Anti-Inflammatory Activity of Glycolipids and a Polyunsaturated Fatty Acid Methyl Ester Isolated from the Marine Dinoflagellate Karenia mikimotoi

 ,
, 

Abstract
1. Introduction
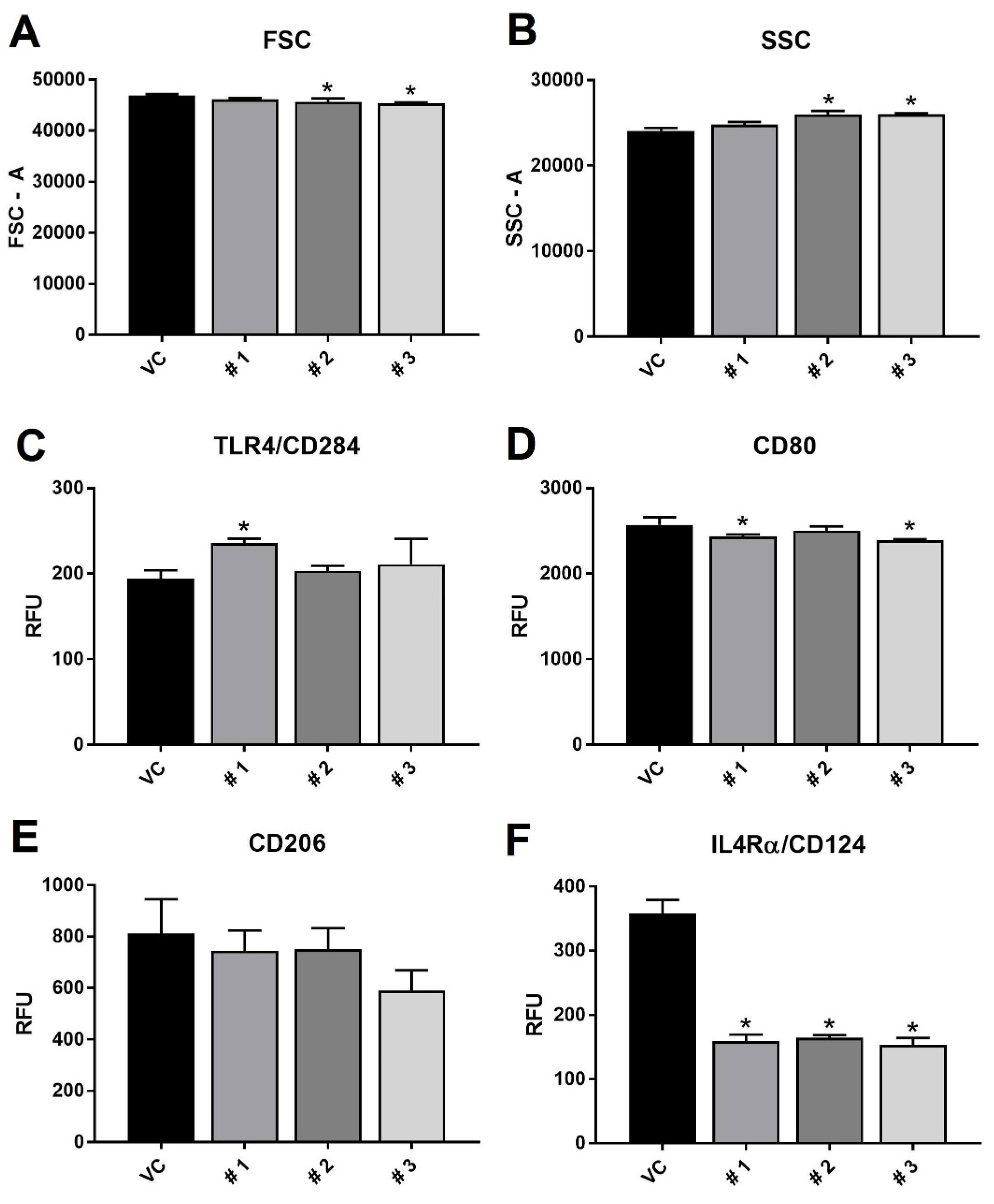
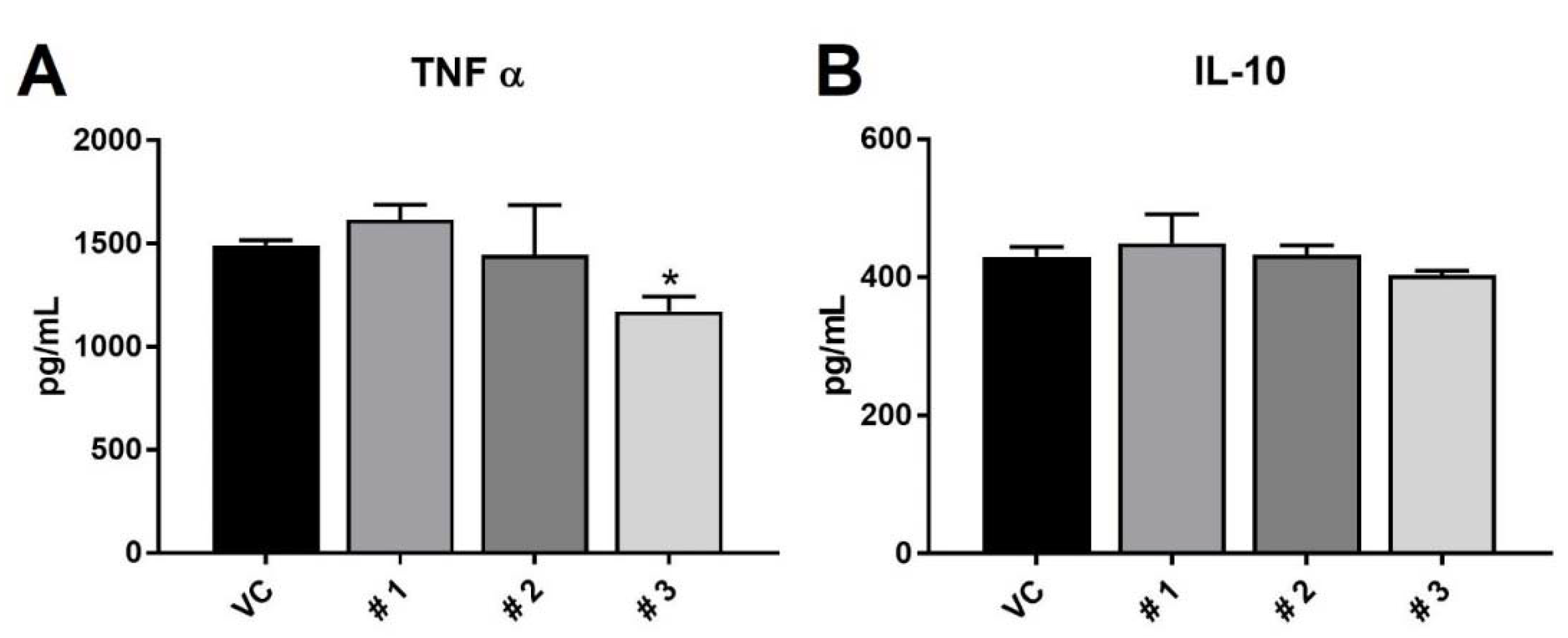
2. Results and Discussion
3. Experimental Section
3.1. Strain and Cultivation
3.2. Extraction and Isolation
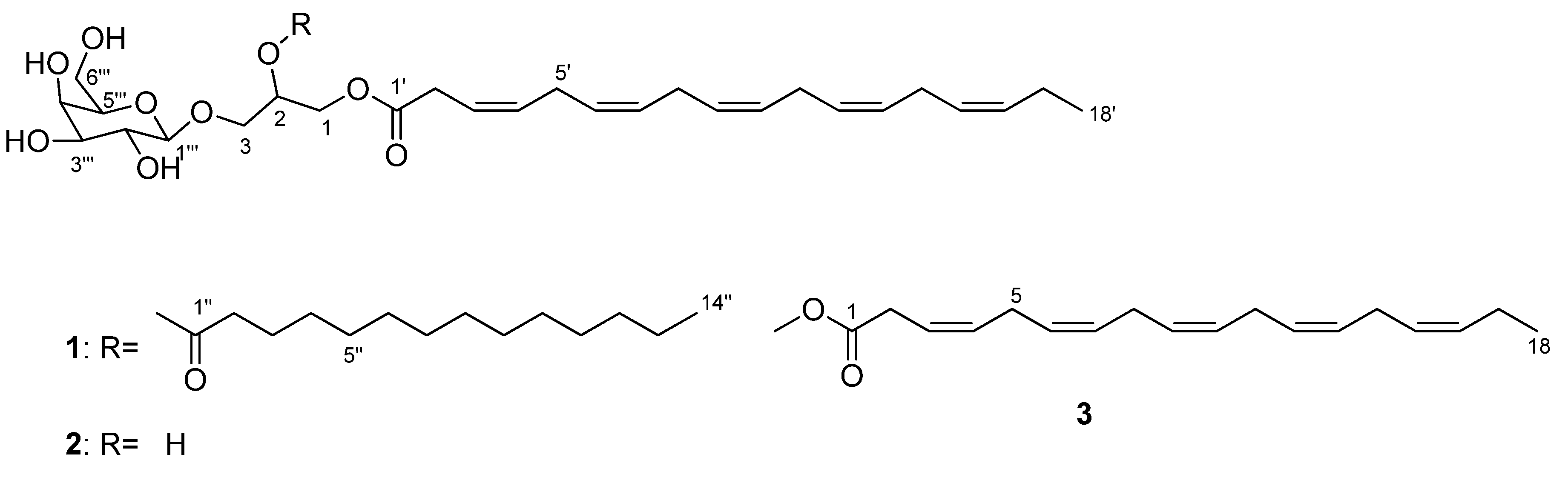
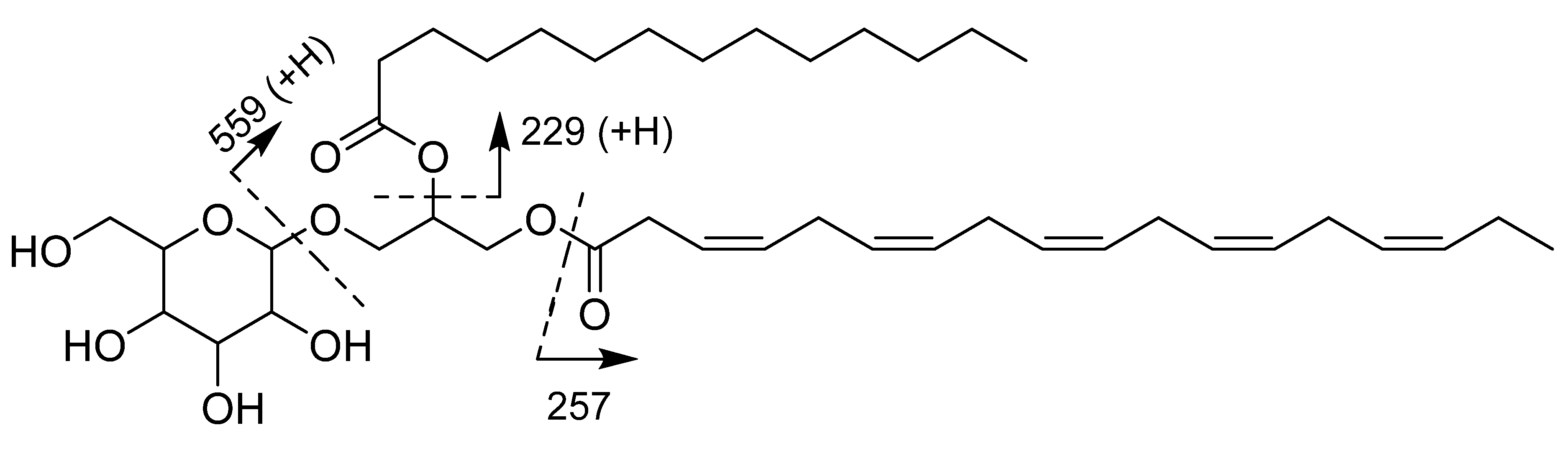
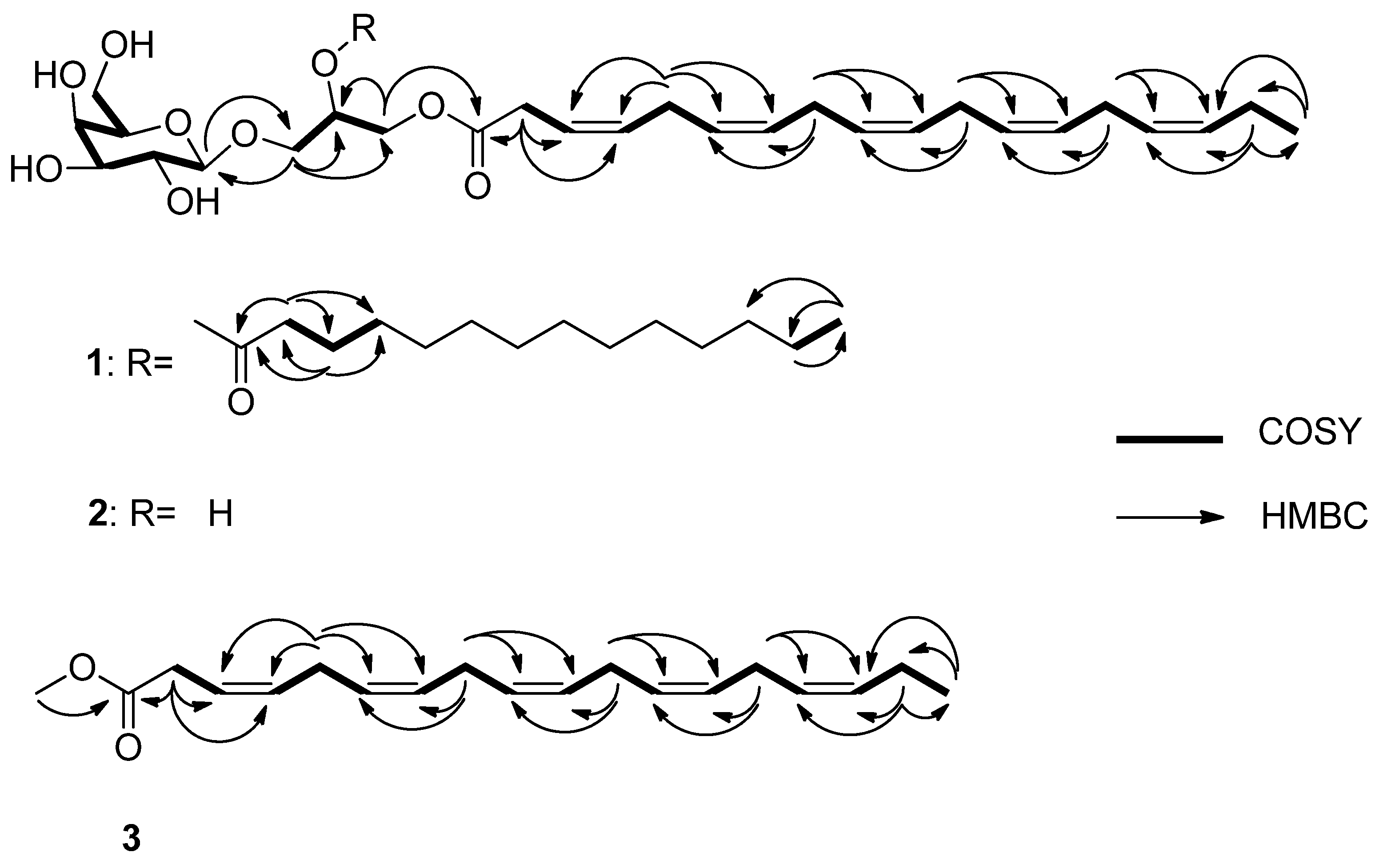
3.3. Structure Elucidation
3.3.1. Monogalactosyldiacylglycerol 1
3.3.2. Monogalactosylmonoacylglycerol 2
3.3.3. Methyl (3Z,6Z,9Z,12Z,15Z)-octadeca-3,6,9,12,15-pentaenoate 3
3.4. Antimicrobial Bioassay Testing
3.5. Flow Cytometry and ELISA Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Imada, C.; Koseki, N.; Kamata, M.; Kobayashi, T.; Hamada-Sato, N. Isolation and characterization of antibacterial substances produced by marine actinomycetes in the presence of seawater. Actinomycetologica 2007, 21, 27–31. [Google Scholar] [CrossRef]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Dahl, E.; Tangen, K. 25 years experience with Gyrodinium aureolum in Norwegian waters 1993. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 15–21. [Google Scholar]
- Davidson, K.; Miller, P.; Wilding, T.A.; Shutler, J.; Bresnan, E.; Kennington, K.; Swan, S. A large and prolonged bloom of Karenia mikimotoi in Scottish waters in 2006. Harmful Algae 2009, 8, 349–361. [Google Scholar] [CrossRef]
- Lindahl, O. On the development of a Gyrodinium aureolum occurrence on the Swedish west coast in 1982. Mar. Biol. 1983, 77, 143–150. [Google Scholar] [CrossRef]
- Raine, R.; O’Boyle, S.; O’Higgins, T.; White, M.; Patching, J.; Cahill, B.; McMahon, T. A satellite and field portrait of a Karenia mikimotoi bloom off the south coast of Ireland, August 1998. Hydrobiologia 2001, 465, 187–193. [Google Scholar] [CrossRef]
- Hodgkiss, I.; Yang, Z. New and dominant species from Sam Xing wan, sai Kung during the 1998 massive fish killing red tide in Hong Kong. Harmful Algal Blooms 2000, 62, 65. [Google Scholar]
- Gentien, P.; Lunven, M.; Lazure, P.; Youenou, A.; Crassous, M.P. Motility and autotoxicity in Karenia mikimotoi (Dinophyceae). Philos. Trans. R Soc. B Biol. Sci. 2007, 362, 1937–1946. [Google Scholar] [CrossRef]
- Tatters, A.O.; Muhlstein, H.I.; Tomas, C.R. The hemolytic activity of Karenia selliformis and two clones of Karenia brevis throughout a growth cycle. J. Appl. Phycol. 2010, 22, 435–442. [Google Scholar] [CrossRef]
- Mooney, B.D.; Nichols, P.D.; De Salas, M.F.; Hallegraeff, G.M. Lipid, fatty acid, and sterol composition of eight species of Kareniaceae (Dinophyta): Chemotaxonomy and putative lipid phycotoxins. J. Phycol. 2007, 43, 101–111. [Google Scholar] [CrossRef]
- Yasumoto, T.; Underdal, B.; Aune, T.; Hormazabal, V.; Skulberg, O.M.; Oshima, Y. Screening for hemolytic and ichthyotoxic components of Chryssochromulina polylepis and Gyrodinium aureolum from Norwegian coastal waters. In Toxic Marine Phytoplankton; Graneli, E., Sundstrom, B., Edler, L., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 436–440. [Google Scholar]
- Matsubara, K.; Matsumoto, H.; Mizushina, Y.; Mori, M.; Nakajima, N.; Fuchigami, M.; Yoshida, H.; Hada, T. Inhibitory effect of glycolipids from spinach on in vitro and ex vivo angiogenesis. Oncol. Rep. 2005, 14, 157–160. [Google Scholar]
- Wang, R.; Furumoto, T.; Motoyama, K.; Okazaki, K.; Kondo, A.; Fukui, H. Possible antitumor promoters in Spinacia oleracea (Spinach) and comparison of their contents among cultivars. Biosci. Biotechnol. Biochem. 2002, 66, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Nakata, K. High resistance to oxygen radicals and heat is caused by a galactoglycerolipid in Microbacterium sp. M874. J. Biochem. 2000, 127, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Nakata, K.; Guo, C.T.; Matsufugi, M.; Yoshimoto, A.; Inagaki, M.; Higuchi, R.; Suzuki, Y. Influenza A virus-binding activity of glycoglycerolipids of aquatic bacteria. J. Biochem. 2000, 127, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Janwitayanuchit, W.; Suwanborirux, K.; Patarapanich, C.; Pummangura, S.; Lipipun, V.; Vilaivan, T. Synthesis and anti-herpes simplex viral activity of monoglycosyl diglycerides. Phytochemistry 2003, 64, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.; Rossi, C.; Marcolongo, G.; Di Lena, A.; Venzo, A.; Barrie, C.P.; Corda, D. Selective in vivo anti-inflammatory action of the galactolipid monogalactosyldiacylglycerol. Eur. J. Pharmacol. 2005, 524, 159–168. [Google Scholar] [CrossRef]
- Tanaka, R.; Sakano, Y.; Nagatsu, A.; Shibuya, M.; Ebizuka, Y.; Goda, Y. Synthesis of digalactosyl diacylglycerols and their structure–inhibitory activity on human lanosterol synthase. Bioorg. Med. Chem. Lett. 2005, 15, 159–162. [Google Scholar] [CrossRef]
- Nagatsu, A.; Watanabe, M.; Ikemoto, K.; Hashimoto, M.; Murakami, N.; Sakakibara, J.; Tokuda, H.; Nishino, H.; Iwashima, A.; Yazawa, K. Synthesis and structure—anti-tumor-promoting activity relationship of monogalactosyl diacylglycerols. Bioorg. Med. Chem. Lett. 1994, 4, 1619–1622. [Google Scholar] [CrossRef]
- Colombo, D.; Compostella, F.; Ronchetti, F.; Scala, A.; Toma, L.; Mukainaka, T.; Nagatsu, A.; Konoshima, T.; Tokuda, H.; Nishino, H. Inhibitory effects of monoacylated 2-O-β-galactosylglycerols on Epstein–Barr virus activation: The significant role of the hexanoyl chain. Cancer Lett. 1999, 143, 1–4. [Google Scholar] [CrossRef]
- Cateni, F.; Bonivento, P.; Procida, G.; Zacchigna, M.; Scialino, G.; Banfi, E. Chemoenzymatic synthesis and in vitro studies on the hydrolysis of antimicrobial monoglycosyl diglycerides by pancreatic lipase. Bioorg. Med. Chem. Lett. 2007, 17, 1971–1978. [Google Scholar] [CrossRef]
- Leblond, J.D.; Dodson, J.; dahmen, J.I. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. VII. Evidence against galactolipid production and plastid presence in the heterotrophic, basal dinoflagellate, Oxyrrhis marina. Eur. J. Phycol. 2013, 48, 309–317. [Google Scholar] [CrossRef]
- Gray, C.G.; Lasiter, A.D.; Li, C.; Leblond, J.D. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. I. Peridinin-containing taxa. Eur. J. Phycol. 2009, 44, 191–197. [Google Scholar] [CrossRef]
- Leblond, J.D.; Lasiter, A.D. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. II. Lepidodinium chlorophorum, Karenia brevis, and Kryptoperidinium foliaceum, three dinoflagellates with aberrant plastids. Eur. J. Phycol. 2009, 44, 199–205. [Google Scholar] [CrossRef][Green Version]
- Hiraga, Y.; Shikano, T.; Widianti, T.; Ohkata, K. Three new glycolipids with cytolytic activity from cultured marine dinoflagellate Heterocapsa circularisquama. Nat. Prod. Res. 2008, 22, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Bond, T.J.; Herald, D.L.; Penny, M.; Doubek, D.L.; Williams, M.D.; Pettit, R.K.; Hooper, J.N.A. Isolation and structure of spongilipid from the Republic of Singapore marine porifera Spongia cf. hispida. Can. J. Chem. 1997, 75, 920–925. [Google Scholar] [CrossRef]
- Ishii, T.; Okino, T.; Mino, Y. A ceramide and cerebroside from the starfish Asterias amurensis Lu1tken and their plant-growth promotion activities. J. Nat. Prod. 2006, 69, 1080–1082. [Google Scholar] [CrossRef]
- Scribe, P.; Guezennec, J.; Dagaut, J.; Pepe, C.; Saliot, A. Identification of the position and the stereochemistry of the double bond in monounsaturated fatty acid methyl esters by gas chromatography/mass spectrometry of dimethyl disulfide derivatives. Anal. Chem. 1988, 60, 928–931. [Google Scholar] [CrossRef]
- Ghioni, C.; Porter, A.E.A.; Sadler, I.H.; Tocher, D.R.; Sargent, J.R. Cultured fish cells metabolize octadecapentaenoic acid (all-cis Δ3,6,9,12,15-18:5) to octadecatetraenoic acid (all-cis Δ6,9,12,15-18:4) via its 2-trans intermediate (trans Δ2, all-cis Δ6,9,12,15-18:5). Lipids 2001, 36, 145–153. [Google Scholar] [CrossRef]
- Kobayashi, M.; Hayashi, K.; Kawazoe, K.; Kitagawa, I. Marine natural products. XXIX. Heterosigma-glycolipids I, II, III, and IV, four diacylglyceroglycolipids possessing ω3-polyunsaturated fatty acid residues, from the raphidopycean dinoflagellate Heterosigma akashiwo. Chem. Pharm. Bull. 1992, 40, 1404–1410. [Google Scholar] [CrossRef]
- Iwagawa, T.; Masuda, T.; Nakatani, M. Polyunsaturated fatty acid methyl esters from a soft Coral Xenia sp. Rep. Fac. Sci. Kagoshima Univ. 1993, 26, 63–67. [Google Scholar]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.; Mardani, M.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Keeler, D.M.; Grandal, M.K.; McCall, J.R. Brevenal, a marine natural product, is anti-inflammatory and an immunomodulator of macrophage and lung epithelial cells. Mar. Drugs 2019, 17, 184. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Grütz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginet, J. Biology of interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef]
- Adolf, J.E.; Place, A.R.; Stoecker, D.K.; Harding, L.W., Jr. Modulation of polyunsaturated fatty acids in mixotrophic Karlodinium veneficum (Dinophyceae) and its prey, Storeatula major (Cryptophyceae). J. Phycol. 2007, 43, 1259–1270. [Google Scholar] [CrossRef]
- Okuyama, H.; Morita, N.; Kogame, K. Occurrence of octadecapentaenoic acid in lipids of a cold stenothermic alga, prymnesiophyte strain B. J. Phycol. 1992, 28, 465–472. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | ||||
| No. | δC, mult. b | δH (J in Hz) | COSY | HMBC |
|---|---|---|---|---|
| 1, Gly | 64.5, CH2 | 4.25, dd (12.1, 6.3) | 3, 2 | 2, 1’ |
| 4.43, dd (12.1, 3.0) | ||||
| 2, Gly | 71.8, CH | 5.27, m | 1, 3 | |
| 3, Gly | 68.8, CH2 | 3.74, m | 2, 1 | 2, 1, 1‴ |
| 4.00, dd (10.7, 4.0) | ||||
| 1’ | 173.1, C | |||
| 2’ | 33.2, CH2 | 3.14, d (4.9) | 3’ | 1’, 3’, 4’ |
| 3’ | 132.6, CH | 5.55, t (4.9) | 2’ | |
| 4’ | 122.4, CH | 5.55, t (4.9) | 5’ | |
| 6’-7’ | 128.3/129.7, CH | 5.36, m | 5’/8’ | |
| 9’-10’ | 128.3/129.7, CH | 5.36, m | 8’/11’ | |
| 12’-13’ | 128.3/129.7, CH | 5.36, m | 11’/14’ | |
| 15’ | 129.3, CH | 5.36, m | 14’ | |
| 16’ | 132.9, CH | 5.36, m | 17’ | |
| 17’ | 21.6, CH2 | 2.09, quint (7.6) | 16’, 18’ | 15’, 16’, 18’ |
| 18’ | 14.7, CH3 | 0.98, t (7.6) | 17’ | 16’, 17’ |
| 5’ | 26.8c, CH2 | 2.84, m | 4’, 6’ | 3’, 4’, 6’, 7’ |
| 8’ | 26.7c, CH2 | 2.84, m | 7’, 9’ | 6’, 7’, 9’, 10’ |
| 11’ | 26.6c, CH2 | 2.84, m | 10’, 12’ | 9’, 10’, 12’, 13’ |
| 14’ | 26.6c, CH2 | 2.84, m | 13’, 15’ | 12’, 13’, 15’, 16’ |
| 1″ | 174.6, C | |||
| 2″ | 35.2, CH2 | 2.33, t (7.4) | 3″ | 1″, 3″, 4″ |
| 3″ | 26.1, CH2 | 1.61, m | 2″, 4″ | 1″, 2″, 4″ |
| 4″ | 30.6, CH2 | 1.33, m | 3″ | |
| 5″-11″ | 30.2-30.8, CH2 | 1.33, m | ||
| 12″ | 33.1, CH2 | 1.33, m | ||
| 13″ | 23.8, CH2 | 1.33, m | 14″ | 14″ |
| 14″ | 14.5, CH3 | 0.90, t (7.3) | 13″ | 12″, 13″ |
| 1‴ | 105.4, CH | 4.22, d (7.5) | 2‴ | 3, 2‴ |
| 2‴ | 72.5, CH | 3.52, m | 1‴ | 3‴ |
| 3‴ | 75.0, CH | 3.45, dd (9.7, 3.2) | 4‴ | 2‴ |
| 4‴ | 70.3, CH | 3.82, brd (3.2) | 3‴, 5‴ | 2‴ |
| 5‴ | 76.9, CH | 3.51, m | 4‴, 6‴ | 1‴, 4‴, 6‴ |
| 6‴ | 62.5, CH2 | 3.75, m | 5‴ | 4‴, 5‴ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leutou, A.S.; McCall, J.R.; York, R.; Govindapur, R.R.; Bourdelais, A.J. Anti-Inflammatory Activity of Glycolipids and a Polyunsaturated Fatty Acid Methyl Ester Isolated from the Marine Dinoflagellate Karenia mikimotoi. Mar. Drugs 2020, 18, 138. https://doi.org/10.3390/md18030138
Leutou AS, McCall JR, York R, Govindapur RR, Bourdelais AJ. Anti-Inflammatory Activity of Glycolipids and a Polyunsaturated Fatty Acid Methyl Ester Isolated from the Marine Dinoflagellate Karenia mikimotoi. Marine Drugs. 2020; 18(3):138. https://doi.org/10.3390/md18030138
Chicago/Turabian StyleLeutou, Alain S., Jennifer R. McCall, Robert York, Rajeshwar R. Govindapur, and Andrea J. Bourdelais. 2020. "Anti-Inflammatory Activity of Glycolipids and a Polyunsaturated Fatty Acid Methyl Ester Isolated from the Marine Dinoflagellate Karenia mikimotoi" Marine Drugs 18, no. 3: 138. https://doi.org/10.3390/md18030138
APA StyleLeutou, A. S., McCall, J. R., York, R., Govindapur, R. R., & Bourdelais, A. J. (2020). Anti-Inflammatory Activity of Glycolipids and a Polyunsaturated Fatty Acid Methyl Ester Isolated from the Marine Dinoflagellate Karenia mikimotoi. Marine Drugs, 18(3), 138. https://doi.org/10.3390/md18030138