Establishment of Novel High-Standard Chemiluminescent Assay for NTPase in Two Protozoans and Its High-Throughput Screening

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Amino Acid Sequences of TgNTPase and NcNTPase and Structural Information of Their Active Mutants
2.2. Establishment of Novel High-Standard Assay to Determine NTPase Activity by Combination of Classical Enzymatic Assay and Fluorescent Assay to Measure ADP Content
2.3. HTS Using a Synthetic Compound Library, Which is Provided by the University of Tokyo, in Order to Identify Compounds Inhibiting NcNTPase
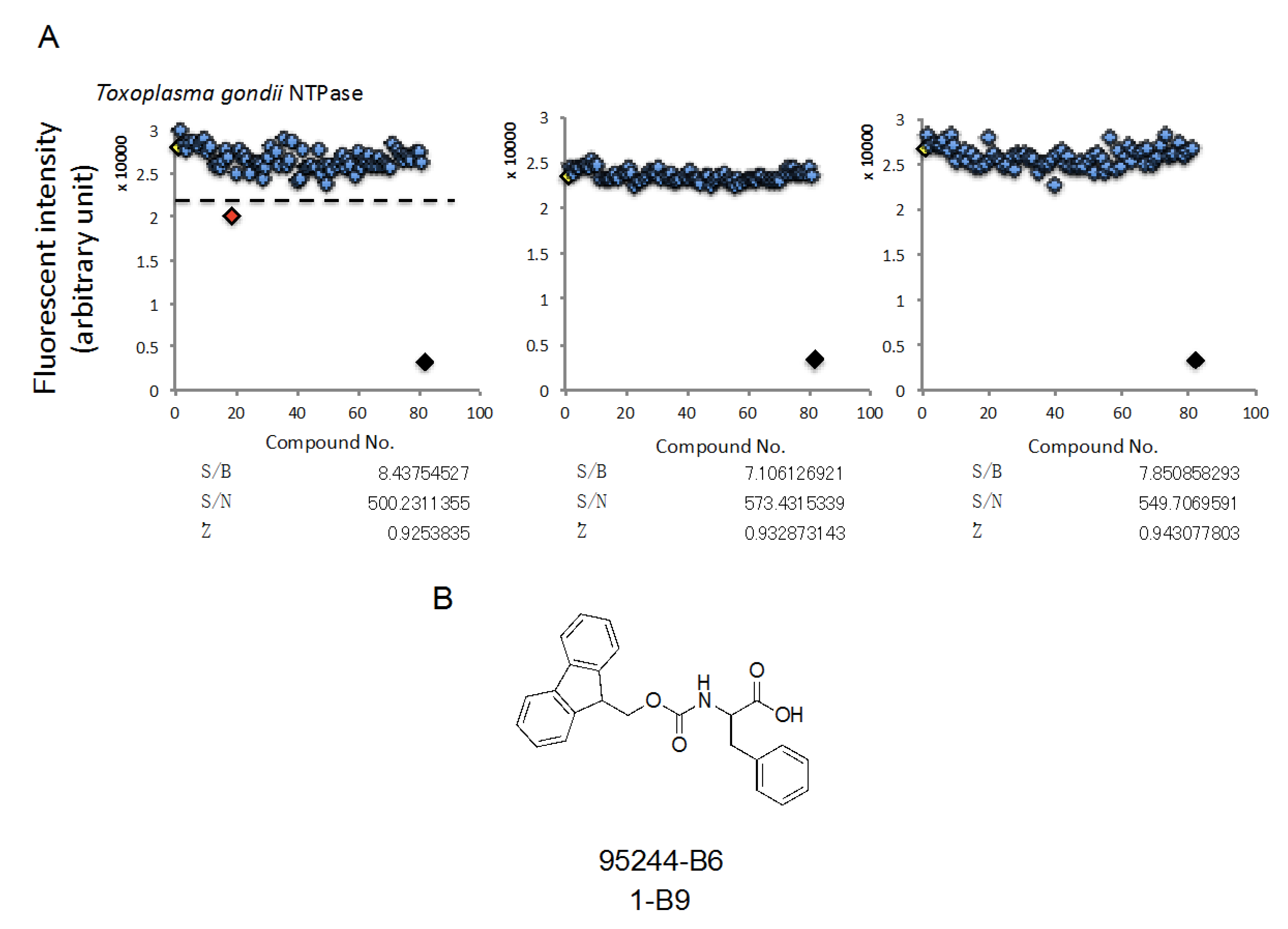
2.4. HTS Using a Synthetic Compound Library, Which is Provided by Nagasaki University, in Order to Identify Compounds Inhibiting TgNTPase
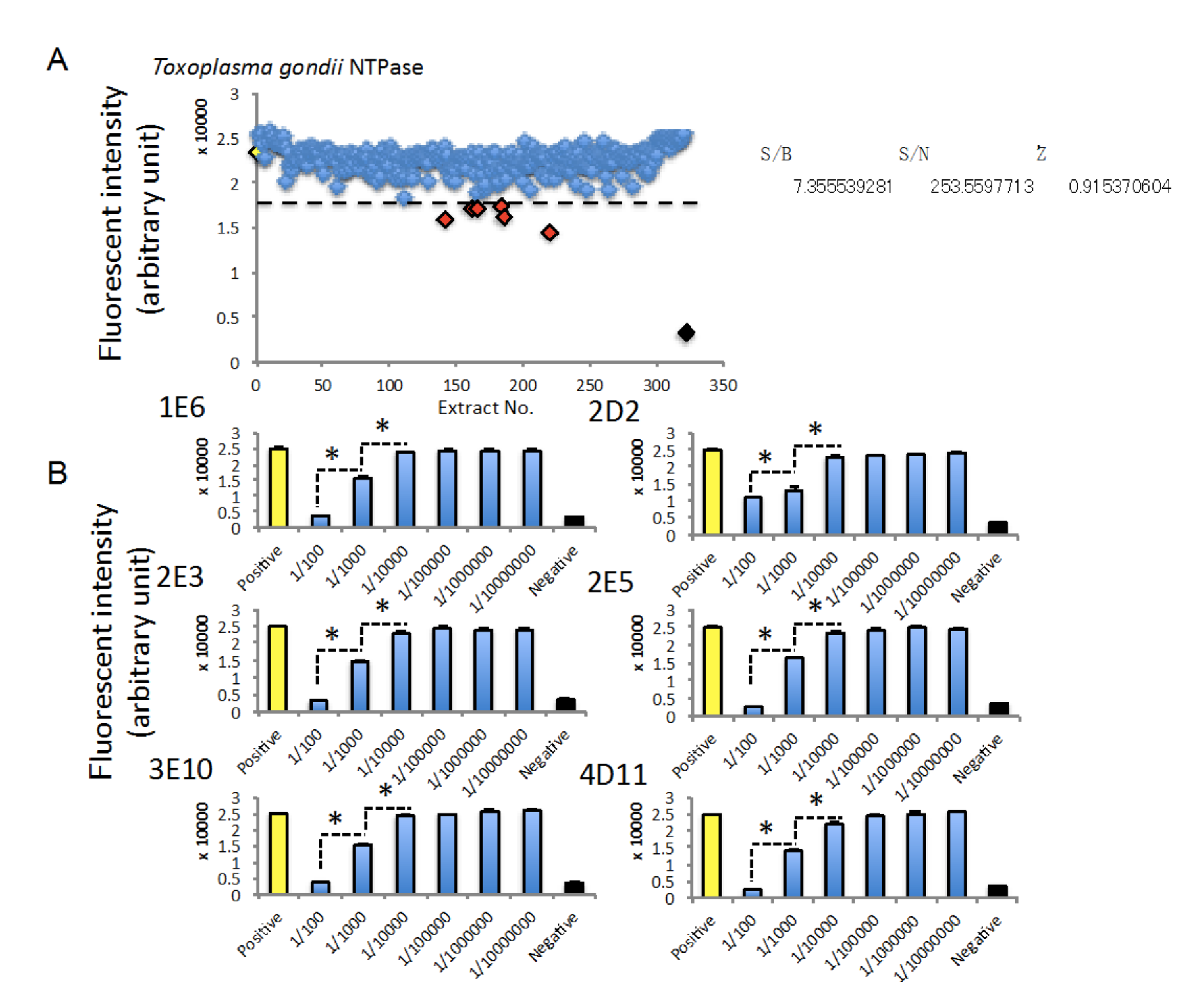
2.5. HTS Using an Extract Library Derived from Marine Bacteria in Order to Identify Extracts Inhibiting TgNTPase
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Extracts from Marine Bacteria
4.3. Classical Enzymatic Assay for NTPase
4.4. Combination Classical Enzymatic Assay with Fluorescent Assay for ADP
4.5. HTS Using Robot Arm
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P. Toxoplasmosis of Animals and Man; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar]
- Michael, C.; Sneller, H.; Clifford, L. Infections in the Immunocompromised Host in Clinical Immunology, 3rd ed.; Elsevirer: Amsterdam, The Netherland, 2008. [Google Scholar]
- Schäfer, G.; Hoffmann, C.; Arasteh, K.; Schürmann, D.; Stephan, C.; Jensen, B.; Stoll, M.; Bogner, J.R.; Faetkenheuer, G.; Rockstroh, J.; et al. Immediate versus deferred antiretroviral therapy in HIV-infected patients presenting with acute AIDS-defining events (toxoplasmosis, Pneumocystis jirovecii-pneumonia): A prospective, randomized, open-label multicenter study (IDEAL-study). AIDS Res. Ther. 2019, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, P.; Scherger, S.; Benamu, E.; Bajrovic, V.; Jackson, W.; Hage, C.A.; Hakki, M.; Baddley, J.W.; Abidi, M.Z. Toxoplasmosis in non-cardiac solid organ transplant recipients: A case series and review of literature. Transpl. Infect. Dis. 2019, 26, e13218. [Google Scholar] [CrossRef]
- Rivera, E.M.; Lavayén, S.N.; Sánchez, P.; Martins, C.M.A.; Gómez, E.; Rodríguez, J.P.; Arias, M.E.; Silva, A.P.; Angel, S.O. Toxoplasma gondii seropositivity associated to peri-urban living places in pregnant women in a rural area of Buenos Aires province, Argentina. Parasite Epidemiol. Control. 2019, 7, e00121. [Google Scholar] [CrossRef] [PubMed]
- Donahoe, S.L.; Lindsay, S.A.; Krockenberger, M.; Phalen, D.; Šlapeta, J. A review of neosporosis and pathologic findings of Neospora caninum infection in wildlife. Int. J. Parasitol. Parasites Wildl. 2015, 4, 216–238. [Google Scholar] [CrossRef] [PubMed]
- Crookshanks, J.L.; Taylor, S.M.; Haines, D.M.; Shelton, G.D. Treatment of canine pediatric Neospora caninum myositis following immunohistochemicalidentification of tachyzoites in muscle biopsies. Can. Vet. J. 2007, 48, 506–508. [Google Scholar] [PubMed]
- Bartner, L.R.; McGrath, S.; Drury, A.; Chen, A.V.; Morris, A.; Brewer, M.; Hall, M.; Lappin, M.R. Testing for Bartonella ssp. DNA in cerebrospinal fluid of dogs with inflammatory central nervous system disease. J. Vet. Intern. Med. 2018, 32, 1983–1988. [Google Scholar] [CrossRef] [PubMed]
- Changoluisa, D.; Rivera-Olivero, I.A.; Echeverria, G.; Garcia-Bereguiain, M.A.; de Waard, J.H. Working group “Applied Microbiology” of the School of Biological Sciences and Engineering at Yachay Tech University. Serology for Neosporosis, Q fever and Brucellosis to assess the cause of abortion in two dairy cattleherds in Ecuador. BMC Vet. Res. 2019, 15, 194. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Martínez, M.E.; Cisterna, C.A.B.; Romero, R.C.E.; Huacho, M.A.Q.; Bermabé, A.M.; Albornoz, L.A.L. Evaluation of abortions spontaneously induced by Neospora caninum and risk factors in dairy cattle from Lima, Peru. Rev. Bras. Parasitol. Vet. 2019, 28, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Matoba, K.; Shiba, T.; Takeuchi, T.; Sibley, L.D.; Seiki, M.; Kikyo, F.; Horiuchi, T.; Asai, T.; Harada, S. Crystallization and preliminary X-ray structural analysis of nucleoside triphosphate hydrolases from Neospora caninum and Toxoplasma gondii. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1445–1448. [Google Scholar] [CrossRef] [PubMed]
- Nakaar, V.; Beckers, C.J.; Polotsky, V.; Joiner, K.A. Basis for substrate specificity of the Toxoplasma gondii nucleoside triphosphate hydrolase. Mol. Biochem. Parasitol. 1998, 97, 209–220. [Google Scholar] [CrossRef]
- Pastor-Fernández, I.; Regidor-Cerrillo, J.; Álvarez-García, G.; Marugán-Hernández, V.; García-Lunar, P.; Hemphill, A.; Ortega-Mora, L.M. The tandemly repeated NTPase (NTPDase) from Neospora caninum is a canonical dense granuleprotein whose RNA expression, protein secretion and phosphorylation coincides with the tachyzoite egress. Parasit. Vectors 2016, 9, 352. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.A.; Qi, H.; Riehl, A.; Beckers, C.; Nakaar, V.; Joiner, K.A. Induced activation of the Toxoplasma gondii nucleoside triphosphate hydrolase leads to depletion of host cell ATP levels and rapid exit of intracellular parasites from infected cells. J. Biol. Chem. 1998, 273, 12352–12359. [Google Scholar] [CrossRef] [PubMed]
- Olias, P.; Sibley, L.D. Functional Analysis of the Role of Toxoplasma gondii Nucleoside Triphosphate Hydrolases I and II in Acute Mouse Virulence and Immune Suppression. Infect. Immun. 2016, 84, 1994–2001. [Google Scholar] [CrossRef] [PubMed]
- Leineweber, M.; Spekker-Bosker, K.; Ince, V.; Schares, G.; Hemphill, A.; Eller, S.K.; Däubener, W. First Characterization of the Neospora caninum Dense Granule Protein GRA9. Biomed. Res. Int. 2017, 2017, 6746437. [Google Scholar] [CrossRef] [PubMed]
- Krug, U.; Zebisch, M.; Krauss, M.; Sträter, N. Structural insight into activation mechanism of Toxoplasma gondii nucleoside triphosphatediphosphohydrolases by disulfide reduction. J. Biol. Chem. 2012, 287, 3051–3066. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, K.; Kojima, H.; Okabe, T.; Nagano, T. Development of a highly sensitive, high-throughput assay for glycosyltransferases using enzyme-coupled fluorescence detection. Anal. Biochem. 2014, 447, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Takeuchi, T.; Diffenderfer, J.; Sibley, L.D. Identification of small-molecule inhibitors of nucleoside triphosphate hydrolase in Toxoplasmagondii. Antimicrob. Agents Chemother. 2002, 46, 2393–2399. [Google Scholar] [CrossRef] [PubMed]
- Nakajima-Nakano, K.; Makioka, A.; Yamashita, N.; Matsuo, N.; Asai, T. Evaluation of serodiagnosis of toxoplasmosis by using the recombinant nucleoside triphosphate hydrolase isoforms expressed in Escherichia coli. Parasitol. Int. 2000, 48, 215–222. [Google Scholar] [CrossRef]
- Kurata, R.; Kumagai, A.; Cui, X.; Harada, M.; Nagai, J.; Yoshida, Y.; Ozaki, K.I.; Tanaka, Y.; Yonezawa, T. Establishment of Novel Reporter Cells Stably Maintaining Transcription Factor-driven Human Secreted Alkaline Phosphatase Expression. Curr. Pharm. Biotechnol. 2018, 19, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Chung, T.D.Y.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plate | Enzyme | S/B | S/N | Z’-Factor | Measurement | Number of Sample |
|---|---|---|---|---|---|---|
| 96 well | NcNTPase | 4.13 | 3.91 | 0.074 | Absorbance | n = 3 |
| TgNTPase | 7.71 | 38.4 | 0.077 | Absorbance | n = 3 | |
| NcNTPase | 7.60 | 234.9 | 0.960 | Fluorescence | n = 3 | |
| TgNTPase | 11.28 | 342.4 | 0.876 | Fluorescence | n = 3 | |
| 384 well | NcNTPase | 15.3 | 551.3 | 0.970 | Fluorescence | n = 16 |
| NcNTPase | 12.7 | 344.3 | 0.970 | Fluorescence | n = 16 | |
| TgNTPase | 8.44 | 500.2 | 0.925 | Fluorescence | n = 16 | |
| TgNTPase | 8.28 | 394.1 | 0.890 | Fluorescence | n = 16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harada, M.; Nagai, J.; Kurata, R.; Shimizu, K.; Cui, X.; Isagawa, T.; Semba, H.; Ishihara, J.; Yoshida, Y.; Takeda, N.; et al. Establishment of Novel High-Standard Chemiluminescent Assay for NTPase in Two Protozoans and Its High-Throughput Screening. Mar. Drugs 2020, 18, 161. https://doi.org/10.3390/md18030161
Harada M, Nagai J, Kurata R, Shimizu K, Cui X, Isagawa T, Semba H, Ishihara J, Yoshida Y, Takeda N, et al. Establishment of Novel High-Standard Chemiluminescent Assay for NTPase in Two Protozoans and Its High-Throughput Screening. Marine Drugs. 2020; 18(3):161. https://doi.org/10.3390/md18030161
Chicago/Turabian StyleHarada, Masamitsu, Jun Nagai, Riho Kurata, Kenji Shimizu, Xiaofeng Cui, Takayuki Isagawa, Hiroaki Semba, Jun Ishihara, Yasuhiro Yoshida, Norihiko Takeda, and et al. 2020. "Establishment of Novel High-Standard Chemiluminescent Assay for NTPase in Two Protozoans and Its High-Throughput Screening" Marine Drugs 18, no. 3: 161. https://doi.org/10.3390/md18030161
APA StyleHarada, M., Nagai, J., Kurata, R., Shimizu, K., Cui, X., Isagawa, T., Semba, H., Ishihara, J., Yoshida, Y., Takeda, N., Maemura, K., & Yonezawa, T. (2020). Establishment of Novel High-Standard Chemiluminescent Assay for NTPase in Two Protozoans and Its High-Throughput Screening. Marine Drugs, 18(3), 161. https://doi.org/10.3390/md18030161