
Solid-Phase Extraction Embedded Dialysis (SPEED), an Innovative Procedure for the Investigation of Microbial Specialized Metabolites

 , and
, and 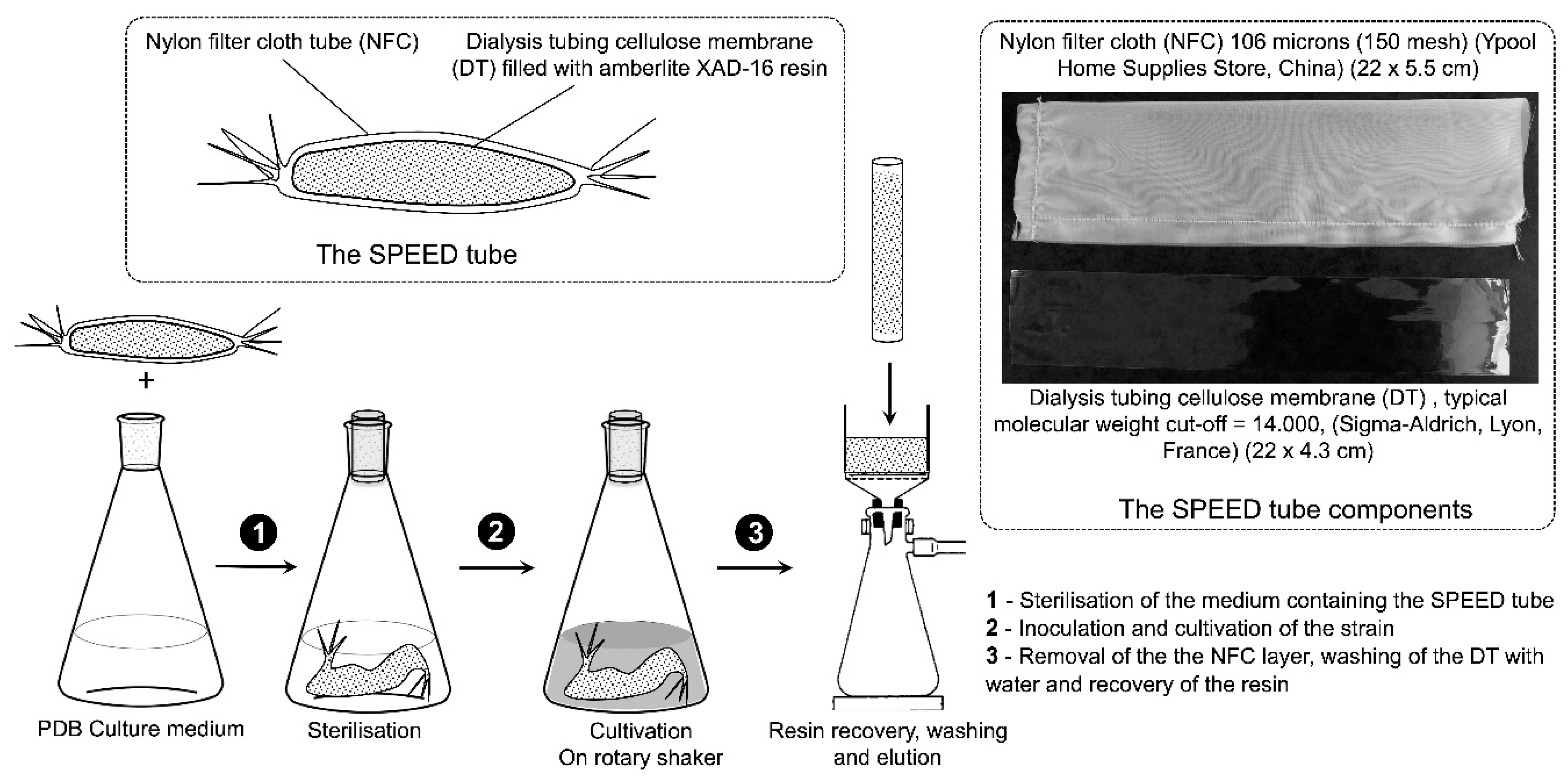{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussions
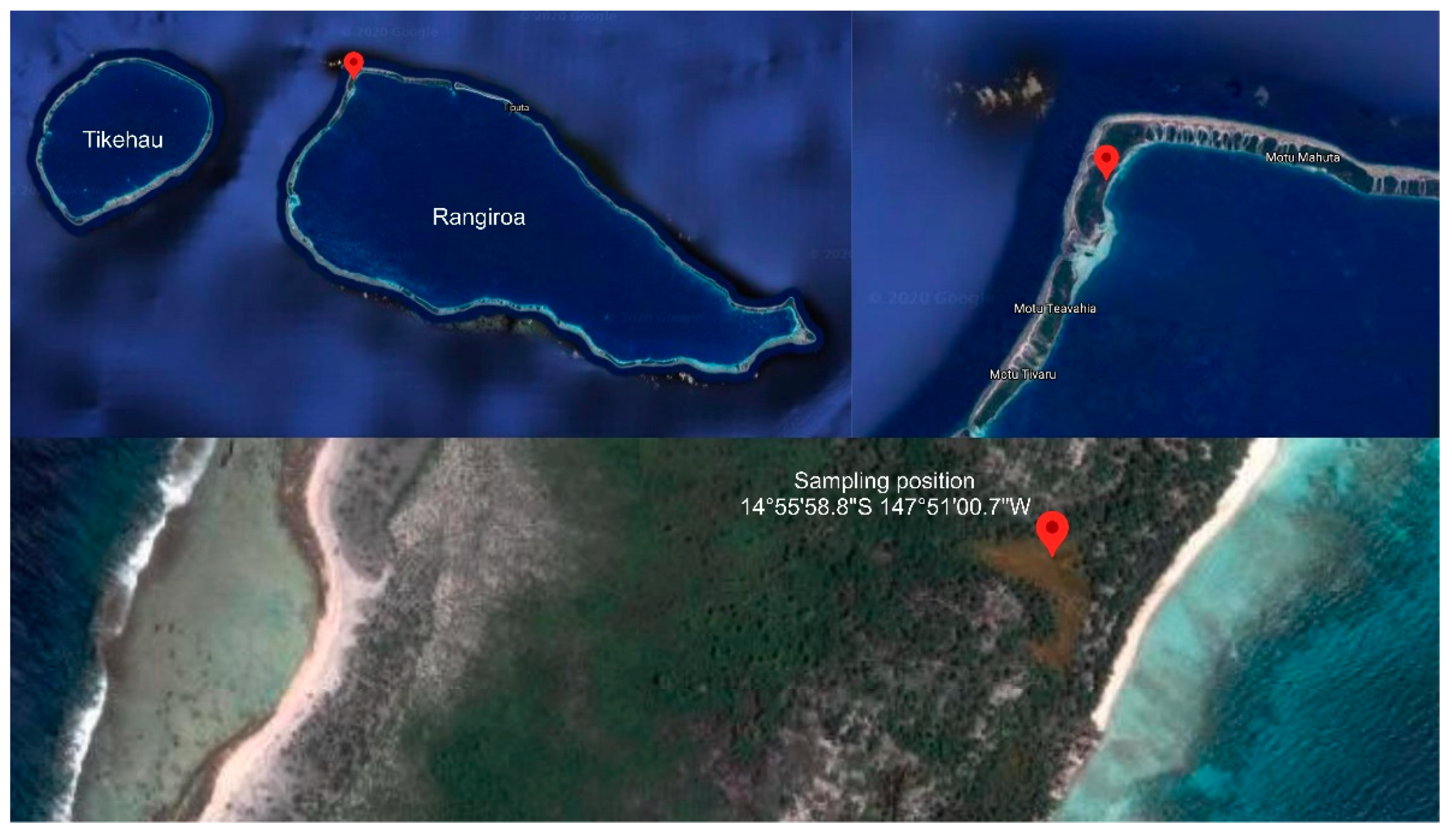
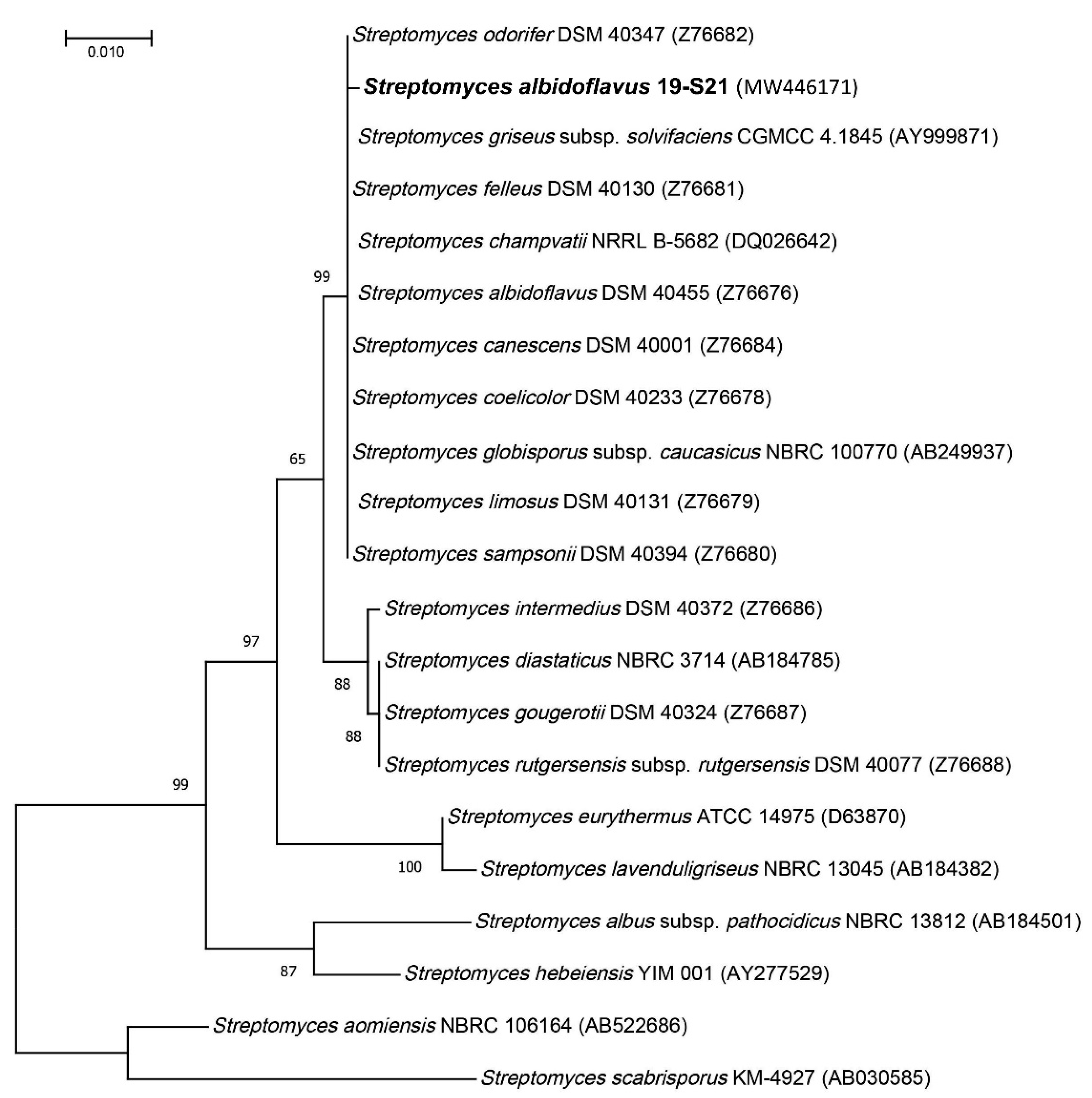
2.1. Collection Site and Strain Identification
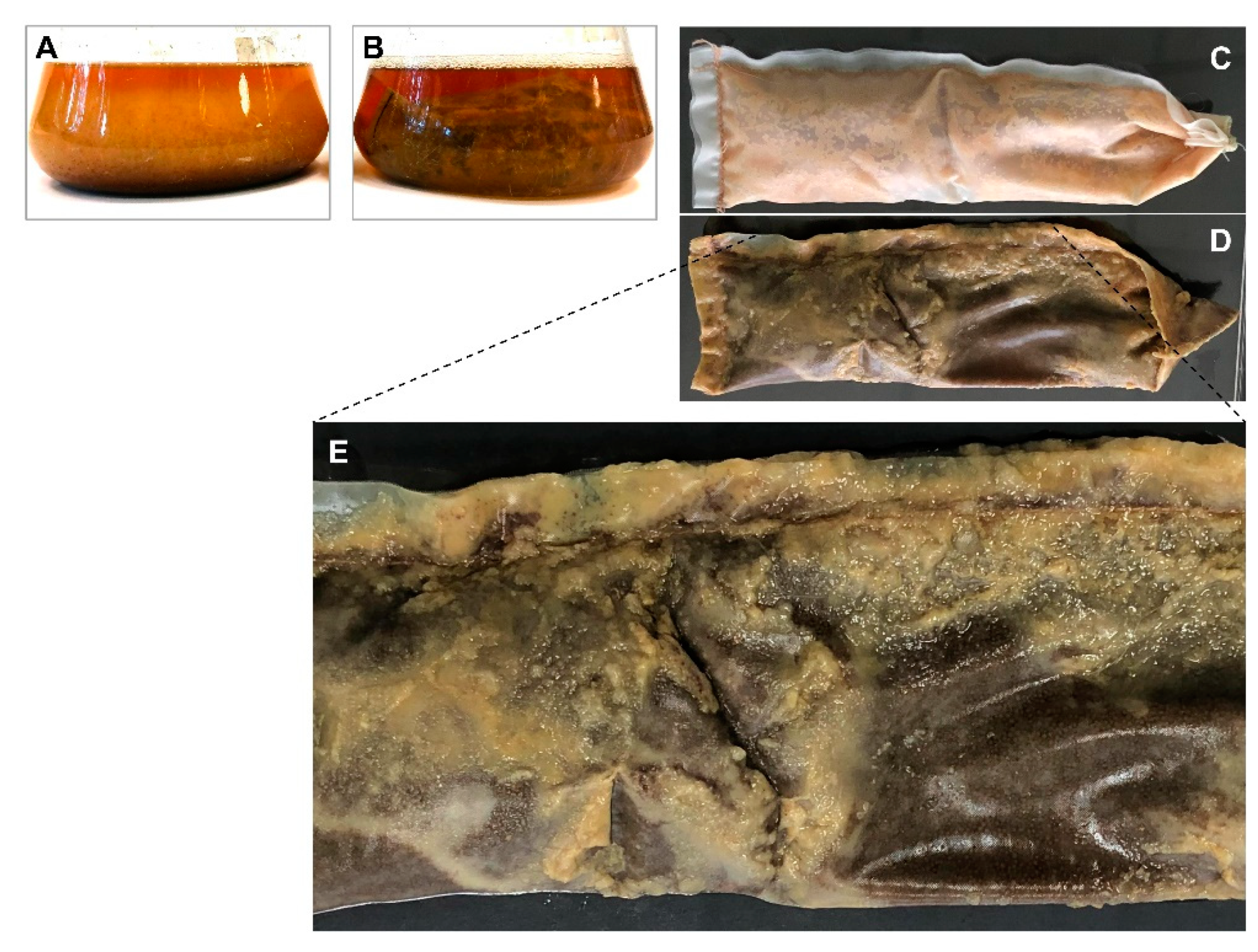
2.2. S. albidoflavus 19-S21 Cultivation according to SPEED Technology
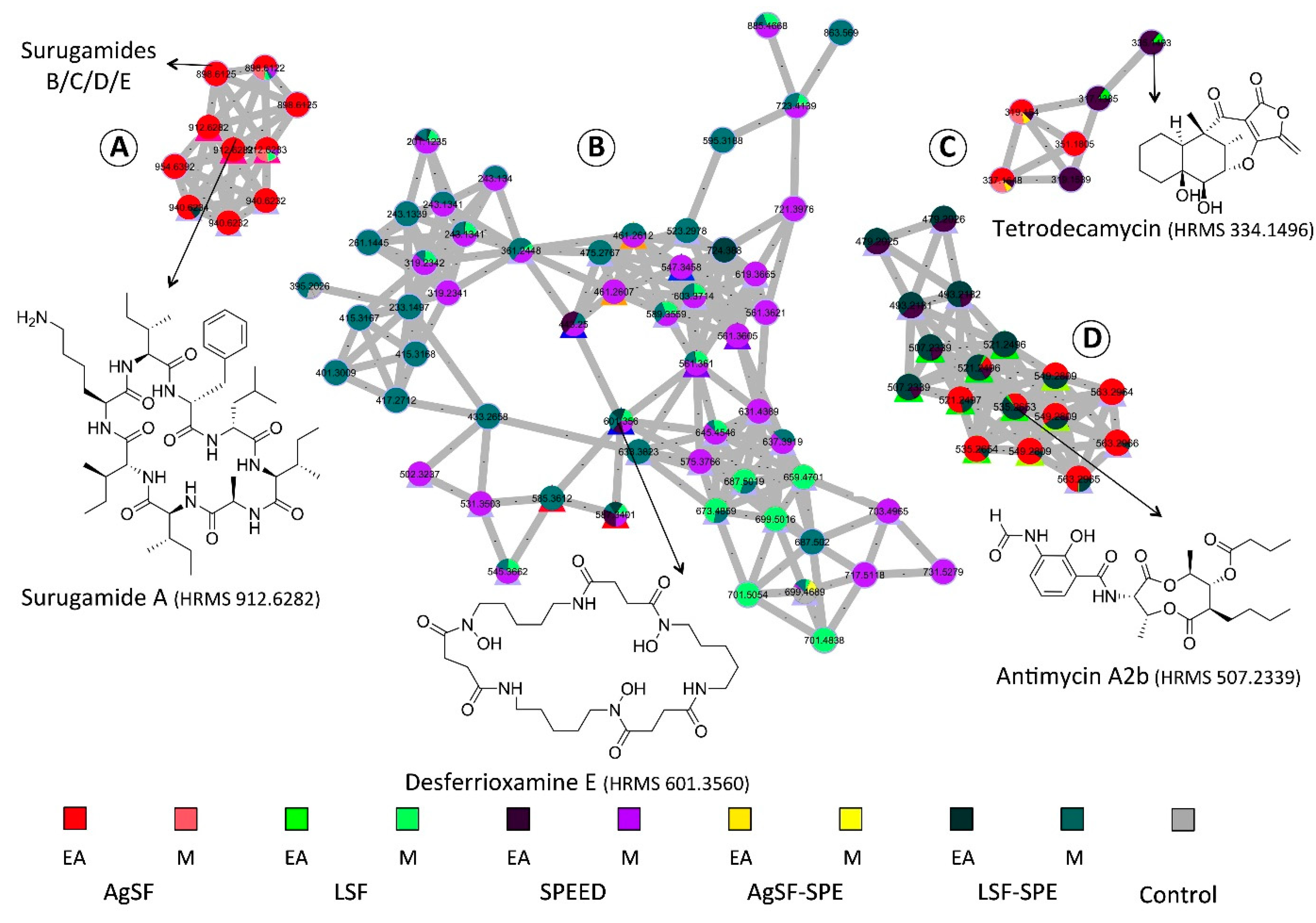
2.3. Molecular Networking-based Chemical Exploration of S. albidoflavus 19-S21 Specialized Metabolites
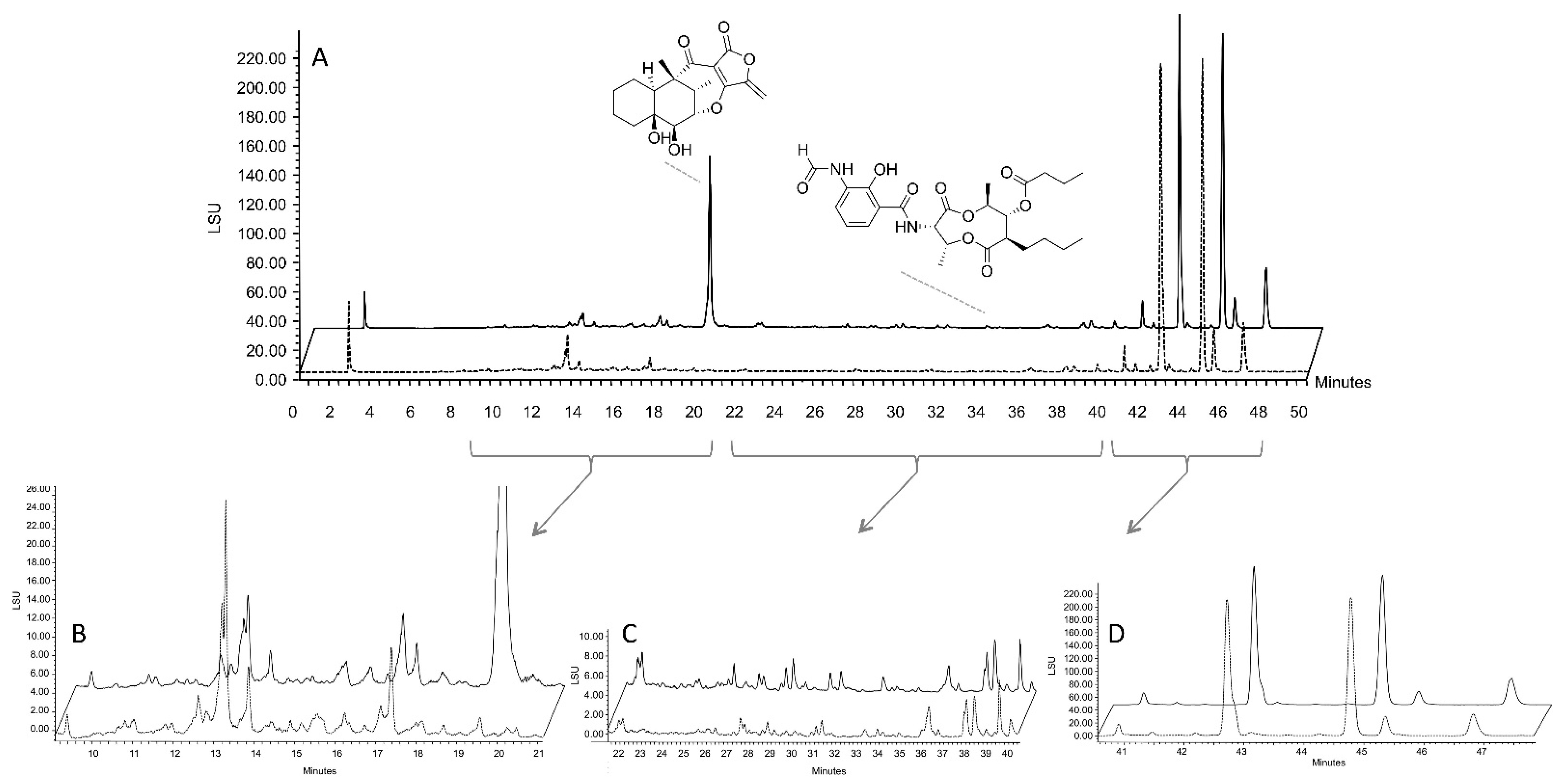
2.4. Isolation of Representative Compounds
3. Discussion
4. Materials and Methods
4.1. Strain Isolation
4.2. Phylogeny Investigation
4.3. Strain Cultivation with In-Situ SPEED Technology
- -
- Agar-state fermentation (AgSF);
- -
- Liquid-state fermentation (LSF);
- -
- SPEED cultivation;
- -
- Agar-state fermentation coupled to SPE (AgSF-SPE) [11];
- -
- Liquid-state fermentation coupled to SPE (LSF-SPE).
4.4. Extraction/Purification Procedures
4.5. Characterization, Isolation, and Structural Elucidation Experiments
4.6. Data Dependent LC-ESI-HRMS2 Analysis
4.7. MS Data Processing and Feature-Based Molecular Networking—GNPS
4.8. Molecular Networking Parameters
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Romano, J.D.; Tatonetti, N.P. Informatics and Computational Methods in Natural Product Drug Discovery: A Review and Perspectives. Front. Genet. 2019, 10, 368. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, T.; Lewis, K.; Epstein, S.S. Isolating “Uncultivable” Microorganisms in Pure Culture in a Simulated Natural Environment. Science 2002, 296, 1127–1129. [Google Scholar] [CrossRef]
- Hernandez, A.; Nguyen, L.T.; Dhakal, R.; Murphy, B.T. The need to innovate sample collection and library generation in microbial drug discovery: A focus on academia. Nat. Prod. Rep. 2021, 38, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Komaraiah, P.; Ramakrishna, S.V.; Reddanna, P.; Kavi Kishor, P.B. Enhanced production of plumbagin in immobilized cells of Plumbago rosea by elicitation and in situ adsorption. J. Biotechnol. 2003, 101, 181–187. [Google Scholar] [CrossRef]
- Klvana, M.; Legros, R.; Jolicoeur, M. In situ extraction strategy affects benzophenanthridine alkaloid production fluxes in suspension cultures of Eschscholtzia californica. Biotechnol. Bioeng. 2005, 89, 280–289. [Google Scholar] [CrossRef]
- Vlachou, P.; Le Goff, G.; Alonso, C.; Alvarez, P.A.; Gallard, J.F.; Fokialakis, N.; Ouazzani, J. Innovative Approach to Sustainable Marine Invertebrate Chemistry and a Scale-Up Technology for Open Marine Ecosystems. Mar. Drugs 2018, 16, 152. [Google Scholar] [CrossRef]
- Bojko, B.; Onat, B.; Boyaci, E.; Psillakis, E.; Dailianis, T.; Pawliszyn, J. Application of in situ Solid-Phase Microextraction on Mediterranean Sponges for Untargeted Exometabolome Screening and Environmental Monitoring. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Le Goff, G.; Martin, M.T.; Iorga, B.I.; Adelin, E.; Servy, C.; Cortial, S.; Ouazzani, J. Isolation and characterization of unusual hydrazides from Streptomyces sp. impact of the cultivation support and extraction procedure. J. Nat. Prod. 2013, 76, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, G.; Martin, M.T.; Servy, C.; Cortial, S.; Lopes, P.; Bialecki, A.; Smadja, J.; Ouazzani, J. Isolation and characterization of alpha,beta-unsaturated gamma-lactono-hydrazides from Streptomyces sp. J. Nat. Prod. 2012, 75, 915–919. [Google Scholar] [CrossRef]
- Le Goff, G.; Adelin, E.; Cortial, S.; Servy, C.; Ouazzani, J. Application of solid-phase extraction to agar-supported fermentation. Bioprocess. Biosyst. Eng. 2013, 36, 1285–1290. [Google Scholar] [CrossRef]
- Adelin, E.; Servy, C.; Martin, M.T.; Arcile, G.; Iorga, B.I.; Retailleau, P.; Bonfill, M.; Ouazzani, J. Bicyclic and tetracyclic diterpenes from a Trichoderma symbiont of Taxus baccata. Phytochemistry 2014, 97, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Dallery, J.F.; Le Goff, G.; Adelin, E.; Iorga, B.I.; Pigne, S.; O’Connell, R.J.; Ouazzani, J. Deleting a Chromatin Remodeling Gene Increases the Diversity of Secondary Metabolites Produced by Colletotrichum higginsianum. J. Nat. Prod. 2019, 82, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Adelin, E.; le Goff, G.; Retailleau, P.; Bonfill, M.; Ouazzani, J. Isolation of the antibiotic methyl (R,E)-3-(1-hydroxy-4-oxocyclopent-2-en-1-yl)-acrylate EA-2801 from Trichoderma atroviridae. J. Antibiot. 2017, 70, 1053–1056. [Google Scholar] [CrossRef]
- Le Goff, G.; Lopes, P.; Arcile, G.; Vlachou, P.; van Elslande, E.; Retailleau, P.; Gallard, J.F.; Weis, M.; Benayahu, Y.; Fokialakis, N.; et al. Impact of the Cultivation Technique on the Production of Secondary Metabolites by Chrysosporium lobatum TM-237-S5, Isolated from the Sponge Acanthella cavernosa. Mar. Drugs 2019, 17, 678. [Google Scholar] [CrossRef]
- Samy, M.N.; Le Goff, G.; Lopes, P.; Georgousaki, K.; Gumeni, S.; Almeida, C.; Gonzalez, I.; Genilloud, O.; Trougakos, I.; Fokialakis, N.; et al. Osmanicin, a Polyketide Alkaloid Isolated from Streptomyces osmaniensis CA-244599 Inhibits Elastase in Human Fibroblasts. Molecules 2019, 24, 2239. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Menendez, V.; Crespo, G.; Toro, C.; Martin, J.; de Pedro, N.; Tormo, J.R.; Genilloud, O. Extending the Metabolite Diversity of the Endophyte Dimorphosporicola tragani. Metabolites 2019, 9, 197. [Google Scholar] [CrossRef]
- Gonzalez-Menendez, V.; Asensio, F.; Moreno, C.; de Pedro, N.; Monteiro, M.C.; de la Cruz, M.; Vicente, F.; Bills, G.F.; Reyes, F.; Genilloud, O.; et al. Assessing the effects of adsorptive polymeric resin additions on fungal secondary metabolite chemical diversity. Mycology 2014, 5, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Hwang, S.; Kim, J.; Cho, S.; Palsson, B.; Cho, B.K. Mini review: Genome mining approaches for the identification of secondary metabolite biosynthetic gene clusters in Streptomyces. Comput. Struct. Biotechnol. J. 2020, 18, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Defarge, C.; Trichet, J.; Maurin, A.; Hucher, M. Kopara in Polynesian atolls: Early stages of formation of calcareous stromatolites. Sediment. Geol. 1994, 89, 9–23. [Google Scholar] [CrossRef]
- Richert, L.; Roland, L.; Annie, H.; Claude, P. Cyanobacterial populations that build ‘kopara’ microbial mats in Rangiroa, Tuamotu Archipelago, French Polynesia. Eur. J. Phycol. 2006, 41, 259–279. [Google Scholar] [CrossRef]
- Che, L.M.; Andréfouet, S.; Bothorel, V.; Guezennec, M.; Rougeaux, H.; Guezennec, J.; Deslandes, E.; Trichet, J.; Matheron, R.; Campion, T.L.; et al. Physical, chemical, and microbiological characteristics of microbial mats (KOPARA) in the South Pacific atolls of French Polynesia. Can. J. Microbiol. 2001, 47, 994–1012. [Google Scholar] [CrossRef] [PubMed]
- Simon-Colin, C.; Raguénès, G.; Crassous, P.; Moppert, X.; Guezennec, J. A novel mcl-PHA produced on coprah oil by Pseudomonas guezennei biovar. tikehau, isolated from a “kopara” mat of French Polynesia. Int. J. Biol. Macromol. 2008, 43, 176–181. [Google Scholar] [CrossRef]
- Guézennec, J.; Moppert, X.; Raguenes, G.; Richert, L.; Costa, B.; Simon-Colin, C. Microbial mats in French Polynesia and their biotechnological applications. Process. Biochem. 2011, 46, 16–22. [Google Scholar] [CrossRef]
- Fox Ramos, A.E.; Evanno, L.; Poupon, E.; Champy, P.; Beniddir, M.A. Natural products targeting strategies involving molecular networking: Different manners, one goal. Nat. Prod. Rep. 2019, 36, 960–980. [Google Scholar] [CrossRef]
- Rong, X.; Guo, Y.; Huang, Y. Proposal to reclassify the Streptomyces albidoflavus clade on the basis of multilocus sequence analysis and DNA-DNA hybridization, and taxonomic elucidation of Streptomyces griseus subsp. solvifaciens. Syst. Appl. Microbiol. 2009, 32, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Labeda, D.P.; Goodfellow, M.; Brown, R.; Ward, A.C.; Lanoot, B.; Vanncanneyt, M.; Swings, J.; Kim, S.B.; Liu, Z.; Chun, J.; et al. Phylogenetic study of the species within the family Streptomycetaceae. Antonie Van Leeuwenhoek 2012, 101, 73–104. [Google Scholar] [CrossRef]
- Lamichhane, S.; Sen, P.; Dickens, A.M.; Hyötyläinen, T.; Orešič, M. An Overview of Metabolomics Data Analysis: Current Tools and Future Perspectives. Compr. Anal. Chem. 2018, 82, 387–413. [Google Scholar] [CrossRef]
- Canada, P.; Pereira, A.; Nogueira, N.; Png-Gonzalez, L.; Andrade, C.; Xavier, R. Analysis of bacterial microbiome associated with nylon and copper nets in an aquaculture context. Aquaculture 2020, 516, 734540. [Google Scholar] [CrossRef]
- Zhong, H.; Wang, H.; Tian, Y.; Liu, X.; Yang, Y.; Zhu, L.; Yan, S.; Liu, G. Treatment of polluted surface water with nylon silk carrier-aerated biofilm reactor (CABR). Bioresour. Technol. 2019, 289, 121617. [Google Scholar] [CrossRef] [PubMed]
- Venable, M.E.; Podbielski, M.R. Impact of substrate material on algal biofilm biomass growth. Environ. Sci. Pollut. Res. 2019, 26, 7256–7262. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.F.; Petras, D.; Schmid, R.; Duhrkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-Based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Almeida, E.L.; Kaur, N.; Jennings, L.K.; Carrillo Rincon, A.F.; Jackson, S.A.; Thomas, O.P.; Dobson, A.D.W. Genome Mining Coupled with OSMAC-Based Cultivation Reveal Differential Production of Surugamide A by the Marine Sponge Isolate Streptomyces sp. SM17 When Compared to Its Terrestrial Relative S. albidoflavus J1074. Microorganisms 2019, 7, 394. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, X.; Kim, S.J.; Zhang, W. Antimycin-type depsipeptides: Discovery, biosynthesis, chemical synthesis, and bioactivities. Nat. Prod. Rep. 2016, 33, 1146–1165. [Google Scholar] [CrossRef]
- Li, H.; Huang, H.; Hou, L.; Ju, J.; Li, W. Discovery of Antimycin-Type Depsipeptides from a wbl Gene Mutant Strain of Deepsea-Derived Streptomyces somaliensis SCSIO ZH66 and Their Effects on Pro-inflammatory Cytokine Production. Front. Microbiol. 2017, 8, 678. [Google Scholar] [CrossRef]
- Tsuchida, T.; Iinuma, H.; Nishida, C.; Kinoshita, N.; Sawa, T.; Hamada, M.; Takeuchi, T. Tetrodecamycin and dihydrotetrodecamycin, new antimicrobial antibiotics against Pasteurella piscicida produced by Streptomyces nashvillensis MJ885-mF8. I. Taxonomy, fermentation, isolation, characterization and biological activities. J. Antibiot. 1995, 48, 1104–1109. [Google Scholar] [CrossRef][Green Version]
- Richardson, M.B.; Williams, S.J. A practical synthesis of long-chain iso-fatty acids (iso-C12-C19) and related natural products. Beilstein J. Org. Chem. 2013, 9, 1807–1812. [Google Scholar] [CrossRef]
- Barrow, C.J.; Oleynek, J.J.; Marinelli, V.; Sun, H.H.; Kaplita, P.; Sedlock, D.M.; Gillum, A.M.; Chadwick, C.C.; Cooper, R. Antimycins, inhibitors of ATP-citrate lyase, from a Streptomyces sp. J. Antibiot. 1997, 50, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Inai, M.; Nishii, T.; Tanaka, A.; Kaku, H.; Horikawa, M.; Tsunoda, T. Total Synthesis of the (+)-Antimycin A Family. Eur. J. Org. Chem. 2011, 2011, 2719–2729. [Google Scholar] [CrossRef]
- Mai, P.Y.; Levasseur, M.; Buisson, D.; Touboul, D.; Eparvier, V. Identification of Antimicrobial Compounds from Sandwithia guyanensis-Associated Endophyte Using Molecular Network Approach. Plants 2019, 9, 47. [Google Scholar] [CrossRef]
- Flardh, K.; Buttner, M.J. Streptomyces morphogenetics: Dissecting differentiation in a filamentous bacterium. Nat. Rev. Microbiol. 2009, 7, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Cary, J.W. Association of fungal secondary metabolism and sclerotial biology. Front. Microbiol. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Rahman, T.M.A.; Tharwat, N.A.; Abo El-Souad, S.M.S.; El-Beih, A.A.; El-Diwany, A.I. Biological activities and variation of symbiotic fungi isolated from Coral reefs collected from Red Sea in Egypt. Mycology 2020, 11, 243–255. [Google Scholar] [CrossRef]
- Calabon, M.S.; Sadaba, R.B.; Campos, W.L. Fungal diversity of mangrove-associated sponges from New Washington, Aklan, Philippines. Mycology 2019, 10, 6–21. [Google Scholar] [CrossRef]
- Brescia, F.; Marchetti-Deschmann, M.; Musetti, R.; Perazzolli, M.; Pertot, I.; Puopolo, G. The rhizosphere signature on the cell motility, biofilm formation and secondary metabolite production of a plant-associated Lysobacter strain. Microbiol. Res. 2020, 234, 126424. [Google Scholar] [CrossRef]
- Rieusset, L.; Rey, M.; Muller, D.; Vacheron, J.; Gerin, F.; Dubost, A.; Comte, G.; Prigent-Combaret, C. Secondary metabolites from plant-associated Pseudomonas are overproduced in biofilm. Microb. Biotechnol. 2020, 13, 1562–1580. [Google Scholar] [CrossRef]
- Timmermans, M.L.; Picott, K.J.; Ucciferri, L.; Ross, A.C. Culturing marine bacteria from the genus Pseudoalteromonas on a cotton scaffold alters secondary metabolite production. MicrobiologyOpen 2019, 8, e00724. [Google Scholar] [CrossRef]
- Letsiou, S.; Bakea, A.; Le Goff, G.; Lopes, P.; Gardikis, K.; Alonso, C.; Alvarez, P.A.; Ouazzani, J. In vitro protective effects of marine-derived Aspergillus puulaauensis TM124-S4 extract on H2O2-stressed primary human fibroblasts. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2020, 66, 104869. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- Myers, O.D.; Sumner, S.J.; Li, S.; Barnes, S.; Du, X. One Step Forward for Reducing False Positive and False Negative Compound Identifications from Mass Spectrometry Metabolomics Data: New Algorithms for Constructing Extracted Ion Chromatograms and Detecting Chromatographic Peaks. Anal. Chem. 2017, 89, 8696–8703. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mai, P.-Y.; Le Goff, G.; Poupon, E.; Lopes, P.; Moppert, X.; Costa, B.; Beniddir, M.A.; Ouazzani, J. Solid-Phase Extraction Embedded Dialysis (SPEED), an Innovative Procedure for the Investigation of Microbial Specialized Metabolites. Mar. Drugs 2021, 19, 371. https://doi.org/10.3390/md19070371
Mai P-Y, Le Goff G, Poupon E, Lopes P, Moppert X, Costa B, Beniddir MA, Ouazzani J. Solid-Phase Extraction Embedded Dialysis (SPEED), an Innovative Procedure for the Investigation of Microbial Specialized Metabolites. Marine Drugs. 2021; 19(7):371. https://doi.org/10.3390/md19070371
Chicago/Turabian StyleMai, Phuong-Y., Géraldine Le Goff, Erwan Poupon, Philippe Lopes, Xavier Moppert, Bernard Costa, Mehdi A. Beniddir, and Jamal Ouazzani. 2021. "Solid-Phase Extraction Embedded Dialysis (SPEED), an Innovative Procedure for the Investigation of Microbial Specialized Metabolites" Marine Drugs 19, no. 7: 371. https://doi.org/10.3390/md19070371
APA StyleMai, P.-Y., Le Goff, G., Poupon, E., Lopes, P., Moppert, X., Costa, B., Beniddir, M. A., & Ouazzani, J. (2021). Solid-Phase Extraction Embedded Dialysis (SPEED), an Innovative Procedure for the Investigation of Microbial Specialized Metabolites. Marine Drugs, 19(7), 371. https://doi.org/10.3390/md19070371