Fucoxanthin Inhibits Myofibroblast Differentiation and Extracellular Matrix Production in Nasal Polyp-Derived Fibroblasts via Modulation of Smad-Dependent and Smad-Independent Signaling Pathways


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
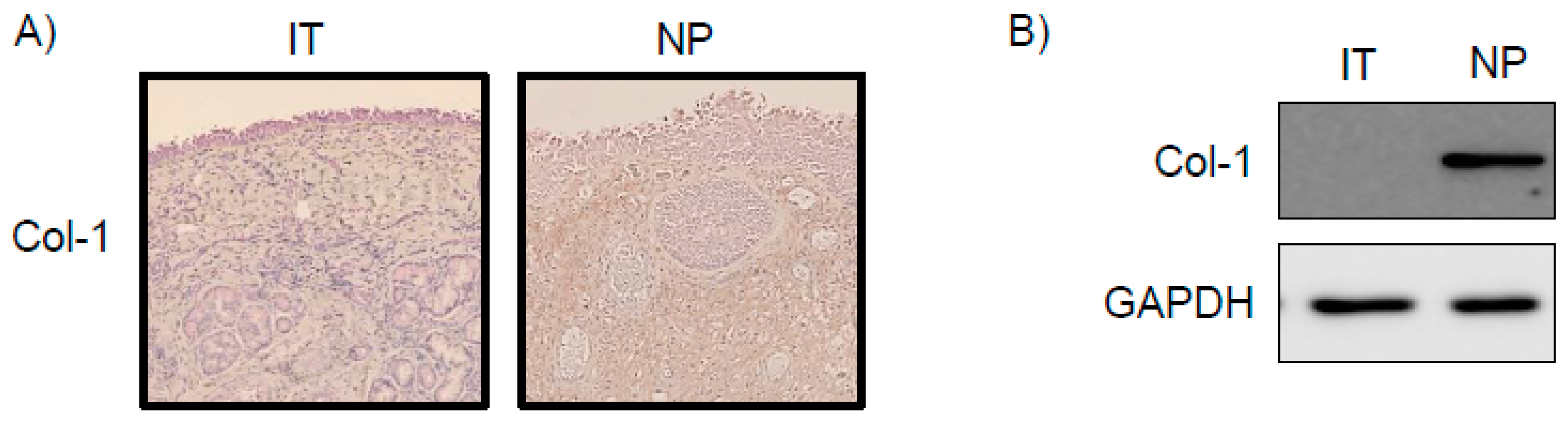
2.1. Expression of Col-1 in NP Tissues
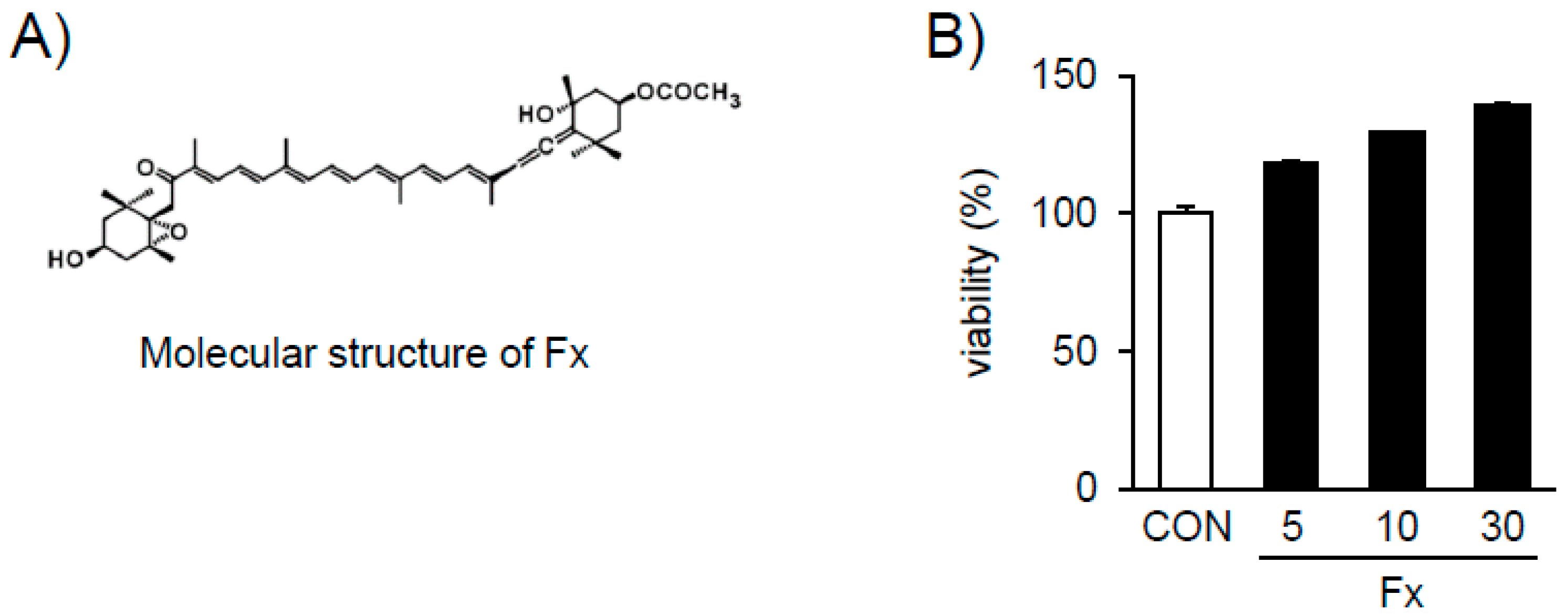
2.2. Effects of Fx on the Viability of NPDFs
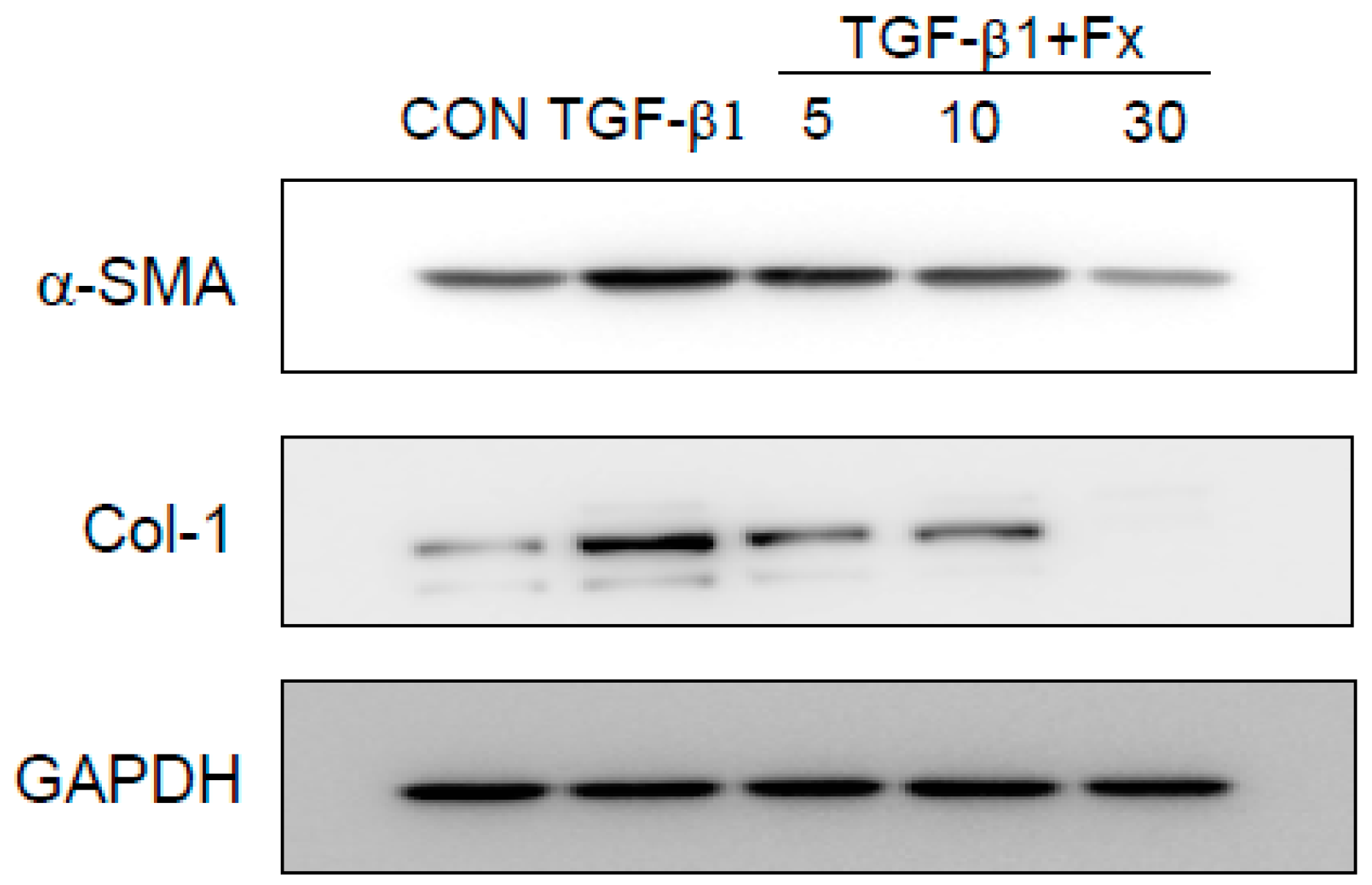
2.3. Effects of Fx on the Production of α-SMA and Col-1 in TGF-β1-Stimulated NPDFs
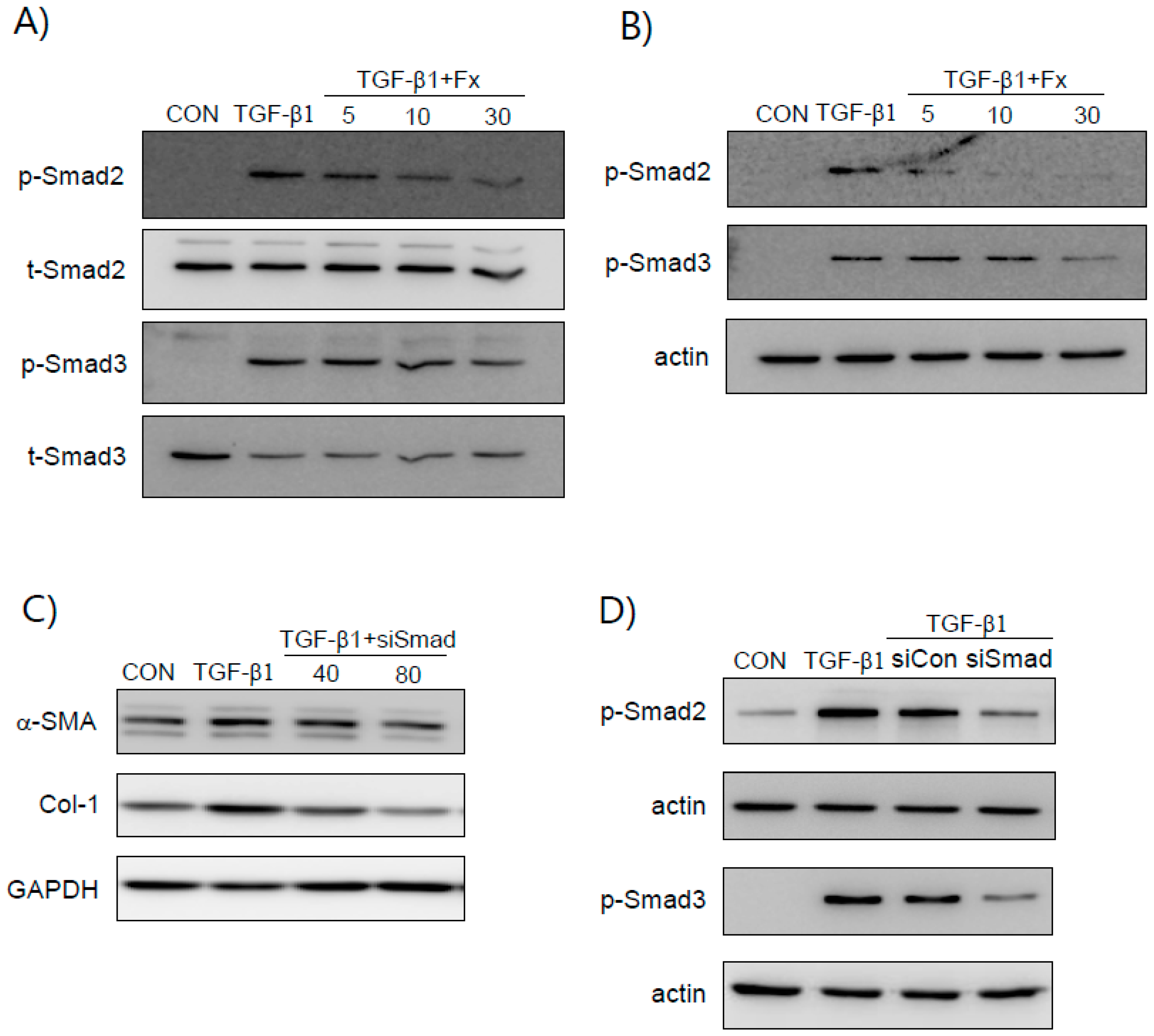
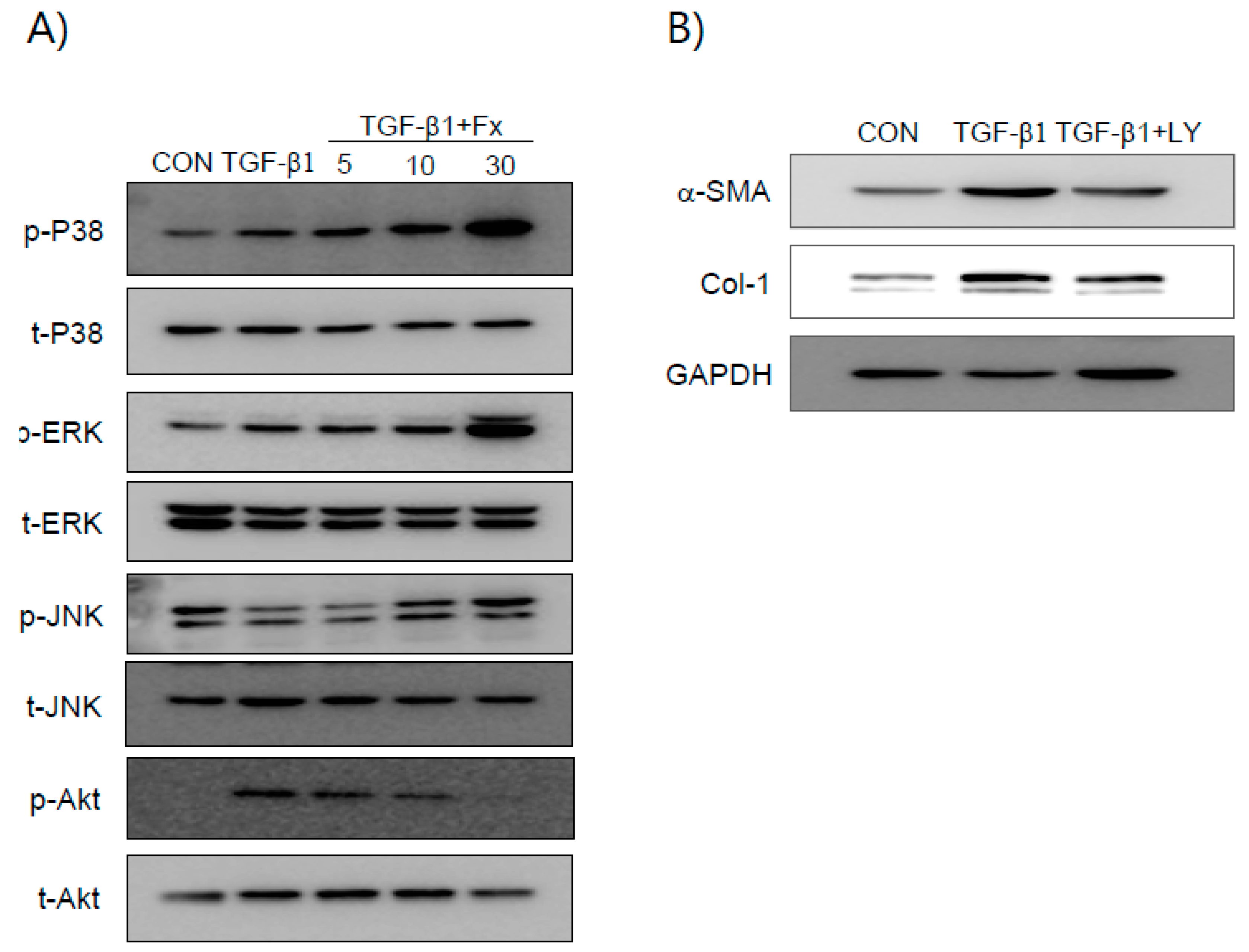
2.4. Fx Inhibits TGF-β1-Stimulated Smad-2/3-Dependent and Smad-2/3-Independent Signaling Pathways
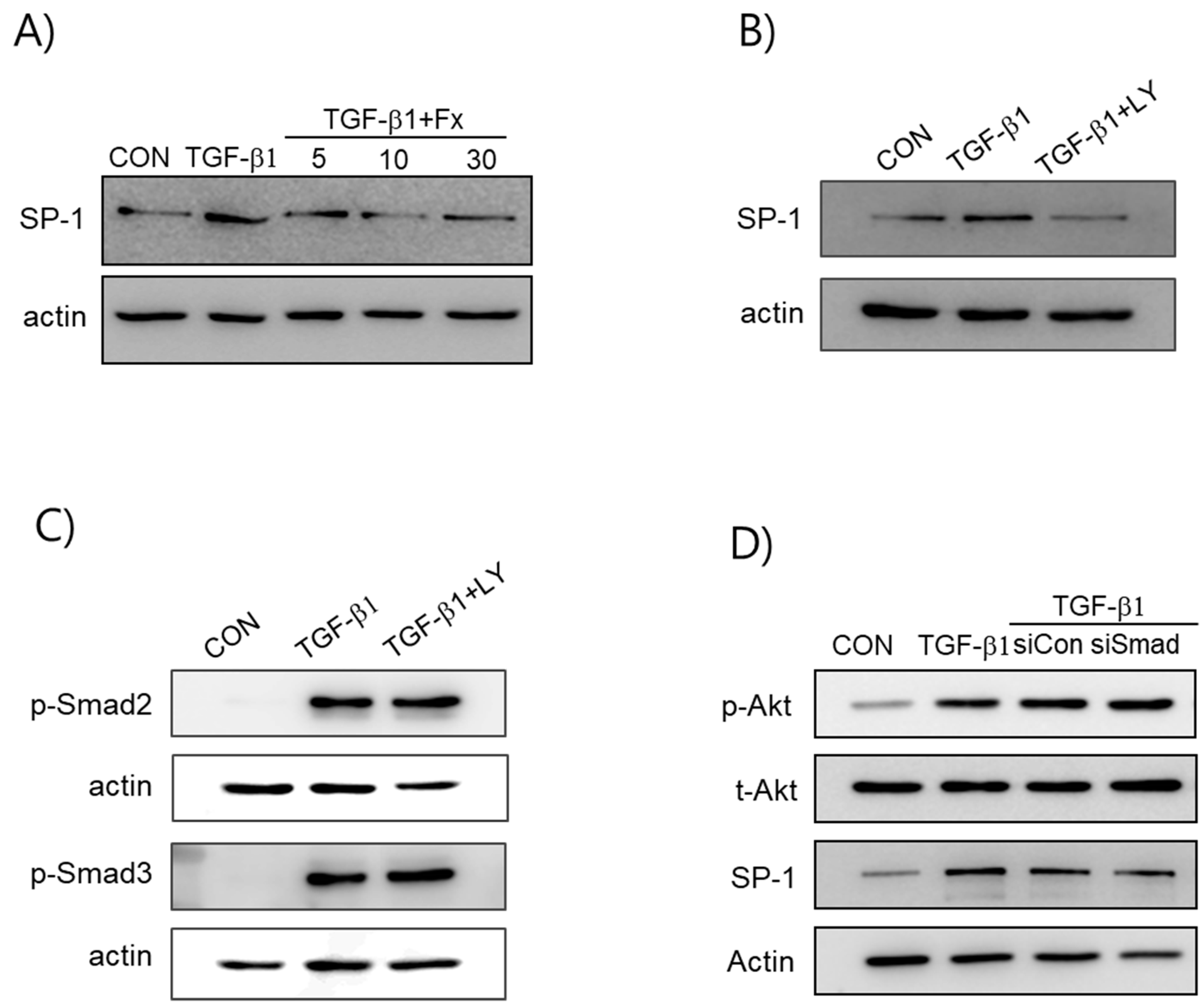
2.5. Fx Inhibits TGF-β1-Induced Signal Protein-1 (SP-1) Activation
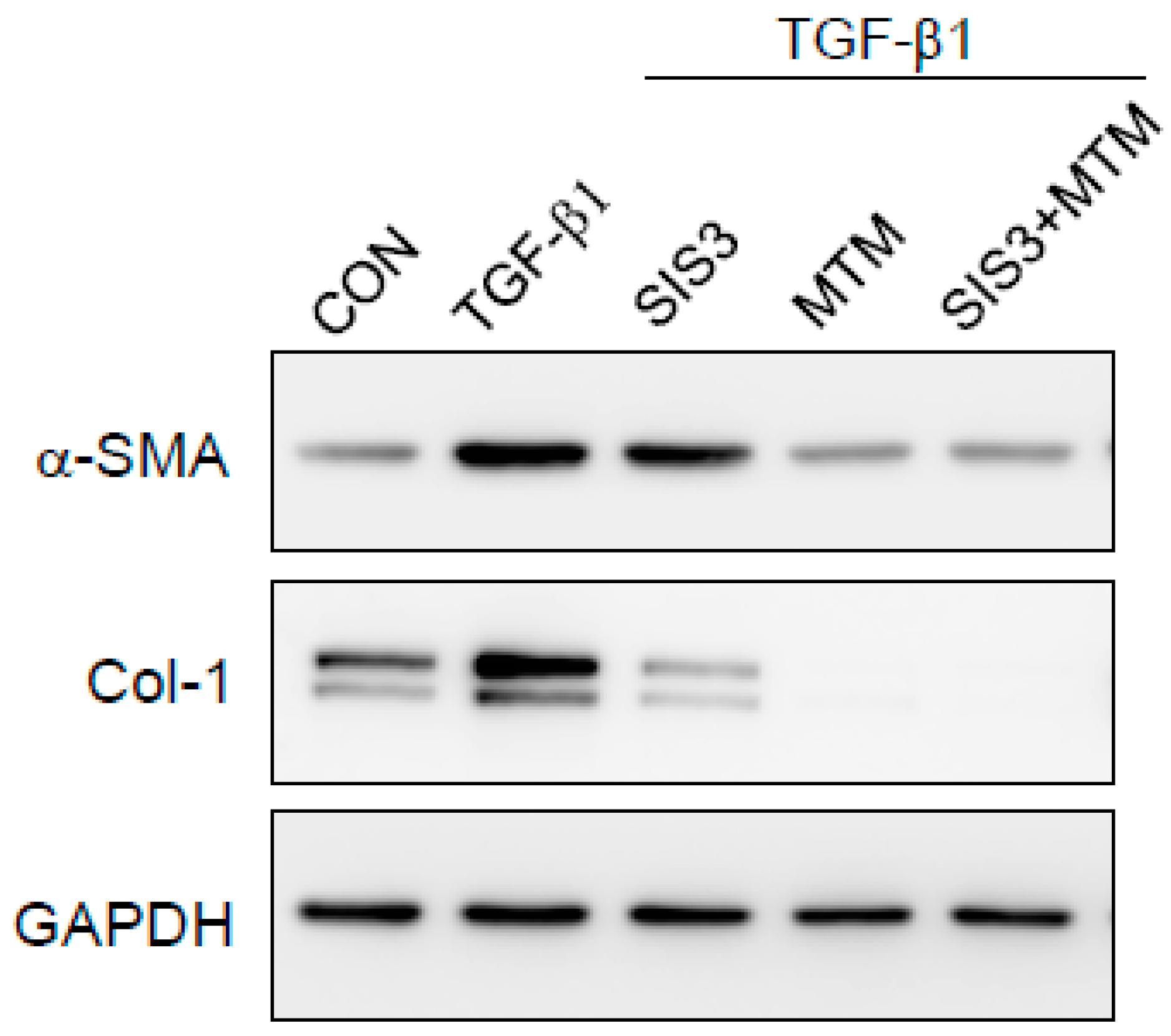
2.6. Synergistic Effects of Smad and SP-1 Signal Pathways on the Production of α-SMA and Col-1 Proteins
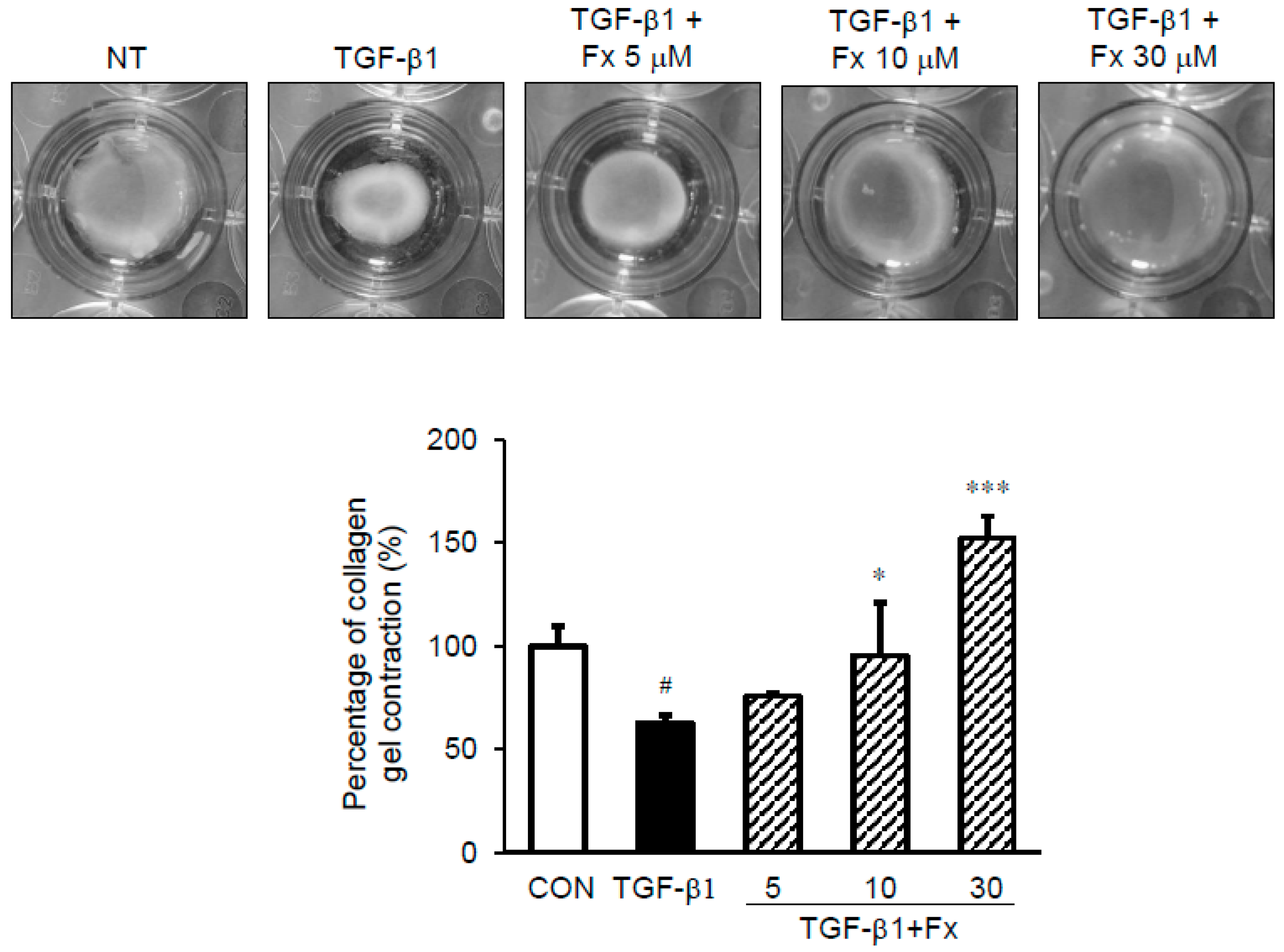
2.7. Effects of Fx on the Fibroblast Contractile Activity
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. NP-Derived Fibroblast Culture
4.3. Immunohistochemistry
4.4. Determining Cell Viability
4.5. Western Blot Analysis
4.6. Silencing of Smad 2/3 by Synthetic siRNAs
4.7. Rat Tail Type I Collagen Gel Contraction Assay
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Park, I.H.; Park, S.J.; Cho, J.S.; Moon, Y.M.; Kim, T.H.; Lee, S.H.; Lee, H.M. Role of reactive oxygen species in transforming growth factor beta1-induced alpha smooth-muscle actin and collagen production in nasal polyp-derived fibroblasts. Int. Arch. Allergy Immunol. 2012, 159, 278–286. [Google Scholar] [CrossRef] [PubMed]
- El Banhawy, O.; Al Abri, R.; Khalil, Y.; Shafy, I.; Fayaz, F. Update of pathogenesis and management of nasal polyposis. Menoufia Med. J. 2016, 29, 469–477. [Google Scholar] [CrossRef]
- Zaravinos, A.; Soufla, G.; Bizakis, J.; Spandidos, D.A. Expression analysis of VEGFA, FGF2, TGFbeta1, EGF and IGF1 in human nasal polyposis. Oncol. Rep. 2008, 19, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Soy, F.K.; Pinar, E.; Imre, A.; Calli, C.; Calli, A.; Oncel, S. Histopathologic parameters in chronic rhinosinusitis with nasal polyposis: Impact on quality of life outcomes. Int. Forum Allergy Rhinol. 2013, 3, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Hellquist, H.B. Nasal polyps update. Histopathology. Allergy Asthma Proc. 1996, 17, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ryu, N.H.; Shin, J.M.; Um, J.Y.; Park, I.H.; Lee, H.M. Wogonin inhibits transforming growth factor beta1-induced extracellular matrix production via the p38/activator protein 1 signaling pathway in nasal polyp-derived fibroblasts. Am. J. Rhinol. Allergy 2016, 30, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Muluk, N.B.; Arikan, O.K.; Atasoy, P.; Kilic, R.; Yalcinozan, E.T. The role of MMP-2, MMP-9, and TIMP-1 in the pathogenesis of nasal polyps: Immunohistochemical assessment at eight different levels in the epithelial, subepithelial, and deep layers of the mucosa. Ear Nose Throat J. 2015, 94, E1–E13. [Google Scholar] [PubMed]
- Buckley, C.D.; Pilling, D.; Lord, J.M.; Akbar, A.N.; Scheel-Toellner, D.; Salmon, M. Fibroblasts regulate the switch from acute resolving to chronic persistent inflammation. Trends Immunol. 2001, 22, 199–204. [Google Scholar] [CrossRef]
- Kendall, R.T.; Feghali-Bostwick, C.A. Fibroblasts in fibrosis: Novel roles and mediators. Front. Pharmacol. 2014, 5, 123. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.; Duffy, H.S. Fibroblasts and myofibroblasts: What are we talking about? J. Cardiovasc. Pharmacol. 2011, 57, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Park, I.H.; Cho, J.S.; Lee, Y.M.; Lee, H.M. The effect of macrolides on myofibroblast differentiation and collagen production in nasal polyp-derived fibroblasts. Am. J. Rhinol. Allergy 2010, 24, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Denney, L.; Byrne, A.J.; Shea, T.J.; Buckley, J.S.; Pease, J.E.; Herledan, G.M.; Walker, S.A.; Gregory, L.G.; Lloyd, C.M. Pulmonary Epithelial Cell-Derived Cytokine TGF-beta1 Is a Critical Cofactor for Enhanced Innate Lymphoid Cell Function. Immunity 2015, 43, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Pohlers, D.; Brenmoehl, J.; Loffler, I.; Muller, C.K.; Leipner, C.; Schultze-Mosgau, S.; Stallmach, A.; Kinne, R.W.; Wolf, G. TGF-β and fibrosis in different organs—Molecular pathway imprints. Biochim. Biophys. Acta 2009, 1792, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Neuzillet, C.; Tijeras-Raballand, A.; Cohen, R.; Cros, J.; Faivre, S.; Raymond, E.; de Gramont, A. Targeting the TGFbeta pathway for cancer therapy. Pharmacol. Ther. 2015, 147, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Lefaucheur, J.P.; Wang, Q.P.; Lesprit, E.; Poron, F.; Peynegre, R.; Escudier, E. Expression of the transforming growth factor beta isoforms in inflammatory cells of nasal polyps. Arch. Otolaryngol. Head Neck Surg. 1998, 124, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhou, P.; Liu, Y.; Liu, F.; Yi, X.; Liu, S.; Holtappels, G.; Bachert, C.; Zhang, N. The development of nasal polyp disease involves early nasal mucosal inflammation and remodelling. PLoS ONE 2013, 8, e82373. [Google Scholar] [CrossRef] [PubMed]
- Sivagnanam, S.P.; Yin, S.; Choi, J.H.; Park, Y.B.; Woo, H.C.; Chun, B.S. Biological Properties of Fucoxanthin in Oil Recovered from Two Brown Seaweeds Using Supercritical CO2 Extraction. Mar. Drugs 2015, 13, 3422–3442. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J. Fucoxanthin and Its Metabolite Fucoxanthinol in Cancer Prevention and Treatment. Mar. Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [PubMed]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. TGF-beta signaling in fibrosis. Growth Factors 2011, 29, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Sebe, A.; Leivonen, S.K.; Fintha, A.; Masszi, A.; Rosivall, L.; Kahari, V.M.; Mucsi, I. Transforming growth factor-beta-induced alpha-smooth muscle cell actin expression in renal proximal tubular cells is regulated by p38beta mitogen-activated protein kinase, extracellular signal-regulated protein kinase1,2 and the Smad signalling during epithelial-myofibroblast transdifferentiation. Nephrol. Dial. Transplant. 2008, 23, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Wick, G.; Backovic, A.; Rabensteiner, E.; Plank, N.; Schwentner, C.; Sgonc, R. The immunology of fibrosis: Innate and adaptive responses. Trends Immunol. 2010, 31, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Jin, Y.D.; Park, Y.K.; Yeon, S.H.; Xu, J.; Han, R.N.; Rha, K.S.; Kim, Y.M. IL-25-induced activation of nasal fibroblast and its association with the remodeling of chronic rhinosinusitis with nasal polyposis. PLoS ONE 2017, 12, e0181806. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Little, S.C.; Early, S.B.; Woodard, C.R.; Shonka, D.C., Jr.; Han, J.K.; Borish, L.; Steinke, J.W. Dual action of TGF-beta1 on nasal-polyp derived fibroblasts. Laryngoscope 2008, 118, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Kang, J.H.; Shin, J.M.; Park, I.H.; Lee, H.M. Inhibitory Effect of Delphinidin on Extracellular Matrix Production via the MAPK/NF-kappaB Pathway in Nasal Polyp-Derived Fibroblasts. Allergy Asthma Immunol. Res. 2015, 7, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Stroschein, S.L.; Wang, W.; Luo, K. Cooperative binding of Smad proteins to two adjacent DNA elements in the plasminogen activator inhibitor-1 promoter mediates transforming growth factor beta-induced smad-dependent transcriptional activation. J. Biol. Chem. 1999, 274, 9431–9441. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Moon, Y.M.; Park, I.H.; Um, J.Y.; Moon, J.H.; Park, S.J.; Lee, S.H.; Kang, H.J.; Lee, H.M. Epigenetic regulation of myofibroblast differentiation and extracellular matrix production in nasal polyp-derived fibroblasts. Clin. Exp. Allergy 2012, 42, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Bauge, C.; Cauvard, O.; Leclercq, S.; Galera, P.; Boumediene, K. Modulation of transforming growth factor beta signalling pathway genes by transforming growth factor beta in human osteoarthritic chondrocytes: Involvement of Sp1 in both early and late response cells to transforming growth factor beta. Arthritis Res. Ther. 2011, 13, R23. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, H.; Fricovsky, E.; Ihm, S.; Schimke, M.; Maya-Ramos, L.; Aroonsakool, N.; Ceballos, G.; Dillmann, W.; Villarreal, F.; Ramirez-Sanchez, I. Role for high-glucose-induced protein O-GlcNAcylation in stimulating cardiac fibroblast collagen synthesis. Am. J. Physiol. Cell Physiol. 2014, 306, C794–C804. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xiao, J.; Qing, X.; Xing, J.; Xia, Y.; Qi, J.; Liu, X.; Zhang, S.; Sheng, X.; Zhang, X.; et al. Sp1 Mediates a Therapeutic Role of MiR-7a/b in Angiotensin II-Induced Cardiac Fibrosis via Mechanism Involving the TGF-beta and MAPKs Pathways in Cardiac Fibroblasts. PLoS ONE 2015, 10, e0125513. [Google Scholar] [CrossRef] [PubMed]
- Yuge, A.; Nasu, K.; Matsumoto, H.; Nishida, M.; Narahara, H. Collagen gel contractility is enhanced in human endometriotic stromal cells: A possible mechanism underlying the pathogenesis of endometriosis-associated fibrosis. Hum. Reprod. 2007, 22, 938–944. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, H.; Lee, D.-S.; Park, S.K.; Choi, J.S.; Jung, W.-K.; Park, W.S.; Choi, I.-W. Fucoxanthin Inhibits Myofibroblast Differentiation and Extracellular Matrix Production in Nasal Polyp-Derived Fibroblasts via Modulation of Smad-Dependent and Smad-Independent Signaling Pathways. Mar. Drugs 2018, 16, 323. https://doi.org/10.3390/md16090323
Jung H, Lee D-S, Park SK, Choi JS, Jung W-K, Park WS, Choi I-W. Fucoxanthin Inhibits Myofibroblast Differentiation and Extracellular Matrix Production in Nasal Polyp-Derived Fibroblasts via Modulation of Smad-Dependent and Smad-Independent Signaling Pathways. Marine Drugs. 2018; 16(9):323. https://doi.org/10.3390/md16090323
Chicago/Turabian StyleJung, Hyun, Dae-Sung Lee, Seong Kook Park, Jung Sik Choi, Won-Kyo Jung, Won Sun Park, and Il-Whan Choi. 2018. "Fucoxanthin Inhibits Myofibroblast Differentiation and Extracellular Matrix Production in Nasal Polyp-Derived Fibroblasts via Modulation of Smad-Dependent and Smad-Independent Signaling Pathways" Marine Drugs 16, no. 9: 323. https://doi.org/10.3390/md16090323
APA StyleJung, H., Lee, D.-S., Park, S. K., Choi, J. S., Jung, W.-K., Park, W. S., & Choi, I.-W. (2018). Fucoxanthin Inhibits Myofibroblast Differentiation and Extracellular Matrix Production in Nasal Polyp-Derived Fibroblasts via Modulation of Smad-Dependent and Smad-Independent Signaling Pathways. Marine Drugs, 16(9), 323. https://doi.org/10.3390/md16090323