
The Marine-Derived Natural Product Epiloliolide Isolated from Sargassum horneri Regulates NLRP3 via PKA/CREB, Promoting Proliferation and Anti-Inflammatory Effects of Human Periodontal Ligament Cells


Abstract
:1. Introduction
2. Results
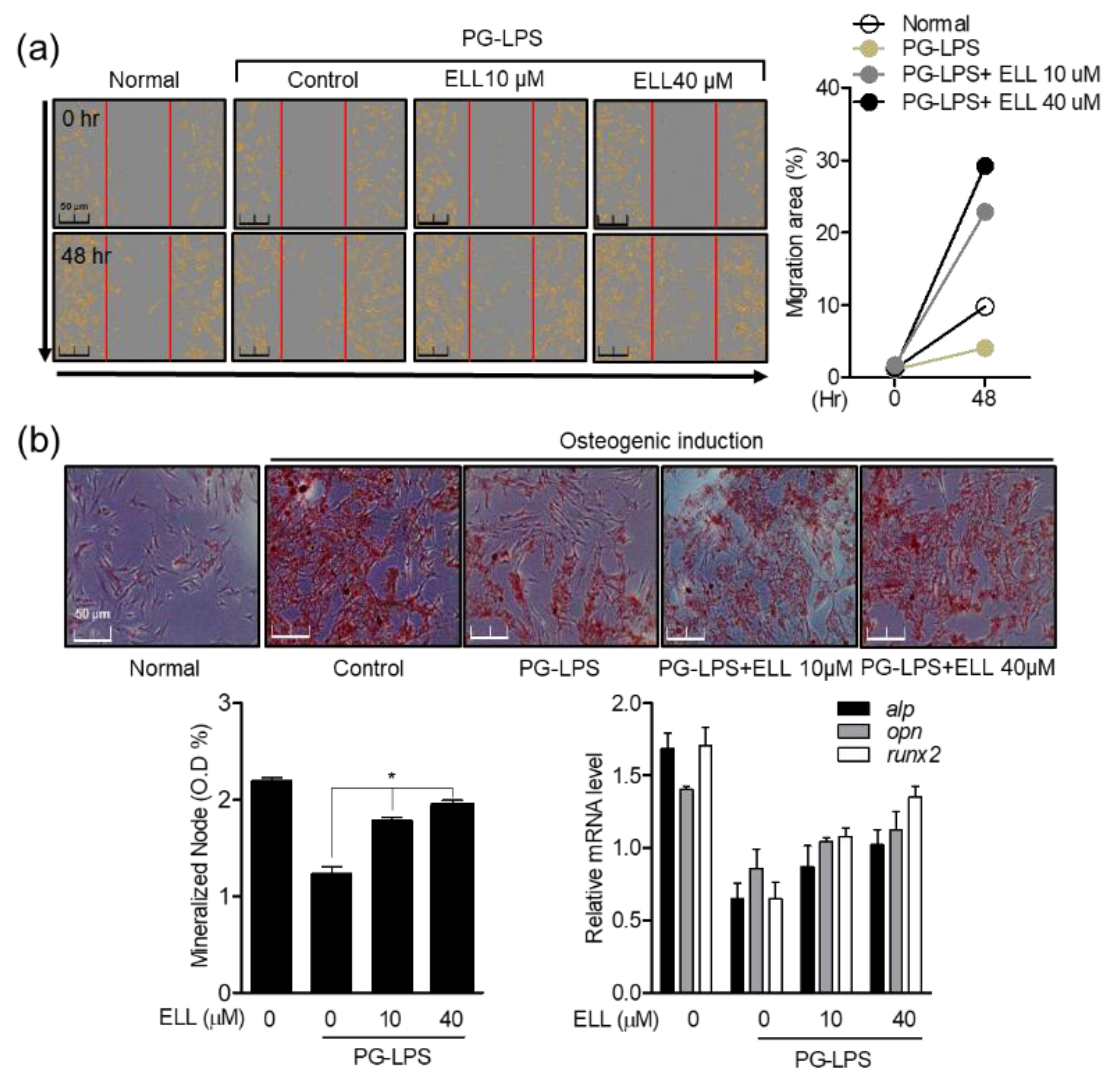
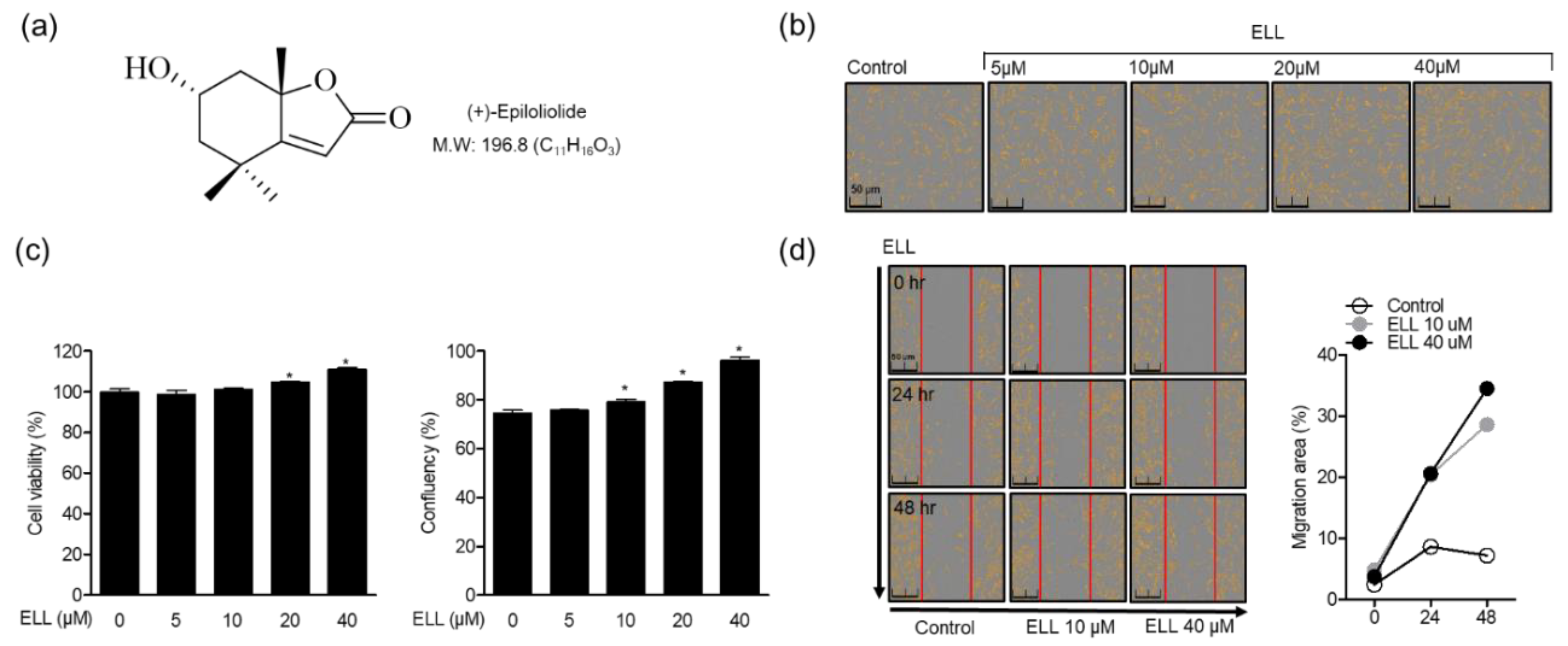
2.1. Effects of Epiloliolide on Proliferation and Migration of Human Periodontal Ligament Cells
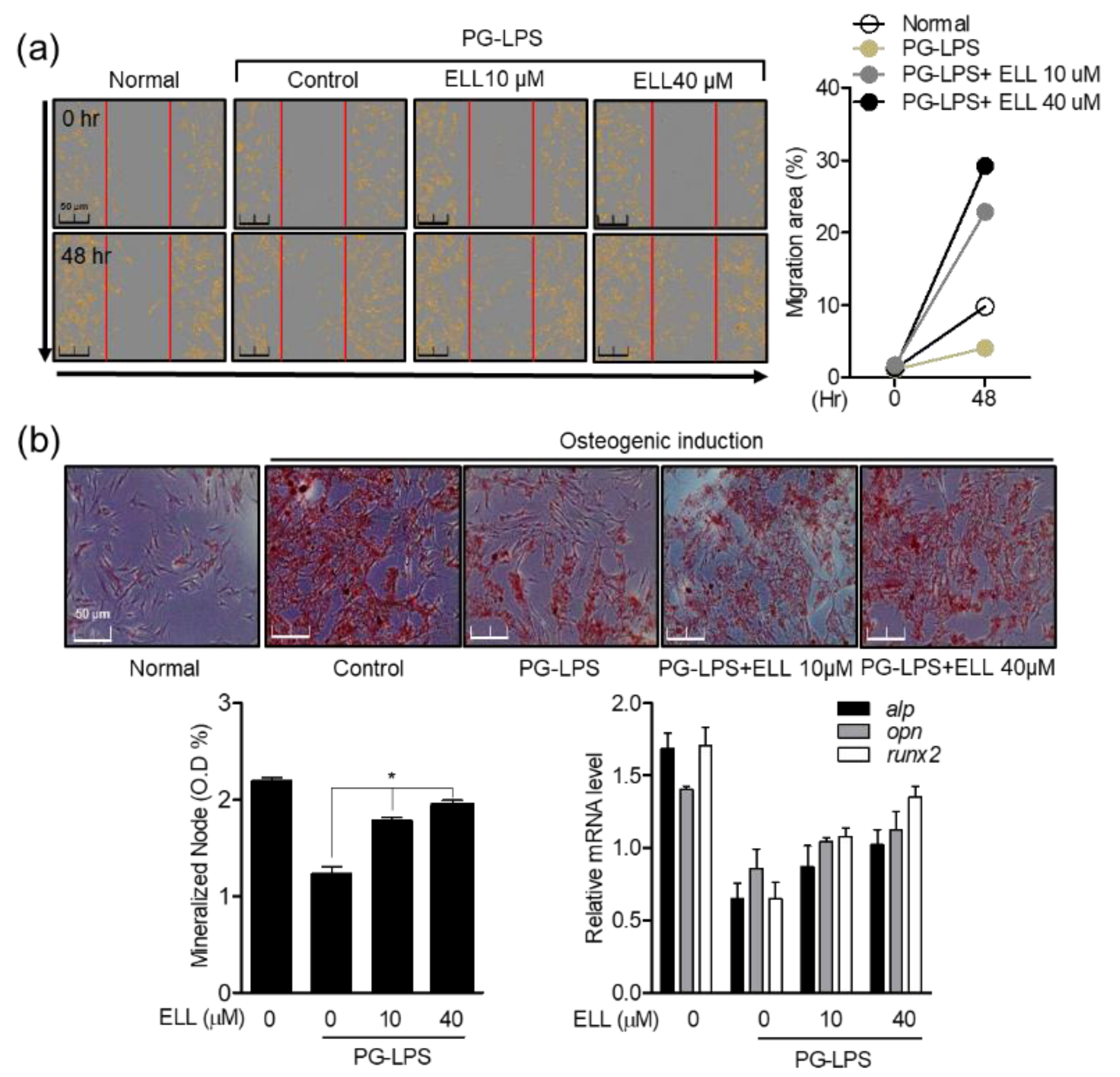
2.2. Effect of Epiloliolide on Wound Healing and Osteoblast Differentiation in HPDL Cells Stimulated with PG-LPS
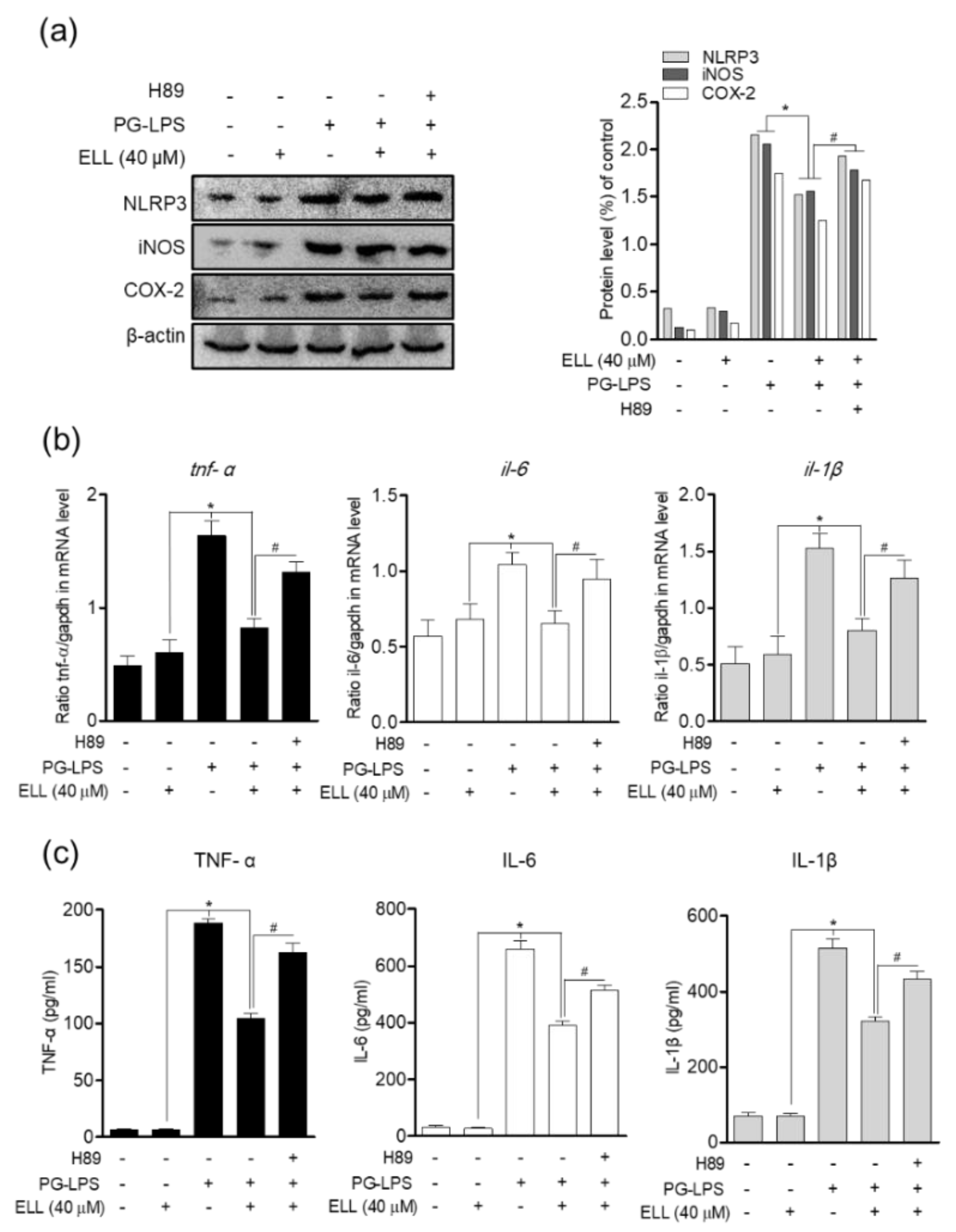
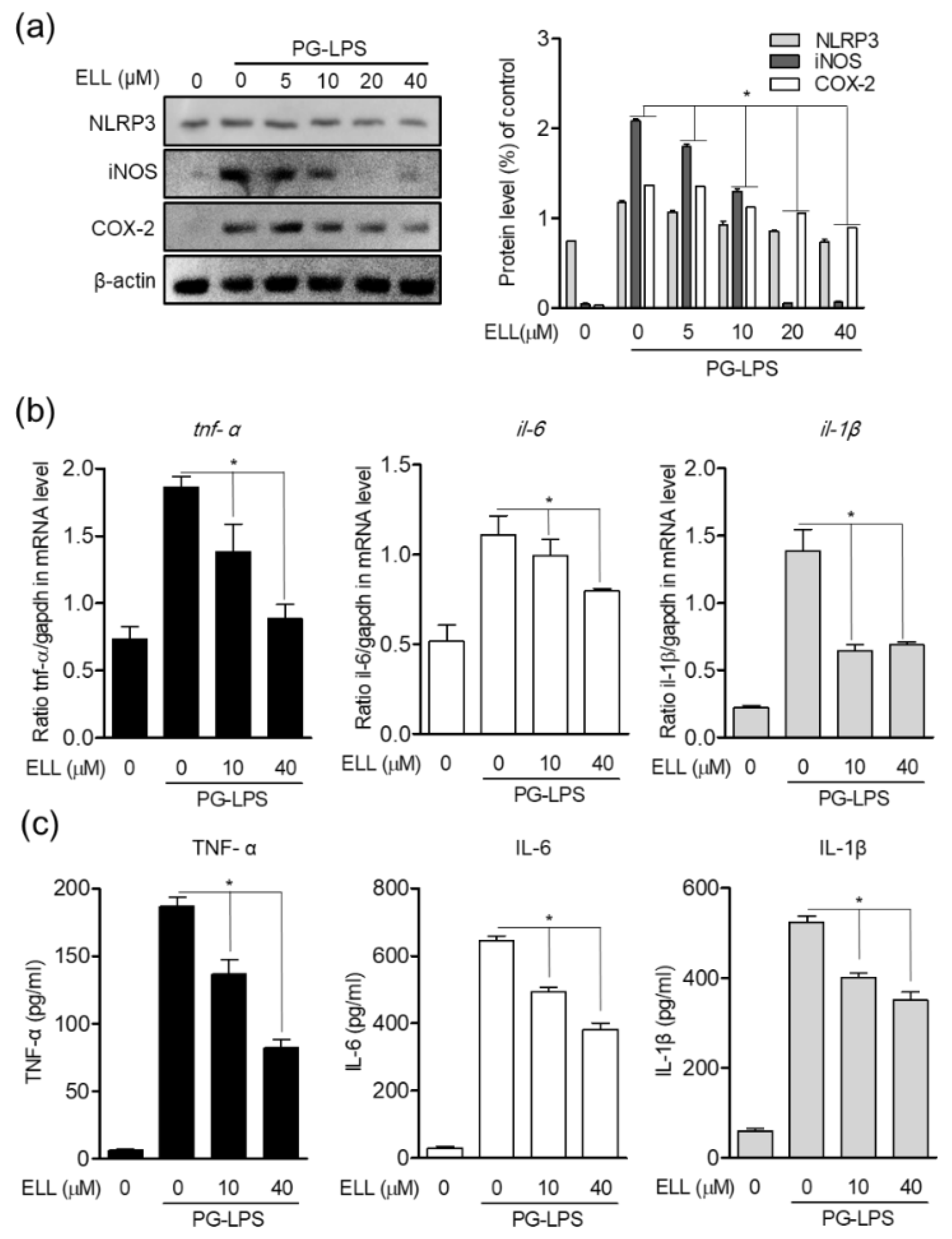
2.3. Epiloliolide Downregulates the Expression of NLRP3 Inflammasome and Pro-Inflammatory Mediators in HPDL Cells Stimulated with PG-LPS
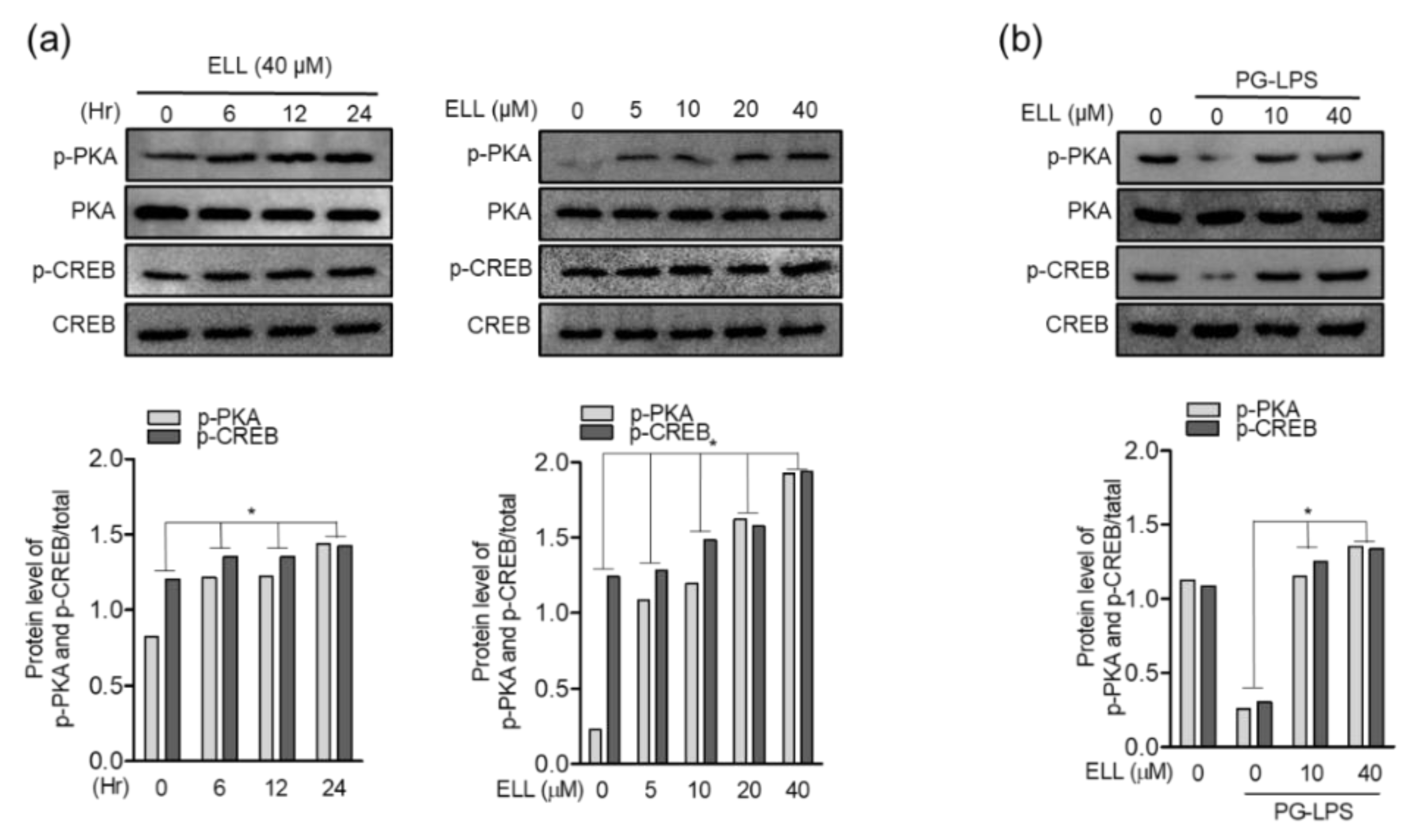
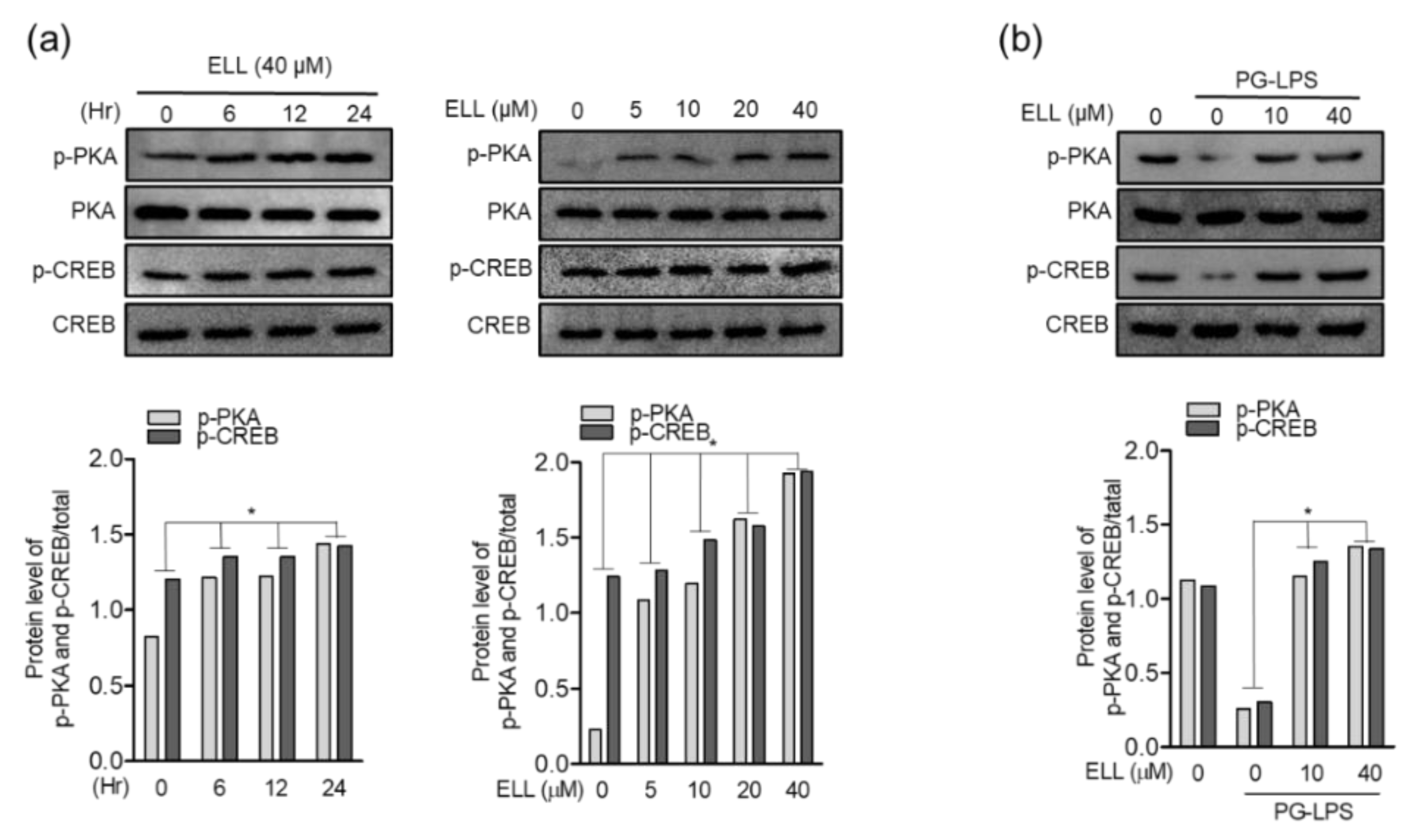
2.4. Epiloliolide Upregulates Phosphorylation of the PKA/CREB Pathway
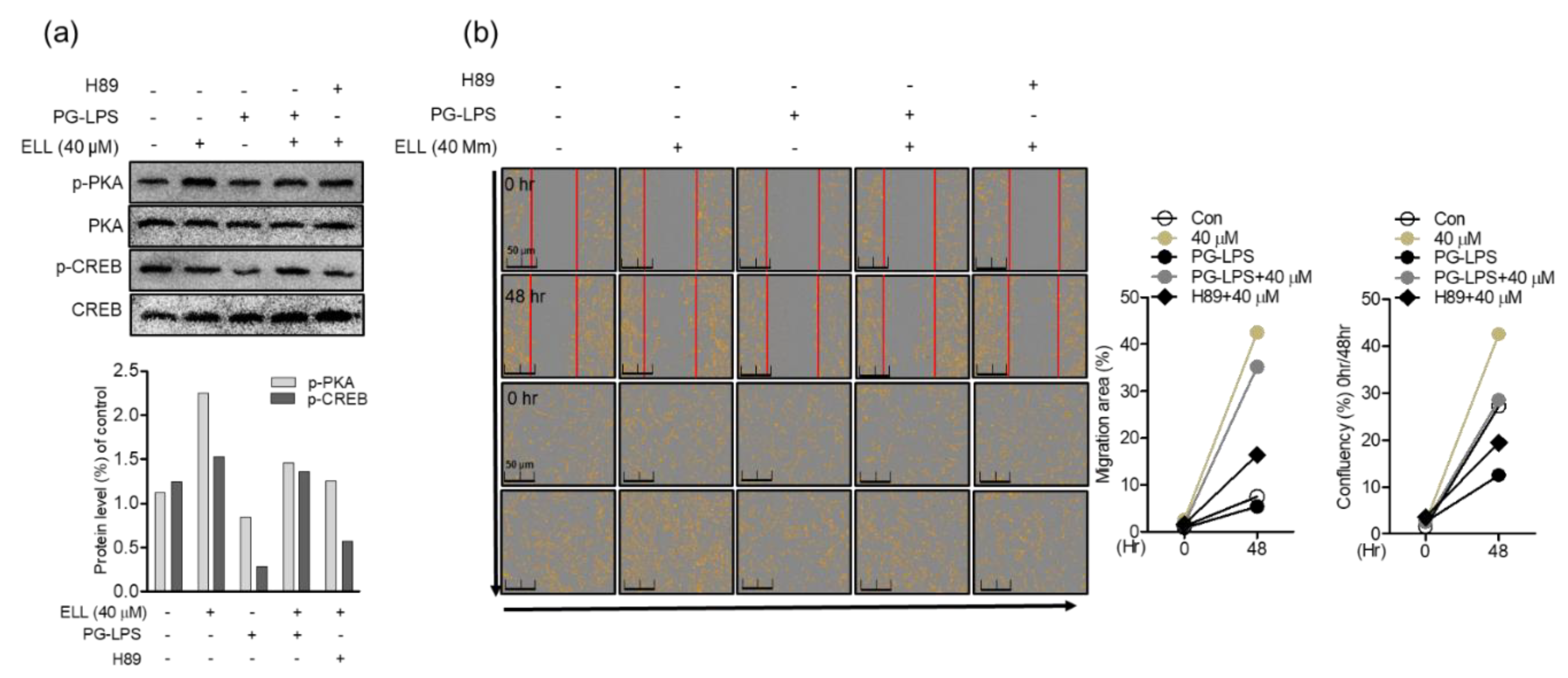
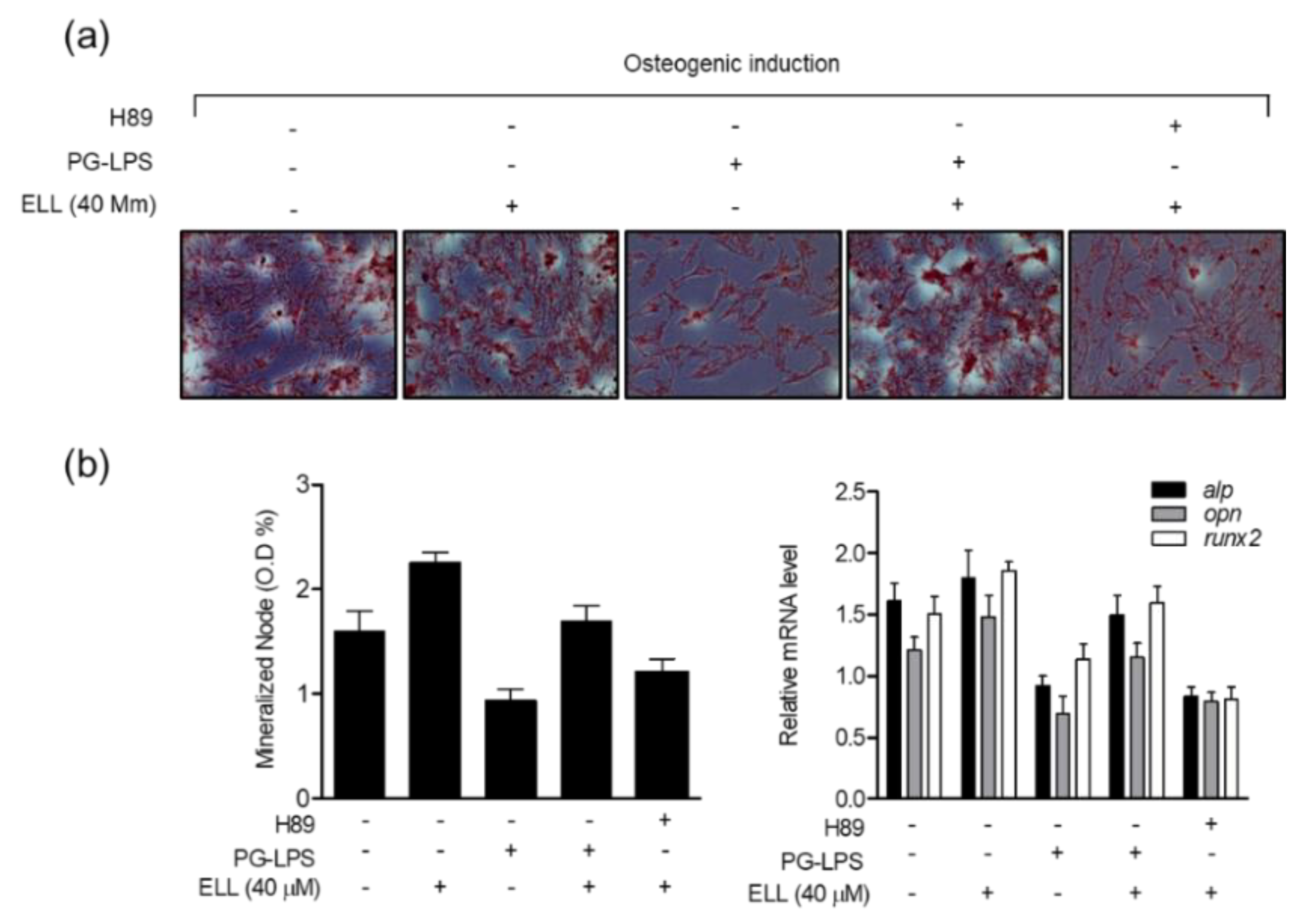
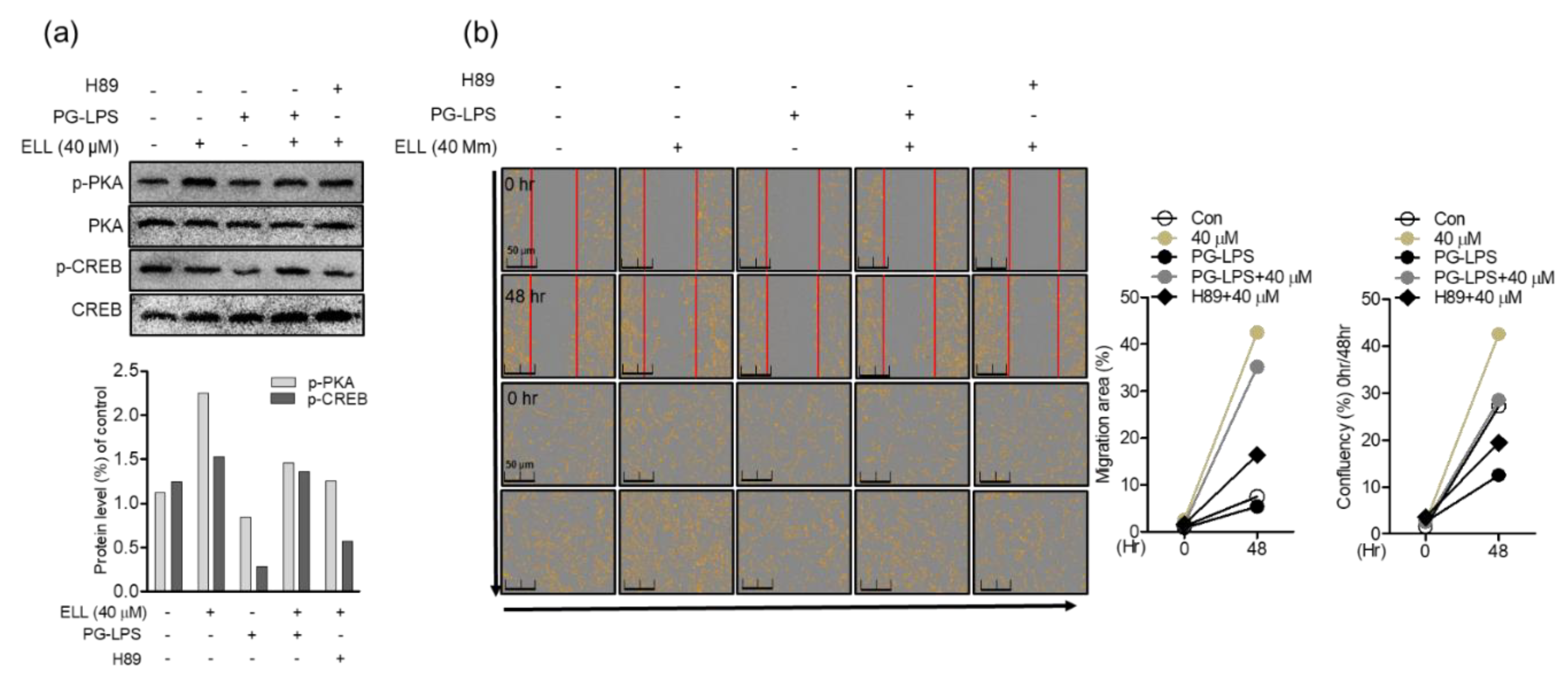
2.5. Anti-Inflammatory Effect of Epiloliolide through the PKA/CREB Signaling Pathway
2.6. Epiloliolide Induces the Proliferation of HPDL Cells through the Regulation of Phosphorylation of the PKA/CREB Signaling Pathway
2.7. Epiloliolide Induces Osteoblast Differentiation of HPDL Cells through Regulation of the PKA/CREB Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Materials
4.3. Extraction and Isolation
4.4. Structural Identification of the Compound
4.5. Preparation and Culture of Human Periodontal Ligament Cells
4.6. MTT and Coefficient Assays
4.7. Wound Healing Assays
4.8. Mineralization Assay
4.9. Western Blot Analysis
4.10. Quantitative Real-Time PCR Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graves, D.T.; Oskoui, M.; Volejnikova, S.; Naguib, G.; Cai, S.; Desta, T.; Kakouras, A.; Jiang, Y. Tumor necrosis factor modulates fibroblast apoptosis, PMN recruitment, and osteoclast formation in response to P. gingivalis infection. J. Dent. Res. 2001, 80, 1875–1879. [Google Scholar] [CrossRef]
- Graves, D.; Cochran, D. The Contribution of Interleukin-1 and Tumor Necrosis Factor to Periodontal Tissue Destruction. J. Periodontol. 2003, 74, 391–401. [Google Scholar] [CrossRef]
- Kuboniwa, M.; Lamont, R.J. Subgingival biofilm formation. Periodontol. 2000 2009, 52, 38–52. [Google Scholar] [CrossRef]
- Uchiyama, M.; Nakamichi, Y.; Nakamura, M.; Kinugawa, S.; Yamada, H.; Udagawa, N.; Miyazawa, H. Dental Pulp and Periodontal Ligament Cells Support Osteoclastic Differentiation. J. Dent. Res. 2009, 88, 609–614. [Google Scholar] [CrossRef]
- Cho, M.-I.; Garant, P.R. Development and general structure of the periodontium. Periodontol. 2000 2000, 24, 9–27. [Google Scholar] [CrossRef]
- Van Opdenbosch, N.; Gurung, P.; Walle, L.V.; Fossoul, A.; Kanneganti, T.-D.; Lamkanfi, M. Activation of the NLRP1b inflammasome independently of ASC-mediated caspase-1 autoproteolysis and speck formation. Nat. Commun. 2014, 5, 3209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Yang, X.; Ni, J.; Xie, B.; Liu, Y.; Xuan, D.; Zhang, J. Hyperglucose Contributes to Periodontitis: Involvement of the NLRP3 Pathway by Engaging the Innate Immunity of Oral Gingival Epithelium. J. Periodontol. 2015, 86, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Bostanci, N.; Emingil, G.; Saygan, B.; Turkoglu, O.; Atilla, G.; Curtis, M.A.; Belibasakis, G.N. Expression and regulation of the NALP3 inflammasome complex in periodontal diseases. Clin. Exp. Immunol. 2009, 157, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Pasqualucci, L.; Kitaura, Y.; Gu, H.; Dalla-Favera, R. PKA-mediated phosphorylation regulates the function of activation-induced deaminase (AID) in B cells. Proc. Natl. Acad. Sci. USA 2006, 103, 395–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skalhegg, B.S.; Tasken, K. Specificity in the cAMP/PKA signaling pathway. Differential expression, regulation, and subcellular localization of subunits of PKA. Front. Biosci. 2000, 5, 678–693. [Google Scholar]
- Lo, K.W.-H.; Kan, H.M.; Ashe, K.M.; Laurencin, C.T. The small molecule PKA-specific cyclic AMP analogue as an inducer of osteoblast-like cells differentiation and mineralization. J. Tissue Eng. Regen. Med. 2011, 6, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Siddappa, R.; Martens, A.; Doorn, J.; Leusink, A.; Olivo, C.; Licht, R.; van Rijn, L.; Gaspar, C.; Fodde, R.; Janssen, F.; et al. cAMP/PKA pathway activation in human mesenchymal stem cells in vitro results in robust bone formation in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 7281–7286. [Google Scholar] [CrossRef] [Green Version]
- Umezaki, I. Ecological studies of Sargassum horneri (TURNER) C. AGARDH in Obama Bay, Japan Sea. Nippon. Suisan Gakkaishi 1984, 50, 1193–1200. [Google Scholar] [CrossRef]
- Kim, H.S.; Priyan, I.; Fernando, S.; Lee, S.H.; Ko, S.C.; Kang, M.C.; Ahn, G.; Je, J.G.; Shin, H.J.; Lee, W.W.; et al. Isolation and characterization of anti-inflammatory compounds from Sargassum horneri via high-performance centrifugal partition chromatography and high-performance liquid chromatograph. Algal. Res. 2021, 54, 102209. [Google Scholar] [CrossRef]
- Shao, P.; Chen, X.; Sun, P. Chemical characterization, antioxidant and antitumor activity of sulfated polysaccharide from Sargassum horneri. Carbohydr. Polym. 2014, 105, 260–269. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.; Fernando, I.P.; Kim, E.A.; Ahn, G.; Jee, Y.; Jeon, Y.J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-S.; Sung, N.-Y.; Park, S.-Y.; Kim, G.; Eom, J.; Yoo, J.-G.; Seo, I.-R.; Han, I.-J.; Cho, Y.-B.; Kim, K.-A. Immunomodulating activity of Sargassum horneri extracts in RAW264.7 macrophages. J. Nutr. Health 2018, 51, 507–514. [Google Scholar] [CrossRef]
- Park, S.; Thomas, S.S.; Cha, Y.S.; Kim, K.A. Inhibitory effects of Sargassum horneri extract against endoplasmic reticulum stress in HepG2 cells. J. Nutr. Health 2020, 53, 583–595. [Google Scholar] [CrossRef]
- Lee, B.J. Development of Functional Food Using Fermented Marine Organism. Food Sci. Nutr. 2013, 18, 8–12. [Google Scholar]
- Yu, T.J.; Cheng, Y.B.; Lin, L.C.; Tsai, Y.H.; Yao, B.Y.; Tang, J.Y.; Chang, F.R.; Yen, C.H.; Ou-Yang, F.; Chang, H.W. Physalis peruviana-Derived Physapruin A (PHA) Inhibits Breast Cancer Cell Proliferation and Induces Oxidative-Stress-Mediated Apoptosis and DNA Damage. Antioxidants 2021, 10, 393. [Google Scholar] [CrossRef]
- Lin, H.; Zheng, Z.; Yuan, J.; Zhang, C.; Cao, W.; Qin, X. Collagen Peptides Derived from Sipunculus nudus Accelerate Wound Healing. Molecules 2021, 26, 1385. [Google Scholar] [CrossRef]
- Pradhan, B.; Patra, S.; Behera, C.; Nayak, R.; Jit, B.; Ragusa, A.; Jena, M. Preliminary Investigation of the Antioxidant, Anti-Diabetic, and Anti-Inflammatory Activity of Enteromorpha intestinalis Extracts. Molecules 2021, 26, 1171. [Google Scholar] [CrossRef] [PubMed]
- Percot, A.; Yalcin, A.; Aysel, V.; Erdugan, H.; Dural, B.; Guven, K.C.; Yalçın, A.; Güven, K.C. Loliolide in marine algae. Nat. Prod. Res. 2009, 23, 460–465. [Google Scholar] [CrossRef]
- Mori, K.; Khlebnikov, V. Carotenoids and Degraded Carotenoids, VIII–Synthesis of (+)-Dihydroactinidiolide,(+)-and (−)-Actinidiolide,(+)-and (−)-Loliolide as well as (+)-and (−)-Epiloliolide. Liebigs Annalen Chemie 1993, 1993, 77–82. [Google Scholar] [CrossRef]
- Isoe, S.; Hyeon, S.B.; Katsumura, S.; Sakan, T. Photo-oxygenation of carotenoids. II. The absolute configuration of loliolide and dihydroactinidiolide. Tetrahedron Lett. 1972, 13, 2517–2520. [Google Scholar] [CrossRef]
- Grabarczyk, M.; Wińska, K.; Mączka, W.; Potaniec, B.; Anioł, M. Loliolide—The most ubiquitous lactone. Folia Biol. Oecologica 2015, 11, 1–8. [Google Scholar] [CrossRef]
- Park, K.-E.; Kim, Y.A.; Jung, H.A.; Lee, H.-J.; Ahn, J.-W.; Lee, B.; Seo, Y. Three Norisoprenoids compounds isolated from brown algae insects. J. Korean Chem. Soc. 2004, 48, 394–398. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Huang, R.-M.; Lin, X.-P.; Liu, Y.-H. Norisoprenoids from the Brown Alga Sargassum naozhouense Tseng et Lu. Molecules 2018, 23, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuniyoshi, M. Germination Inhibitors from the Brown Alga Sargassum crassifolium (Phaeophyta, Sargassaceae). Bot. Mar. 1985, 28. [Google Scholar] [CrossRef]
- Gangadhar, K.N.; Rodrigues, M.J.; Pereira, H.; Gaspar, H.; Malcata, F.X.; Barreira, L.; Varela, J. Anti-Hepatocellular Carci-noma (HepG2) Activities of Monoterpene Hydroxy Lactones Isolated from the Marine Microalga Tisochrysis Lutea. Mar. Drugs 2020, 18, 567. [Google Scholar] [CrossRef]
- Ko, R.K.; Kang, M.-C.; Kim, S.S.; Oh, T.H.; Kim, G.-O.; Hyun, C.-G.; Hyun, J.W.; Lee, N.H. Anti-melanogenesis Constituents from the Seaweed Dictyota Coriacea. Nat. Prod. Commun. 2013, 8, 427–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenkein, H.A. Host responses in maintaining periodontal health and determining periodontal disease. Periodontol. 2000 2006, 40, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Abais, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.-L. Redox Regulation of NLRP3 Inflammasomes: ROS as Trigger or Effector? Antioxid. Redox Signal. 2015, 22, 1111–1129. [Google Scholar] [CrossRef] [Green Version]
- Bostanci, N.; Belibasakis, G.N. Porphyromonas gingivalis: An invasive and evasive opportunistic oral pathogen. FEMS Microbiol. Lett. 2012, 333, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, D.; Daia, L.; Xie, Z.; Zhou, X.; Liu, X.; Zhang, Y.; Huang, Y.; Chen, Y. Periodontal ligament fibroblasts migration injury via ROS/TXNIP/Nlrp3 inflammasome pathway with Porphyromonas gingivalis lipopolysaccharide. Mol. Immunol. 2018, 103, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hao, L.; Wu, J.; Zhang, J.; Su, J. Linarin promotes osteogenic differentiation by activating the BMP-2/RUNX2 pathway via protein kinase A signaling. Int. J. Mol. Med. 2016, 37, 901–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Cui, Y.; Li, H.; Luan, J.; Zhou, X.; Han, J. Icariin Promotes the Osteogenic Action of BMP2 by Activating the cAMP Signaling Pathway. Molecules 2019, 24, 3875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zhang, L.; Wang, Z.; Li, C.; Li, S.; Li, L.; Fan, Q.; Zheng, L. Melatonin Suppresses Estrogen Deficiency-Induced Os-teoporosis and Promotes Osteoblastogenesis by Inactivating the NLRP3 Inflammasome. Calcif. Tissue Int. 2018, 103, 400–410. [Google Scholar] [CrossRef]
- Ran, S.; Chu, M.; Gu, S.; Wang, J.; Liang, J. Enterococcus faecalis induces apoptosis and pyroptosis of human osteoblastic MG63 cells via the NLRP3 inflammasome. Int. Endod. J. 2019, 52, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Kimura, J.; Maki, N. New Loliolide Derivatives from the Brown Alga Undaria pinnatifida. J. Nat. Prod. 2002, 65, 57–58. [Google Scholar] [CrossRef]
- Seo, B.M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Inves-tigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence (5′→3′) | |
|---|---|---|
| il-6 | Forward | AGTGAGGAACAAGCCAGAGC |
| Reverse | GTCAGGGGTGGTTATTGCAT | |
| il-1β | Forward | AACCTCTTCGAGGCACAAGG |
| Reverse | GTCCTGGAAGGAGCACTTCAT | |
| tnf-α | Forward | GCCTCTTCTCCTTCCTGATCGT |
| Reverse | TGAGGGTTTGCTACAACATGGG | |
| alp | Forward | TGCAGTACGAGCTGAACAGG |
| Reverse | GTCAATTCTGCCTCCTTCCA | |
| opn | Forward | TCAGCTGGATGACCAGAGTG |
| Reverse | TTGGGGTCTACAACCAGCAT | |
| runx2 | Forward | TCTTAGAACAAATTCTGCCCTTT |
| Reverse | TGCTTTGGTCTTGAAATCACA | |
| gapdh | Forward | TGTTCGTCATGGGTGTGAAC |
| Reverse | GTCTTCTGGGTGGCAGTGAT | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-N.; Nabende, W.Y.; Jeong, H.; Hahn, D.; Jeong, G.-S. The Marine-Derived Natural Product Epiloliolide Isolated from Sargassum horneri Regulates NLRP3 via PKA/CREB, Promoting Proliferation and Anti-Inflammatory Effects of Human Periodontal Ligament Cells. Mar. Drugs 2021, 19, 388. https://doi.org/10.3390/md19070388
Kim E-N, Nabende WY, Jeong H, Hahn D, Jeong G-S. The Marine-Derived Natural Product Epiloliolide Isolated from Sargassum horneri Regulates NLRP3 via PKA/CREB, Promoting Proliferation and Anti-Inflammatory Effects of Human Periodontal Ligament Cells. Marine Drugs. 2021; 19(7):388. https://doi.org/10.3390/md19070388
Chicago/Turabian StyleKim, Eun-Nam, Woguti Yvonne Nabende, Hyeyoon Jeong, Dongyup Hahn, and Gil-Saeng Jeong. 2021. "The Marine-Derived Natural Product Epiloliolide Isolated from Sargassum horneri Regulates NLRP3 via PKA/CREB, Promoting Proliferation and Anti-Inflammatory Effects of Human Periodontal Ligament Cells" Marine Drugs 19, no. 7: 388. https://doi.org/10.3390/md19070388
APA StyleKim, E.-N., Nabende, W. Y., Jeong, H., Hahn, D., & Jeong, G.-S. (2021). The Marine-Derived Natural Product Epiloliolide Isolated from Sargassum horneri Regulates NLRP3 via PKA/CREB, Promoting Proliferation and Anti-Inflammatory Effects of Human Periodontal Ligament Cells. Marine Drugs, 19(7), 388. https://doi.org/10.3390/md19070388