A Conantokin Peptide Con-T[M8Q] Inhibits Morphine Dependence with High Potency and Low Side Effects


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
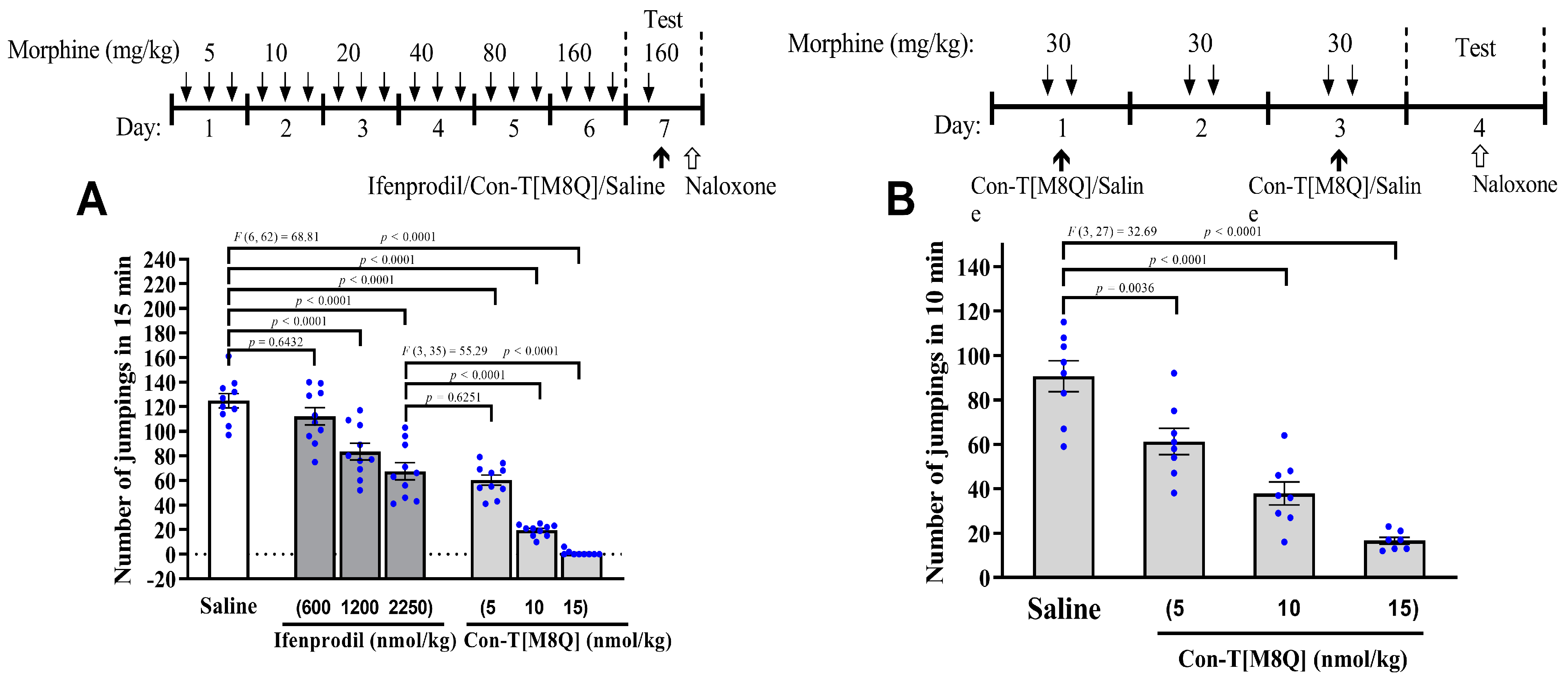
2.1. Con-T[M8Q] Potently Inhibits the Expression and Development of Morphine Physical Dependence in Mice
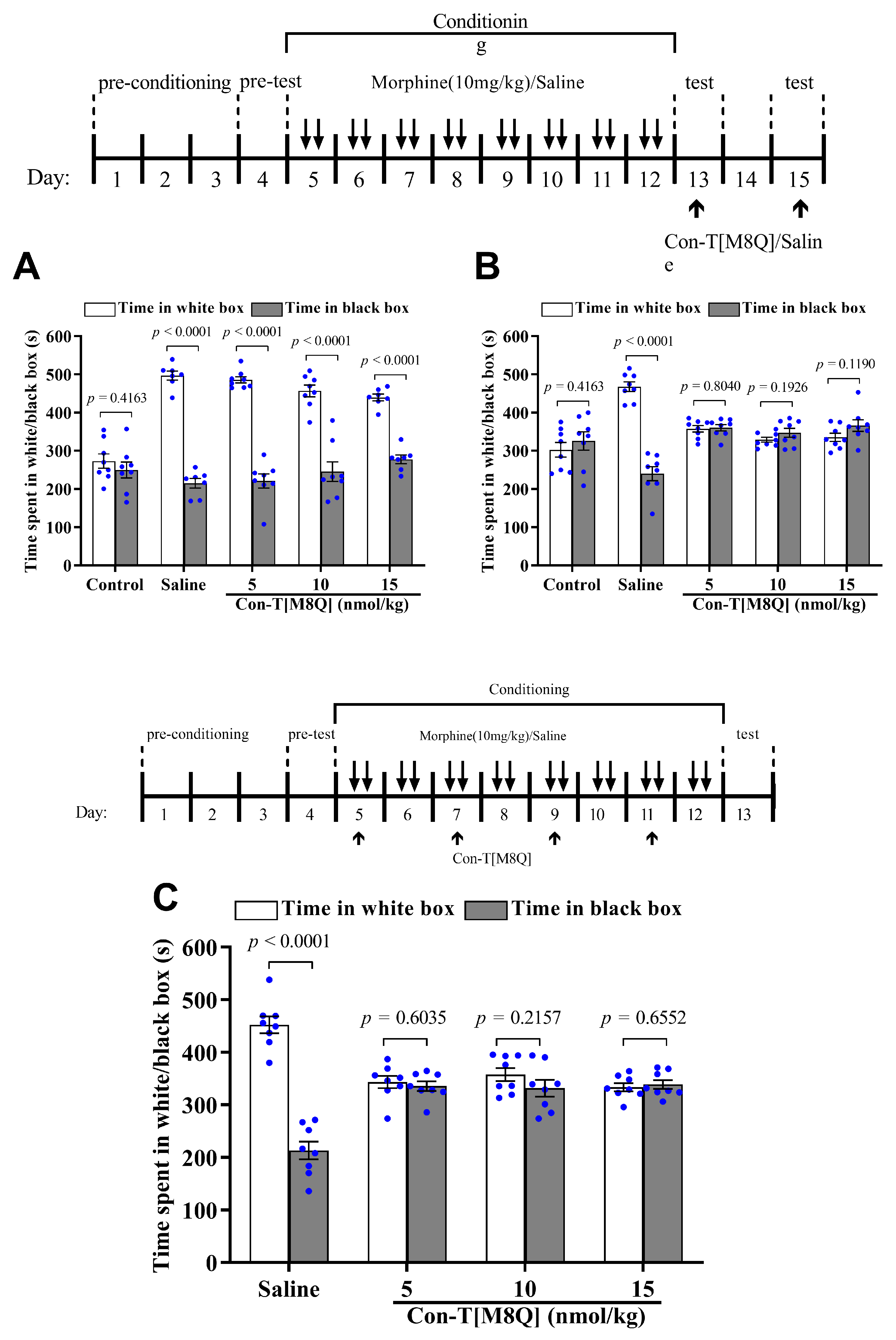
2.2. Con-T[M8Q] Potently Inhibits the Expression and Development of Morphine-Induced CPP in Mice
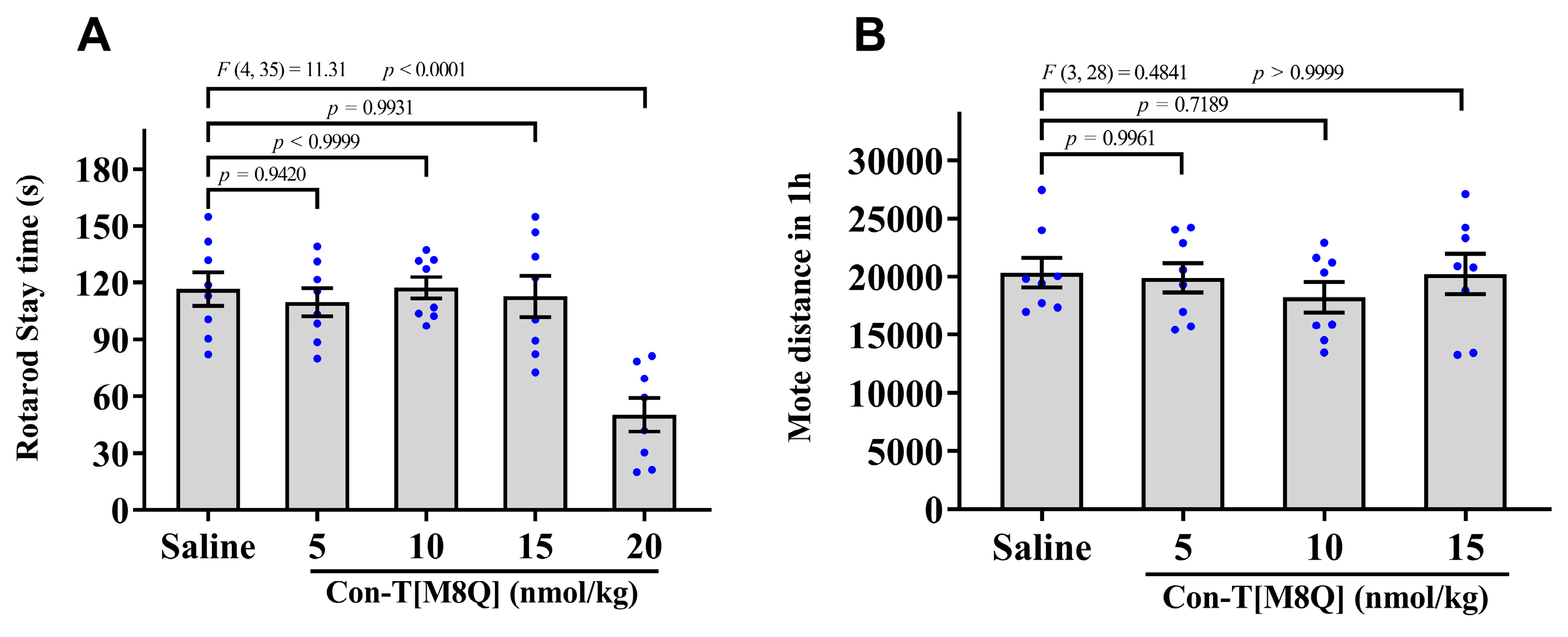
2.3. Con-T[M8Q] Has Little Effects on the Coordinated Locomotion and Spontaneous Locomotor Activity in Mice
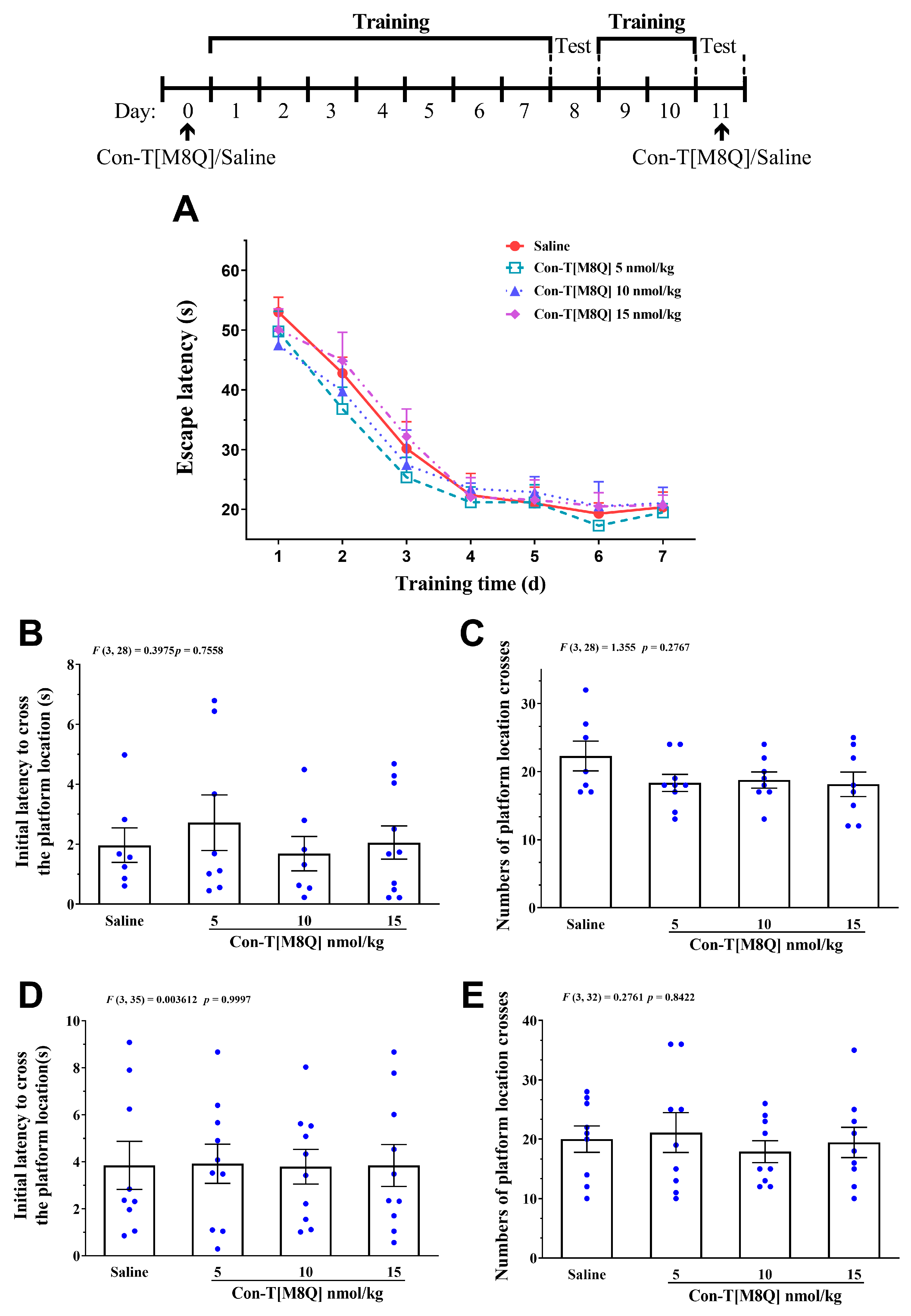
2.4. Con-T[M8Q] Has No Apparent Effects on the Spatial learning Memory and Menory Recall in Mice
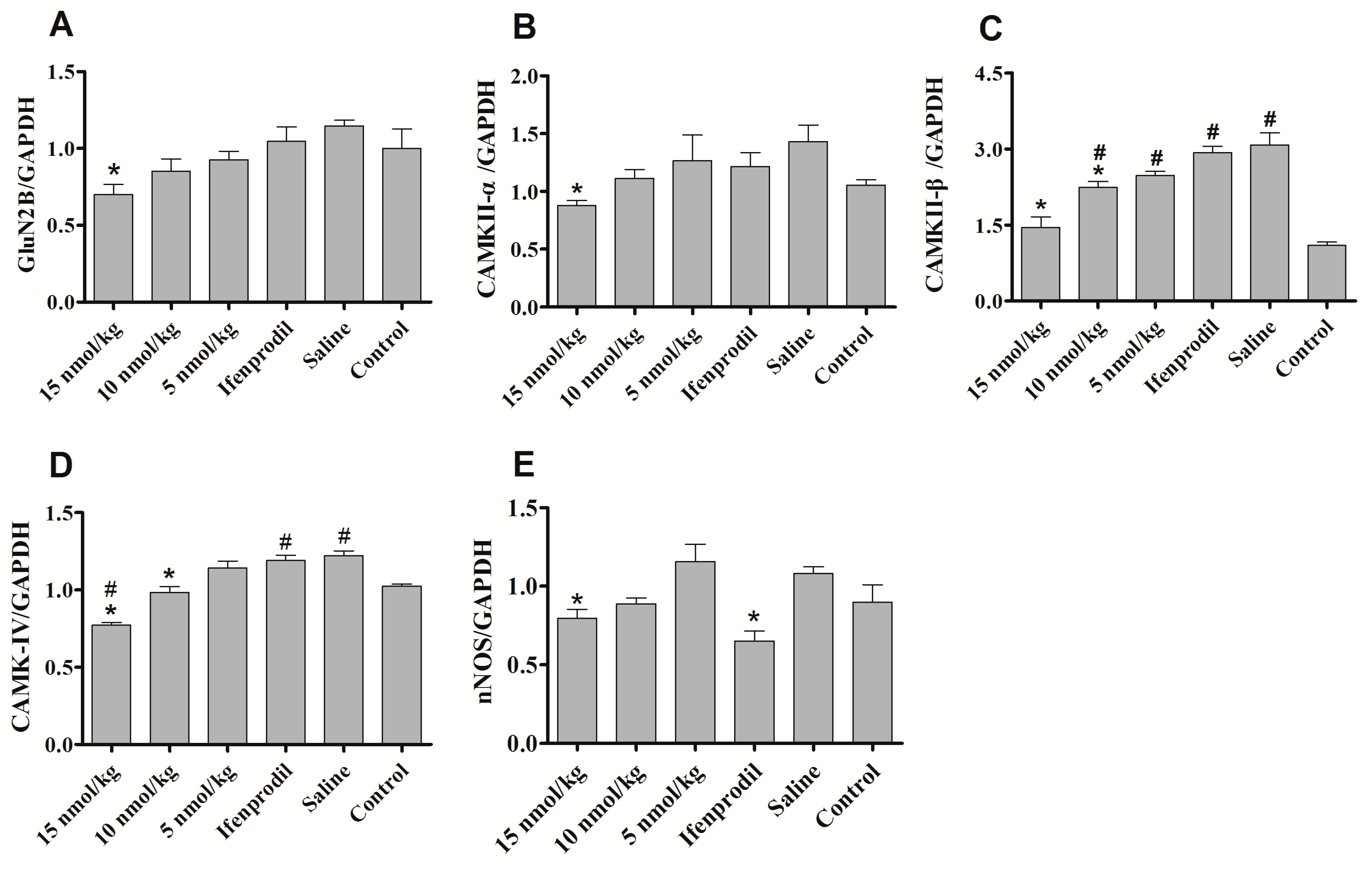
2.5. Con-T[M8Q] Decreases the Levels of GluN2B, CaMKs, nNOS and c-fos mRNA in Morphine-Dependent Mice
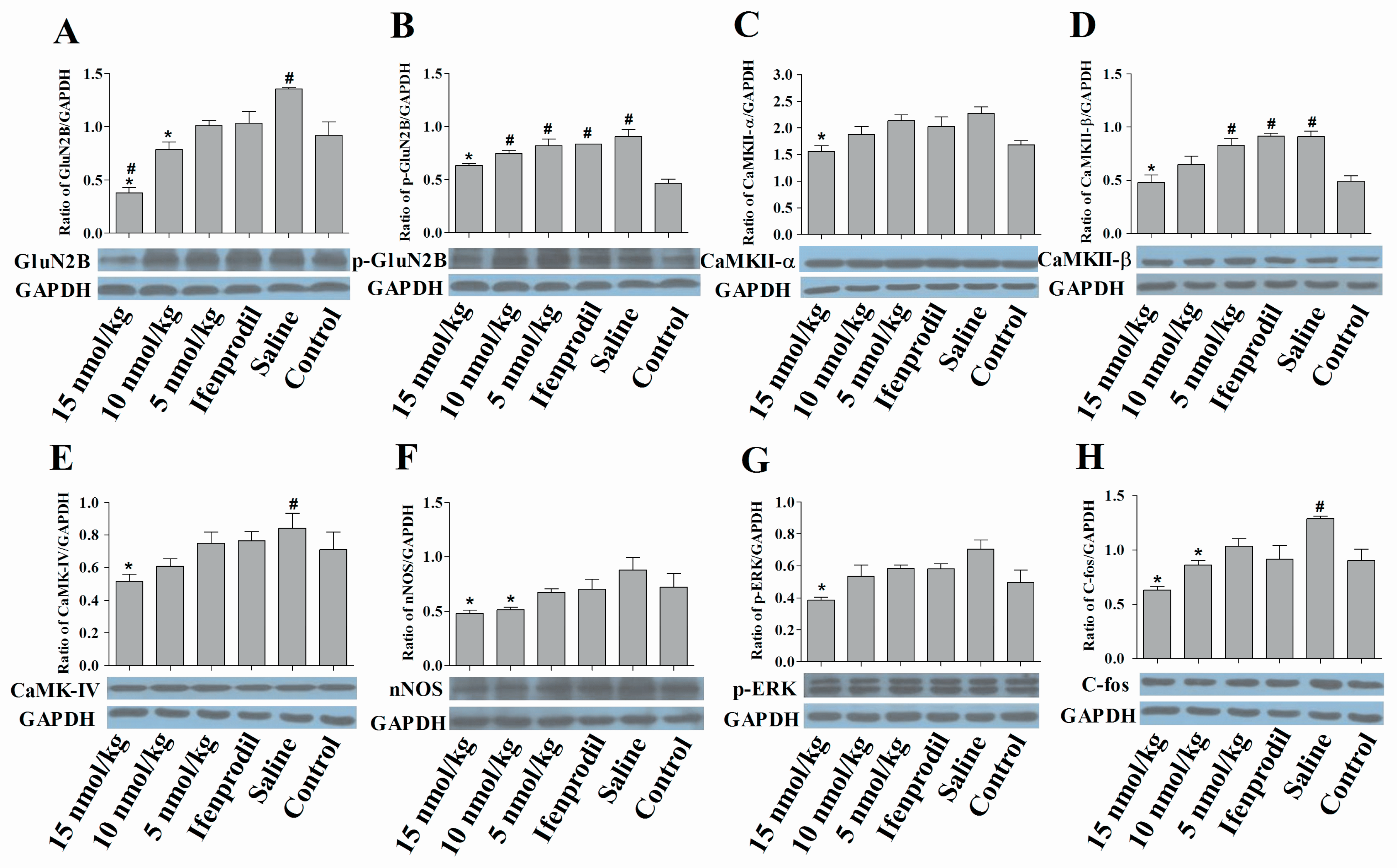
2.6. Con-T[M8Q] Efficiently Decreases the Expressions of GluN2B, p-GluN2B, CaMKs, nNOS, pERK and c-fos Proteins in Morphine-Dependent Mice
3. Discussion
4. Materials and Methods
4.1. Animals and Drugs
4.2. Morphine Physical Dependent Abstinence/Withdrawal
4.3. Morphine Psychological Dependent Abstinence
4.4. Coordinated Locomotion in Rotarod Test
4.5. Spontaneous Locomotor Activity
4.6. Morris Water Maze Test
4.7. mRNA Extraction and Quantitative RT-PCR
4.8. Western Blot Analyses
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furukawa, H.; Singh, S.K.; Mancusso, R.; Gouaux, E. Subunit arrangement and function in NMDA receptors. Nature 2005, 438, 185–192. [Google Scholar] [CrossRef]
- Paoletti, P. Molecular basis of NMDA receptor functional diversity. Eur. J. Neurosci. 2011, 33, 1351–1365. [Google Scholar] [CrossRef]
- Noda, Y.; Nabeshima, T. Opiate physical dependence and N-methyl-d-aspartate receptors. Eur. J. Pharmacol. 2004, 500, 121–128. [Google Scholar] [CrossRef]
- Christie, M.J. Cellular neuroadaptations to chronic opioids: Tolerance, withdrawal and addiction. Br. J. Pharmacol. 2008, 154, 384–396. [Google Scholar] [CrossRef]
- Mony, L.; Kew, J.N.C.; Gunthorpe, M.J.; Paoletti, P. Allosteric modulators of NR2B-containing NMDA receptors: Molecular mechanisms and therapeutic potential. Br. J. Pharmacol. 2009, 157, 1301–1317. [Google Scholar] [CrossRef]
- Tan, H.; Rosen, L.G.; Ng, G.A.; Rushlow, W.J.; LaViolette, S.R.; Addiction Research Group. NMDA Receptor Blockade in the Prelimbic Cortex Activates the Mesolimbic System and Dopamine-Dependent Opiate Reward Signaling. Psychopharmacology 2014, 231, 4669–4679. [Google Scholar] [CrossRef]
- Narita, M.; Aoki, T.; Suzuki, T. Molecular evidence for the involvement of NR2B subunit containing N-methyl-d-aspartate receptors in the development of morphine-induced place preference. Neuroscience 2000, 101, 601–606. [Google Scholar] [CrossRef]
- Narita, M.; Kato, H.; Miyoshi, K.; Aoki, T.; Yajima, Y.; Suzuki, T. Treatment for psychological dependence on morphine: Use-fulness of inhibiting NMDA receptor and its associated protein kinase in the nucleus accumbens. Life Sci. 2005, 77, 2207–2220. [Google Scholar] [CrossRef]
- Garzón-Niño, J.; Rodríguez-Muñoz, M.; Sánchez-Blázquez, P. Direct association of Mu-opioid and NMDA glutamate receptors supports their cross-regulation: Molecular implications for opioid tolerance. Curr. Drug Abuse Rev. 2012, 5, 199–226. [Google Scholar] [CrossRef]
- Suzuki, T.; Kato, H.; Tsuda, M.; Suzuki, H.; Misawa, M. Effects of the non-competitive NMDA receptor antagonist ifenprodil on the morphine-induced place preference in mice. Life Sci. 1999, 64, 151–156. [Google Scholar] [CrossRef]
- Ma, Y.Y.; Yu, P.; Guo, C.Y.; Cui, C.L. Effects of Ifenprodil on Morphine-Induced Conditioned Place Preference and Spatial Learning and Memory in Rats. Neurochem. Res. 2011, 36, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Bespalov, A.Y.; Balster, R.L.; Beardsley, P.M. N-Methyl-D-aspartate receptor antagonists and the development of tolerance to the discriminative stimulus effects of morphine in rats. J. Pharmacol. Exp. Ther. 1999, 290, 20–27. [Google Scholar] [PubMed]
- Ko, S.W.; Wu, L.J.; Shum, F.; Quan, J.; Zhuo, M. Cingulate NMDA NR2B receptors contribute to morphine-induced analgesic tolerance. Mol. Brain 2008, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Chizh, B.A.; Headley, P.M.; Tzschentke, T.M. NMDA receptor antagonists as analgesics: Focus on the NR2B subtype. Trends Pharmacol. Sci. 2001, 22, 636–642. [Google Scholar] [CrossRef]
- Prorok, M.; Castellino, F.J. The molecular basis of conantokin antagonism of NMDA receptor function. Curr. Drug Targets 2007, 8, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Dutt, M.; Dutertre, S.; Jin, A.H.; Lavergne, V.; Alewood, P.F.; Lewis, R.J. Venomics Reveals Venom Complexity of the Piscivorous Cone Snail, Conus tulipa. Mar. Drugs 2019, 17, 71. [Google Scholar] [CrossRef]
- Kunda, S.; Yuan, Y.; Balsara, R.D.; Zajicek, J.; Castellino, F.J. Hydroxyproline-induced Helical Disruption in Conantokin Rl-B Affects Subunit-selective Antagonistic Activities toward Ion Channels of N-Methyl-d-aspartate Receptors. J. Biol. Chem. 2015, 290, 18156–18172. [Google Scholar] [CrossRef]
- Wei, J.; Dong, M.; Xiao, C.; Jiang, F.; Castellino, F.J.; Prorok, M.; Dai, Q. Conantokins and variants derived from cone snail venom inhibit naloxone-induced withdrawal jumping in morphine-dependent mice. Neurosci. Lett. 2006, 405, 137–141. [Google Scholar] [CrossRef]
- Xiao, C.; Huang, Y.; Dong, M.; Hu, J.; Hou, S.; Castellino, F.J.; Prorok, M.; Dai, Q. NR2B-selective conantokin peptide inhibitors of the NMDA receptor display enhanced antinociceptive properties compared to non-selective conantokins. Neuropeptides 2008, 42, 601–609. [Google Scholar] [CrossRef]
- Sheng, Z.; Dai, Q.; Prorok, M.; Castellino, F.J. Subtype-selective antagonism of N-methyl-D-aspartate receptor ion channels by synthetic conantokin peptides. Neuropharmacology 2007, 53, 145–156. [Google Scholar] [CrossRef]
- Ren, B.; Zhou, Z.; Liu, Z.; Li, B.; Ou, J.; Dai, Q. Con-T[M8Q] potently attenuates the expression and development of morphine tolerance in mice. Neurosci. Lett. 2015, 597, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Takahashi, E.; Li, W.; Halt, A.; Wiltgen, B.; Ehninger, D.; Li, G.D.; Hell, J.W.; Kennedy, M.B.; Silva, A.J. Interactions between the NR2B Receptor and CaMKII Modulate Synaptic Plasticity and Spatial Learning. J. Neurosci. 2007, 27, 13843–13853. [Google Scholar] [CrossRef] [PubMed]
- Portugal, G.S.; Al-Hasani, R.; Fakira, A.K.; Gonzalez-Romero, J.L.; Melyan, Z.; McCall, J.G.; Bruchas, M.R.; Morón, J.A. Hippocampal long-term potentiation is disrupted during expression and extinction but is restored after reinstatement of morphine place preference. J. Neurosci. 2014, 34, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Xiang, X.H.; Guo, Y.; Wu, W.R.; Cao, D.Y.; Wang, H.S.; Zhao, Y. Ionotropic glutamatergic neurotransmission in the ventral tegmental area modulates DeltaFosB expression in the nucleus accumbens and abstinence syndrome in morphine withdrawal rats. Eur. J. Pharmacol. 2005, 527, 94–104. [Google Scholar] [CrossRef]
- Harrison, N.J.; Grimwood, S.; Bristow, L.J.; Hutson, P.H. Nucleus accumbens NMDA receptor subunit expression and function is enhanced in morphine-dependent rats. Eur. J. Pharmacol. 2007, 562, 191–197. [Google Scholar] [CrossRef]
- Bossert, J.M.; Adhikary, S.; Laurent, R.S.; Marchant, N.J.; Wang, H.L.; Morales, M.; Shaham, Y. Role of projections from ventral subiculum to nucleus accumbens shell in context-induced reinstatement of heroin seeking in rats. Psychopharmacology 2016, 233, 1991–2004. [Google Scholar] [CrossRef]
- Anderson, E.M.; Reeves, T.; Kapernaros, K.; Neubert, J.K.; Caudle, R.M. Phosphorylation of the N-methyl-d-aspartate receptor is increased in the nucleus accumbens during both acute and extended morphine withdrawal. J. Pharmacol. Exp. Ther. 2015, 355, 496–505. [Google Scholar] [CrossRef]
- Abe, T.; Matsumura, S.; Katano, T.; Mabuchi, T.; Takagi, K.; Xu, L.; Yamamoto, A.; Hattori, K.; Yagi, T.; Watanabe, M.; et al. Fyn kinase-mediated phosphorylation of NMDA receptor NR2B subunit at Tyr1472 is essential for maintenance of neuropathic pain. Eur. J. Neurosci. 2005, 22, 1445–1454. [Google Scholar] [CrossRef]
- Matsumura, S.; Kunori, S.; Mabuchi, T.; Katano, T.; Nakazawa, T.; Abe, T.; Watanabe, M.; Yamamoto, T.; Okuda-Ashitaka, E.; Ito, S. Impairment of CaMKII activation and attenuation of neuropathic pain in mice lacking NR2B phosphorylated at Tyr1472. Eur. J. Neurosci. 2010, 32, 798–810. [Google Scholar] [CrossRef]
- Katano, T.; Nakazawa, T.; Nakatsuka, T.; Watanabe, M.; Yamamoto, T.; Ito, S. Involvement of spinal phosphorylation cascade of Tyr1472-NR2B, Thr286-CaMKII, and Ser831-GluR1 in neuropathic pain. Neuropharmacology 2011, 60, 609–616. [Google Scholar] [CrossRef]
- Yu, Z.; Huang, Y.Y.; Liu, Z.G.; Dai, Q.Y. Inhibitory effect of conantokins and their variants on morphine dependent devel-op-ment in mice. Lett. Biotechnol. 2010, 21, 551–554. [Google Scholar]
- Wei, X.L.; Su, R.B.; Lu, X.Q.; Liu, Y.; Yu, S.Z.; Yuan, B.L.; Li, J. Inhibition by agmatine on morphine-induced conditioned place preference in rats. Eur. J. Pharmacol. 2005, 515, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Gao, Y.; Yang, L.; Kong, X.; Yu, J.; Hou, W.; Hua, B. The mechanism of μ-opioid receptor (MOR)-TRPV1 crosstalk in TRPV1 activation involves morphine anti-nociception, tolerance and dependence. Channels (Austin) 2015, 9, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Reflections on: “A general role for adaptations in G-Proteins and the cyclic AMP system in mediating the chronic actions of morphine and cocaine on neuronal function”. Brain Res. 2016, 1645, 71–74. [Google Scholar] [CrossRef]
- Wright, V.L.; Georgiou, P.; Bailey, A.; Heal, D.J.; Bailey, C.P.; Wonnacott, S. Inhibition of alpha7 nicotinic receptors in the ventral hippocampus selectively attenuates reinstatement of morphine-conditioned place preference and associated changes in AMPA receptor binding. Addict. Biol. 2019, 24, 590–603. [Google Scholar] [CrossRef]
- Elliott, K.; Minami, N.; Kolesnikov, Y.A.; Pasternak, G.W.; Inturrisi, C.E. The NMDA Receptor antagonists, LY274614 and MK-801, and the nitric oxide synthase inhibitor, NG-nitro-L-arginine, attenuate analgesic tolerance to the mu-opioid morphine but not to kappa opioids. Pain 1994, 56, 69–75. [Google Scholar] [CrossRef]
- Fan, G.H.; Wang, L.Z.; Qiu, H.C.; Ma, L.; Pei, G. Inhibition of calcium/calmodulin-dependent protein kinase II in rat hippocampus attenuates morphine tolerance and dependence. Mol. Pharmacol. 1999, 56, 39–45. [Google Scholar] [CrossRef]
- Park, C.S.; Elgersma, Y.; Grant, S.G.N.; Morrison, J.H. α-Isoform of calcium-calmodulin-dependent protein kinase II and postsynaptic density protein 95 differentially regulate synaptic expression of NR2A– and NR2B–containing N-methyl-d-aspartate receptors in hippocampus. Neuroscience 2008, 151, 43–55. [Google Scholar] [CrossRef][Green Version]
- Almela, P.; Milanés, M.V.; Laorden, M.L. Activation of the ERK signalling pathway contributes to the adaptive changes in rat hearts during naloxone-induced morphine withdrawal. Br. J. Pharmacol. 2007, 151, 787–797. [Google Scholar] [CrossRef]
- McPhie, A.A.; Barr, G.A. Regional Fos expression induced by morphine withdrawal in the 7-day-old rat. Dev. Psychobiol. 2009, 51, 544–552. [Google Scholar] [CrossRef]
- Barria, A.; Malinow, R. NMDA Receptor Subunit Composition Controls Synaptic Plasticity by Regulating Binding to CaMKII. Neuron 2005, 48, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Marie, H.; Morishita, W.; Yu, X.; Calakos, N.; Malenka, R.C. Generation of Silent Synapses by Acute In Vivo Expression of CaMKIV and CREB. Neuron 2005, 45, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.W.; Jia, Y.; Xu, H.; Yim, S.J.; Jang, D.H.; Lee, Y.S.; Zhao, M.G.; Toyoda, H.; Wu, L.J.; Chatila, T.; et al. Evidence for a role of CaMKIV in the development of opioid analgesic tolerance. Eur. J. Neurosci. 2006, 23, 2158–2168. [Google Scholar] [CrossRef] [PubMed]
- Williams, K. Ifenprodil, a Novel NMDA Receptor Antagonist: Site and Mechanism of Action. Curr. Drug Targets 2001, 2, 285–298. [Google Scholar] [CrossRef]
- Gitto, R.; De Luca, L.; Ferro, S.; Occhiuto, F.; Samperi, S.; De Sarro, G.; Russo, E.; Ciranna, L.; Costa, L.; Chimirri, A. Computational studies to discover a new NR2B/NMDA receptor antagonist and evaluation of pharmacological profile. ChemMedChem 2008, 3, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Tomitori, H.; Suganami, A.; Saiki, R.; Mizuno, S.; Yoshizawa, Y.; Masuko, T.; Tamura, Y.; Nishimura, K.; Toida, T.; Williams, K.; et al. Structural changes of regulatory domain heterodimer of N-methyl-D-aspartate receptor subunits GluN1 and GluN2B through the binding of spermine and ifenprodil. J. Pharmacol. Exp. Ther. 2012, 343, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Skolnick, P.; Boje, K.; Miller, R.; Pennington, M.; Maccecchini, M.L. Noncompetitive inhibition of N-methyl-D-aspartate by conantokin-G: Evidence for an allosteric interactin at polyamins sites. J. Neurochem. 1992, 59, 1516–1521. [Google Scholar] [CrossRef]
- Sheng, Z.; Liang, Z.; Geiger, J.H.; Prorok, M.; Castellino, F.J. The selectivity of conantokin-G for ion channel inhibition of NR2B subunit-containing NMDA receptors is regulated by amino acid residues in the S2 region of NR2B. Neuropharmacology 2009, 57, 127–136. [Google Scholar] [CrossRef]
- Hackos, D.H.; Hanson, J.E. Diverse modes of NMDA receptor positive allosteric modulation: Mechanisms and consequences. Neuropharmacology 2017, 112, 34–45. [Google Scholar] [CrossRef]
- Warder, S.E.; Blandl, T.; Klein, R.C.; Castellino, F.J.; Prorok, M. Amino acid determinants for NMDA receptor inhibition by conantokin-T. J. Neurochem. 2001, 77, 812–822. [Google Scholar] [CrossRef]
- Tsai, V.W.; Dodd, P.R.; Lewis, R.J. The effects of alanine-substituted conantokin-G and ifenprodil on the human spermine-activated N-methyl-D-aspartate receptor. Neuroscience 2005, 130, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Sheng, Z.; Geiger, J.H.; Castellino, F.J.; Prorok, M. Helix-Helix Interactions between Homo- and Heterodimeric γ-Carboxyglutamate-containing Conantokin Peptides and Their Derivatives. J. Biol. Chem. 2007, 282, 12641–12649. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liang, L.; Ning, H.; Cai, F.; Liu, Z.; Zhang, L.; Zhou, L.; Dai, Q. Cloning, Synthesis and Functional Characterization of a Novel α-Conotoxin Lt1.3. Mar. Drugs 2018, 16, 112. [Google Scholar] [CrossRef]
- Popik, P.; Skolnick, P. The NMDA antagonist memantine blocks the expression and maintenance of morphine dependence. Pharmacol. Biochem. Behav. 1996, 53, 791–797. [Google Scholar] [CrossRef]
- Popik, P.; Mamczarz, J.; Fraczek, M.; Widła, M.; Hesselink, M.; Danysz, W. Inhibition of reinforcing effects of morphine and naloxone-precipitated opioid withdrawal by novel glycine site and uncompetitive NMDA receptor antagonists. Neuropharmacology 1998, 37, 1033–1042. [Google Scholar] [CrossRef]
- Xu, S.Y.; Bian, R.L.; Chen, X. Experimental Methodology of Pharmacology, 3rd ed.; People’s Medical Publishing House: Beijing, China, 2001; 252p. [Google Scholar]
- Malmberg, A.B.; Gilbert, H.; McCabe, R.T.; Basbaum, A.I. Powerful antinociceptive effects of the cone snail venom-derived subtype-selective NMDA receptor antagonists conantokins G and T. Pain 2003, 101, 109–116. [Google Scholar] [CrossRef]
- Yu, Y.Z.; Liu, S.; Wang, H.C.; Shi, D.Y.; Xu, Q.; Zhou, X.W.; Sun, Z.W.; Huang, P.T. A novel recombinant 6Aβ15-THc-C chimeric vaccine (rCV02) mitigates Alzheimer’s disease-like pathology, cognitive decline and synaptic loss in aged 3 × Tg-AD mice. Sci. Rep. 2016, 6, 27175. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Yu, Z.; Yu, S.; Zhu, C.; Dong, M.; Mao, W.; Hu, J.; Prorok, M.; Su, R.; Dai, Q. A Conantokin Peptide Con-T[M8Q] Inhibits Morphine Dependence with High Potency and Low Side Effects. Mar. Drugs 2021, 19, 44. https://doi.org/10.3390/md19010044
Liu Z, Yu Z, Yu S, Zhu C, Dong M, Mao W, Hu J, Prorok M, Su R, Dai Q. A Conantokin Peptide Con-T[M8Q] Inhibits Morphine Dependence with High Potency and Low Side Effects. Marine Drugs. 2021; 19(1):44. https://doi.org/10.3390/md19010044
Chicago/Turabian StyleLiu, Zhuguo, Zheng Yu, Shuo Yu, Cui Zhu, Mingxin Dong, Wenxiang Mao, Jie Hu, Mary Prorok, Ruibin Su, and Qiuyun Dai. 2021. "A Conantokin Peptide Con-T[M8Q] Inhibits Morphine Dependence with High Potency and Low Side Effects" Marine Drugs 19, no. 1: 44. https://doi.org/10.3390/md19010044
APA StyleLiu, Z., Yu, Z., Yu, S., Zhu, C., Dong, M., Mao, W., Hu, J., Prorok, M., Su, R., & Dai, Q. (2021). A Conantokin Peptide Con-T[M8Q] Inhibits Morphine Dependence with High Potency and Low Side Effects. Marine Drugs, 19(1), 44. https://doi.org/10.3390/md19010044