
Structure Diversity and Properties of Some Bola-like Natural Products




Abstract
1. Introduction
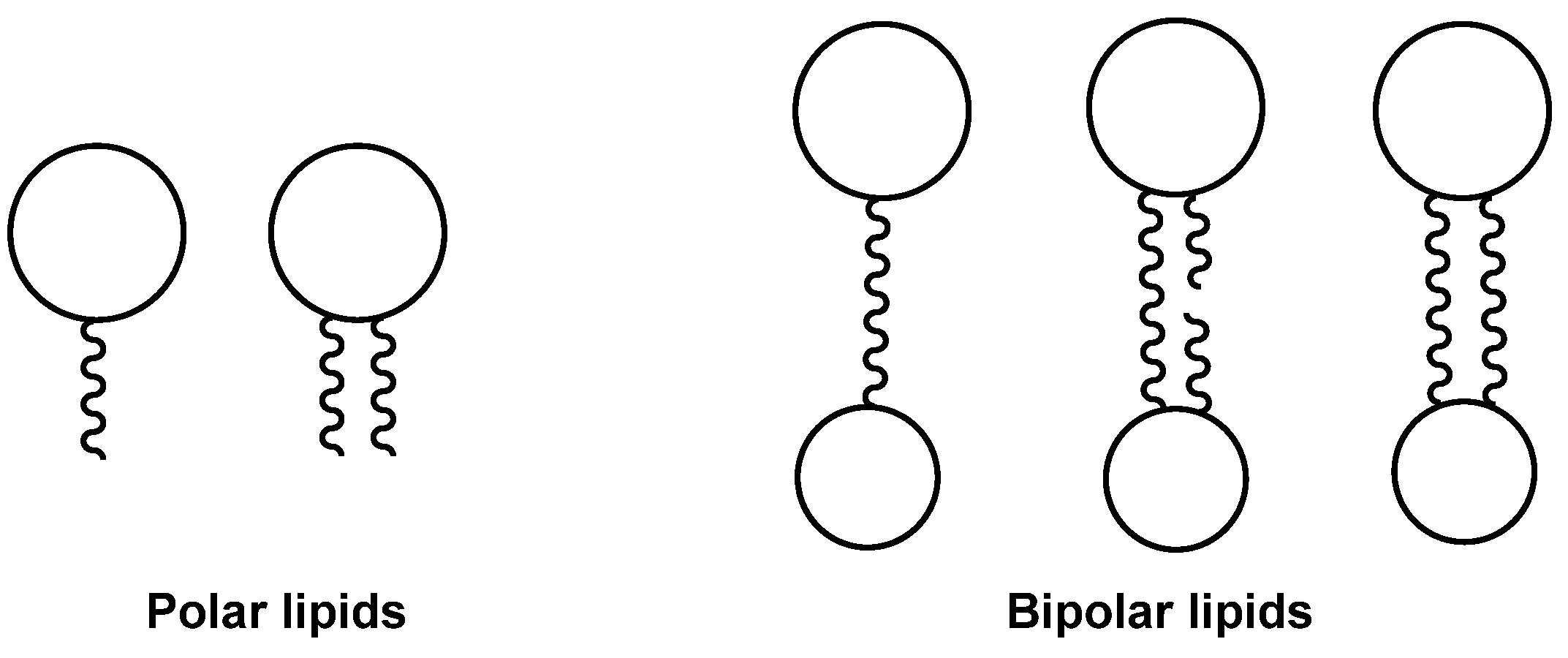
2. Bipolar Lipids of Archaea and Some Results of the Studies on Their Structures and Properties
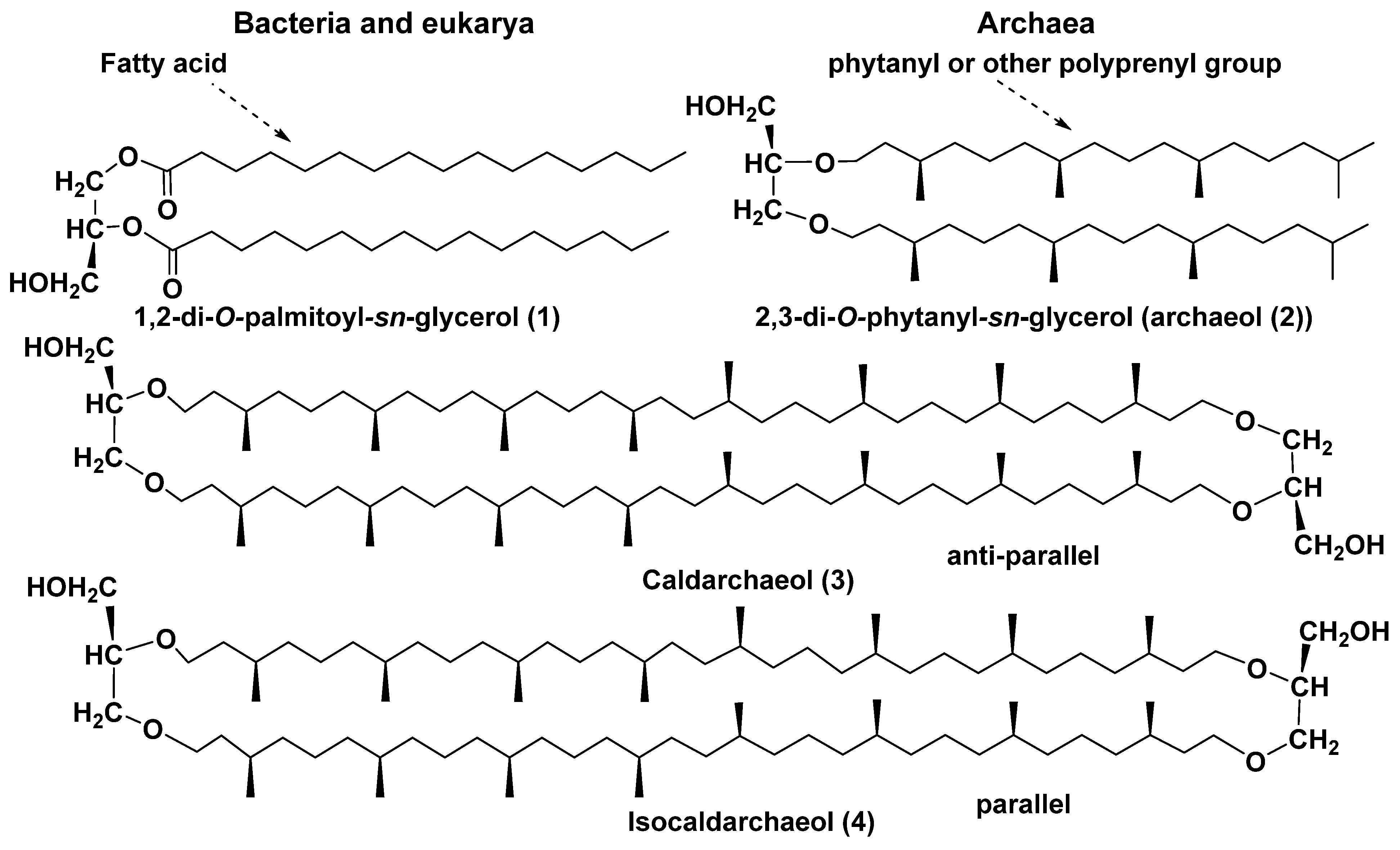
2.1. General Information About Archaea and Their Lipids
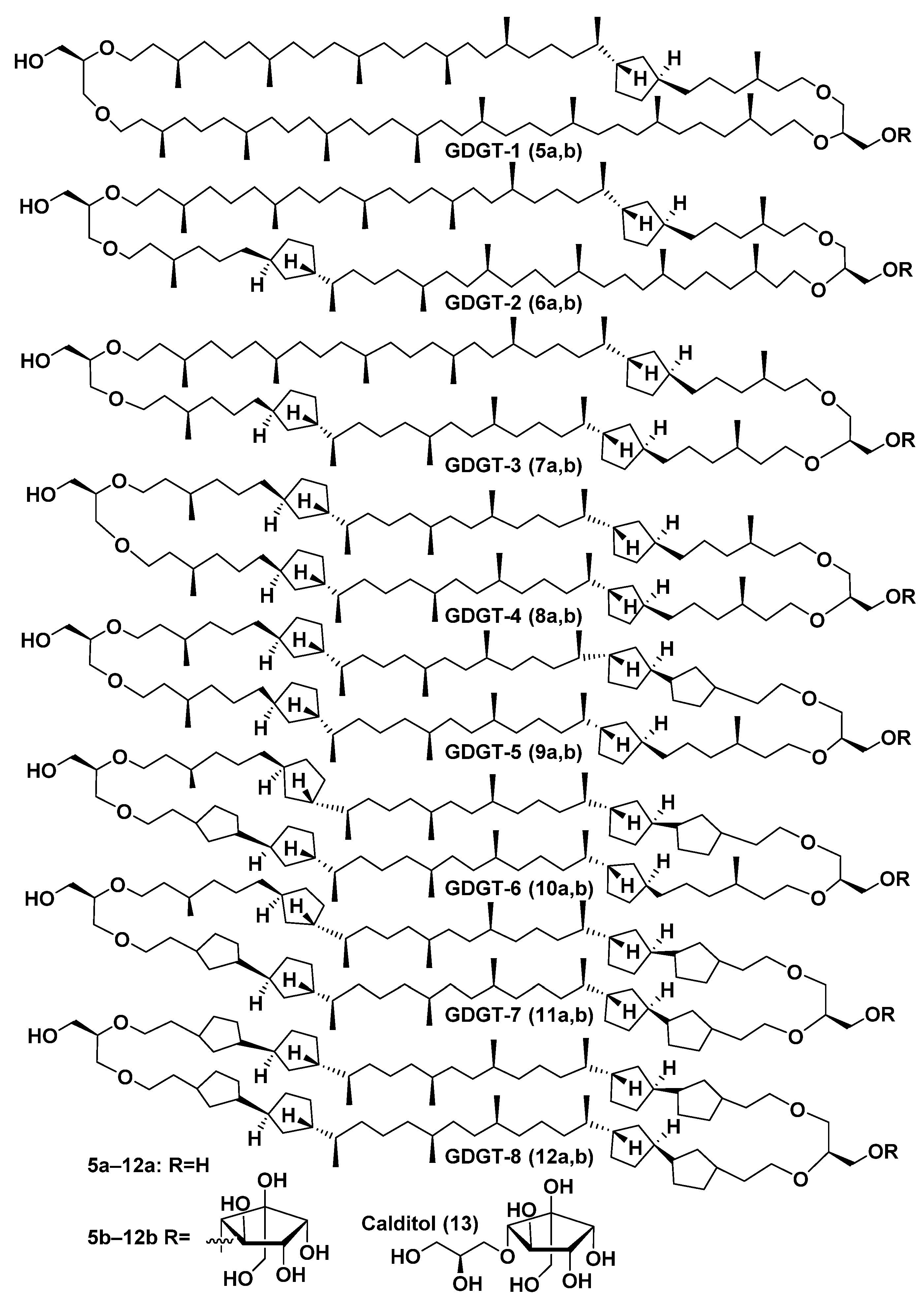
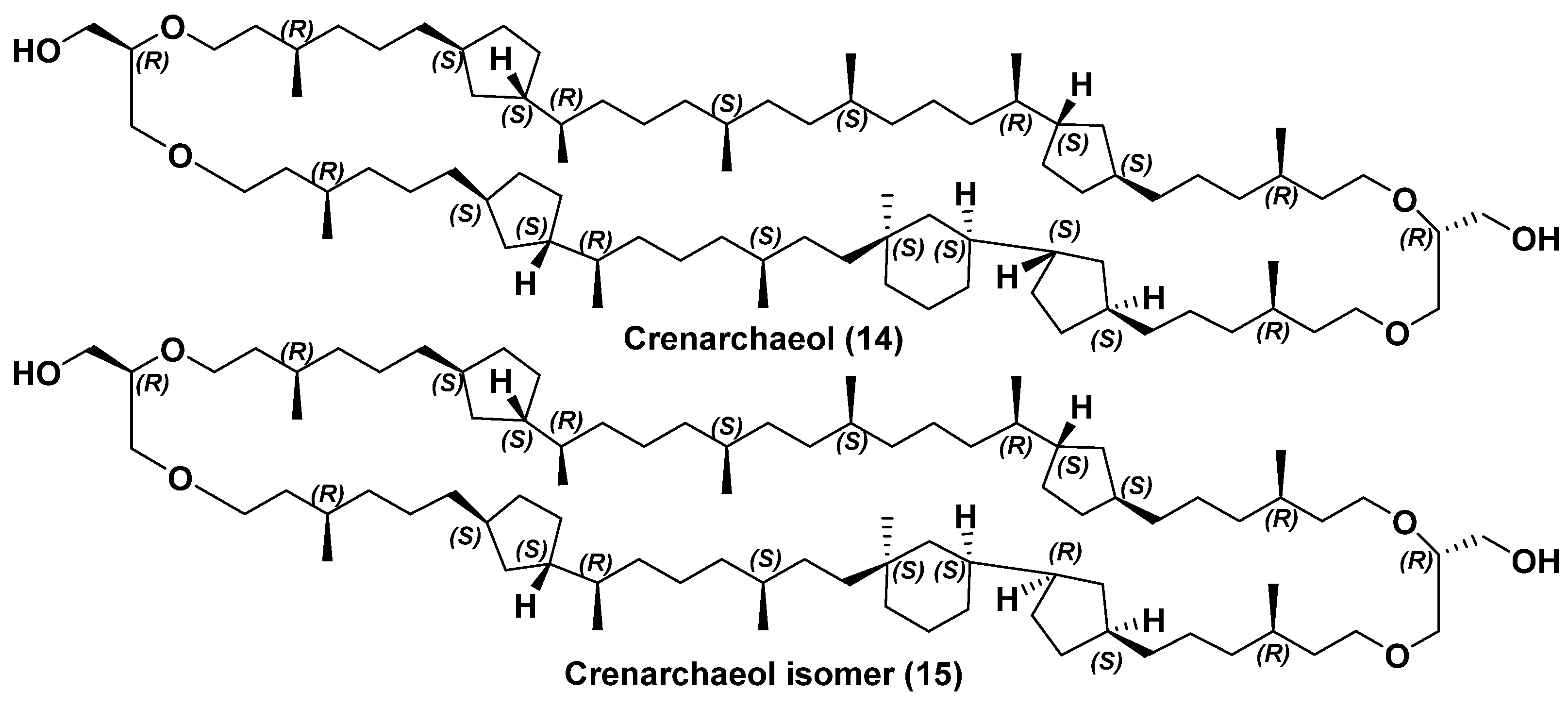
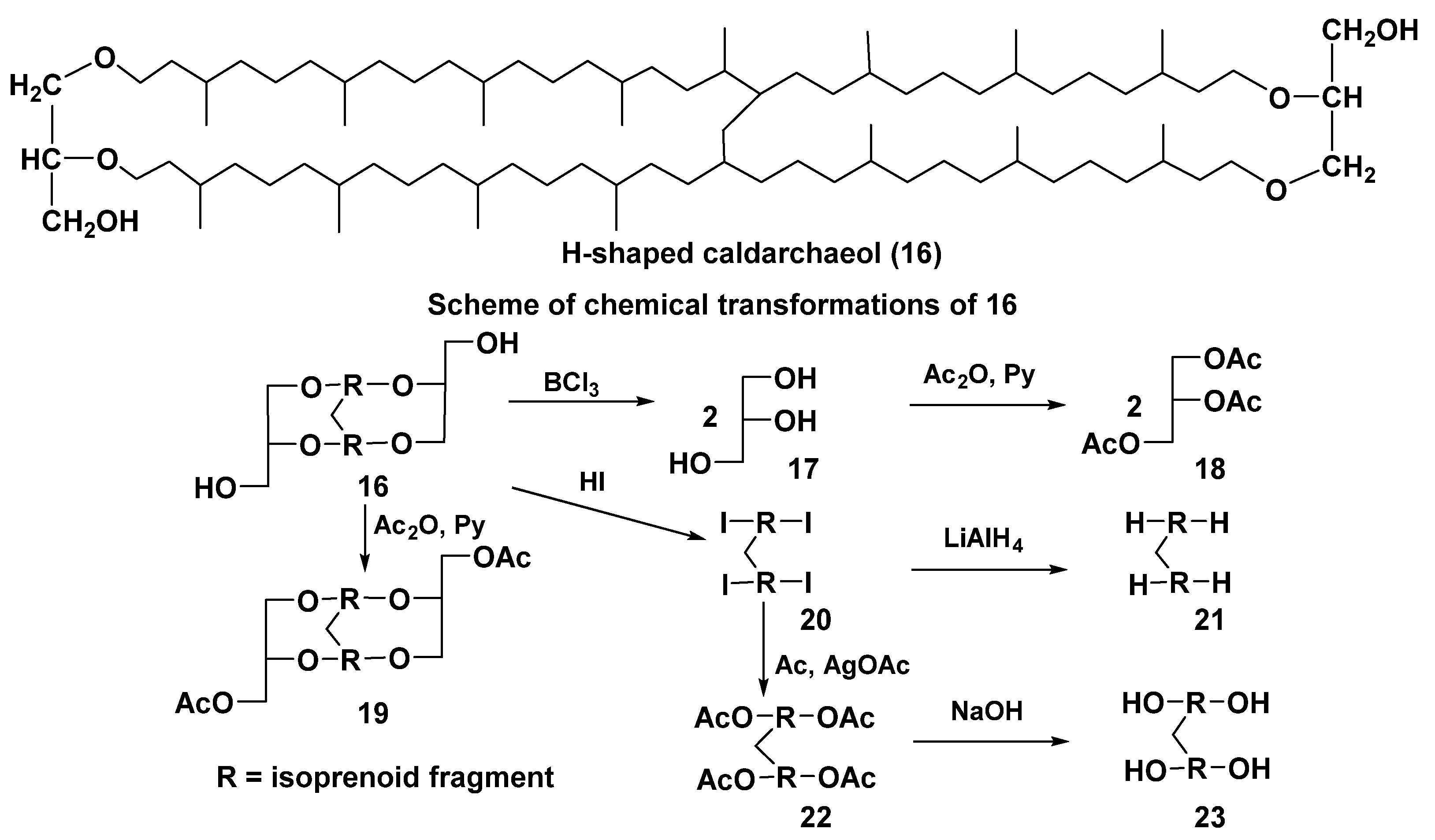
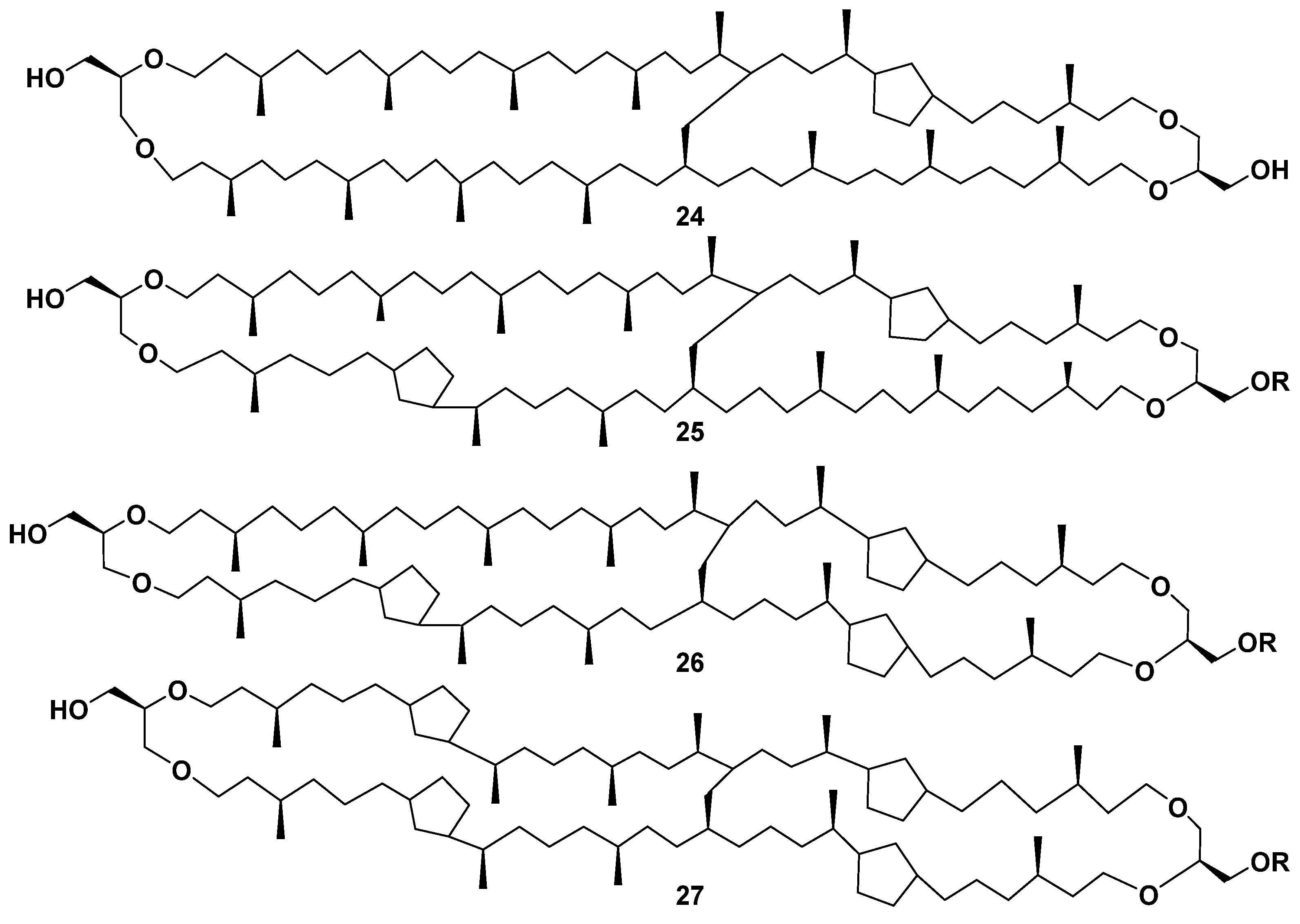
2.2. Structure and Taxonomic Distribution of Archaeal Bipolar Lipids with Emphasis on Recent Studies
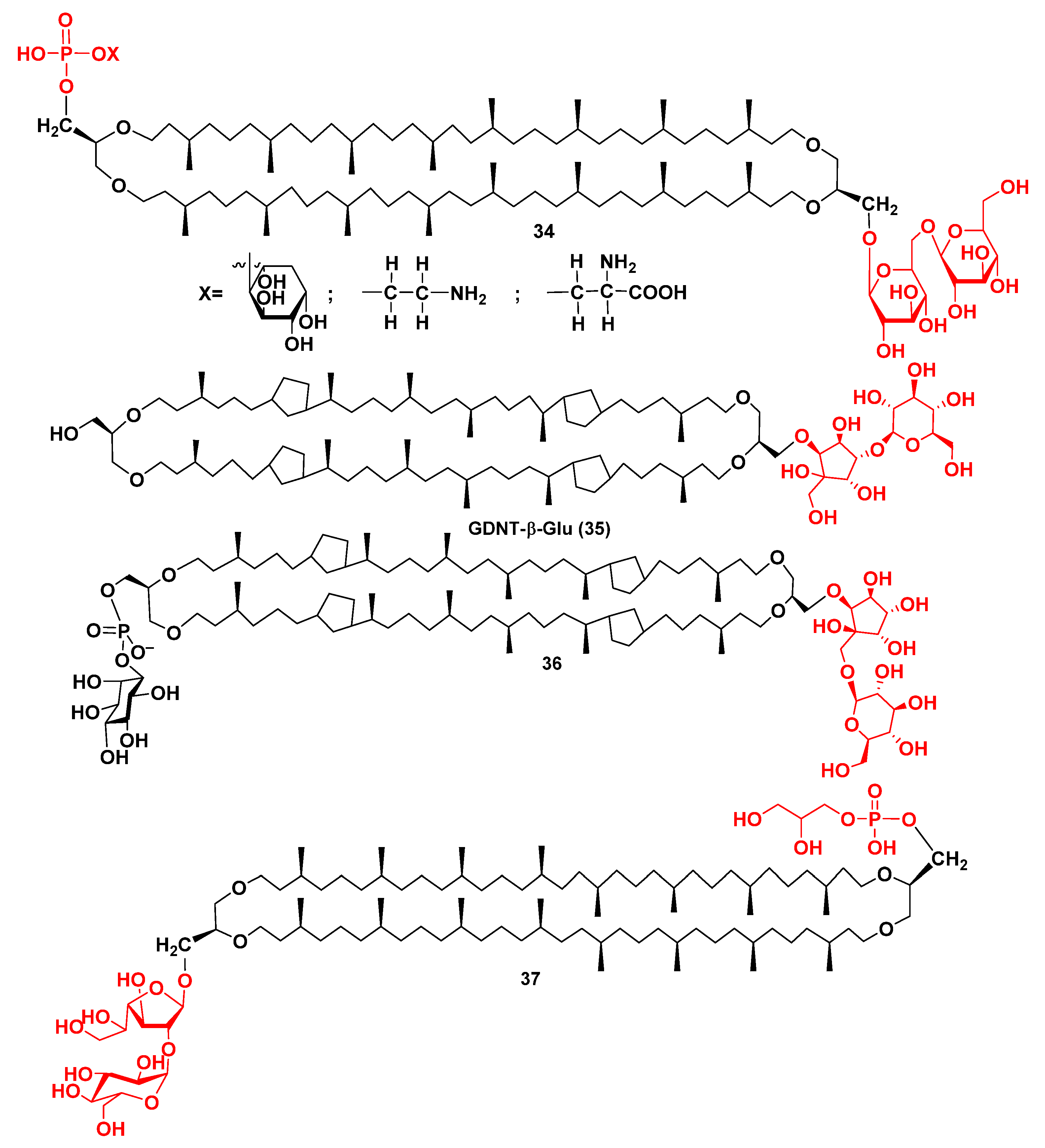
2.3. Intact Membrane Bipolar Lipids from Archaea
2.4. Taxonomic Distribution of Bola-like Metabolites in Archaea, Some of Their Properties and Application, Archaeosomes
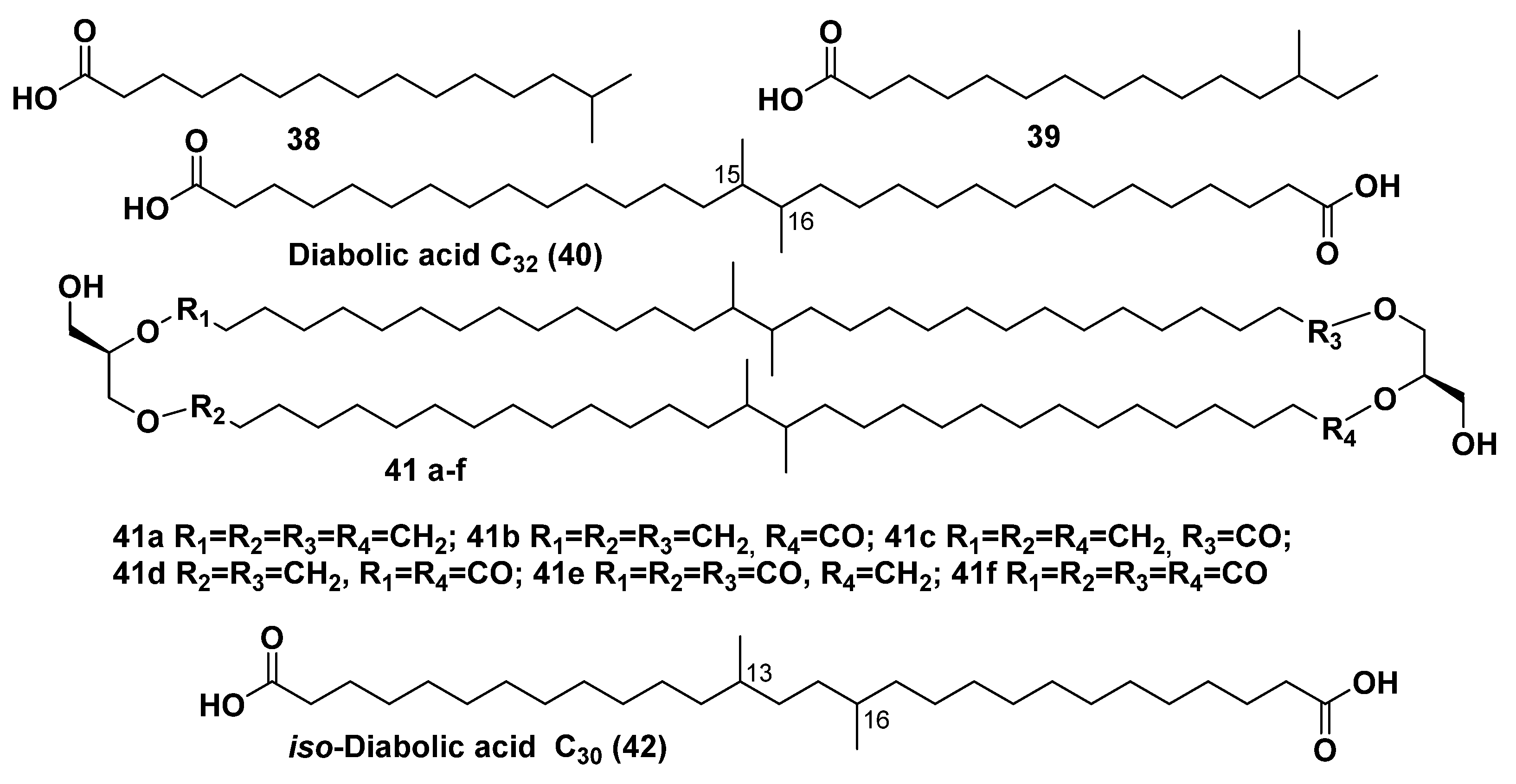
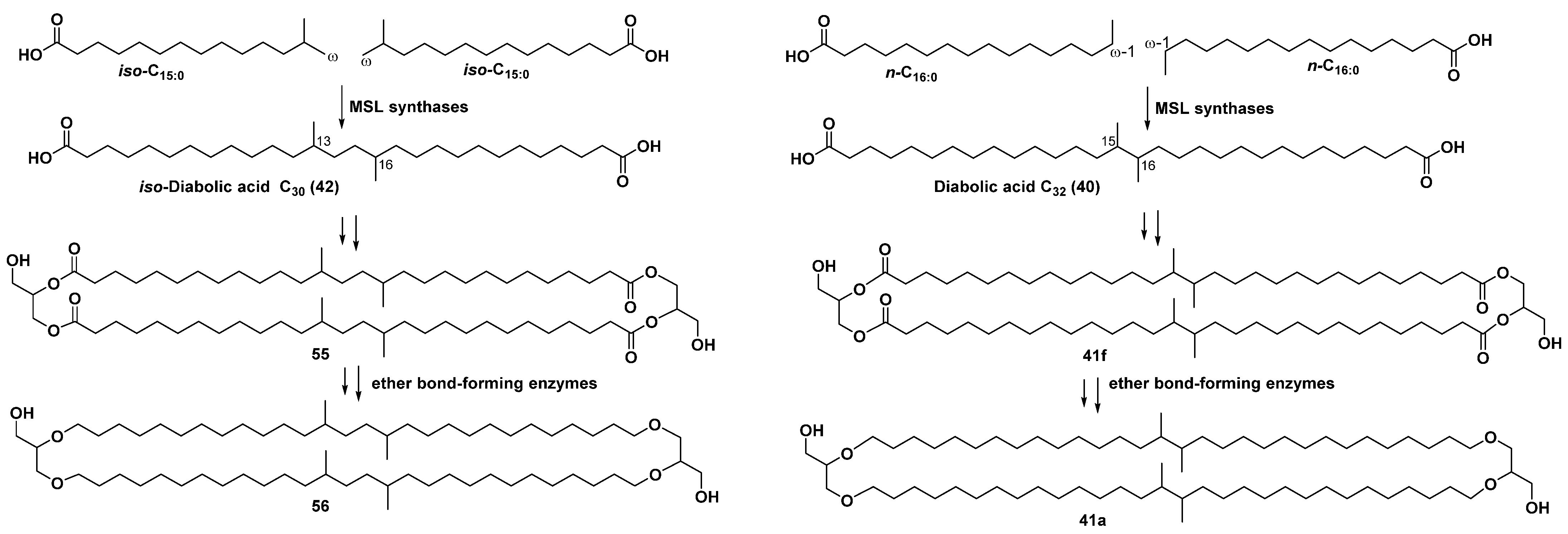
3. Natural Bola-Amphiphilic Compounds of Some Anaerobic Bacteria, Peat and Soil
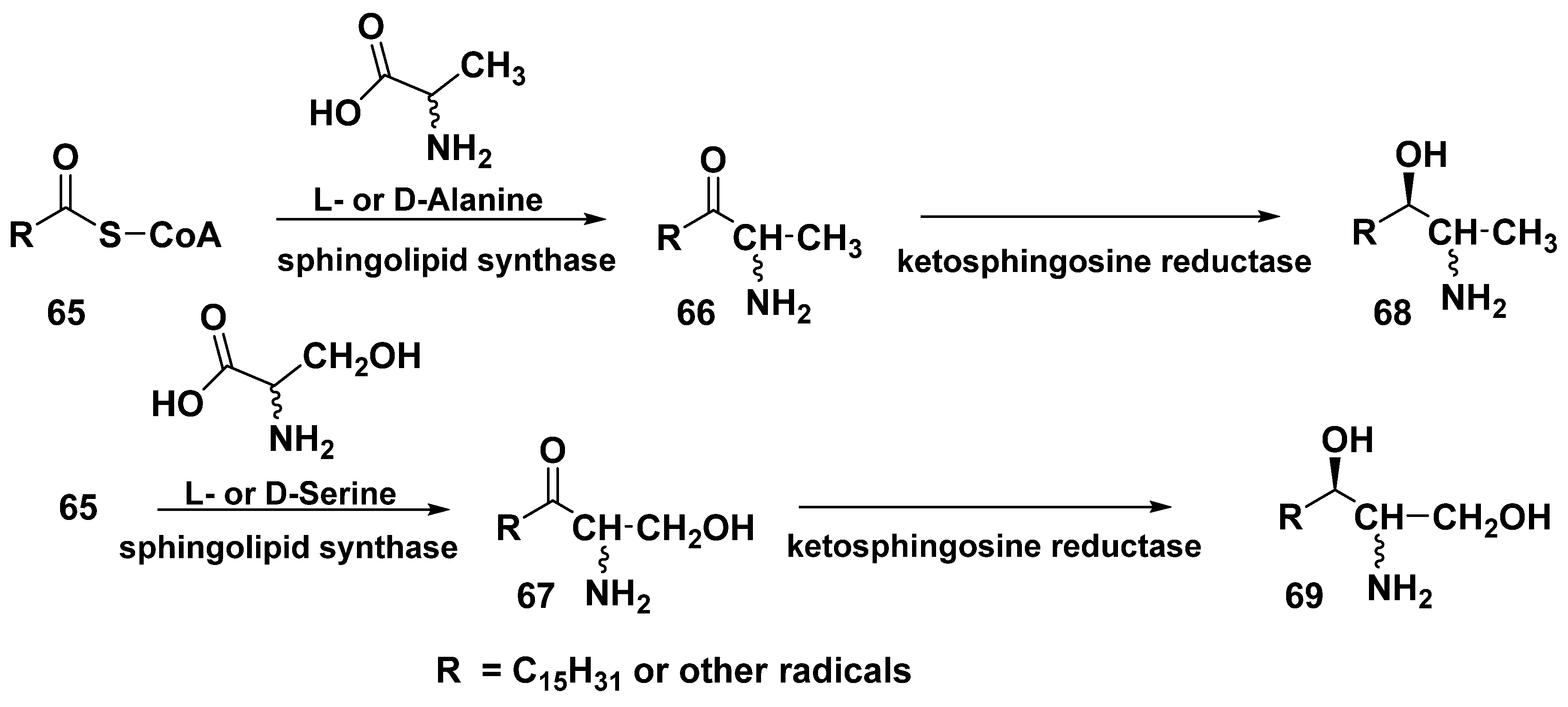
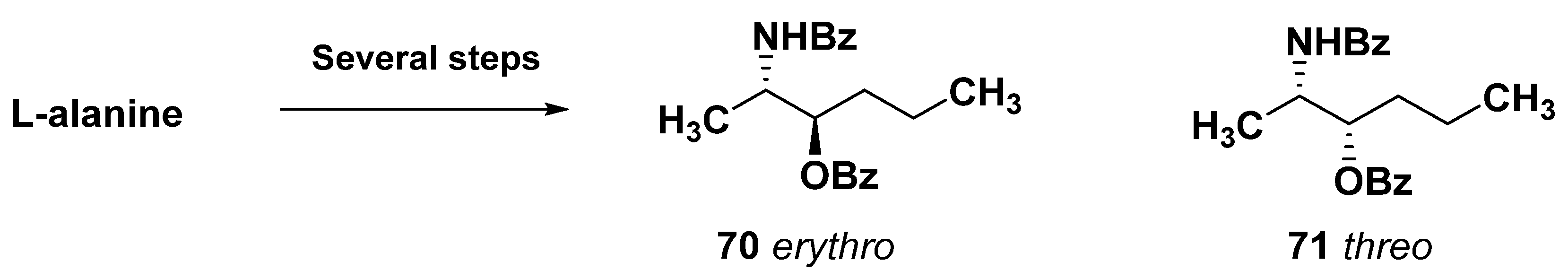
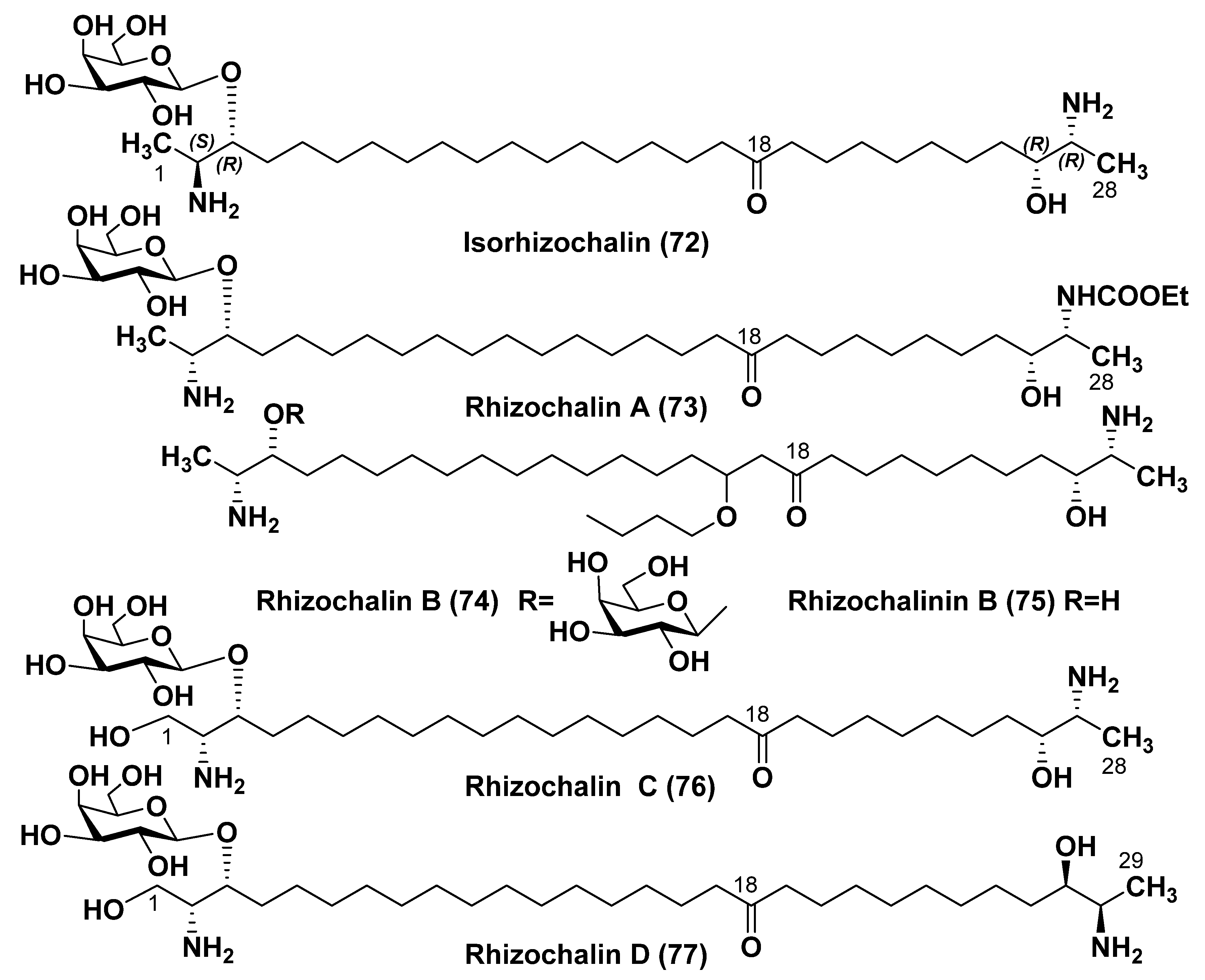
4. Sphingolipid-like Bolaamphiphiles from Marine Invertebrates
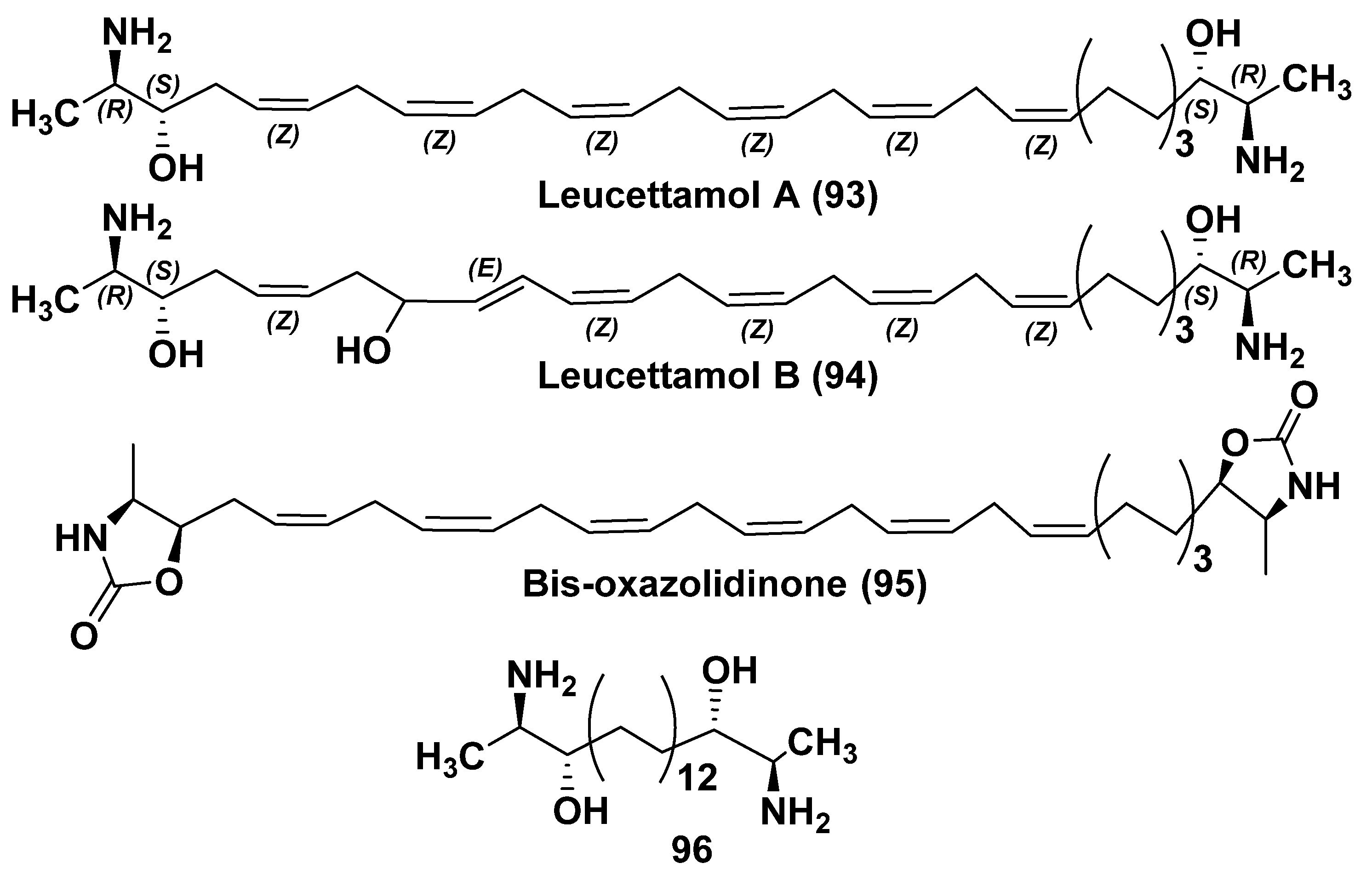
4.1. Sphingolipid-like Bolaamphiphiles from Sponges
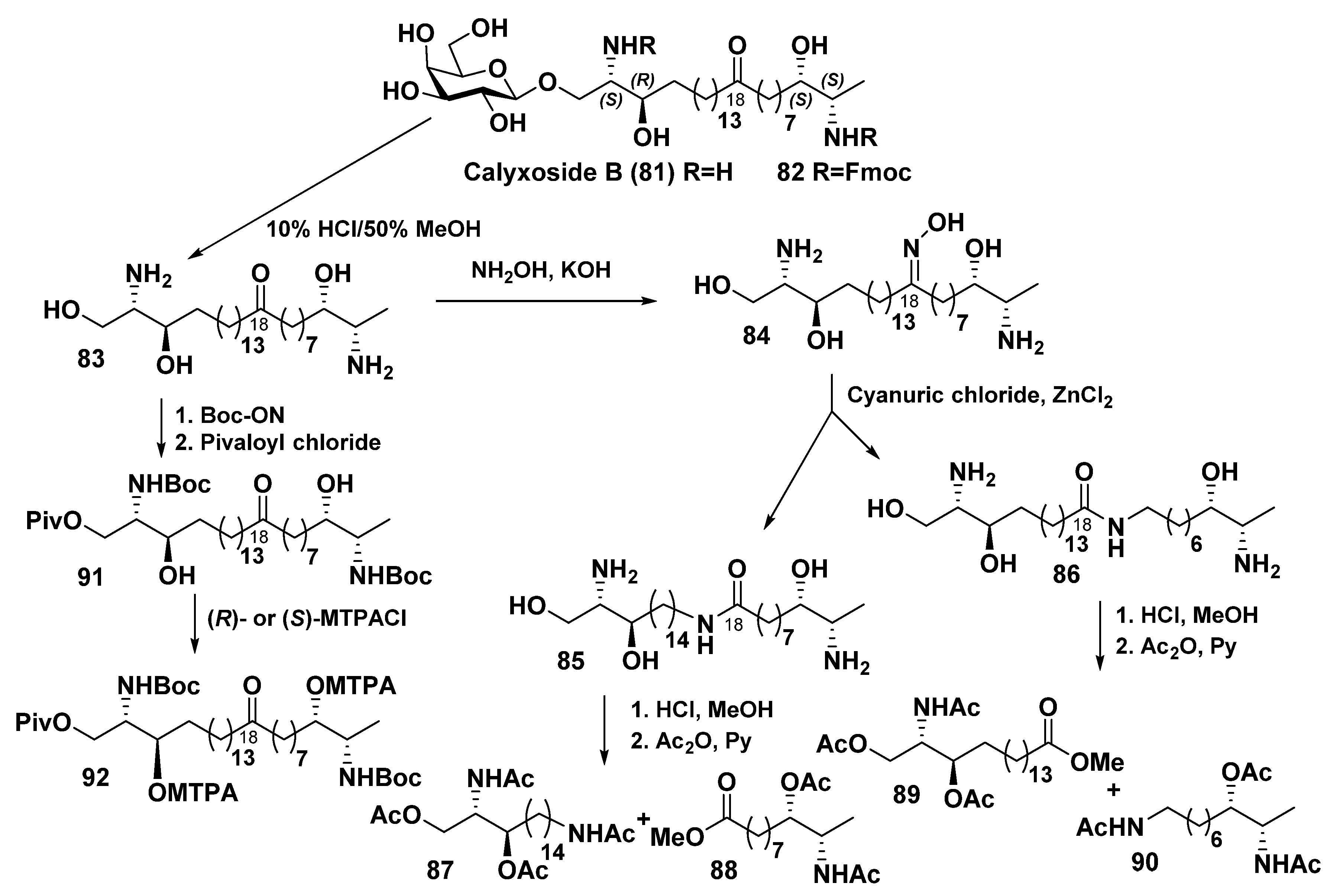
4.2. Bipolar Alkaloidolipids from Sponges
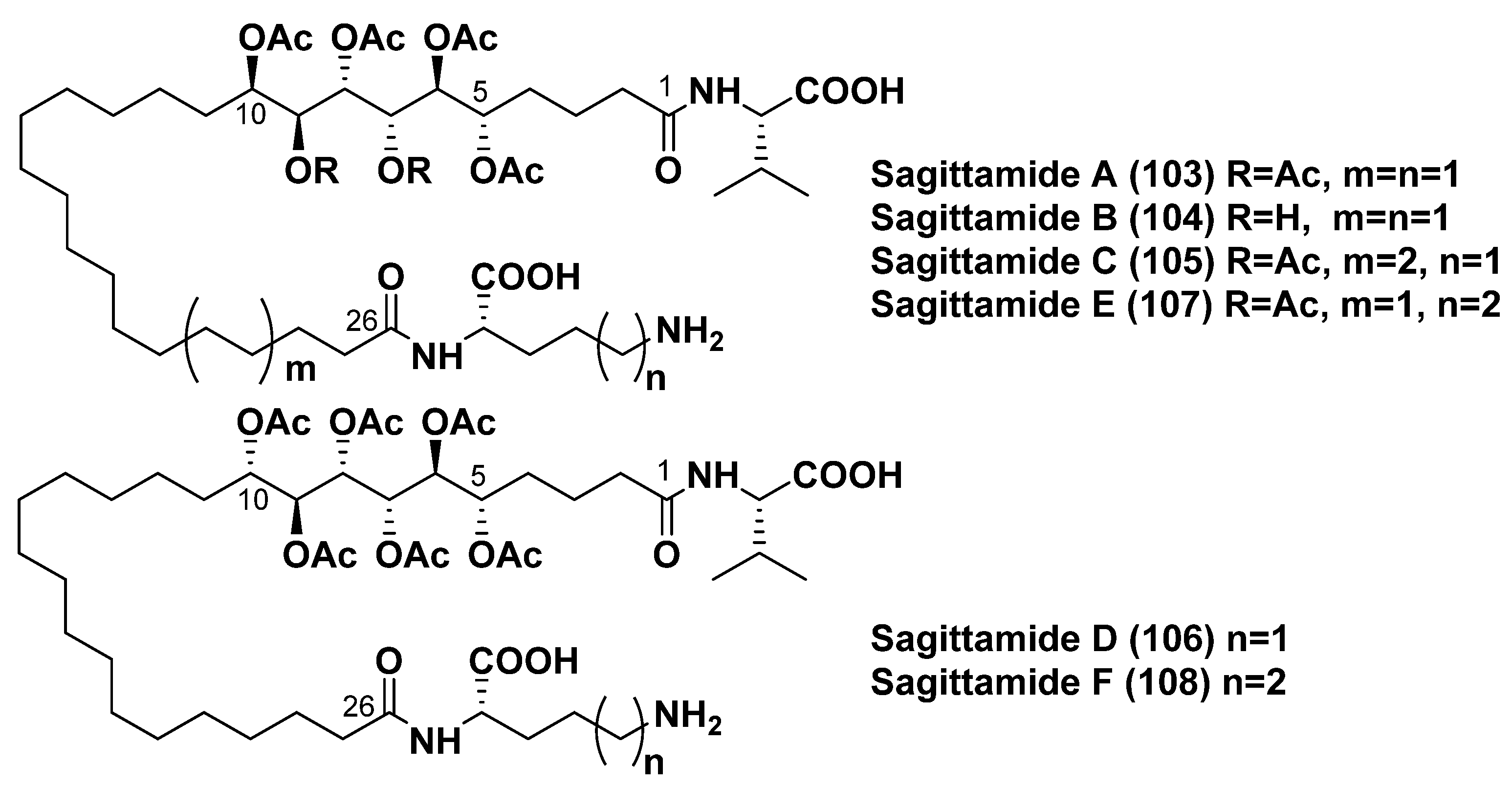
4.3. Sagittamides

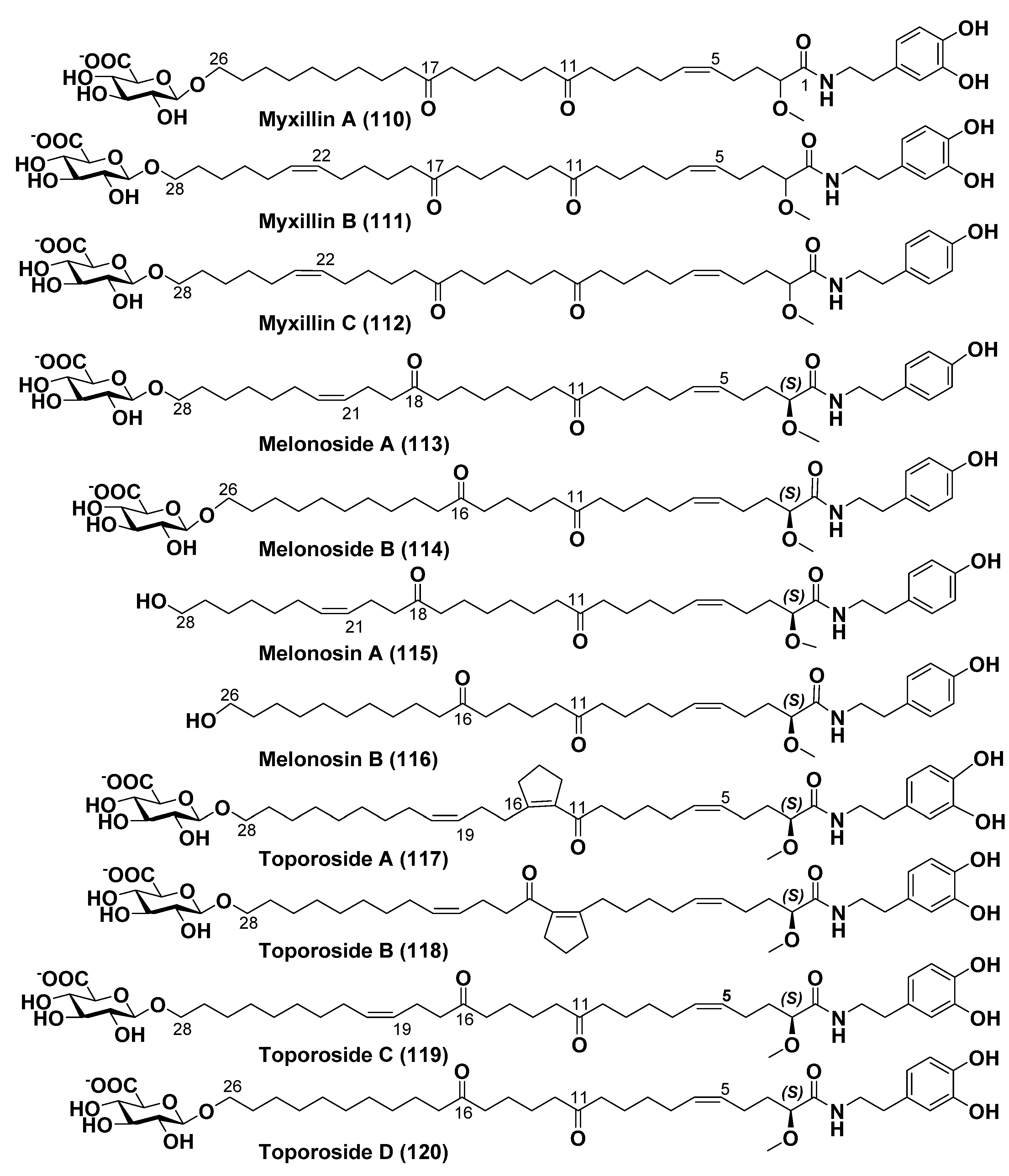
5. Other Bola-like Metabolites from Sponges
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fariya, M.; Jain, A.; Dhawan, V.; Shah, S.; Nagarsenker, M.S. Bolaamphiphiles: A pharmaceutical review. Adv. Pharm. Bull. 2014, 4, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Woese, C.R.; Fox, G.E. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proc. Natl. Acad. Sci. USA 1977, 74, 5088–5090. [Google Scholar] [CrossRef]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yafremava, L.S.; Caetano-Anolles, D.; Mittenthal, J.E.; Caetano-Anolles, G. Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world. Genome Res. 2007, 17, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-G.; Jung, M.-Y.; Park, S.-J.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S.; Madsen, E.L.; Min, D.; Kim, J.-S.; Kim, G.-J.; Rhee, S.-K. Cultivation of a highly enriched ammonia-oxidizing archaeon of thaumarchaeotal group I.1b from an agricultural soil. Environ. Microbiol. 2012, 14, 1528–1543. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Guo, W.; Liu, H. Basin scale variation on the composition and diversity of Archaea in the Pacific Ocean. Front. Microbiol. 2017, 8, 2057. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.E.; Richter, R.A.; Dupont, C.L. Planktonic marine archaea. Ann. Revs. Mar. Sci. 2019, 11, 131–158. [Google Scholar] [CrossRef]
- Lincoln, S.A.; Wai, B.; Eppley, J.M.; Church, M.J.; Summons, R.E.; DeLong, E.F. Planktonic Euryarchaeota are a significant source of archaeal tetraether lipids in the ocean. Proc. Natl. Acad. Sci. USA 2014, 111, 9858–9863. [Google Scholar] [CrossRef] [PubMed]
- Karner, M.B.; DeLong, E.F.; Karl, D.M. Archaeal dominance in the mesopelagic zone of the Pacific Ocean. Nature 2001, 409, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Lipp, J.S.; Morono, Y.; Inagaki, F.; Hinrichs, K.-U. Significant contribution of Archaea to extant biomass in marine subsurface sediments. Nature 2008, 454, 991–994. [Google Scholar] [CrossRef]
- Moissl-Eichinger, C.; Pausan, M.; Taffner, J.; Berg, G.; Bang, C.; Schmitz, R.A. Archaea are interactive components of complex microbiomes. Trends Microbiol. 2018, 26, 70–85. [Google Scholar] [CrossRef]
- Volmer, J.G.; McRae, H.; Morrison, M. The evolving role of methanogenic archaea in mammalian microbiomes. Front. Microbiol. 2023, 14, 1268451. [Google Scholar] [CrossRef] [PubMed]
- Wrede, C.; Dreier, A.; Kokoschka, S.; Hoppert, M. Archaea in symbioses. Archaea 2012, 1, 596846. [Google Scholar] [CrossRef] [PubMed]
- Moissl-Eichinger, C.; Huber, H. Archaeal symbionts and parasites. Curr. Opin. Microbiol. 2011, 14, 364–370. [Google Scholar] [CrossRef]
- Koga, Y. From promiscuity to the lipid divide: On the evolution of distinct membranes in Archaea and Bacteria. J. Mol. Evol. 2014, 78, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.; Meyerdierks, A.; Glockner, F.O.; Amann, R.; Widdel, F.; Kube, M.; Reinhardt, R.; Kahnt, J.; Böcher, R.; Thauer, R.K.; et al. A conspicuous nickel protein in microbial mats that oxidize methane anaerobically. Nature 2003, 426, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Takano, Y.; Kaneko, M.; Kahnt, J.; Imachi, H.; Shima, S.; Ohkouchi, N. Detection of coenzyme F430 in deep sea sediments: A key molecule for biological methanogenesis. Org. Geochem. 2013, 58, 137–140. [Google Scholar] [CrossRef]
- Kates, M.; Yengoyan, L.S.; Sastry, P.S. A diether analog of phosphatidyl glycerophosphate in Halobacterium cutirubrum. Biochim. Biophys. Acta 1965, 98, 252–269. [Google Scholar] [CrossRef] [PubMed]
- Langworthy, T.A. Long-chain diglycerol tetraethers from Thermoplasma acidophilum. Biochim. Biophys. Acta 1977, 487, 37–50. [Google Scholar] [CrossRef]
- Gräther, O.; Arigoni, D. Detection of regioisomeric macrocyclic tetraethers in the lipids of Methanobacterium thermoautotrophicum and other archaeal organisms. J. Chem. Soc. Chem. Comm. 1995, 4, 405–406. [Google Scholar] [CrossRef]
- DeRosa, M.; Gambacorta, A.; Nicolaus, B.; Sodano, S.; Bu’Lock, J.D. Structural regularities in tetraether lipids of Caldariella and their biosynthetic and phyletic implications. Phytochem 1980, 19, 833–836. [Google Scholar] [CrossRef]
- DeRosa, M.; Gambacorta, A.; Gliozzi, A. Structure, biosynthesis, and physicochemical properties of archaebacterial lipids. Microbiol. Rev. 1986, 50, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Hanford, M.J.; Peeples, T.L. Archaeal tetraether lipids: Unique structures and applications. Appl. Biochem. Biotechnol. 2002, 97, 45–62. [Google Scholar] [CrossRef]
- Chong, P.L.-G. Archaebacterial bipolar tetraether lipids: Physico-chemical and membrane properties. Chem. Phys. Lipids 2010, 163, 253–265. [Google Scholar] [CrossRef]
- Pearson, A. Resolving a piece of the archaeal lipid puzzle. Proc. Natl. Acad. Sci. USA 2019, 116, 22423–22425. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Kyselová, L.; Murphy, D.J. Archaeal lipids. Prog. Lipid Res. 2023, 91, 101237. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Chen, H.; Yang, H.; Chen, Y.; Yang, W.; Xi, F.; Pei, X.; Welander, P.V. Identification of a protein responsible for the synthesis of archaeal membrane-spanning GDGT lipids. Nat. Commun. 2022, 13, 1545. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, C.T.; Iwig, D.F.; Wang, B.; Cossu, M.; Metcalf, W.W.; Boal, A.K.; Booker, S.J. Discovery, structure and mechanism of a tetraether lipid synthase. Nature 2022, 609, 197–203. [Google Scholar] [CrossRef]
- Eguchi, T.; Ibaragi, K.; Kakinuma, K. Total synthesis of archaeal 72-membered macrocyclic tetraether lipids. J. Org. Chem. 1998, 63, 2689–2698. [Google Scholar] [CrossRef]
- Andringa, R.L.H.; de Kok, N.A.W.; Driessen, A.J.M.; Minnaard, A.J. A unified approach for the total synthesis of cyclo-archaeol, iso-caldarchaeol, caldarchaeol, and mycoketide. Angew. Chem. 2021, 133, 17638–17644. [Google Scholar] [CrossRef]
- Sita, L.R. Convenient highly stereoselective syntheses of (3R, 7R, 11R)-and (3S, 7R, 11R)-3, 7, 11, 15-tetramethylhexadecanoic acid (phytanic acid) and the corresponding 3, 7, 11, 15-tetramethylhexadecan-1-ols. J. Org. Chem. 1993, 58, 5285–5287. [Google Scholar] [CrossRef]
- Wang, A.; Wüstenberg, B.; Pfaltz, A. Enantio- and diastereoselective hydrogenation of farnesol and O-protected derivatives: Stereocontrol by changing the C=C bond configuration. Angew. Chem. 2008, 120, 2330–2332. [Google Scholar] [CrossRef]
- Sinninghe Damsté, J.S.; Schouten, S.; Hopmans, E.C.; van Duin, A.C.T.; Geenevasen, J.A.J. Crenarchaeol: The characteristic core glycerol dibiphytanyl glycerol tetraether membrane lipid of cosmopolitan pelagic crenarchaeota. J. Lipid Res. 2002, 43, 1641–1651. [Google Scholar] [CrossRef]
- Takano, Y.; Chikaraishi, Y.; Ogawa, N.O.; Nomaki, H.; Morono, Y.; Inagaki, F.; Kitazato, H.; Hinrichs, K.-U.; Ohkouchi, N. Sedimentary membrane lipids recycled by deep-sea benthic archaea. Nat. Geosci. 2010, 3, 858–861. [Google Scholar] [CrossRef]
- Knappy, C.S.; Chong, J.P.; Keely, B.J. Rapid discrimination of archaeal tetraether lipid cores by liquid chromatography-tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2009, 20, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Knappy, C.S.; Barillà, D.; De Blaquiere, J.P.A.; Morgan, H.W.; Nunn, C.E.M.; Suleman, M.; Tan, C.H.W.; Keely, B.J. Structural complexity in isoprenoid glycerol dialkyl glycerol tetraether lipid cores of Sulfolobus and other archaea revealed by liquid chromatography-tandem mass spectrometry. Chem. Phys. Lipids 2012, 165, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Blѐriot, Y.; Untersteller, E.; Fritz, B.; Sinaÿ, P. Total synthesis of calditol: Structural clarification of this typical component of Archaea order Sulfolobales. Chem. A Eur. J. 2002, 8, 240–246. [Google Scholar] [CrossRef]
- Siliakus, M.F.; van der Oost, J.; Kengen, S.W.M. Adaptations of archaeal and bacterial membranes to variations in temperature, pH and pressure. Extremophiles 2017, 21, 651–670. [Google Scholar] [CrossRef]
- Zeng, Z.; Liu, X.-L.; Wei, J.H.; Summons, R.E.; Welander, P.V. Calditol-linked membrane lipids are required for acid tolerance in Sulfolobus acidocaldarius. Proc. Natl. Acad. Sci. USA 2018, 115, 12932–12937. [Google Scholar] [CrossRef]
- Zeng, Z.; Liu, X.-L.; Farley, K.R.; Wei, J.H.; Metcalf, W.W.; Summons, R.E.; Welander, P.V. GDGT cyclization proteins identify the dominant archaeal sources of tetraether lipids in the ocean. Proc. Natl. Acad. Sci. USA 2019, 116, 22505–22511. [Google Scholar] [CrossRef] [PubMed]
- Holzheimer, M.; Sinninghe Damsté, J.S.; Schouten, S.; Havenith, R.W.A.; Cunha, A.V.; Minnaard, A.J. Total synthesis of the alleged structure of crenarchaeol enables structure revision. Angew. Chem. 2021, 133, 17645–17654. [Google Scholar] [CrossRef]
- Sinninghe Damsté, J.S.; Rijpstra, W.I.C.; Hopmans, E.C.; den Uijl, M.J.; Weijers, J.W.H.; Schouten, S. The enigmatic structure of the crenarchaeol isomer. Org. Geochem. 2018, 124, 22–28. [Google Scholar] [CrossRef]
- Summons, R.E.; Welander, P.V.; Gold, D.A. Lipid biomarkers: Molecular tools for illuminating the history of microbial life. Nat. Rev. Microbiol. 2022, 20, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Schouten, S.; Hopmans, E.S.; Sinninghe Damsté, J.S. The organic geochemistry of glycerol dialkyl glycerol tetraether lipids: A review. Org. Geochem. 2013, 54, 19–61. [Google Scholar] [CrossRef]
- Schouten, S.; Hopmans, E.C.; Schefuß, E.; Sinninghe Damsté, J.S. Distributional variations in marine crenarchaeotal membrane lipids: A new tool for reconstructing ancient sea water temperatures? Earth Planet. Sci. Lett. 2002, 204, 265–274. [Google Scholar] [CrossRef]
- Kim, J.-H.; van der Meer, J.; Schouten, S.; Helmke, P.; Willmott, V.; Sangiorgi, F.; Koc, N.; Hopmans, E.C.; Sinninghe Damsté, J.S. New indices and calibrations derived from the distribution of crenarchaeal isoprenoid tetraether lipids: Implications for past sea surface temperature reconstructions. Geochim. Cosmochim. Acta 2010, 74, 4639–4654. [Google Scholar] [CrossRef]
- Ho, S.L.; Mollenhauer, G.; Fietz, S.; Martínez-Garcia, A.; Lamy, F.; Rueda, G.; Schipper, K.; Meheust, M.; Rosell-Mele, A.; Stein, R.; et al. Appraisal of TEX86 and TEX86L thermometries in subpolar and polar regions. Geochim. Cosmochim. Acta 2014, 131, 213–226. [Google Scholar] [CrossRef]
- Fietz, S.; Ho, S.L.; Huguet, C. Archaeal membrane lipid-based paleothermometry for applications in polar oceans. Oceanography 2020, 33, 104–114. [Google Scholar] [CrossRef]
- Morii, H.; Eguchi, T.; Nishihara, M.; Kakinuma, K.; König, H.; Koga, Y. A novel ether core lipid with H-shaped C80-isoprenoid hydrocarbon chain from the hyperthermophilic methanogen Methanothermus fervidus. Biochem. Biophys. Acta 1998, 1390, 339–345. [Google Scholar] [CrossRef]
- Schouten, S.; Baas, M.; Hopmans, E.C.; Reysenbach, A.-L.; Sinninghe Damsté, J.S. Tetraether membrane lipids of Candidatus “Aciduliprofundum boonei”, a cultivated obligate thermoacidophilic euryarchaeote from deep-sea hydrothermal vents. Extremophiles 2008, 12, 119–124. [Google Scholar] [CrossRef]
- Zhu, C.; Meador, T.B.; Dummann, W.; Hinrichs, K.-U. Identification of unusual butanetriol dialkyl glycerol tetraether and pentanetriol dialkyl glycerol tetraether lipids in marine sediments. Rapid Commun. Mass Spectrom. 2014, 28, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.W.; Elling, F.J.; Yoshinaga, M.Y.; Söllinger, A.; Urich, T.; Hinrichs, K.-U. Unusual butane- and pentanetriol-based tetraether lipids in Methanomassiliicoccus luminyensis, a representative of the seventh order of methanogens. Appl. Environ. Microbiol. 2016, 82, 4505–4516. [Google Scholar] [CrossRef]
- Coffinet, S.; Meador, T.B.; Mühlena, L.; Becker, K.W.; Schröder, J.; Zhu, Q.-Z.; Lipp, J.S.; Heuer, V.B.; Crump, M.P.; Hinrichs, K.-U. Structural elucidation and environmental distributions of butanetriol and pentanetriol dialkyl glycerol tetraethers (BDGTs and PDGTs). Biogeosciences 2020, 17, 317–330. [Google Scholar] [CrossRef]
- Coffinet, S.; Mühlena, L.; Lipp, J.S.; Weil, M.; Neubauer, C.; Urich, T.; Hinrichs, K.-U. Evidence for enzymatic backbone methylation of the main membrane lipids in the archaeon Methanomassiliicoccus luminyensis. Appl. Environ. Microbiol. 2022, 88, e0215421. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-L.; Lipp, J.S.; Simpson, J.H.; Lin, Y.-S.; Summons, R.E.; Hinrichs, K.-U. Mono-and dihydroxyl glycerol dibiphytanyl glycerol tetraethers in marine sediments: Identification of both core and intact polar lipid forms. Geochim. Cosmochim. Acta 2012, 89, 102–115. [Google Scholar] [CrossRef]
- Elling, F.J.; Könneke, M.; Nicol, G.W.; Stieglmeier, M.; Bayer, B.; Spieck, E.; de la Torre, J.R.; Becker, K.W.; Thomm, M.J.; Prosser, J.I.; et al. Chemotaxonomic characterisation of the thaumarchaeal lipidome. Environ. Microbiol. 2017, 19, 2681–2700. [Google Scholar] [CrossRef]
- Xiao, W.; Xu, Y.; Zhang, C.; Lin, J.; Wu, W.; Xiaoxia, L.; Tan, J.; Zhang, X.; Zheng, F.; Song, X.; et al. Disentangling effects of sea surface temperature and water depth on hydroxylated isoprenoid GDGTs: Insights from the Hadal zone and global sediments. Geophys. Res. Lett. 2023, 50, e2023GL103109. [Google Scholar] [CrossRef]
- Koga, Y.; Nishihara, M.; Morii, H.; Akagawa-Matsushita, M. Ether polar lipids of methanogenic bacteria: Structures, comparative aspects, and biosyntheses. Microbiol. Rev. 1993, 57, 164–182. [Google Scholar] [CrossRef]
- Scholte, A.; Hübner, C.; Ströhl, D.; Scheufler, O.; Czich, S.; Börke, J.M.; Hildebrand, G.; Liefeith, K. First isolation and structure elucidation of GDNT-β-Glu—Tetraether lipid fragment from archaeal Sulfolobus strains. Chem. Open 2021, 10, 889–895. [Google Scholar] [CrossRef]
- Řezanka, T.; Kolouchova, I.; Gharwalova, L.; Palyzova, A.; Sigler, K. Lipidomic analysis: From archaea to mammals. Lipids 2018, 53, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Law, K.P.; Zhang, C.L. Current progress and future trends in mass spectrometry-based archaeal lipidomics. Org. Geochem. 2019, 134, 45–61. [Google Scholar] [CrossRef]
- Lobasso, S.; Lopalco, P.; Angelini, R.; Vitale, R.; Huber, H.; Müller, V.; Corcelli, A. Coupled TLC and MALDI-TOF/MS analyses of the lipid extract of the hyperthermophilic archaeon Pyrococcus furiosus. Archaea 2012, 2012, 957852. [Google Scholar] [CrossRef] [PubMed]
- Tourte, M.; Kuentz, V.; Schaeffer, P.; Grossi, V.; Cario, A.; Oger, P.M. Novel intact polar and core lipid compositions in the Pyrococcus model species, P. furiosus and P. yayanosii, reveal the largest lipid diversity amongst Thermococcales. Biomolecules 2020, 10, 830. [Google Scholar] [CrossRef] [PubMed]
- Tourte, M.; Schaeffer, P.; Grossi, V.; Oger, P.M. Functionalized membrane domains: An ancestral feature of Archaea? Front. Microbiol. 2020, 11, 526. [Google Scholar] [CrossRef]
- Cario, A.; Grossi, V.; Schaeffer, P.; Oger, P.M. Membrane homeoviscous adaptation in the piezo-hyperthermophilic archaeon Thermococcus barophilus. Front. Microbiol. 2015, 6, 1152. [Google Scholar] [CrossRef]
- Bauersachs, T.; Weidenbach, K.; Schmitz, R.A.; Schwark, L. Distribution of glycerol ether lipids in halophilic, methanogenic and hyperthermophilic archaea. Org. Geochem. 2015, 83, 101–108. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, W.; He, W.; Xiao, W.; Chen, Y.; Zhu, Y.; Zheng, F.; Zhang, C. Lipidomic chemotaxonomy aligned with phylogeny of Halobacteria. Front. Microbiol. 2023, 14, 1297600. [Google Scholar] [CrossRef]
- Naafs, B.D.A.; McCormick, D.; Inglis, G.N.; Pancost, R.D. Archaeal and bacterial H-GDGTs are abundant in peat and their relative abundance is positively correlated with temperature. Geochim. Cosmochim. Acta 2018, 227, 156–170. [Google Scholar] [CrossRef]
- Chugunov, A.O.; Volynsky, P.E.; Krylov, N.A.; Boldyrev, I.A.; Efremov, R.G. Liquid but durable: Molecular dynamics simulations explain the unique properties of archaeal-like membranes. Sci. Rep. 2014, 4, 7462. [Google Scholar] [CrossRef]
- Rastädter, K.; Wurm, D.J.; Spadiut, O.; Quehenberger, J. The cell membrane of Sulfolobus spp. Homeoviscous adaption and biotechnological applications. Int. J. Mol. Sci. 2020, 21, 3935. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, B.; Gupta, R.; Gulati, M.; Singh, S.K.; Khursheed, R.; Gupta, M. The Why, Where, Who, How, and What of the vesicular delivery systems. Adv. Colloid Interface Sci. 2019, 271, 101985. [Google Scholar] [CrossRef]
- Jacquemet, A.; Barbeau, J.; Lemiègre, L.; Benvegnu, T. Archaeal tetraether bipolar lipids: Structures, functions and applications. Biochimie 2009, 91, 711–717. [Google Scholar] [CrossRef]
- Kashyap, K. Archaeosomes: Revolutionary technique for both cell-based and drug-based delivery applications. Int. J. Pharm. Sci. Med. 2021, 6, 102–127. [Google Scholar] [CrossRef]
- Kaur, G.; Garg, T.; Rath, G.; Goyal, A.K. Archaeosomes: An excellent carrier for drug and cell delivery. Drug Deliv. 2016, 23, 2497–2512. [Google Scholar] [CrossRef] [PubMed]
- Santhosh, P.B.; Genova, J. Archaeosomes: New generation of liposomes based on archaeal lipids for drug delivery and biomedical applications. ACS Omega 2022, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chong, P.L.-G.; Chang, A.; Yu, A.; Mammedova, A. Vesicular and planar membranes of archaea lipids: Unusual physical properties and biomedical applications. Int. J. Mol. Sci. 2022, 23, 7616. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, N.; Krawczyk, K.T.; Locht, C.; Kowalewicz-Kulbat, M. Archaeosomes and gas vesicles as tools for vaccine development. Front. Immunol. 2021, 12, 746235. [Google Scholar] [CrossRef]
- Ayesa, U.; Chong, P.L.-G. Polar lipid fraction E from Sulfolobus acidocaldarius and dipalmitoylphosphatidylcholine can form stable yet thermo-sensitive tetraether/diester hybrid archaeosomes with controlled release capability. Int. J. Mol. Sci. 2020, 21, 8388. [Google Scholar] [CrossRef]
- Satyanarayana, T.; Raghukumar, C.; Shivaji, S. Extremophilic microbes: Diversity and perspectives. Curr. Sci. 2005, 89, 78–90. Available online: http://www.jstor.org/stable/24110434 (accessed on 20 December 2024).
- Koga, Y. Thermal adaptation of the archaeal and bacterial lipid membranes. Archaea 2012, 2012, 1222012. [Google Scholar] [CrossRef]
- Miura, Y. The biological significance of ω-oxidation of fatty acids. Proc. Jpn. Acad. Ser. B 2013, 89, 370–382. [Google Scholar] [CrossRef]
- Kaneda, T. Iso-and anteiso-fatty acids in bacteria: Biosynthesis, function, and taxonomic significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Clarke, N.G.; Hazlewood, G.P.; Dawson, R.M.C. Structure of diabolic acid-containing phospholipids isolated from Butyrivibrio sp. Biochem. J. 1980, 191, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Sinninghe Damsté, J.S.; Rijpstra, W.I.C.; Hopmans, E.C.; Schouten, S.; Balk, M.; Stams, A.J. Structural characterization of diabolic acid-based tetraester, tetraether and mixed ether/ester, membrane-spanning lipids of bacteria from the order Thermotogales. Arch. Microbiol. 2007, 188, 629–641. [Google Scholar] [CrossRef]
- Sahonero-Canavesi, D.X.; Villanueva, L.; Bale, N.J.; Bosviel, J.; Koenen, M.; Hopmans, E.C.; Sinninghe Damsté, J.S. Changes in the distribution of membrane lipids during growth of Thermotoga maritima at different temperatures: Indications for the potential mechanism of biosynthesis of ether-bound diabolic acid (membrane-spanning) lipids. Appl. Environ. Microbiol. 2022, 88, e01763-21. [Google Scholar] [CrossRef]
- Sinninghe Damsté, J.S.; Rijpstra, W.I.C.; Hopmans, E.C.; Weijers, J.W.; Foesel, B.U.; Overmann, J.; Dedysh, S.N. 13,16-Dimethyl octacosanedioic acid (iso-diabolic acid), a common membrane-spanning lipid of Acidobacteria subdivisions 1 and 3. Appl. Environ. Microbiol. 2011, 77, 4147–4154. [Google Scholar] [CrossRef] [PubMed]
- Weijers, J.W.; Schouten, S.; Hopmans, E.C.; Geenevasen, J.A.; David, O.R.; Coleman, J.M.; Pancost, R.D.; Sinninghe Damsté, J.S. Membrane lipids of mesophilic anaerobic bacteria thriving in peats have typical archaeal traits. Environ. Microbiol. 2006, 8, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Weijers, J.W.; Panoto, E.; van Bleijswijk, J.; Schouten, S.; Rijpstra, W.I.C.; Balk, M.; Stams, J.W.H.; Rijpstra, W.I.; Sinninghe Damsté, J.S. Constraints on the biological source(s) of the orphan branched tetraether membrane lipids. Geomicrobiol. J. 2009, 26, 402–414. [Google Scholar] [CrossRef]
- De Jonge, C.; Hopmans, E.C.; Stadnitskaia, A.; Rijpstra, W.I.C.; Hofland, R.; Tegelaar, E.; Sinninghe Damsté, J.S. Identification of novel penta-and hexamethylated branched glycerol dialkyl glycerol tetraethers in peat using HPLC–MS2, GC–MS and GC–SMB-MS. Org. Geochem. 2013, 54, 78–82. [Google Scholar] [CrossRef]
- Peterse, F.; van der Meer, J.; Schouten, S.; Weijers, J.W.; Fierer, N.; Jackson, R.B.; Kim, J.H.; Sinninghe Damsté, J.S. Revised calibration of the MBT–CBT paleotemperature proxy based on branched tetraether membrane lipids in surface soils. Geochim. Cosmochim. Acta 2012, 96, 215–229. [Google Scholar] [CrossRef]
- Zhao, B.; Russell, J.M.; Tsai, V.C.; Blaus, A.; Parish, M.C.; Liang, J.; Wilk, A.; Du, X.; Bush, M.B. Evaluating global temperature calibrations for lacustrine branched GDGTs: Seasonal variability, paleoclimate implications, and future directions. Quat. Sci. Rev. 2023, 310, 108124. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, F.; Yang, H.; Yang, W.; Wu, R.; Liu, X.; Liang, H.; Chen, H.; Pei, H.; Zhang, C.; et al. The production of diverse brGDGTs by an Acidobacterium providing a physiological basis for paleoclimate proxies. Geochim. Cosmochim. Acta 2022, 337, 155–165. [Google Scholar] [CrossRef]
- Sahonero-Canavesi, D.X.; Siliakus, M.F.; Abdala Asbun, A.; Koenen, M.; von Meijenfeldt, F.B.; Boeren, S.; Bale, N.J.; Engelman, J.C.; Fiege, K.; von Schijndel, L.S.; et al. Disentangling the lipid divide: Identification of key enzymes for the biosynthesis of membrane-spanning and ether lipids in Bacteria. Sci. Adv. 2022, 8, eabq8652. [Google Scholar] [CrossRef]
- Peterse, F.; Hopmans, E.C.; Schouten, S.; Mets, A.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S. Identification and distribution of intact polar branched tetraether lipids in peat and soil. Org. Geochem. 2011, 42, 1007–1015. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Denisenko, V.A.; Stonik, V.A.; Milgrom, Y.N.; Rashkes, Y.V. Rhizochalin, a novel secondary metabolite of mixed biosynthesis from the sponge Rhizochalina incrustata. Tetrahedron Lett. 1989, 30, 6581–6584. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Dmitrenok, P.S.; Zakharenko, A.M.; Denisenko, V.A.; Guzii, A.G.; Li, R.; Skepper, C.R.; Molinski, T.F.; Stonik, V.A. Rhizochalins C and D from the sponge Rhizochalina incrustata. A rare threo-sphingolipid and a facile method for determination of the carbonyl position in α,ω-bifunctionalized ketosphingolipids. J. Nat. Prod. 2007, 70, 1991–1998. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.J.; Dunn, T.M.; Campopiano, D.J. Sphingolipid biosynthesis in man and microbes. Nat. Prod. Rep. 2018, 35, 921–954. [Google Scholar] [CrossRef]
- Molinski, T.F.; Makarieva, T.N.; Stonik, V.A. (−)-Rhizochalin is a dimeric enantiomorphic (2R)-sphingolipid: Absolute configuration of pseudo-C2v-symmetric bis-2-amino-3-alkanols by CD. Angew. Chem. 2000, 112, 4242–4245. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Zakharenko, A.M.; Dmitrenok, P.S.; Guzii, A.G.; Denisenko, V.A.; Savina, A.S.; Dalisay, D.S.; Molinski, T.F.; Stonik, V.A. Isorhizochalin: A minor unprecedented bipolar sphingolipid of stereodivergent biogenesis from the Rhizochalina incrustata. Lipids 2009, 44, 1155–1162. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Guzii, A.G.; Denisenko, V.A.; Dmitrenok, P.S.; Santalova, E.A.; Pokanevich, E.V.; Molinski, T.F.; Stonik, V.A. Rhizochalin A, a novel two-headed sphingolipid from the sponge Rhizochalina incrustata. J. Nat. Prod. 2005, 68, 255–257. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Guzii, A.G.; Denisenko, V.A.; Dmitrenok, P.S.; Stonik, V.A. New two-headed sphingolipid-like compounds from the marine sponge Oceanapia sp. Russ. Chem. Bull. 2008, 57, 669–673. [Google Scholar] [CrossRef]
- Jin, J.O.; Shastina, V.; Park, J.I.; Han, J.Y.; Makarieva, T.; Fedorov, S.; Rasskazov, V.; Stonik, V.; Kwak, J.Y. Differential induction of apoptosis of leukemic cells by rhizochalin, two headed sphingolipids from sponge and its derivatives. Biol. Pharm. Bull. 2009, 32, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A.; Otte, K.; Tabakmakher, K.M.; Hauschild, J.; Makarieva, T.N.; Shubina, L.K.; Fedorov, S.N.; Bokemeyer, C.; Stonik, V.A.; von Amsberg, G. Synthesis and anticancer activity of the derivatives of marine compound rhizochalin in castration resistant prostate cancer. Oncotarget 2018, 9, 16962–16973. [Google Scholar] [CrossRef]
- Khanal, P.; Kang, B.S.; Yun, H.J.; Cho, H.G.; Makarieva, T.N.; Choi, H.S. Aglycon of rhizochalin from the Rhizochalina incrustata induces apoptosis via activation of AMP-activated protein kinase in HT-29 colon cancer cells. Biol. Pharm. Bull. 2011, 34, 1553–1558. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, S.N.; Makarieva, T.N.; Guzii, A.G.; Shubina, L.K.; Kwak, J.Y.; Stonik, V.A. Marine two-headed sphingolipid-like compound rhizochalin inhibits EGF-induced transformation of JB6 P+ Cl 41 cells. Lipids 2009, 44, 777–785. [Google Scholar] [CrossRef]
- Sibirtsev, J.T.; Shastina, V.V.; Menzorova, N.I.; Makarieva, T.N.; Rasskazov, V.A. Ca2+, Mg2+-dependent DNAase involvement in apoptotic effects in spermatozoa of sea urchin Strongylocentrotus intermedius induced by two-headed sphingolipid rhizochalin. Mar. Biotechnol. 2011, 13, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Makarieva, T.N.; Ivanchina, N.V.; Stonik, V.A. Application of oxidative and reductive transformations in the structure determination of marine natural products. J. Nat. Prod. 2020, 83, 1314–1333. [Google Scholar] [CrossRef]
- Nicholas, G.M.; Hong, T.W.; Molinski, T.F.; Lerch, M.L.; Cancilla, M.T.; Lebrilla, C.B. Oceanapiside, an antifungal bis-α,ω-amino alcohol glycoside from the marine sponge Oceanapia phillipensis. J. Nat. Prod. 1999, 62, 1678–1681. [Google Scholar] [CrossRef]
- Nicholas, G.M.; Molinski, T.F. Enantiodivergent biosynthesis of the dimeric sphingolipid oceanapiside from the marine sponge Oceanapia phillipensis. Determination of remote stereochemistry. J. Am. Chem. Soc. 2000, 122, 4011–4019. [Google Scholar] [CrossRef]
- Dalisay, D.S.; Rogers, E.W.; Molinski, T.F. Oceanapiside, a marine natural product, targets the sphingolipid pathway of fluconazole-resistant Candida glabrata. Mar. Drugs 2021, 19, 126. [Google Scholar] [CrossRef]
- Zhou, B.N.; Mattern, M.P.; Johnson, R.K.; Kingston, D.G. Structure and stereochemistry of a novel bioactive sphingolipid from a Calyx sp. Tetrahedron 2001, 57, 9549–9554. [Google Scholar] [CrossRef]
- Sugawara, K.; Watarai, H.; Ise, Y.; Yokose, H.; Morii, Y.; Yamawaki, N.; Matsunaga, S. Structure elucidation of calyxoside B, a bipolar sphingolipid from a marine sponge Cladocroce sp. through the use of Beckmann rearrangement. Mar. Drugs 2021, 19, 287. [Google Scholar] [CrossRef]
- Kong, F.; Faulkner, D.J. Leucettamols A and B, two antimicrobial lipids from the calcareous sponge Leucetta microraphis. J. Org. Chem. 1993, 58, 970–971. [Google Scholar] [CrossRef]
- Dalisay, D.S.; Tsukamoto, S.; Molinski, T.F. Absolute configuration of the α,ω-bifunctionalized sphingolipid leucettamol A from Leucetta microrhaphis by deconvoluted exciton coupled CD. J. Nat. Prod. 2009, 72, 353–359. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Takeuchi, T.; Rotinsulu, H.; Mangindaan, R.E.; van Soest, R.W.; Ukai, K.; Kobayashi, H.; Namikoshi, M.; Ohtsa, T.; Yokosawa, H. Leucettamol A: A new inhibitor of Ubc13-Uev1A interaction isolated from a marine sponge, Leucetta aff. microrhaphis. Bioorg. Med. Chem. Lett. 2008, 18, 6319–6320. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G.; Voets, T.; Peters, J.A. Transient receptor potential cation channels in disease. Physiol. Rev. 2007, 87, 165–217. [Google Scholar] [CrossRef]
- Chianese, G.; Fattorusso, E.; Putra, M.Y.; Calcinai, B.; Bavestrello, G.; Moriello, A.S.; De Petrocellis, D.; Di Marzo, V.; Taglialatela-Scafati, O. Leucettamols, bifunctionalized marine sphingoids, act as modulators of TRPA1 and TRPM8 channels. Mar. Drugs 2012, 10, 2435–2447. [Google Scholar] [CrossRef] [PubMed]
- Makarieva, T.N.; Denisenko, V.A.; Dmitrenok, P.S.; Guzii, A.G.; Santalova, E.A.; Stonik, V.A.; MacMillan, J.B.; Molinski, T.F. Oceanalin A, a hybrid α,ω-bifunctionalized sphingoid tetrahydroisoquinoline β-glycoside from the marine sponge Oceanapia sp. Org. Lett. 2005, 7, 2897–2900. [Google Scholar] [CrossRef] [PubMed]
- Makarieva, T.N.; Ivanchina, N.V.; Dmitrenok, P.S.; Guzii, A.G.; Stonik, V.A.; Dalisay, D.S.; Molinski, T.F. Oceanalin B, a hybrid α,ω-bifunctionalized sphingoid tetrahydroisoquinoline β-glycoside from the marine sponge Oceanapia sp. Mar. Drugs 2021, 19, 635. [Google Scholar] [CrossRef] [PubMed]
- Lievens, S.C.; Molinski, T.F. Sagittamides A and B. Polyacetoxy long-chain acyl amino acids from a Didemnid Ascidian. Org. Lett. 2005, 7, 2281–2284. [Google Scholar] [CrossRef]
- Lievens, S.C.; Molinski, T.F. Progressive−convergent elucidation of stereochemistry in complex polyols. The absolute configuration of (−)-sagittamide A. J. Am. Chem. Soc. 2006, 128, 11764–11765. [Google Scholar] [CrossRef]
- Seike, H.; Ghosht, I.; Kishi, Y. Stereochemistry of sagittamide A: Prediction and confirmation. Org. Lett. 2006, 8, 3865–3868. [Google Scholar] [CrossRef] [PubMed]
- Schuetz, A.; Junker, J.; Leonov, A.; Lange, O.F.; Molinski, T.F.; Griesinger, C. Stereochemistry of sagittamide A from residual dipolar coupling enhanced NMR. J. Am. Chem. Soc. 2007, 129, 15114–15115. [Google Scholar] [CrossRef] [PubMed]
- Lievens, S.C.; Morinaka, B.I.; Molinski, T.F. Stereochemical elucidation of new sagittamides C–F from a Didemnid ascidian. Aust. J. Chem. 2010, 63, 935–941. [Google Scholar] [CrossRef]
- Humbert, A.; Plé, K.; Harakat, D.; Martinez, A.; Haudrechy, A. A further contribution to the study of sagittamide A: Synthesis of a pivotal intermediate belonging to a rare L-series. Molecules 2012, 17, 7709–7721. [Google Scholar] [CrossRef]
- Whitson, E.L.; Bugni, T.S.; Chockalingam, P.S.; Concepcion, G.P.; Feng, X.; Jin, G.; Herper, M.K.; Mangalindon, G.C.; McDonald, L.A.; Ireland, C.M. Fibrosterol sulfates from the Philippine sponge Lissodendoryx (Acanthodoryx) fibrosa: Sterol dimers that inhibit PKCζ. J. Org. Chem. 2009, 74, 5902–5908. [Google Scholar] [CrossRef] [PubMed]
- Ushiyama, S.; Umaoka, H.; Kato, H.; Suwa, Y.; Morioka, H.; Rotinsulu, H.; Losung, F.; Mangindaan, R.E.P.; De Foogd, N.J.; Yokosawa, H.; et al. Manadosterols A and B, sulfonated sterol dimers inhibiting the Ubc13–Uev1A interaction, isolated from the marine sponge Lissodendryx fibrosa. J. Nat. Prod. 2012, 75, 1495–1499. [Google Scholar] [CrossRef]
- Einarsdottir, E.; Liu, H.-B.; Freysdottir, J.; Gotfredsen, C.H.; Omarsdottir, S. Immunomodulatory N-acyl dopamine glycosides from the icelandic marine sponge Myxilla incrustans collected at a hydrothermal vent site. Planta Med. 2016, 82, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Guzii, A.G.; Makarieva, T.N.; Denisenko, V.A.; Dmitrenok, P.S.; Popov, R.S.; Kuzmich, A.S.; Dyshlovoy, S.A.; von Amsberg, G.; Krasokhin, V.B.; Stonik, V.A. Melonoside A: An ω-glycosylated fatty acid amide from the Far Eastern marine sponge Melonanchora kobjakovae. Org. Lett. 2016, 18, 3478–3481. [Google Scholar] [CrossRef]
- Guzii, A.G.; Makarieva, T.N.; Denisenko, V.A.; Dmitrenok, P.S.; Popov, R.S.; Kuzmich, A.S.; Krasokhin, V.B.; Kim, N.Y.; Stonik, V.A. Melonoside B and melonosins A and B, lipids containing multifunctionalized ω-hydroxy fatty acid amides from the far eastern marine sponge Melonanchora kobjakovae. J. Nat. Prod. 2018, 81, 2763–2767. [Google Scholar] [CrossRef] [PubMed]
- Guzii, A.G.; Makarieva, T.N.; Fedorov, S.N.; Menshov, A.S.; Denisenko, V.A.; Popov, R.S.; Iarotsckaia, V.V.; Kim, N.Y.; Stonik, V.A. Toporosides A and B, cyclopentenyl-containing ω-glycosylated fatty acid amides, and toporosides C and D from the Northwestern Pacific marine sponge Stelodoryx toporoki. J. Nat. Prod. 2022, 86, 1186–1191. [Google Scholar] [CrossRef] [PubMed]
- Fuhrhop, J.H.; Wang, T. Bolaamphiphiles. Chem. Rev. 2004, 104, 2901–2938. [Google Scholar] [CrossRef] [PubMed]
- Nuraje, N.; Bai, H.; Su, K. Bolaamphiphilic molecules: Assembly and applications. Prog. Polym. Sci. 2013, 8, 302–343. [Google Scholar] [CrossRef]
- Dhasaiyan, P.; Prasad, B.L.V. Self-Assembly of bolaamphiphilic molecules. Chem. Rec. 2017, 17, 597–610. [Google Scholar] [CrossRef]
- Goergen, N.; Wojcik, M.; Drescher, S.; Pinnapireddy, S.R.; Brüßler, J.; Bakowsky, U.; Jedelská, J. The use of artificial gel forming bolalipids as novel formulations in antimicrobial and antifungal therapy. Pharmaceutics 2019, 11, 307. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
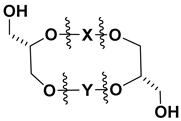 | ||
| Number | Side Chains (X, Y) | Refs. |
| 43 (Ia) | 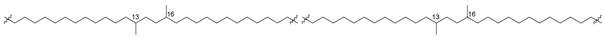 | [87,88] |
| 44 (Ib) | 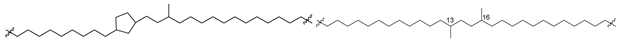 | [87,88] |
| 45 (Ic) | 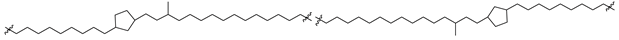 | [87,88] |
| 46 (IIa) |  | [87,88] |
| 47 (IIb) | 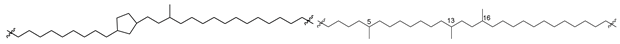 | [87,88] |
| 48 (IIc) |  | [87,88] |
| 49 (IIIa) | 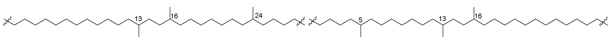 | [87,88] |
| 50 (IIIb) | 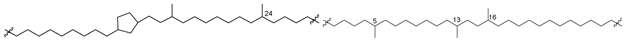 | [87,88] |
| 51 (Шс) | 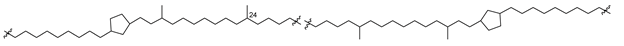 | [87,88] |
| 52 | 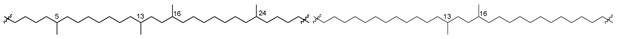 | [89] |
| 53 | 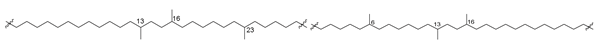 | [89] |
| 54 | 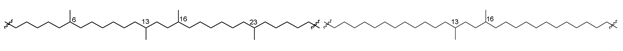 | [89] |
| Bioactivity | Compound | Refs. |
|---|---|---|
| Anticancer | rhizochalin (57) | [102] |
| rhizochalinin (58) | [104] | |
| isorhizochalin (72) | [99] | |
| melonoside A (113) | [129] | |
| Antifungal | oceanapiside (78) | [108] |
| oceanalin A (97) | [118] | |
| oceanalin B (102) | [119] | |
| Antimicrobial | leucettamol A (93) | [113] |
| leucettamol B (94) | ||
| Cancer preventive | melonosin A (115) | [130] |
| rhizochalin (57) | [105] | |
| Cardioprotective | toporoside A (117) | [131] |
| toporoside C (119) | ||
| toporoside D (120) | ||
| Drug delivery system | caldarchaeol (3) | [73,74,75,76,77,78] |
| GDGTs | ||
| crenarchaeol (14) | ||
| Inhibitor of Ubc13-Uev1A interaction | leucettamol A (93) | [115] |
| Immunomodulatory | myxillin A (110) | [128] |
| myxillin C (112) | ||
| Modulators of TRPA1 and TRPM8 channels | leucettamol A (93) | [117] |
| leucettamol B (94) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stonik, V.A.; Makarieva, T.N.; Shubina, L.K.; Guzii, A.G.; Ivanchina, N.V. Structure Diversity and Properties of Some Bola-like Natural Products. Mar. Drugs 2025, 23, 3. https://doi.org/10.3390/md23010003
Stonik VA, Makarieva TN, Shubina LK, Guzii AG, Ivanchina NV. Structure Diversity and Properties of Some Bola-like Natural Products. Marine Drugs. 2025; 23(1):3. https://doi.org/10.3390/md23010003
Chicago/Turabian StyleStonik, Valentin A., Tatyana N. Makarieva, Larisa K. Shubina, Alla G. Guzii, and Natalia V. Ivanchina. 2025. "Structure Diversity and Properties of Some Bola-like Natural Products" Marine Drugs 23, no. 1: 3. https://doi.org/10.3390/md23010003
APA StyleStonik, V. A., Makarieva, T. N., Shubina, L. K., Guzii, A. G., & Ivanchina, N. V. (2025). Structure Diversity and Properties of Some Bola-like Natural Products. Marine Drugs, 23(1), 3. https://doi.org/10.3390/md23010003