.; Quiñones, J.; Dong, Z.; Morandini, A.C.; Preciado, M.; et al. Jellyfish Fisheries in the Americas: Origin, State of the Art, and Perspectives on New Fishing Grounds. Rev. Fish Biol. Fish. 2017, 27, 1–29. [Google Scholar] [CrossRef]
- Brotz, L. Jellyfish Fisheries in the World. Ph.D. Thesis, The University of British Columbia, Vancouver, BC, Canada, 2016. [Google Scholar]
- Kumawat, T.; Saravanan, R.; Vinod, K.; Jaiswar, A.K.; Deshmukhe, G.; Shenoy, L.; Divu, D.; Joshi, K.K.; Gopalakrishnan, A. Jellyfish Fisheries along the Gujarat Coast, India: Status and Challenges. Mar. Policy 2023, 150, 105554. [Google Scholar] [CrossRef]
- Nishikawa, J.; Thu, N.T.; Ha, T.M.; Thu, P.T. Jellyfish Fisheries in Northern Vietnam. Plankton Benthos Res. 2008, 3, 227–234. [Google Scholar] [CrossRef]
- Torri, L.; Tuccillo, F.; Bonelli, S.; Piraino, S.; Leone, A. The Attitudes of Italian Consumers towards Jellyfish as Novel Food. Food Qual. Prefer. 2020, 79, 103782. [Google Scholar] [CrossRef]
- Leone, A.; Lecci, R.M.; Milisenda, G.; Piraino, S. Mediterranean Jellyfish as Novel Food: Effects of Thermal Processing on Antioxidant, Phenolic, and Protein Contents. Eur. Food Res. Technol. 2019, 245, 1611–1627. [Google Scholar] [CrossRef]
- Leone, A. European Jellyfish Cookbook—Prime Ricette a Base Di Meduse in Stile Occidentale; © Cnr Edizioni: Rome, Italy, 2020; ISBN 978-88-8080-395-9. [Google Scholar]
- Jadach, B.; Mielcarek, Z.; Osmałek, T. Use of Collagen in Cosmetic Products. Curr. Issues Mol. Biol. 2024, 46, 2043–2070. [Google Scholar] [CrossRef]
- Amirrah, I.N.; Lokanathan, Y.; Zulkiflee, I.; Wee, M.F.M.R.; Motta, A.; Fauzi, M.B. A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold. Biomedicines 2022, 10, 2307. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Powell, L.C.; Matin, N.; Mearns-Spragg, A.; Thornton, C.A.; Khan, I.M.; Francis, L.W. Jellyfish Collagen: A Biocompatible Collagen Source for 3d Scaffold Fabrication and Enhanced Chondrogenicity. Mar. Drugs 2021, 19, 405. [Google Scholar] [CrossRef]
- Cadar, E.; Pesterau, A.-M.; Sirbu, R.; Negreanu-Pirjol, B.S.; Tomescu, C.L. Jellyfishes—Significant Marine Resources with Potential in the Wound-Healing Process: A Review. Mar. Drugs 2023, 21, 201. [Google Scholar] [CrossRef]
- Rachmawati, R.; Hidayat, M.; Permatasari, N.; Widyarti, S. Jellyfish (Aurelia Aurita) Collagen Scaffolds Potential in Alveolar Bone Regeneration. F1000Research 2021, 10, 318. [Google Scholar] [CrossRef]
- Widdowson, J.P.; Picton, A.J.; Vince, V.; Wright, C.J.; Mearns-Spragg, A. In Vivo Comparison of Jellyfish and Bovine Collagen Sponges as Prototype Medical Devices. J. Biomed. Mater. Res. Part B Appl. Biomater. 2018, 106, 1524–1533. [Google Scholar] [CrossRef]
- Paradiso, F.; Fitzgerald, J.; Yao, S.; Barry, F.; Taraballi, F.; Gonzalez, D.; Conlan, R.S.; Francis, L. Marine Collagen Substrates for 2D and 3D Ovarian Cancer Cell Systems. Front. Bioeng. Biotechnol. 2019, 7, 343. [Google Scholar] [CrossRef]
- Flaig, I.; Radenković, M.; Najman, S.; Pröhl, A.; Jung, O.; Barbeck, M. In Vivo Analysis of the Biocompatibility and Immune Response of Jellyfish Collagen Scaffolds and Its Suitability for Bone Regeneration. Int. J. Mol. Sci. 2020, 21, 4518. [Google Scholar] [CrossRef] [PubMed]
- Mearns-Spragg, A.; Tilman, J.; Tams, D.; Barnes, A. The Biological Evaluation of Jellyfish Collagen as a New Research Tool for the Growth and Culture of iPSC Derived Microglia. Front. Mar. Sci. 2020, 7, 689. [Google Scholar] [CrossRef]
- Riacci, L.; Sorriento, A.; Ricotti, L. Genipin-Based Crosslinking of Jellyfish Collagen 3D Hydrogels. Gels 2021, 7, 238. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Cervantes, I.; Rodríguez-Fuentes, N.; León-Deniz, L.V.; Alcántara Quintana, L.E.; Cervantes-Uc, J.M.; Herrera Kao, W.A.; Cerón-Espinosa, J.D.; Cauich-Rodríguez, J.V.; Castaño-Meneses, V.M. Cell-Free Scaffold from Jellyfish Cassiopea Andromeda (Cnidaria; Scyphozoa) for Skin Tissue Engineering. Mater. Sci. Eng. C 2020, 111, 110748. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; Sarhan, E.M.; Ibrahim, A.A.; Abo-Elwafa, A.H.; Yahia, A.M.; Salah, A.; Maher, R.M.; Wagdy, M.; El-Sherbeny, G.A.; El-Sawah, A.A. One-Step Green Synthesis of Collagen Nanoparticles Using Ulva Fasciata, Network Pharmacology and Functional Enrichment Analysis in Hepatocellular Carcinoma Treatment. Int. J. Biol. Macromol. 2025, 294, 139244. [Google Scholar] [CrossRef]
- Charoenchokpanich, W.; Muangrod, P.; Roytrakul, S.; Rungsardthong, V.; Wonganu, B.; Charoenlappanit, S.; Casanova, F.; Thumthanaruk, B. Exploring the Model of Cefazolin Released from Jellyfish Gelatin-Based Hydrogels as Affected by Glutaraldehyde. Gels 2024, 10, 271. [Google Scholar] [CrossRef]
- Alkildani, S.; Jung, O.; Barbeck, M. In Vitro Investigation of Jellyfish Collagen as a Tool in Cell Culture and (Bone) Tissue Engineering. Anticancer Res. 2021, 41, 707–717. [Google Scholar] [CrossRef]
- Carvalho, D.N.; Williams, D.S.; Sotelo, C.G.; Pérez-Martín, R.I.; Mearns-Spragg, A.; Reis, R.L.; Silva, T.H. Marine Origin Biomaterials Using a Compressive and Absorption Methodology as Cell-Laden Hydrogel Envisaging Cartilage Tissue Engineering. Biomater. Adv. 2022, 137, 212843. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liu, L.; Xiong, X.; Krastev, R.; Smeets, R.; Rimashevskiy, D.; Schnettler, R.; Alkildani, S.; Emmert, S.; Jung, O.; et al. Suitability of R. Pulmo Jellyfish-Collagen-Coated Well Plates for Cytocompatibility Analyses of Biomaterials. Int. J. Mol. Sci. 2023, 24, 3007. [Google Scholar] [CrossRef]
- Rigogliuso, S.; Salamone, M.; Barbarino, E.; Barbarino, M.; Nicosia, A.; Ghersi, G. Production of Injectable Marine Collagen-Based Hydrogel for the Maintenance of Differentiated Chondrocytes in Tissue Engineering Applications. Int. J. Mol. Sci. 2020, 21, 5798. [Google Scholar] [CrossRef]
- Salthouse, D.; Goulding, P.D.; Reay, S.L.; Jackson, E.L.; Xu, C.; Ahmed, R.; Mearns-Spragg, A.; Novakovic, K.; Hilkens, C.M.U.; Ferreira, A.M. Amine-Reactive Crosslinking Enhances Type 0 Collagen Hydrogel Properties for Regenerative Medicine. Front. Bioeng. Biotechnol. 2024, 12, 1391728. [Google Scholar] [CrossRef] [PubMed]
- León-Campos, M.I.; Rodríguez-Fuentes, N.; Claudio-Rizo, J.A.; Cabrera-Munguía, D.A.; Becerra-Rodríguez, J.J.; Herrera-Guerrero, A.; Soriano-Corral, F.; Alcántara-Quintana, L.E. Development and in Vitro Evaluation of a Polymeric Matrix of Jellyfish Collagen-Human Stem Cell Secretome-Polyurethane for Wound Healing. J. Mater. Sci. 2023, 58, 8047–8060. [Google Scholar] [CrossRef]
- Brotz, L.; Cisneros-Montemayor, A.M.; Cisneros-Mata, M.Á. The Race for Jellyfish: Winners and Losers in Mexico’s Gulf of California. Mar. Policy 2021, 134, 104775. [Google Scholar] [CrossRef]
- María Esther, C.-C.; Miguel Ángel, C.-M.; Gabriela, M.-M.; Ileana, E.; Andrés Miguel, C.-M.; Luis, M.-C. Analysis of the Gulf of California Cannonball Jellyfish Fishery as a Complex System. Ocean Coast. Manag. 2021, 207, 105610. [Google Scholar] [CrossRef]
- Kitamura, M.; Omori, M. Synopsis of Edible Jellyfishes Collected from Southeast Asia, with Notes on Jellyfish Fisheries. Plankton Benthos Res. 2010, 5, 106–118. [Google Scholar] [CrossRef]
- Behera, P.R.; Raju, S.S.; Jishnudev, M.A.; Ghosh, S.; Saravanan, R. Emerging Jellyfish Fisheries along Central South East Coast of India. Ocean Coast. Manag. 2020, 191, 105183. [Google Scholar] [CrossRef]
- Jia, C.; Fujimori, Y.; Wang, X.; Guan, C. Performance of Large-Scale Stow Nets for Investigating Jellyfish. Fish Sci. 2023, 89, 595–603. [Google Scholar] [CrossRef]
- Edelist, D.; Angel, D.L.; Canning-Clode, J.; Gueroun, S.K.M.; Aberle, N.; Javidpour, J.; Andrade, C. Jellyfishing in Europe: Current Status, Knowledge Gaps, and Future Directions towards a Sustainable Practice. Sustainability 2021, 13, 12445. [Google Scholar] [CrossRef]
- Elliott, A.; Hobson, V.; Tang, K.W. Balancing Fishery and Conservation: A Case Study of the Barrel Jellyfish Rhizostoma Octopus in South Wales. ICES J. Mar. Sci. 2017, 74, 234–241. [Google Scholar] [CrossRef]
- Jellagen. Available online: https://jellagen.co.uk/ (accessed on 3 March 2025).
- You, K.; Bian, Y.; Ma, C.; Chi, X.; Liu, Z.; Zhang, Y. Study on the Carry Capacity of Edible Jellyfish Fishery in Liaodong Bay. J. Ocean Univ. China 2016, 15, 471–479. [Google Scholar] [CrossRef]
- Mghili, B.; Analla, M.; Aksissou, M. Estimating the Economic Damage Caused by Jellyfish to Fisheries in Morocco. Afr. J. Mar. Sci. 2022, 44, 271–277. [Google Scholar] [CrossRef]
- Palmieri, M.G.; Barausse, A.; Luisetti, T.; Turner, K. Jellyfish Blooms in the Northern Adriatic Sea: Fishermen’s Perceptions and Economic Impacts on Fisheries. Fish. Res. 2014, 155, 51–58. [Google Scholar] [CrossRef]
- Conley, K.R.; Sutherland, K.R. Commercial Fishers’ Perceptions of Jellyfish Interference in the Northern California Current. ICES J. Mar. Sci. 2015, 72, 1565–1575. [Google Scholar] [CrossRef]
- D’Ambra, I.; Merquiol, L. Jellyfish from Fisheries By-Catches as a Sustainable Source of High-Value Compounds with Biotechnological Applications. Mar. Drugs 2022, 20, 266. [Google Scholar] [CrossRef]
- COLMED: Colágeno de Medusas, Un Recurso Marino Innovador Del Mediterráneo En Biotecnología Azul|Pleamar. Available online: https://www.programapleamar.es/proyectos/colmed-colageno-de-medusas-un-recurso-marino-innovador-del-mediterraneo-en-biotecnologia (accessed on 3 March 2025).
- Fernández-Alías, A.; Marcos, C.; Pérez-Ruzafa, A. The Unpredictability of Scyphozoan Jellyfish Blooms. Front. Mar. Sci. 2024, 11, 1349956. [Google Scholar] [CrossRef]
- Purcell, J.E.; Baxter, E.J.; Fuentes, V.L. Jellyfish as Products and Problems of Aquaculture. In Advances in Aquaculture Hatchery Technology; Elsevier: Amsterdam, The Netherlands, 2013; pp. 404–430. ISBN 978-0-85709-119-2. [Google Scholar]
- Duarte, I.M.; Marques, S.C.; Leandro, S.M.; Calado, R. An Overview of Jellyfish Aquaculture: For Food, Feed, Pharma and Fun. Rev. Aquac. 2022, 14, 265–287. [Google Scholar] [CrossRef]
- Ballesteros, A.; Páez, D.; Santín, A.; García, A.; Martín, Y.; Alonso, E.; Jourdan, E.; Gili, J.-M. Successful Culture of Pelagia Noctiluca (Cnidaria: Scyphozoa) Over Time: A Continuous Supply of the Holoplanktonic Jellyfish for Research and Industrial Applications. Front. Mar. Sci. 2022, 9, 911383. [Google Scholar] [CrossRef]
- De Domenico, S.; Toso, A.; De Rinaldis, G.; Mammone, M.; Fumarola, L.M.; Piraino, S.; Leone, A. Wild or Reared? Cassiopea Andromeda Jellyfish as a Potential Biofactory. Mar. Drugs 2025, 23, 19. [Google Scholar] [CrossRef]
- Dong, J.; Jiang, L.; Tan, K.; Liu, H.; Purcell, J.E.; Li, P.; Ye, C. Stock Enhancement of the Edible Jellyfish (Rhopilema Esculentum Kishinouye) in Liaodong Bay, China: A Review. Hydrobiologia 2009, 616, 113–118. [Google Scholar] [CrossRef]
- Yang, S.-Q.; Zhang, W.-C. The cultural technique of jellyfish. Fish Sci. 2004, 23, 25–26. [Google Scholar]
- JellyFarmer. Available online: https://jellyfarmer.com/ (accessed on 16 January 2025).
- Ballesteros, A.; Siles, P.; Jourdan, E.; Gili, J.-M. Versatile Aquarium for Jellyfish: A Rearing System for the Biomass Production of Early Life Stages in Flow-through or Closed Systems. Front. Mar. Sci. 2022, 9, 942094. [Google Scholar] [CrossRef]
- Duarte, I.; Leandro, S.; Ferreira, M.; Pinto, C.; Marques, D.; Falcão, J.; Maranhão, P.; Marques, S. Effect of Diet on Growth and Development of the Jellyfish Phyllorhiza Punctata (Cnidaria, Scyphozoa) in Laboratory Conditions. In Proceedings of the XX Iberian Symposium on Marine Biology Studies (SIEBM XX), Braga, Portugal, 9–12 September 2019; Volume 6. [Google Scholar]
- Raskoff, K.A.; Sommer, F.A.; Hamner, W.M.; Cross, K.M. Collection and Culture Techniques for Gelatinous Zooplankton. Biol. Bull. 2003, 204, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Crow, G.; Howard, M.; Levesque, V.; Matsushige, L.; Spina, S.; Schaadt, M.; Sowinski, N.; Widmer, C.; Upton, B. Jellyfish (Cnidaria/Ctenophora) Care Manual; Association of Zoos and Aquariums: Silver Spring, MD, USA, 2013. [Google Scholar] [CrossRef]
- Morata, D.P.M. Extraction, Analysis, and Use of Collagen from Cassiopea spp. (Cnidaria: Scyphozoa) in Biomedical Applications. Master’s Thesis, Catholic University of Valencia, Valencia, Spain, 2022. [Google Scholar]
- Pierce, J. A System for Mass Culture of Upside-down Jellyfish Cassiopea Spp as a Potential Food Item for Medusivores in Captivity. Int. Zoo Yearb. 2005, 39, 62–69. [Google Scholar] [CrossRef]
- Lisboa, R.; Gil, F.; Mascarenhas, P.; Morais, Z.; Morandini, A. Catostylus Tagi: Survival and Maintenance Trials of Planula and Polyps. Ann. Med. 2019, 51, 78. [Google Scholar] [CrossRef]
- Gueroun, S.K.M.; Torres, T.M.; Santos, A.D.; Vasco-Rodrigues, N.; Canning-Clode, J.; Andrade, C. Catostylus Tagi (Class: Scyphozoa, Order: Discomedusae, Suborder: Rhizostomida, Family: Catostylidae) Life Cycle and First Insight into Its Ecology. PeerJ 2021, 9, e12056. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballesteros, A.; Torres, R.; Pascual-Torner, M.; Revert-Ros, F.; Tena-Medialdea, J.; García-March, J.R.; Lloret, J.; Gili, J.-M. Jellyfish Collagen in the Mediterranean Spotlight: Transforming Challenges into Opportunities. Mar. Drugs 2025, 23, 200. https://doi.org/10.3390/md23050200
Ballesteros A, Torres R, Pascual-Torner M, Revert-Ros F, Tena-Medialdea J, García-March JR, Lloret J, Gili J-M. Jellyfish Collagen in the Mediterranean Spotlight: Transforming Challenges into Opportunities. Marine Drugs. 2025; 23(5):200. https://doi.org/10.3390/md23050200
Chicago/Turabian StyleBallesteros, Ainara, Raquel Torres, Maria Pascual-Torner, Francisco Revert-Ros, Jose Tena-Medialdea, José Rafael García-March, Josep Lloret, and Josep-Maria Gili. 2025. "Jellyfish Collagen in the Mediterranean Spotlight: Transforming Challenges into Opportunities" Marine Drugs 23, no. 5: 200. https://doi.org/10.3390/md23050200
APA StyleBallesteros, A., Torres, R., Pascual-Torner, M., Revert-Ros, F., Tena-Medialdea, J., García-March, J. R., Lloret, J., & Gili, J.-M. (2025). Jellyfish Collagen in the Mediterranean Spotlight: Transforming Challenges into Opportunities. Marine Drugs, 23(5), 200. https://doi.org/10.3390/md23050200