

The Effects of Agaro-Oligosaccharides Produced by Marine Bacteria (Rheinheimera sp. (HY)) Possessing Agarose-Degrading Enzymes on Myotube Function

Abstract
1. Introduction
2. Results
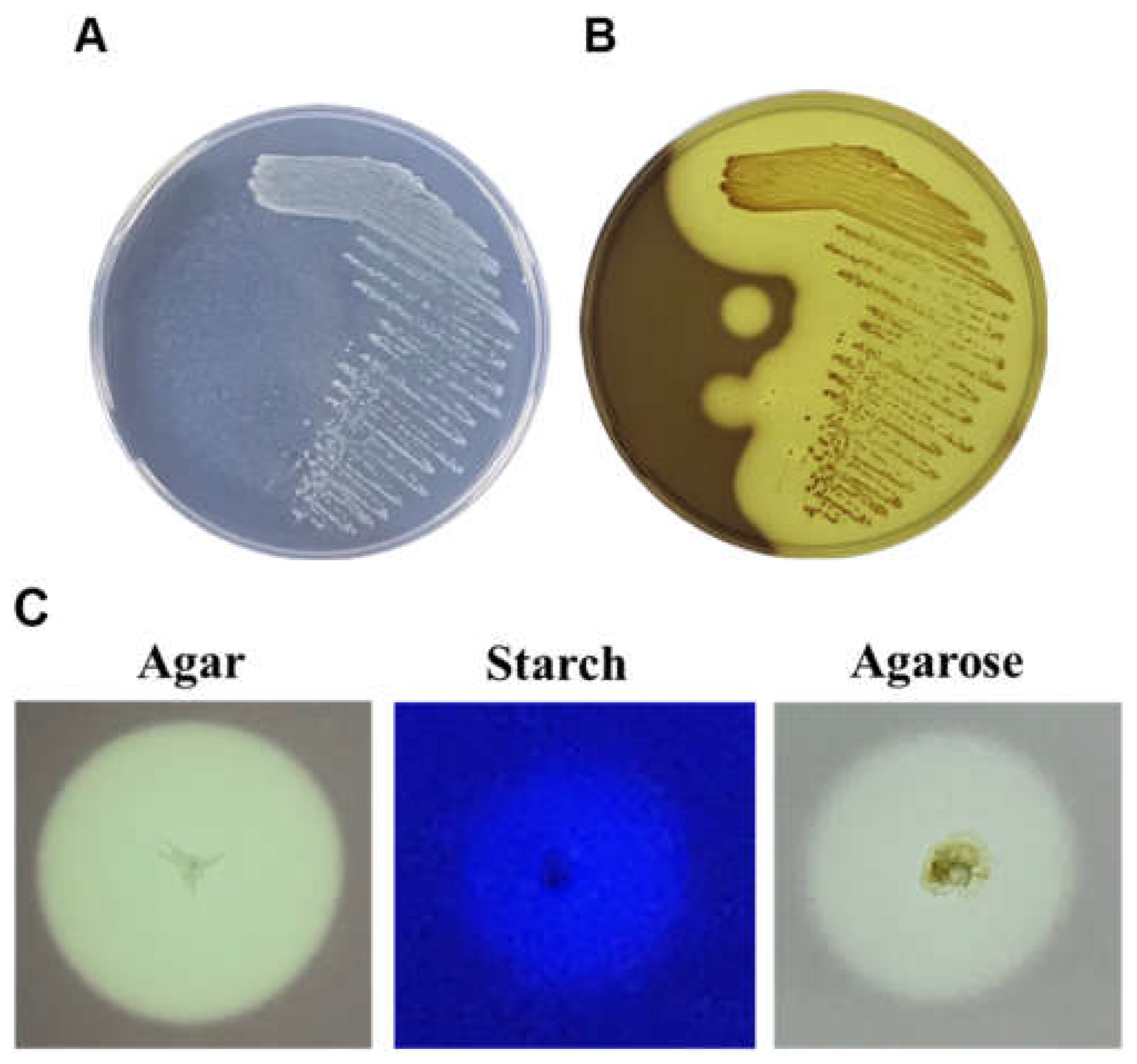
2.1. The Hydrolytic Ability of Rheinheimera sp. (HY) on Polysaccharides
2.2. Biochemical Characterization of Crude Enzyme
2.2.1. Substrate Specificity of Crude Enzyme
2.2.2. Optimal Temperature and pH of Crude Enzyme Activity
2.2.3. Effects of Chemicals on Crude Enzyme Activity
2.2.4. Analysis of Hydrolytic Activity of Agarase in Crude Enzymes
2.3. Agaro-Oligosaccharides (AOS) on Mouse C2C12 Myotubes
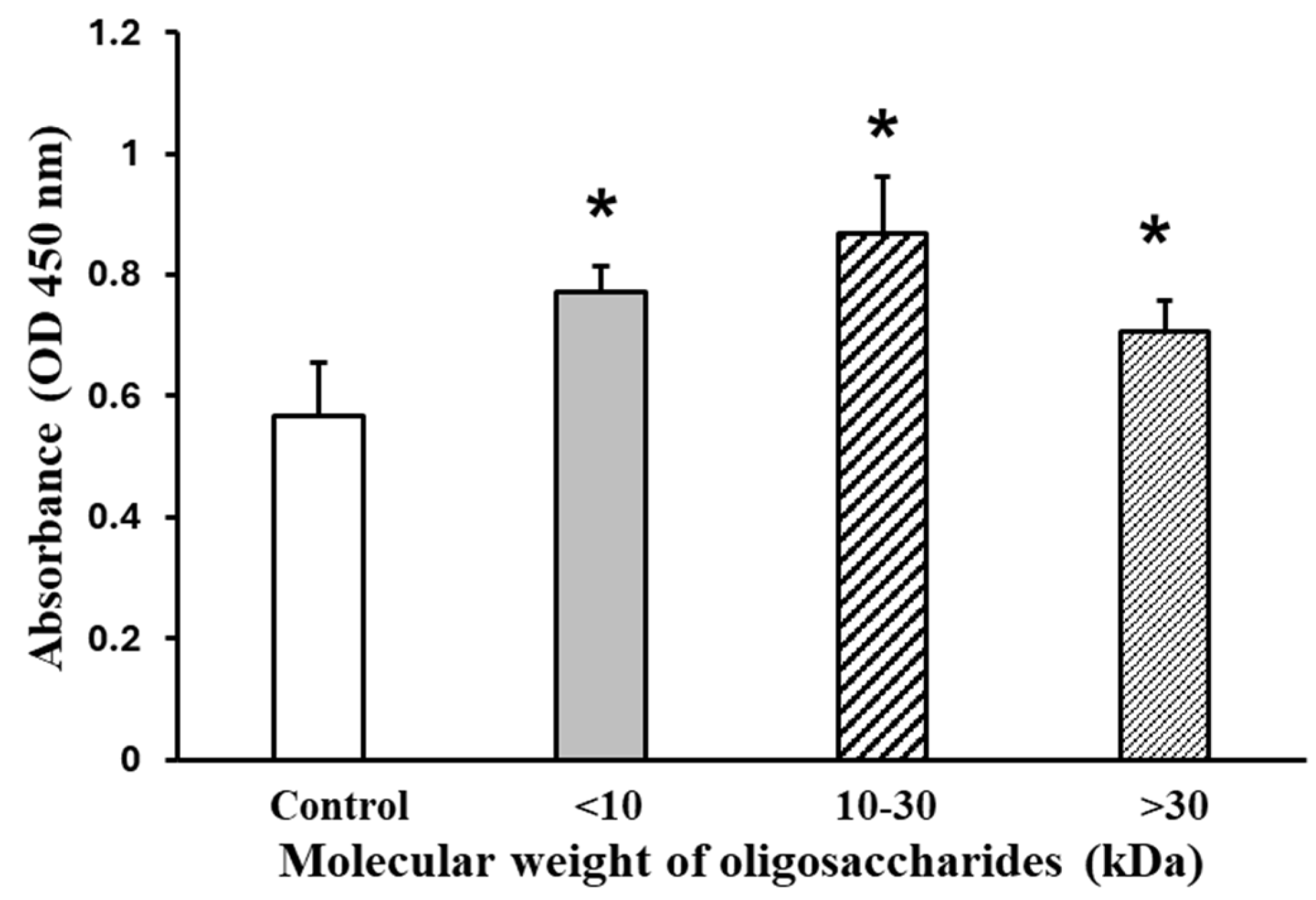
2.3.1. AOS on C2C12 Cell Viability
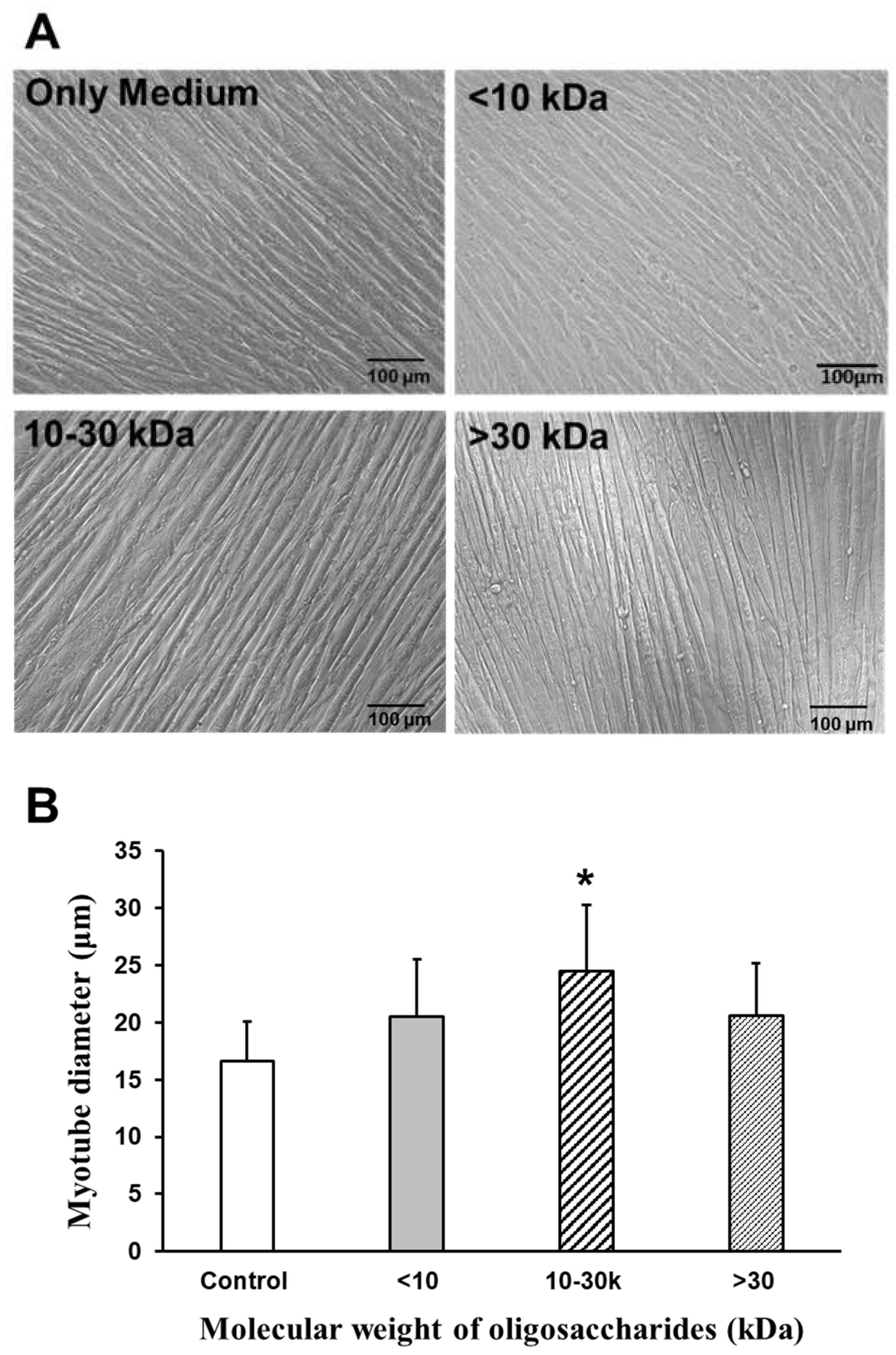
2.3.2. Effect of AOS on Diameters of Myotubes
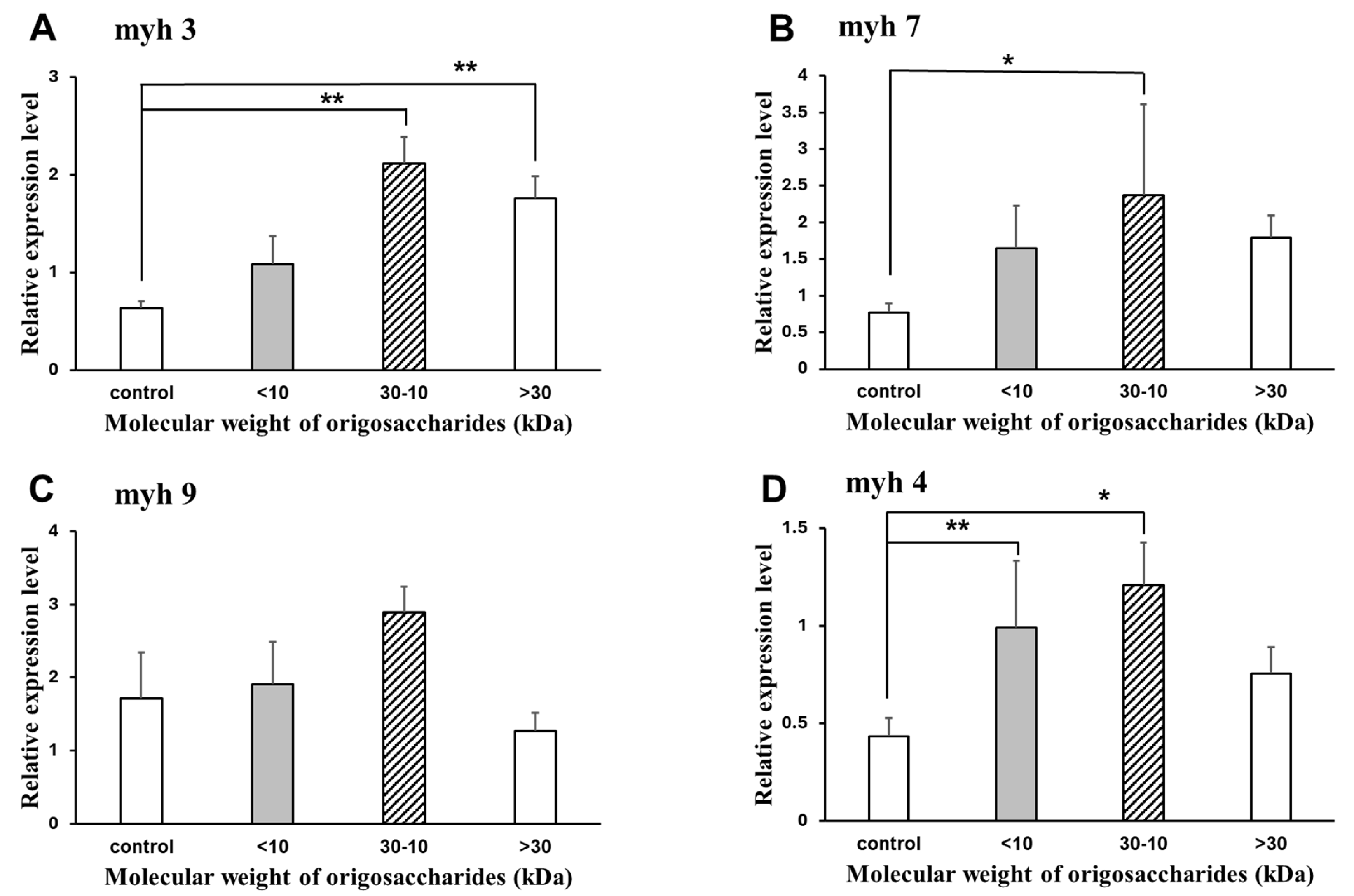
2.3.3. Relative mRNA Expression of the Genes Related to Muscle Synthesis
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolation and Identification from Seawater
4.2. Hydrolytic Ability of Bacteria
4.3. 3,5-Dinitrosalicylic Acid (DNS) Assay
4.4. Production of Crude Enzyme
4.5. Substrate Specificity of Crude Enzyme
4.6. Optimal Temperature of Crude Enzyme
4.7. Optimal pH of Crude Enzyme
4.8. Effect of Chemicals on the Crude Enzyme
4.9. Glycosidic Bonds of Crude Enzyme Activity
4.10. Preparation of Agaro-Oligosaccharide
4.11. Cell Culture
4.12. Cell Viability Assay
4.13. Measurement of Myotube Diameter
4.14. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| AOS | Agaro-oligosaccharides |
| BLAST | Basic Local Alignment Search Tool |
| CCK-8 | Cell counting kit-8 |
| DNS | 3,5-dinitrosalicylic acid |
| DMEM | Dulbecco’s modified Eagle medium |
| EDTA | ethylenediaminetetraacetic acid |
| FBS | fetal bovine serum |
| FeSO4 | ferrous sulfate |
| GLUT2 | glucose transporter 2 |
| HBSS | Hanks’ Balanced Salt Solution |
| HS | horse serum |
| KCl | potassium chloride |
| MnSO4 | manganese sulfate |
| myh3 | Myosin Heavy Chain 3 |
| myh4 | Myosin Heavy Chain 4 |
| myh7 | Myosin Heavy Chain 7 |
| myh9 | Myosin Heavy Chain 9 |
| MyHC | Myosin heavy chain |
| NAOS | neoagaro-oligosaccharides |
| NaCl | sodium chloride |
| NCBI | National Center for Biotechnology Information |
| OD | Optical density |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| SDS | sodium dodecyl sulfate |
| SGLT1 | sodium–glucose-linked transporter 1 |
References
- Sinha, V.R.; Kumria, R. Polysaccharides in colon-specific drug delivery. Int. J. Pharm. 2001, 224, 19–38. [Google Scholar] [CrossRef]
- Fu, X.T.; Kim, S.M. Agarase: Review of major sources, categories, purification method, enzyme characteristics and applications. Mar. Drugs 2010, 8, 200–218. [Google Scholar] [CrossRef] [PubMed]
- Potin, P.; Richard, C.; Rochas, C.; Kloareg, B. Purification and characterization of the α-agarase from Alteromonas agarlyticus (Cataldi) comb. nov., strain GJ1B. Eur. J. Biochem. 1993, 214, 599–607. [Google Scholar] [PubMed]
- Kirimura, K.; Masuda, N.; Iwasaki, Y.; Nakagawa, H.; Kobayashi, R.; Usami, S. Purification and characterization of a novel β-agarase from an alkalophilic bacterium, Alteromonas sp. E-1. J. Biosci. Bioeng. 1999, 87, 436–441. [Google Scholar] [PubMed]
- Chen, X.; Fu, X.; Huang, L.; Xu, J.; Gao, X. Agar oligosaccharides: A review of preparation, structures, bioactivities and application. Carbohydr. Polym. 2021, 265, 118076. [Google Scholar]
- Xu, S.; Kan, J.; Hu, Z.; Liu, Y.; Du, H.; Pang, G.C.; Cheong, K.L. Quantification of Neoagaro-Oligosaccharide Production through Enzymatic Hydrolysis and Its Anti-Oxidant Activities. Molecules 2018, 23, 1354. [Google Scholar] [CrossRef]
- Kim, K.H.; Baek, J.H.; Wenting, R.; Jeon, C.O. Rheinheimera maricola sp. nov., isolated from seawater of the Yellow Sea. Int. J. Syst. Evol. Microbiol. 2022, 72, 005423. [Google Scholar]
- Manderfeld, E.; Thamaraiselvan, C.; Kleinberg, M.N.; Jusufagic, L.; Arnusch, C.J.; Rosenhahn, A. Bacterial surface attachment and fouling assay on polymer and carbon surfaces using Rheinheimera sp. identified using bacteria community analysis of brackish water. Biofouling 2022, 38, 40–951. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, S.; Cheng, Y.; Ding, L.; Li, S.; Peng, N.; He, S. Rheinheimera mangrovi sp. nov., a bacterium isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol. 2020, 70, 6188–6194. [Google Scholar] [CrossRef]
- Fu, J.; Zhong, C.; Zhang, P.; Gao, Q.; Zong, G.; Zhou, Y.; Cao, G. A Novel Mobile Element ICERspD18B in Rheinheimera sp. D18 Contributes to Antibiotic and Arsenic Resistance. Front. Microbiol. 2020, 11, 616364. [Google Scholar] [CrossRef]
- Yu, Q.; Qi, Y.; Zhang, H.; Pu, J. Rheinheimera sediminis sp. nov., a marine bacterium isolated from coastal sediment. Int. J. Syst. Microbiol. 2020, 70, 1282–1287. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, Z.; Zou, R.; Zhang, X. The application of agar oligosaccharides in directly acidified milk drinks. Food Hydrocoll. 2018, 77, 421–426. [Google Scholar] [CrossRef]
- Yun, E.J.; Yu, S.; Kim, Y.-A.; Liu, J.-J.; Kang, N.J.; Jin, Y.-S.; Kim, K.H. In Vitro Prebiotic and Anti-Colon Cancer Activities of Agar-Derived Sugars from Red Seaweeds. Mar. Drugs 2021, 19, 213. [Google Scholar] [CrossRef]
- Lee, M.H.; Jang, J.-H.; Yoon, G.Y.; Lee, S.J.; Lee, M.-G.; Kang, T.H.; Han, H.D.; Kim, H.S.; Choi, W.S.; Park, W.S.; et al. Neoagarohexaose-mediated activation of dendritic cells via Toll-like receptor 4 leads to stimulation of natural killer cells and enhancement of antitumor immunity. BMB Rep. 2017, 50, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Yun, E.J.; Lee, S.; Kim, J.H.; Kim, B.B.; Kim, H.T.; Lee, S.H.; Pelton, J.G.; Kang, N.J.; Choi, I.-G.; Kim, K.H. Enzymatic production of 3,6-anhydro-l-galactose from agarose and its purification and in vitro skin whitening and anti-inflammatory activities. Appl. Microbiol. Biotechnol. 2013, 97, 2961–2970. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, Q.; Lin, F.; Zheng, B.; Huang, Y.; Yang, Y.; Xue, C.; Xiao, M.; Ye, J. Neoagarotetraose alleviates high fat diet induced obesity via white adipocytes browning and regulation of gut microbiota. Carbohydr. Polym. 2022, 296, 119903. [Google Scholar] [CrossRef]
- Zhang, N.; Mao, X.; Li, R.W.; Hou, E.; Wang, Y.; Xue, C.; Tang, Q. Neoagarotetraose protects mice against intense exercise-induced fatigue damage by modulating gut microbial composition and function. Mol. Nutr. Food Res. 2017, 61, 1600585. [Google Scholar] [CrossRef]
- Chen, H.; Yan, X.; Zhu, P.; Lin, J. Antioxidant activity and hepatoprotective potential of agaro-oligosaccharides in vitro and in vivo. Nutr. J. 2006, 5, 31. [Google Scholar] [CrossRef]
- Hu, B.; Gong, Q.; Wang, Y.; Ma, Y.; Li, J.; Yu, W. Prebiotic effects of neoagaro-oligosaccharides prepared by enzymatic hydrolysis of agarose. Anaerobe 2006, 12, 260–266. [Google Scholar] [CrossRef]
- Shirai, I.; Sakai, T.; Shiba, K.; Uzuhashi, Y.; Karasawa, K. Agaro-Oligosaccharides Prevent Myostatin Hyperexpression and Myosin Heavy Chain Protein Degradation in C2C12 Myotubes Induced by Tumor Necrosis Factor-α. CellBio 2018, 6, 23–24. [Google Scholar]
- Cheong, K.-L.; Qiu, H.-M.; Du, H.; Liu, Y.; Khan, B.M. Oligosaccharides Derived from Red Seaweed: Production, Properties, and Potential Health and Cosmetic Applications. Molecules 2018, 23, 2451. [Google Scholar] [CrossRef] [PubMed]
- Tanji, M.; Wada, K.; Sakamoto, K.; Ono, Y.; Inui, M. Digoxigenin-labeled RNA probes for untranslated regions enable the isoform-specific gene expression analysis of myosin heavy chains in whole-mount in situ hybridization. Dev. Growth Differ. 2022, 65, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zhang, T.; Xu, Y.; Mao, X. Characterization of a GH50 β-Agarase: A Biotechnological Tool for Preparing Oligosaccharides from Agarose and Porphyran. Biotechnol. Biol. Transform. 2022, 70, 9931–9940. [Google Scholar] [CrossRef] [PubMed]
- Alkotaini, B.; Han, N.S.; Kim, B.S. Fusion of agarase and neoagarobiose hydrolase f or mono-sugar production from agar. Appl. Microbiol. Biotechnol. 2017, 101, 1573–1580. [Google Scholar] [CrossRef]
- Liu, G.; Wu, S.; Jin, W.; Sun, C. Amy63, a novel type of marine bacterial multifunctional enzyme possessing amylase, agarase and carrageenase activities. Sci. Rep. 2016, 6, 18726. [Google Scholar] [CrossRef]
- Veerakumar, S.; Manian, R. Agarase, Amylase and Xylanase from Halomonas meridiana: A Study on Optimization of Coproduction for Biomass Saccharification. Fermentation 2022, 8, 479. [Google Scholar] [CrossRef]
- Roseline, T.L.; Sachindra, N.M. Purification and Characterization of Agarase from Marine Bacteria Acinetobacter sp. PS12B and Its Use for Preparing Bioactive Hydrolysate from Agarophyte Red Seaweed Gracilaria verrucosa. Appl. Biochem. Biotechnol. 2018, 186, 66–84. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Ma, X.; Zhang, L.; Li, F.; Liu, Z.; Mao, X. Biochemical Characterization and Substrate Degradation Mode of a Novel Exotype β-Agarase from Agarivorans gilvus WH0801. J. Agric. Food Chem. 2017, 65, 7982–7988. [Google Scholar] [CrossRef]
- Li, R.K.; Ying, X.J.; Chen, Z.L.; Ng, T.B.; Zhou, Z.M.; Ye, X.Y. Expression and Characterization of a GH16 Family β-Agarase Derived from the Marine Bacterium Microbulbifer sp. BN3 and Its Efficient Hydrolysis of Agar Using Raw Agar-Producing Red Seaweeds Gracilaria sjoestedtii and Gelidium amansii as Substrates. Catalysts 2020, 10, 885. [Google Scholar] [CrossRef]
- Sun, H.; Gao, L.; Xue, C.; Mao, X. Marine-polysaccharide degrading enzymes: Status and prospects. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2767–2796. [Google Scholar] [CrossRef]
- Aziz, G.M.; Ali, H.M. Purification and Characterization of Agarase from Bacillus sp., H12. Curr. Res. J. Biol. Sci. 2013, 5, 13–18. [Google Scholar] [CrossRef]
- Lin, Y.; Xia, J.; Yang, Z.; Wenju, J. Effect of MnSO4 on the Removal of SO2 by Manganese-Modified Activated Coke. Ind. Eng. Chem. Res. 2015, 54, 1689–1696. [Google Scholar]
- Xie, H.; Han, B.; Dong, W.; Yang, Y.; Chang, J.; Peng, Y.; Liu, W. Isolation and characterization of a marine agarase. Acta Microbiol. Sin. 2009, 49, 896–901. [Google Scholar]
- Chen, H.M.; Yan, X.J. Antioxidant activities of agaro-oligosaccharides with different degrees of polymerization in cell-based system. Biochim. Biophys. Acta 2004, 1722, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Sano, R.; Shinozaki, Y.; Ohta, T. Sodium-glucose cotransporters: Functional properties and pharmaceutical potential. J. Diabetes Investig. 2020, 11, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, S.; Chanderbhan, R.F. Chapter 20—The Biology of Nutrients: Genetic and Molecular Principles. Nutraceuticals 2021, 2021, 273–288. [Google Scholar] [CrossRef]
- Hong, S.J.; Lee, J.-H.; Kim, E.J.; Yang, H.J.; Chang, Y.-K.; Park, J.-S.; Hong, S.-K. In vitro and in vivo investigation for biological activities of neoagarooligosaccharides prepared by hydrolyzing agar with β-agarase. Biotechnol. Bioprocess Eng. 2017, 22, 489–496. [Google Scholar] [CrossRef]
- Deshavath, N.N.; Mukherjee, G.; Goud, V.V.; Veeranki, V.D.; Sastri, C.V. Pitfalls in the 3,5-dinitrosalicylic acid (DNS) assay for the reducing sugars: Interference of furfural and 5-hydroxymethylfurfural. Int. J. Biol. Macromol. 2020, 156, 180–185. [Google Scholar] [CrossRef]
- Sun, L.; Miyaji, N.; Yang, M.; Mills, E.M.; Taniyama, S.; Uchida, T.; Nikawa, T.; Li, J.; Shi, J.; Tachibana, K.; et al. Astaxanthin prevents atrophy in slow muscle fibers by inhibiting mitochondrial reactive oxygen species via a mitochondria-mediated apoptosis pathway. Nutrients 2021, 13, 379. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Relative Activity (%) a,b,c |
|---|---|
| Agarose | 100.00 ± 0.01 |
| Agar | 68.51 ± 2.50 d |
| Starch | 28.85 ± 3.58 d |
| Reagents | Relative Activity (%) a,b,c |
|---|---|
| Concentration (5 mM) | |
| None | 100.00 ± 0.01 |
| KCl | 22.59 ± 4.50 d |
| MnSO4 | 184.98 ± 12.55 e |
| FeSO4 | 580.73 ± 144.45 e |
| NaCl | 54.63 ± 9.07 |
| EDTA | 30.13 ± 4.12 d |
| SDS | 77.34 ± 5.70 |
| Urea | 51.86 ± 8.51 |
| Artificial Chromogenic Substrates | Absorbance (OD 420 nm) a |
|---|---|
| 48 h | |
| p-nitrophenyl-α-d-galactopyranoside | 0.095 ± 0.010 |
| p-nitrophenyl-β-d-galactopyranoside | 0.071 ± 0.014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Hirose, T.; Tsai, J.-M.; Hirasaka, K. The Effects of Agaro-Oligosaccharides Produced by Marine Bacteria (Rheinheimera sp. (HY)) Possessing Agarose-Degrading Enzymes on Myotube Function. Mar. Drugs 2024, 22, 515. https://doi.org/10.3390/md22110515
Huang Y, Hirose T, Tsai J-M, Hirasaka K. The Effects of Agaro-Oligosaccharides Produced by Marine Bacteria (Rheinheimera sp. (HY)) Possessing Agarose-Degrading Enzymes on Myotube Function. Marine Drugs. 2024; 22(11):515. https://doi.org/10.3390/md22110515
Chicago/Turabian StyleHuang, Youshi, Takuya Hirose, Jyh-Ming Tsai, and Katsuya Hirasaka. 2024. "The Effects of Agaro-Oligosaccharides Produced by Marine Bacteria (Rheinheimera sp. (HY)) Possessing Agarose-Degrading Enzymes on Myotube Function" Marine Drugs 22, no. 11: 515. https://doi.org/10.3390/md22110515
APA StyleHuang, Y., Hirose, T., Tsai, J.-M., & Hirasaka, K. (2024). The Effects of Agaro-Oligosaccharides Produced by Marine Bacteria (Rheinheimera sp. (HY)) Possessing Agarose-Degrading Enzymes on Myotube Function. Marine Drugs, 22(11), 515. https://doi.org/10.3390/md22110515