


Laurequinone, a Lead Compound against Leishmania

 , , ,
, , ,  ,
,  ,
, 

Abstract
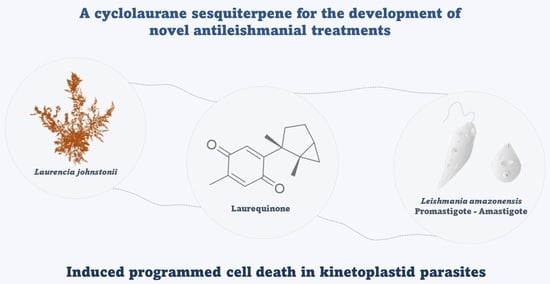
1. Introduction
2. Results
2.1. Isolation, Identification, and Semisynthesis of Active Compounds
2.2. Leishmanicidal Activity
2.3. Cell Death Mechanisms
3. Discussion
4. Materials and Methods
4.1. Extraction and Isolation of Metabolites
4.2. Conversion of Laurinterol (2) into Laurequinone (1)
4.3. Cell Cultures
4.4. Antileishmanial Activity
4.5. Cytotoxic Activity
4.6. Cell Death Mechanisms
4.6.1. ATP Level Analysis
4.6.2. Mitochondrial Membrane Potential Analysis
4.6.3. Phosphatidylserine Externalization
4.6.4. Reactive Oxygen Species Analysis
4.6.5. Chromatin Condensation Analysis
4.6.6. Plasma Membrane Permeability
4.7. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Neglected Tropical Diseases (NTDs). 2021. Available online: https://www.who.int/health-topics/neglected-tropical-diseases#tab=tab_1 (accessed on 3 April 2023).
- Pan American Health Organization—World Health Organization. Leishmaniasis—Fact Sheet for Health Workers. 2017. Available online: https://www.paho.org/en (accessed on 3 April 2023).
- Pradhan, S.; Scwartz, R.A.; Patil, A.; Grabbe, S.; Goldust, M. Treatment options for leishmaniasis. Cli. Exp. Dermatol. 2022, 47, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Marinho, F.D.E.A.; Gonçalves, K.C.; Oliveira, S.S.; Oliveira, A.C.; Bellio, M.; d’Avila-Levy, C.M.; Santos, A.L.; Branquinha, M.H. Miltefosine induces programmed cell death in Leishmania amazonensis promastigotes. Mem. Inst. Oswaldo Cruz 2011, 106, 507–509. [Google Scholar] [CrossRef] [PubMed]
- Khademvatan, S.; Gharavi, M.J.; Rahim, F.; Saki, J. Miltefosine-induced apoptotic cell death on Leishmania major and L. tropica strains. Korean J. Parasitol 2011, 49, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Santiago, A.S.; da Rocha Pita, S.S.; Guimarães, E.T. Tratamento da leishmaniose, limitações da terapêutica atual ea necessidade de novas alternativas: Uma revisão narrativa. Res. Soc. Dev. 2021, 10, e29510716543. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–808. [Google Scholar] [CrossRef]
- Marine Pharmacology. Approved Marine Drugs. 2022. Available online: https://www.marinepharmacology.org/approved (accessed on 3 April 2023).
- Tempone, A.G.; Pieper, P.; Borborema, S.E.T.; Thevenard, F.; Lago, J.H.G.; Croft, S.L.; Anderson, E.A. Marine alkaloids as bioactive agents against protozoal neglected tropical diseases and malaria. Nat. Prod. Rep. 2021, 38, 2214–2235. [Google Scholar] [CrossRef]
- Young, R.M.; Pesce, E.R. Screening for antiparasitic marine natural products. In Marine Biomedicine from Beach to Bedside; CRC Press: Boca Raton, FL, USA, 2015; Chapter 9. [Google Scholar] [CrossRef]
- García-Davis, S.; Sifaoui, I.; Reyes-Batlle, M.; Viveros-Valdez, E.; Piñero, J.E.; Lorenzo-Morales, J.; Fernández, J.J.; Díaz-Marrero, A.R. Anti-Acanthamoeba Activity of Brominated Sesquiterpenes from Laurencia johnstonii. Mar. Drugs 2018, 16, 443. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Díaz-Marrero, A.R.; Cen-Pacheco, F.; Sifaoui, I.; Reyes-Batlle, M.; Souto, M.L.; Hernández Daranas, A.; Piñero, J.E.; Fernández, J.J. Evaluation of Oxasqualenoids from the Red Alga Laurencia viridis against Acanthamoeba. Mar. Drugs 2019, 17, 420. [Google Scholar] [CrossRef]
- Díaz-Marrero, A.R.; López-Arencibia, A.; Bethencout-Estrella, C.J.; Cen-Pacheco, F.; Sifaoui, I.; Hernández Creus, A.; Duque-Ramírez, M.C.; Souto, M.L.; Hernández Daranas, A.; Lorenzo-Morales, J.; et al. Antiprotozoal activities of marine polyether triterpenoids. Bioorg. Chem. 2019, 92, 103276. [Google Scholar] [CrossRef]
- Arberas-Jiménez, I.; García-Davis, S.; Rizo-Liendo, A.; Sifaoui, I.; Reyes-Batlle, M.; Chiboub, O.; Rodríguez-Expósito, R.L.; Díaz-Marrero, A.R.; Piñero, J.E.; Fernández, J.J.; et al. Laurinterol from Laurencia johnstonii eliminates Naegleria fowleri triggering PCD by inhibition of ATPases. Sci. Rep. 2020, 10, 17731. [Google Scholar] [CrossRef]
- Arberas-Jiménez, I.; García-Davis, S.; Rizo-Liendo, A.; Sifaoui, I.; Morales, E.Q.; Piñero, J.E.; Lorenzo-Morales, J.; Díaz-Marrero, A.R.; Fernández, J.J. Cyclolauranes as plausible chemical scaffold against Naegleria fowleri. Biomed. Pharmacother. 2022, 149, 112816. [Google Scholar] [CrossRef]
- Shizuri, Y.; Yamada, A.; Yamada, K. Laurequinone, a ciclolaurane sesquiterpene from the red algae Laurencia nidifica. Phytochemistry 1984, 23, 2672–2673. [Google Scholar] [CrossRef]
- Irie, T.; Suzuki, M.; Kurosawa, E.; Masamune, T. Laurinterol and debromolaurinterol, constituennts from Laurencia intermedia. Tetrahedron Lett. 1966, 17, 1837–1840. [Google Scholar] [CrossRef]
- Irie, T.; Suzuki, M.; Kurosawa, E.; Masamune, T. Laurinterol, debromolaurinterol and isolaurinterol, constituents of Laurencia intermedia Yamada. Tetrahedron 1970, 26, 3271–3277. [Google Scholar] [CrossRef]
- Yamamura, S.; Hirata, Y. Structures of aplysin and aplysinol, naturally occurring bromocompounds. Tetrahedron 1963, 19, 1485–1496. [Google Scholar] [CrossRef]
- Tchokouaha Yamthe, L.R.; Appiah-Opong, R.; Tsouh Fokou, P.V.; Tsabang, N.; Fekam Boyom, F.; Nyarko, A.K.; Wilson, M.D. Marine algae as source of novel antileishmanial drugs: A review. Mar. Drugs 2017, 15, 323. [Google Scholar] [CrossRef]
- Ishii, T.; Nagamine, T.; Nguyen, B.C.; Tawata, S. Insecticidal and repellent activities of laurinterol from the Okinawan red alga Laurencia nidifica. Rec. Nat. Prod. 2017, 11, 63–68. [Google Scholar]
- Protopapa, M.; Kotsiri, M.; Mouratidis, S.; Roussis, V.; Ioannou, E.; Dedos, S.G. Evaluation of antifouling potential and ecotoxicity of secondary metabolites derived from red algae of the genus Laurencia. Mar. Drugs 2019, 17, 646. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Kawamoto, T.; Miwa, H.; Suzuki, M. Potent antibacterial activity of halogenated compounds against antibiotic-resistant bacteria. Planta Med. 2004, 70, 1087–1090. [Google Scholar] [CrossRef]
- García-Davis, S.; Leal-López, K.; Molina-Torres, C.A.; Vera-Cabrera, L.; Díaz-Marrero, A.R.; Fernández, J.J.; Carranza-Rosales, P.; Viveros-Valdez, E. Antimycobacterial activity of laurinterol and aplysin from Laurencia johnstonii. Mar. Drugs 2020, 18, 287. [Google Scholar] [CrossRef]
- Kladi, M.; Xenaki, H.; Vagias, C.; Papazafiri, P.; Roussis, V. New cytotoxic sesquiterpenes from the red algae Laurencia obtuse and Laurencia microcladia. Tetrahedron 2006, 62, 182–189. [Google Scholar] [CrossRef]
- García-Davis, S.; Viveros-Valdez, E.; Díaz-Marrero, A.R.; Fernández, J.J.; Valencia-Mercado, D.; Esquivel-Hernández, O.; Carranza-Rosales, P.; Carranza-Torres, I.E.; Guzmán-Delgado, N.E. Antitumoral Effect of Laurinterol on 3D Culture of Breast Cancer Explants. Mar. Drugs 2019, 17, 201. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, Y.; Nitanda, N.; Ojika, M.; Sakagami, Y. Aplysiallene, a new bromoallene as an Na, K-ATPase inhibitor from the sea hare, Aplysia kurodai. Biosci. Biotechnol. Biochem. 2001, 65, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Nwaka, S.; Hudson, A. Innovative lead discovery strategies for tropical diseases. Nat. Rev. Drug. Discov. 2006, 5, 941–955. [Google Scholar] [CrossRef]
- Nwaka, S.; Ramírez, B.; Brun, R.; Maes, L.; Douglas, F.; Ridley, R. Advancing drug innovation for neglected diseases-criteria for lead progression. PLoS Negl. Trop. Dis. 2009, 3, e440. [Google Scholar] [CrossRef]
- Álvarez-Bardón, M.; Pérez-Pertejo, Y.; Ordóñez, C.; Sepúlveda-Crespo, D.; Carballeira, N.M.; Tekwani, B.L.; Murugesan, S.; Martinez-Valladares, M.; García-Estrada, C.; Reguera, R.M.; et al. Screening marine natural products for new drug leads against trypanosomatids and malaria. Mar. Drugs 2020, 18, 187. [Google Scholar] [CrossRef]
- Santos, A.; Britta, E.A.; Bianco, E.M.; Ueda-Nakamura, T.; Filho, B.; Pereira, R.C.; Nakamura, C.V. 4-Acetoxydolastane diterpene from the Brazilian brown alga Canistrocarpus cervicornis as antileishmanial agent. Mar. Drugs 2011, 9, 2369–2383. [Google Scholar] [CrossRef]
- Sharma, N.; Shukla, A.K.; Das, M.; Dubey, V.K. Evaluation of plumbagin and its derivative as potential modulators of redox thiol metabolism of Leishmania parasite. Parasitol. Res. 2012, 110, 341–348. [Google Scholar] [CrossRef]
- Ichiba, T.; Higa, T. New cuparene-derived sesquiterpenes with unprecedented oxygenation patterns from the sea hare Aplysia dactylomela. J. Org. Chem. 1986, 51, 3364–3366. [Google Scholar] [CrossRef]
- Silverman, R.B.; Holladay, M.W. Chapter 6—DNA-Interactive Agents; Third, E., Ed.; Academic Press: Boston, MA, USA, 2014; pp. 275–331. [Google Scholar]
- Imperatore, C.; Gimmelli, R.; Persico, M.; Casertano, M.; Guidi, A.; Saccoccia, F.; Ruberti, G.; Luciano, P.; Aiello, A.; Parapini, S.; et al. Investigating the Antiparasitic Potential of the Marine Sesquiterpene Avarone, Its Reduced form Avarol, and the Novel Semisynthetic Thiazinoquinone Analogue Thiazoavarone. Mar. Drugs 2020, 18, 112. [Google Scholar] [CrossRef]
- Bethencourt-Estrella, C.J.; Delgado-Hernández, S.; López-Arencibia, A.; San Nicolás-Hernández, D.; Tejedor, D.; García-Tellado, F.; Lorenzo-Morales, J.; Piñero, J.E. In vitro activity and cell death mechanism induced by acrylonitrile derivatives against Leishmania amazonensis. Bioorg. Chem. 2022, 124, 105872. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Sahu, R.; Walker, L.A.; Tekwani, B.L. A parasite rescue and transformation assay for antileishmanial screening against intracellular Leishmania donovani amastigotes in THP1 human acute monocytic leukemia cell line. J. Vis. Exp. JOVE 2012, 70, 4054. [Google Scholar] [CrossRef]
- Sifaoui, I.; Reyes-Batlle, M.; López-Arencibia, A.; Chiboub, O.; Betherncourt-Estrella, C.J.; San Nicolás-Hernández, D.; Rodríguez-Expósito, R.L.; Rizo-Liendo, A.; Piñero, J.E.; Lorenzo-Morales, J. Screening of the pathogen box for the identification of anti-Acanthamoeba agents. Exp. Parasitol. 2019, 201, 90–92. [Google Scholar] [CrossRef] [PubMed]
- López-Arencibia, A.; Martín-Navarro, C.; Sifaoui, I.; Reyes-Batlle, M.; Wagner, C.; Lorenzo-Morales, J.; Maciver, S.K.; Piñero, J.E. Perifosine mechanisms of action in Leishmania sp. Antimicrob. Agents Chemother. 2017, 61, e02127-16. [Google Scholar] [CrossRef]
- López-Arencibia, A.; Bethencourt-Estrella, C.J.; Freijo, M.B.; Reyes-Batlle, M.; Sifaoui, I.; Nicolás-Hernández, D.S.; McNaughton-Smith, G.; Lorenzo-Morales, J.; Abad-Grillo, T.; Piñero, J.E. New phenalenone analogues with improved activity against Leishmania species. Biomed. Pharmacother. 2020, 132, 110814. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CC50, µM | IC50, µM Promastigotes | SI | IC50, µM Amastigotes | SI |
|---|---|---|---|---|---|
| Laurequinone (1) | 42.07 ± 1.30 | 1.87 ± 0.17 | 22.5 | 16.72 ± 6.08 | 2.5 |
| Laurinterol (2) | 80.11 ± 7.79 | 34.45 ± 6.44 | 2.3 | 34.72 ± 4.06 | 2.3 |
| Debromolaurinterol (3) | 70.13 ± 12.94 | 12.48 ± 0.05 | 5.6 | 24.32 ± 5.09 | 2.9 |
| Isolaurinterol (4) | 24.56 ± 2.37 | 10.09 ± 1.69 | 2.4 | Nd | Nd |
| Aplysin (5) | 1096.44 ± 40.65 | 54.13 ± 6,44 | 20.3 | Nd | Nd |
| Miltefosine | 72.18 ± 3.07 | 6.48 ± 0.25 | 11.1 | 3.12 ± 0.29 | 23.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Davis, S.; López-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Viveros-Valdez, E.; Díaz-Marrero, A.R.; Fernández, J.J.; Lorenzo-Morales, J.; Piñero, J.E. Laurequinone, a Lead Compound against Leishmania. Mar. Drugs 2023, 21, 333. https://doi.org/10.3390/md21060333
García-Davis S, López-Arencibia A, Bethencourt-Estrella CJ, San Nicolás-Hernández D, Viveros-Valdez E, Díaz-Marrero AR, Fernández JJ, Lorenzo-Morales J, Piñero JE. Laurequinone, a Lead Compound against Leishmania. Marine Drugs. 2023; 21(6):333. https://doi.org/10.3390/md21060333
Chicago/Turabian StyleGarcía-Davis, Sara, Atteneri López-Arencibia, Carlos J. Bethencourt-Estrella, Desirée San Nicolás-Hernández, Ezequiel Viveros-Valdez, Ana R. Díaz-Marrero, José J. Fernández, Jacob Lorenzo-Morales, and José E. Piñero. 2023. "Laurequinone, a Lead Compound against Leishmania" Marine Drugs 21, no. 6: 333. https://doi.org/10.3390/md21060333
APA StyleGarcía-Davis, S., López-Arencibia, A., Bethencourt-Estrella, C. J., San Nicolás-Hernández, D., Viveros-Valdez, E., Díaz-Marrero, A. R., Fernández, J. J., Lorenzo-Morales, J., & Piñero, J. E. (2023). Laurequinone, a Lead Compound against Leishmania. Marine Drugs, 21(6), 333. https://doi.org/10.3390/md21060333