Biotechnological and Pharmacological Applications of Biotoxins and Other Bioactive Molecules from Dinoflagellates




Abstract
1. Introduction
2. Characterization of Main Dinoflagellate Bioactive (Potential) Applications
2.1. Saxitoxin (and Analogues)
2.2. Tetrodotoxin
2.3. Okadaic Acid and Dinophysistoxin
2.4. Yessotoxin
2.5. Pectenotoxin
2.6. Ciguatoxin
2.7. Maitotoxin
2.8. Palytoxin and Ostreocin
2.9. Gambierol
2.10. Brevetoxin
2.11. Azaspiracid
2.12. Gymnocin
2.13. Karlotoxin
2.14. Cyclic Imine Toxins (Spirolide and Gymonodimine)
2.15. Gambieric Acid (Bioactive)
2.16. Goniodomin A (Bioactive)
2.17. Amphidinolide (Bioactive)
2.18. Amphidinol (Bioactive)
3. Biotoxin and Bioactive Production
3.1. Dinoflagellate Bioactive Supply
3.2. Culture of Dinoflagellates and Biotoxin Production
3.2.1. Nutritional Requirements
3.2.2. Culture Light Provision
3.2.3. Bioreactor Culture and Design
4. Final Considerations
Acknowledgments
Conflicts of Interest
References
- Glaser, K.B.; Mayer, A.M.S. A renaissance in marine pharmacology: From preclinical curiosity to clinical reality. Biochem. Pharmacol. 2009, 78, 440–448. [Google Scholar] [CrossRef] [PubMed]
- De Jesus Raposo, M.F.; De Morais, A.M.B.; De Morais, R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; García-Camacho, F.; López-rosales, L.; Chisti, Y.; Molina-Grima, E. Bioactives from microalgal dinoflagellates. Biotechnol. Adv. 2012, 30, 1673–1684. [Google Scholar] [CrossRef] [PubMed]
- Taylor, F.J.R. Dinoflagellates: An introduction. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell Scientific Publications: Oxford, UK, 1987; pp. 1–13. [Google Scholar]
- Gaines, G.; Elbrachter, M. Heterotrophic nutrition. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell Scientific Publications: Oxford, UK, 1987; pp. 224–247. [Google Scholar]
- Bralewska, J.M.; Witek, Z. Heterotrophic dinoflagellates in the ecosystem of the Gulf of Gdansk. Mar. Ecol. Prog. Ser. 1995, 117, 241–248. [Google Scholar] [CrossRef]
- Hickman, C.P.; Roberts, L.S.; Larson, A.L. Integrated Principles of Zoology; McGraw-Hill International Edition: New York, NY, USA, 2008; pp. 213–222. [Google Scholar]
- Coffroth, M.; Santos, S. Genetic Diversity of Symbiotic Dinoflagellates in the Genus Symbiodinium. Protist 2005, 156, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-Z. Neurotoxins from Marine Dinoflagellates: A Brief Review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.D. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Marine Biotoxins. In FAO Food and Nutritrion; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004; p. 278. [Google Scholar]
- Smayda, T.J. What is a bloom? A commentary. Limnol. Oceanogr. 1997, 42, 1132–1136. [Google Scholar] [CrossRef]
- Bower, D.J.; Hart, R.J.; Matthews, P.A.; Howden, M.E.H. Nonprotein Neurotoxins. Clin. Toxicol. 1981, 18, 813–863. [Google Scholar] [CrossRef] [PubMed]
- Moestrup, Ø.; Akselmann, R.; Fraga, S.; Hoppenrath, M.; Iwataki, M.; Komárek, J.; Larsen, J.; Lundholm, N.; Zingone, A. IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae (HABs). Available online: http://www.marinespecies.org/hab/ (accessed on 28 October 2017).
- Ciminiello, P.; Fattorusso, E. Bivalve Molluscs as Vectors of Marine Biotoxins Involved in Seafood Poisoning. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 53–82. [Google Scholar]
- Quilliam, M.A. Chemical methods for lipophilic shellfish toxins. In Manual on Harmful Marine Microalgae; Hallegraeff, G., Anderson, D., Cembella, A., Eds.; Intergovernmental Oceanographic Comission (UNESCO): Paris, France, 2003; pp. 211–246. ISBN 92-3-103948-2. [Google Scholar]
- FAO (Food and Agriculture Organization)/WHO (World Health Organization). Toxicity Equivalency Factors for Marine Biotoxins Associated with Bivalve Molluscs; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; ISBN 978-92-5-109345-0. [Google Scholar]
- Kellmann, R.; Stüken, A.; Orr, R.J.S.; Svendsen, H.M.; Jakobsen, K.S. Biosynthesis and molecular genetics of polyketides in marine dinoflagellates. Mar. Drugs 2010, 8, 1011–1048. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, S.P.J.N.; Ahmed, N.; Fichtali, J. Nutraceuticals and Bioactives from Marine Algae. In Handbook of Seafood Quality, Safety and Health Applications; Wiley-Blackwell: Oxford, UK, 2010; pp. 455–463. ISBN 9781444325546. [Google Scholar]
- Sugawara, T.; Yamashita, K.; Sakai, S.; Asai, A.; Nagao, A.; Shiraishi, T.; Imai, I.; Hirata, T. Induction of Apoptosis in DLD-1 Human Colon Cancer Cells by Peridinin Isolated from the Dinoflagellate, Heterocapsa triquetra. Biosci. Biotechnol. Biochem. 2007, 71, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S. Natural guanidine derivatives. Nat. Prod. Rep. 2002, 19, 617–649. [Google Scholar] [CrossRef] [PubMed]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Marine biotoxins in shellfish Marine biotoxins in shellfish—Saxitoxin group. Scientific Opinion of the Panel on Contaminants in the Food chain. EFSA J. 2009, 1109, 1–47. [Google Scholar] [CrossRef]
- Durán-Riveroll, L.M.; Cembella, A.D.; Band-Schmidt, C.J.; Bustillos-Guzmán, J.J.; Correa-Basurto, J. Docking Simulation of the Binding Interactions of Saxitoxin Analogs Produced by the Marine Dinoflagellate Gymnodinium catenatum to the Voltage-Gated Sodium Channel Nav1.4. Toxins 2016, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, K.; Musgrave, I.F.; Humpage, A. Low dose extended exposure to saxitoxin and its potential neurodevelopmental effects: A review. Environ. Toxicol. Pharmacol. 2016, 48, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Thomas, K.; Gibbs, R.; Murphy, C.; Quilliam, M.A. Acute toxicities of saxitoxin, neosaxitoxin, decarbamoyl saxitoxin and gonyautoxins 1&4 and 2&3 to mice by various routes of administration. Toxicon 2013, 76, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Epstein-Barash, H.; Shichor, I.; Kwon, A.H.; Hall, S.; Lawlor, M.W.; Langer, R.; Kohane, D.S. Prolonged duration local anesthesia with minimal toxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 7125–7130. [Google Scholar] [CrossRef] [PubMed]
- Kohane, D.S.; Lu, N.T.; Gökgöl-Kline, A.C.; Shubina, M.; Kuang, Y.; Hall, S.; Strichartz, G.R.; Berde, C.B. The local anesthetic properties and toxicity of saxitonin homologues for rat sciatic nerve block in vivo. Reg. Anesth. Pain Med. 2000, 25, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Microalgae as sources of pharmaceuticals and other biologically active compounds. J. Appl. Phycol. 1995, 7, 3–15. [Google Scholar] [CrossRef]
- Garrido, R.; Lagos, N.; Lattes, K.; Abedrapo, M.; Bocic, G.; Cuneo, A.; Chiong, H.; Jensen, C.; Azolas, R.; Henriquez, A.; et al. Gonyautoxin: New treatment for healing acute and chronic anal fissures. Dis. Colon Rectum 2005, 48, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Lattes, K.; Venegas, P.; Lagos, N.; Lagos, M.; Pedraza, L.; Rodriguez-Navarro, A.; García, C. Local infiltration of gonyautoxin is safe and effective in treatment of chronic tension-type headache. Neurol. Res. 2009, 31, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Moczydlowski, E.G. The molecular mystique of tetrodotoxin. Toxicon 2013, 63, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Sato, S.; Sakamoto, S.; Ogata, T. Occurrence of tetrodotoxin in Alexandrium tamarense, a causative dinoflagellate of paralytic shellfish poisoning. Toxicon 1996, 34, 1101–1105. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, Toxicity, Source, Distribution and Detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.R.; Novick, P.A.; Parsons, W.H.; Mcgregor, M.; Zablocki, J.; Pande, V.S.; Bois, J.D. Marked difference in saxitoxin and tetrodoxin affinity for the human nociceptive voltage-gated sodium. Proc. Natl. Acad. Sci. USA 2012, 109, 18102–18107. [Google Scholar] [CrossRef] [PubMed]
- Chong, H.L.; Ruben, P.C. Interaction between voltage-gated sodium channels and the neurotoxin, tetrodotoxin. Channels 2008, 2, 407–412. [Google Scholar] [CrossRef]
- Lago, J.; Rodriguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an extremely potent marine neurotoxin: Distribution, toxicity, origin and therapeutical uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef] [PubMed]
- Nieto, F.R.; Cobos, E.J.; Tejada, M.Á.; Sánchez-Fernández, C.; González-Cano, R.; Cendán, C.M. Tetrodotoxin (TTX) as a therapeutic agent for pain. Mar. Drugs 2012, 10, 281–305. [Google Scholar] [CrossRef] [PubMed]
- Hagen, N.A.; Fisher, K.M.; Lapointe, B.; Souich, P.; Chary, S.; Moulin, D. An Open-Label, Multi-Dose Efficacy and Safety Study of Intramuscular Tetrodotoxin in Patients with Severe. J. Pain Symptom Manag. 2007, 34, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Hagen, N.A.; Souich, P.; Lapointe, B.; Ong-lam, M.; Dubuc, B.; Walde, D. Tetrodotoxin for Moderate to Severe Cancer Pain: A Randomized, Double Blind, Parallel Design Multicenter Study. J. Pain Symptom Manag. 2008, 35, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Berde, C.B.; Athiraman, U.; Yahalom, B.; Zurakowski, D.; Corfas, G.; Bognet, C. Tetrodotoxin-bupivacaine-epinephrine combinations for prolonged local anesthesia. Mar. Drugs 2011, 9, 2717–2728. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Li, J.; Lu, C.L.; Kang, L.; Xie, L.; Zhang, Y.Y.; Zhou, X.B.; Zhong, S. Tetrodotoxin alleviates acute heroin withdrawal syndrome: A multicentre, randomized, double-blind, placebo-controlled study. Clin. Exp. Pharmacol. Physiol. 2011, 38, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Benford, D.; Cockburn, A.; Dogliotti, E.; Di Domenico, A.; Fernández-Cruz, M.L.; Fürst, P.; Galli, C.; Grandjean, P.; Gzyl, J.; et al. Scientific Opinion of the Panel on Contaminants in the Food chain on a request from the European Commission on marine biotoxins in shellfish—Okadaic acid and analogues. EFSA J. 2008, 589, 1–62. [Google Scholar] [CrossRef]
- Cetinkaya, F.; Mus, T.E. Shellfish Poisoning and Toxins. J. Biol. Environ. Sci. 2012, 6, 115–119. [Google Scholar]
- Hallegraeff, G.F. Harmful algal blooms: A global overview. In Manual of Harmful Marine Microalgae; Hallegraeff, G.F., Anderson, D.M., Cembella, A.D., Eds.; UNESCO Publishing: Paris, France, 2003; pp. 25–45. ISBN 9231039482. [Google Scholar]
- Wera, S.; Hemmingst, B.A. Serine/threonine protein phosphatases. Biochem. J. 1995, 311, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Martín-López, A.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; García-Camacho, F.; Molina-Grima, E. Cytotoxicity of yessotoxin and okadaic acid in mouse T lymphocyte cell line EL-4. Toxicon 2012, 60, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Cruz, P.G.; Norte, M.; Creus, A.H.; Fernández, J.J.; Daranas, A.H. Self-association of okadaic acid: Structural and pharmacological significance. Mar. Drugs 2013, 11, 1866–1877. [Google Scholar] [CrossRef] [PubMed]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Paśaro, E.; Meńdez, J.; Laffon, B. Okadaic Acid: More than a diarrheic toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed]
- Thévenin, C.; Kim, S.J.; Kehrl, J.H. Inhibition of protein phosphatases by okadaic acid induces AP1 in human T cells. J. Biol. Chem. 1991, 266, 9363–9366. [Google Scholar] [PubMed]
- Kamat, P.K.; Rai, S.; Nath, C. Okadaic acid induced neurotoxicity: An emerging tool to study Alzheimer’s disease pathology. Neurotoxicology 2013, 37, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Kamat, P.K.; Tota, S.; Rai, S.; Swarnkar, S.; Shukla, R.; Nath, C. A study on neuroinflammatory marker in brain areas of okadaic acid (ICV) induced memory impaired rats. Life Sci. 2012, 90, 713–720. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Yang, Y.; Xu, H.; Zhang, X.; Li, X.-M. Olanzapine attenuates the okadaic acid-induced spatial memory impairment and hippocampal cell death in rats. Neuropsychopharmacology 2005, 30, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.; Götz, J. A local insult of okadaic acid in wild-type mice induces tau phosphorylation and protein aggregation in anatomically distinct brain regions. Acta Neuropathol. Commun. 2016, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.; Rajasekar, N.; Hanif, K.; Nath, C.; Shukla, R. Sulforaphane Ameliorates Okadaic Acid-Induced Memory Impairment in Rats by Activating the Nrf2/HO-1 Antioxidant Pathway. Mol. Neurobiol. 2016, 53, 5310–5323. [Google Scholar] [CrossRef] [PubMed]
- Re, F.; Braaten, D.; Franke, E.K.; Luban, J. Human immunodeficiency virus type 1 Vpr arrests the cell cycle in G2 by inhibiting the activation of p34cdc2-cyclin B. J. Virol. 1995, 69, 6859–6864. [Google Scholar] [PubMed]
- Liu, J.; Sidell, N. Anti-estrogenic effects of conjugated linoleic acid through modulation of estrogen receptor phosphorylation. Breast Cancer Res. Treat. 2005, 94, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Obanda, D.N.; Ribnicky, D.; Yu, Y.; Stephens, J.; Cefalu, W.T. An extract of Urtica dioica L. mitigates obesity induced insulin resistance in mice skeletal muscle via protein phosphatase 2A (PP2A). Sci. Rep. 2016, 6, 22222. [Google Scholar] [CrossRef] [PubMed]
- López, A.M.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; García-Camacho, F.; Molina-Grima, E. Immunoregulatory potential of marine algal toxins yessotoxin and okadaic acid in mouse T lymphocyte cell line EL-4. Toxicol. Lett. 2011, 207, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, Y.; Kasahara, T.; Yamaguehi, Y.; Kuno, K.; Matsushima, K.; Mukaida, N. Stimulation of interleukin-8 production by okadaic acid and vanadate in a human promyelocyte cell line, an HL-60 subline: Possible role of mitogen-activated protein kinase on the okadaic acid-induced NF-κB activation. J. Biol. Chem. 1997, 272, 15366–15372. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Ahn, K.; Kim, S.; Jeong, J. Okadaic acid promotes angiogenesis via activation of hypoxia-inducible factor-1. Cancer Lett. 2009, 276, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Satake, M.; Yasumoto, T. Antimicrobial activities of polyether compounds of dinoflagellate origins. J. Appl. Phycol. 1990, 2, 305–308. [Google Scholar] [CrossRef]
- Gerssen, A.; Pol-Hofstad, I.E.; Poelman, M.; Mulder, P.P.J.; van den Top, H.J.; Dde Boer, J. Marine toxins: Chemistry, toxicity, occurrence and detection, with special reference to the dutch situation. Toxins 2010, 2, 878–904. [Google Scholar] [CrossRef] [PubMed]
- Paz, B.; Riobó, P.; Luisa Fernández, M.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.; McNabb, P.; de Salas, M.; Briggs, L.; Beuzenberg, V.; Gladstone, M. Yessotoxin production by Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar] [CrossRef]
- Miles, C.O.; Samdal, I.A.; Aasen, J.A.G.; Jensen, D.J.; Quilliam, M.A.; Petersen, D.; Briggs, L.M.; Wilkins, A.L.; Rise, F.; Cooney, J.M.; et al. Evidence for numerous analogs of yessotoxin in Protoceratium reticulatum. Harmful Algae 2005, 4, 1075–1091. [Google Scholar] [CrossRef]
- Murata, M.; Kumagai, M.; Lee, J.S.; Yasumoto, T. Isolation and structure of yessotoxin, a novel polyether compound implicated in diarrhetic shellfish poisoning. Tetrahedron Lett. 1987, 28, 5869–5872. [Google Scholar] [CrossRef]
- Paz, B.; Daranas, A.H.; Norte, M.; Riobó, P.; Franco, J.M.; Fernández, J.J. Yessotoxins, a group of marine polyether toxins: An overview. Mar. Drugs 2008, 6, 73–102. [Google Scholar] [CrossRef] [PubMed]
- Korsnes, M.S.; Røed, S.S.; Tranulis, M.A.; Espenes, A.; Christophersen, B. Yessotoxin triggers ribotoxic stress. Toxicol. In Vitro 2014, 28, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Suárez Korsnes, M.; Espenes, A. Yessotoxin as an apoptotic inducer. Toxicon 2011, 57, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, A.; Vieytes, M.; Botana, L. Yessotoxin, a Promising Therapeutic Tool. Mar. Drugs 2016, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Araujo, A.; Alfonso, A.; Vieytes, M.R.; Botana, L.M. Key role of phosphodiesterase 4A (PDE4A) in autophagy triggered by yessotoxin. Toxicology 2015, 329, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, A.; de la Rosa, L.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. Yessotoxin, a novel phycotoxin, activates phosphodiesterase activity: Effect of yessotoxin on cAMP levels in human lymphocytes. Biochem. Pharmacol. 2003, 65, 193–208. [Google Scholar] [CrossRef]
- Jeon, Y.H.; Heo, Y.-S.; Kim, C.M.; Hyun, Y.-L.; Lee, T.G.; Ro, S.; Cho, J.M. Phosphodiesterase: Overview of protein structures, potential therapeutic applications and recent progress in drug development. Cell. Mol. Life Sci. 2005, 62, 1198–1220. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, L.A.; Alfonso, A.; Vilariño, N.; Vieytes, M.R.; Botana, L.M. Modulation of cytosolic calcium levels of human lymphocytes by yessotoxin, a novel marine phycotoxin. Biochem. Pharmacol. 2001, 61, 827–833. [Google Scholar] [CrossRef]
- Korsnes, M.S.; Hetland, D.L.; Espenes, A.; Tranulis, M.A.; Aune, T. Apoptotic events induced by yessotoxin in myoblast cell lines from rat and mouse. Toxicol. In Vitro 2006, 20, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, C.; Fato, R.; Angelin, A.; Trombetti, F.; Ventrella, V.; Borgatti, A.R.; Fattorusso, E.; Ciminiello, P.; Bernardi, P.; Lenaz, G.; et al. Yessotoxin, a shellfish biotoxin, is a potent inducer of the permeability transition in isolated mitochondria and intact cells. Biochim. Biophys. Acta Bioenerg. 2004, 1656, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Orsi, C.F.; Colombari, B.; Callegari, F.; Todaro, A.M.; Ardizzoni, A.; Rossini, G.P.; Blasi, E.; Peppoloni, S. Yessotoxin inhibits phagocytic activity of macrophages. Toxicon 2010, 55, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Korsnes, M.S.; Hetland, D.L.; Espenes, A.; Aune, T. Cleavage of tensin during cytoskeleton disruption in YTX-induced apoptosis. Toxicol. In Vitro 2007, 21, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Stemmler, M.P. Cadherins in development and cancer. Mol. Biosyst. 2008, 4, 835–850. [Google Scholar] [CrossRef] [PubMed]
- Tubaro, A.; Dell’Ovo, V.; Sosa, S.; Florio, C. Yessotoxins: A toxicological overview. Toxicon 2010, 56, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Korsnes, M.S.; Korsnes, R. Mitotic Catastrophe in BC3H1 Cells following Yessotoxin Exposure. Front. Cell Dev. Biol. 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Tobío, A.; Alfonso, A.; Madera-Salcedo, I.; Botana, L.M.; Blank, U. Yessotoxin, a Marine Toxin, Exhibits Anti-Allergic and Anti-Tumoural Activities Inhibiting Melanoma Tumour Growth in a Preclinical Model. PLoS ONE 2016, 11, e0167572. [Google Scholar] [CrossRef] [PubMed]
- Bloom, G.S. Amyloid-β and Tau The Trigger and Bullet in Alzheimer Disease Pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Terao, K.; Ito, E.; Oarada, M.; Murata, M.; Yasumoto, T. Histopathological studies on experimental marine toxin poisoning—5. The effects in mice of yessotoxin isolated from Patinopecten yessoensis and of a desulfated derivative. Toxicon 1990, 28, 1095–1104. [Google Scholar] [CrossRef]
- Rubiolo, J.A.; López-Alonso, H.; Martínez, P.; Millán, A.; Cagide, E.; Vieytes, M.R.; Vega, F.V.; Botana, L.M. Yessotoxin induces ER-stress followed by autophagic cell death in glioma cells mediated by mTOR and BNIP3. Cell. Signal. 2014, 26, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; González, V.; Martínez, A.; Paz, B.; Lago, J.; Cordeiro, V.; Blanco, L.; Vieites, J.; Cabado, A. Occurrence of Lipophilic Marine Toxins in Shellfish from Galicia (NW of Spain) and Synergies among Them. Mar. Drugs 2015, 13, 1666–1687. [Google Scholar] [CrossRef] [PubMed]
- Espiña, B.; Louzao, M.C.; Ares, I.R.; Fonfría, E.S.; Vilariño, N.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. Impact of the pectenotoxin C-43 oxidation degree on its cytotoxic effect on rat hepatocytes. Chem. Res. Toxicol. 2010, 23, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Allingham, J.S.; Miles, C.O.; Rayment, I. A Structural Basis for Regulation of Actin Polymerization by Pectenotoxins. J. Mol. Biol. 2007, 371, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Espiña, B.; Rubiolo, J.A. Marine toxins and the cytoskeleton: Pectenotoxins, unusual macrolides that disrupt actin. FEBS J. 2008, 275, 6082–6088. [Google Scholar] [CrossRef] [PubMed]
- Ares, I.R.; Louzao, M.C.; Espiña, B.; Vieytes, M.R.; Miles, C.O.; Yasumoto, T.; Botana, L.M. Lactone ring of pectenotoxins: A key factor for their activity on cytoskeletal dynamics. Cell. Physiol. Biochem. 2007, 19, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O.; Kim, M.-O.; Nam, T.-J.; Kim, S.-K.; Choi, Y.H.; Kim, G.-Y. Esculetin inhibits cell proliferation through the Ras/ERK1/2 pathway in human colon cancer cells. Oncol. Rep. 2011, 25, 223–230. [Google Scholar] [CrossRef]
- Jung, J.H.; Sim, C.J.; Lee, C.O. Cytotoxic compounds from a two-sponge association. J. Nat. Prod. 1995, 58, 1722–1726. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.-D.; Choi, T.-S.; Kim, B.-M.; Jung, J.H.; Bang, Y.-J.; Shin, D.Y. Oocyte-based screening of cytokinesis inhibitors and identification of pectenotoxin-2 that induces Bim/Bax-mediated apoptosis in p53-deficient tumors. Oncogene 2005, 24, 4813–4819. [Google Scholar] [CrossRef] [PubMed]
- García-Camacho, F.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Cerón-García, M.C.; Belarbi, E.H.; Chisti, Y.; Grima-Molina, E. Biotechnological significance of toxic marine dinoflagellates. Biotechnol. Adv. 2007, 25, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M. The origin of ciguatera—An update. Ciguatera Inf. Bull. 1992, 2, 8–9. [Google Scholar]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Crump, J.A.; McLay, C.L.; Chambers, S.T. Ciguatera fish poisoning. Postgrad. Med. J. 1999, 75, 678–679. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Holmes, M.J. Origin and transfer of toxins involved in ciguatera. Comp. Biochem. Physiol. C 1993, 106, 615–628. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Scheuer, P.J.; Takahashi, W.; Tsutsumi, J.; Yoshida, T. Ciguatoxin: Isolation and chemical nature. Science 1967, 155, 1267–1268. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic polyether modulators of voltage-gated ion channel function. Mar. Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef]
- Hogg, R.C.; Lewis, R.J.; Adams, D.J. Ciguatoxin-induced oscillations in membrane potential and action potential firing in rat parasympathetic neurons. Eur. J. Neurosci. 2002, 16, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Lombet, A.; Bidard, J.N.; Lazdunski, M. Ciguatoxin and brevetoxins share a common receptor site on the neuronal voltage-dependent Na+ channel. FEBS Lett. 1987, 219, 355–359. [Google Scholar] [CrossRef]
- Mattei, C.; Molgó, J.; Benoit, E. Involvement of both sodium influx and potassium efflux in ciguatoxin-induced nodal swelling of frog myelinated axons. Neuropharmacology 2014, 85, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C.; Dechraoui, M.Y.; Molgó, J.; Meunier, F.; Legrand, A.M.; Benoit, E. Neurotoxins targeting receptor site 5 of voltage-dependent sodium channels increase the nodal volume of myelinated axons. J. Neurosci. Res. 1999, 55, 666–673. [Google Scholar] [CrossRef]
- Cameron, J.; Flowers, A.E.; Capra, M.F. Effects of ciguatoxin on nerve excitability in rats (Part I). J. Neurol. Sci. 1991, 101, 87–92. [Google Scholar] [CrossRef]
- Cameron, J.; Flowers, A.E.; Capra, M.F. Electrophysiological studies on ciguatera poisoning in man (Part II). J. Neurol. Sci. 1991, 101, 93–97. [Google Scholar] [CrossRef]
- Molgó, J.; Comella, J.X.; Legrand, A.M. Ciguatoxin enhances quantal transmitter release from frog motor nerve terminals. Br. J. Pharmacol. 1990, 99, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C.; Wen, P.J.; Nguyen-Huu, T.D.; Alvarez, M.; Benoit, E.; Bourdelais, A.J.; Lewis, R.J.; Baden, D.G.; Molgó, J.; Meunier, F.A. Brevenal inhibits pacific ciguatoxin-1B-induced neurosecretion from bovine chromaffin cells. PLoS ONE 2008, 3, e3448. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, K.; Inoue, M.; Miyazaki, K.; Hirama, M.; Kondo, C.; Kinoshita, E.; Miyoshi, H.; Seyama, I. Synthetic ciguatoxins selectively activate Nav1.8-derived chimeric sodium channels expressed in HEK293 cells. J. Biol. Chem. 2009, 284, 7597–7605. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.; Touska, F.; Hess, A.; Hinsbey, R.; Sattler, S.; Lampert, A.; Sergejeva, M.; Sharov, A.; Collins, L.S.; Eberhardt, M.; et al. Ciguatoxins activate specific cold pain pathways to elicit burning pain from cooling. EMBO J. 2012, 31, 3795–3808. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C.; Legros, C. The voltage-gated sodium channel: A major target of marine neurotoxins. Toxicon 2014, 91, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Eijkelkamp, N.; Linley, J.E.; Baker, M.D.; Minett, M.S.; Cregg, R.; Werdehausen, R.; Rugiero, F.; Wood, J.N. Neurological perspectives on voltage-gated sodium channels. Brain 2012, 135, 2585–2612. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Aversa, R.J. Maitotoxin: An inspiration for synthesis. Isr. J. Chem. 2011, 51, 359–377. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.G.; Sánchez-cárdenas, C.; Acevedo-Castillo, W.; Leyton, P.; López-gonzález, I.; Felix, R.; Gandini, M.A.; Treviño, M.B.; Treviño, C.L. Maitotoxin: An Enigmatic Toxic Molecule with Useful Applications in the Biomedical Sciences. In Seafood and Freshwater Toxins Pharmacology, Physiology, and Detection; Botana, L.M., Ed.; CRC Press: London, UK, 2014; pp. 677–694. ISBN 978-1-4665-0515-5. [Google Scholar]
- Holmes, M.J.; Lewis, R.J. Purification and characterisation of large and small maitotoxins from cultured Gambierdiscus toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Murata, M.; Oshima, Y.; Iwashita, T.; Yasumoto, T. Some chemical properties of maitotoxin, a putative calcium channel agonist isolated from a marine dinoflagellate. J. Biochem. 1988, 104, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.; Harwood, T.; Smith, K.; Argyle, P.; Munday, R. Production of ciguatoxin and maitotoxin by strains of Gambierdiscus australes, G. pacificus and G. polynesiensis (Dinophyceae) isolated from Rarotonga, Cook Islands. Harmful Algae 2014, 39, 185–190. [Google Scholar] [CrossRef]
- Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Hambright, K.D.; Zamor, R.M.; Easton, J.D.; Allison, B. Algae. In Encyclopedia of Toxicology; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 130–141. [Google Scholar]
- Konoki, K.; Hashimoto, M.; Murata, M.; Tachibana, K. Maitotoxin-induced calcium influx in erythrocyte ghosts and rat glioma C6 cells, and blockade by gangliosides and other membrane lipids. Chem. Res. Toxicol. 1999, 12, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; Van Dolah, F.M.; Ramsdell, J.S. Maitotoxin induces a calcium-dependent membrane depolarization in GH4C1 pituitary cells via activation of type L voltage-dependent calcium channels. J. Biol. Chem. 1992, 267, 25025–25031. [Google Scholar] [PubMed]
- Soergel, D.G.; Gusovsky, F.; Yasumoto, T.; Daly, J.W. Stimulatory effects of maitotoxin on insulin release in insulinoma HIT cells: Role of calcium uptake and phosphoinositide breakdown. J. Pharmacol. Exp. Ther. 1990, 255, 1360–1365. [Google Scholar] [PubMed]
- Taglialatela, M.; Amoroso, S.; Yasumoto, T.; Di Renzo, G.; Annunziato, L. Maitotoxin and Bay-K-8644: Two putative calcium channel activators with different effects on endogenous dopamine release from tuberoinfundibular neurons. Brain Res. 1986, 381, 356–358. [Google Scholar] [CrossRef]
- Takahashi, M.; Tatsumi, M.; Ohizumi, Y.; Yasumoto, T. Ca2+ channel activating function of maitotoxin, the most potent marine toxin known, in clonal rat pheochromocytoma cells. J. Biol. Chem. 1983, 258, 10944–10949. [Google Scholar] [PubMed]
- Gusovsky, F.; Daly, J.W.; Yasumoto, T.; Rojas, E. Differential effects of maitotoxin on ATP secretion and on phosphoinositide breakdown in rat pheochromocytoma cells. FEBS Lett. 1988, 233, 139–142. [Google Scholar] [CrossRef]
- Estacion, M.; Schilling, W.P. Maitotoxin-induced membrane blebbing and cell death in bovine aortic endothelial cells. BMC Physiol. 2001, 1, 2. [Google Scholar] [CrossRef]
- Chávez, J.C.; de Blas, G.A.; de la Vega-Beltrán, J.L.; Nishigaki, T.; Chirinos, M.; González-González, M.E.; Larrea, F.; Solís, A.; Darszon, A.; Treviño, C.L. The opening of maitotoxin-sensitive calcium channels induces the acrosome reaction in human spermatozoa: Differences from the zona pellucida. Asian J. Androl. 2011, 13, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Treviño, C.L.; de la Vega-Beltrán, J.L.; Nishigaki, T.; Felix, R.; Darszon, A. Maitotoxin potently promotes Ca2+ influx in mouse spermatogenic cells and sperm, and induces the acrosome reaction. J. Cell. Physiol. 2006, 206, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, P.A.; Kertesy, S.B.; Estacion, M.; Schilling, W.P.; Dubyak, G.R. Maitotoxin induces biphasic interleukin-1beta secretion and membrane blebbing in murine macrophages. Mol. Pharmacol. 2004, 66, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Flores, P.; Rodríguez, E.; Zapata, E.; Carbó, R.; Farías, J.; Martínez, M. Maitotoxin Is a Potential Selective Activator of the Endogenous Transient Receptor Potential Canonical Type 1 Channel in Xenopus laevis Oocytes. Mar. Drugs 2017, 15, 198. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H. Pharmacological Actions of Palytoxin. In Toxins and Biologically Active Compounds from Microalgae, Volume 2: Biological Effects and Risk Management; Rossini, P., Ed.; CRC Press/Taylor and Francis: Boca Raton, FL, USA, 2014; p. 663. [Google Scholar]
- Moore, R.E.; Scheuer, P.J. Palytoxin: A new marine toxin from a coelenterate. Science 1971, 172, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Franchini, A.; Malagoli, D.; Ottaviani, E. Targets and effects of yessotoxin, okadaic acid and palytoxin: A differential review. Mar. Drugs 2010, 8, 658–677. [Google Scholar] [CrossRef] [PubMed]
- Ramos, V.; Vasconcelos, V. Palytoxin and analogs: Biological and ecological effects. Mar. Drugs 2010, 8, 2021–2037. [Google Scholar] [CrossRef] [PubMed]
- Patocka, J.; Gupta, R.C.; Wu, Q.H.; Kuca, K. Toxic potential of palytoxin. J. Huazhong Univ. Sci. Technol.—Med. Sci. 2015, 35, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Pelin, M.; Florio, C.; Ponti, C.; Lucafò, M.; Gibellini, D.; Tubaro, A.; Sosa, S. Pro-inflammatory effects of palytoxin: An in vitro study on human keratinocytes and inflammatory cells. Toxicol. Res. 2016, 5, 1172–1181. [Google Scholar] [CrossRef]
- Louzao, M.C.; Ares, I.R.; Cagide, E. Marine toxins and the cytoskeleton: A new view of palytoxin toxicity. FEBS J. 2008, 275, 6067–6074. [Google Scholar] [CrossRef] [PubMed]
- Ares, I.R.; Cagide, E.; Louzao, M.C.; Espiña, B.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. Ostreocin-d Impact on Globular Actin of Intact Cells. Chem. Res. Toxicol. 2009, 22, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Carmen Louzao, M.; Fraga, M.; Vilariño, N. Pharmacology of palytoxins and ostreocins. In Phycotoxins; Botana, L.M., Amparo, A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 113–135. ISBN 9781118500354. [Google Scholar]
- Bigiani, A. Palytoxin action on the Na+, K+-ATPase and the disruption of ion equilibria in biological systems. Toxicon 2011, 57, 429–439. [Google Scholar] [CrossRef]
- Wattenberg, E.V. Palytoxin: Exploiting a novel skin tumor promoter to explore signal transduction and carcinogenesis. Am. J. Physiol. Cell Physiol. 2007, 292, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Murata, M.; Yasumoto, T. Gambierol: A new toxic polyether compound isolated from the marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1993, 115, 361–362. [Google Scholar] [CrossRef]
- Fuwa, H.; Fukazawa, R.; Sasaki, M. Concise synthesis of the A/BCD-ring fragment of gambieric acid A. Front. Chem. 2015, 2, 116. [Google Scholar] [CrossRef] [PubMed]
- Fuwa, H.; Kainuma, N.; Satake, M.; Sasaki, M. Synthesis and biological evaluation of gambierol analogues. Bioorg. Med. Chem. Lett. 2003, 13, 2519–2522. [Google Scholar] [CrossRef]
- Ito, E.; Suzuki-Toyota, F.; Toshimori, K.; Fuwa, H.; Tachibana, K.; Satake, M.; Sasaki, M. Pathological effects on mice by gambierol, possibly one of the ciguatera toxins. Toxicon 2003, 42, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.; Rubiolo, J.A. Therapeutics of marine toxins. In Phycotoxins: Chemistry and Biochemistry; Botana, L.M., Alfonso, A., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 2015; pp. 181–195. ISBN 9781118500347. [Google Scholar]
- Cao, Z.; George, J.; Gerwick, W.H.; Baden, D.G.; Rainier, J.D.; Murray, T.F. Influence of lipid-soluble gating modifier toxins on sodium influx in neocortical neurons. J. Pharmacol. Exp. Ther. 2008, 326, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, E.; Abdel-Mottaleb, Y.; Kopljar, I.; Rainier, J.D.; Raes, A.L.; Snyders, D.J.; Tytgat, J. Gambierol, a toxin produced by the dinoflagellate Gambierdiscus toxicus, is a potent blocker of voltage-gated potassium channels. Toxicon 2008, 51, 974–983. [Google Scholar] [CrossRef] [PubMed]
- Ghiaroni, V.; Sasaki, M.; Fuwa, H.; Rossini, G.P.; Scalera, G.; Yasumoto, T.; Pietra, P.; Bigiani, A. Inhibition of voltage-gated potassium currents by gambierol in mouse taste cells. Toxicol. Sci. 2005, 85, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Kopljar, I.; Labro, A.J.; de Block, T.; Rainier, J.D.; Tytgat, J.; Snyders, D.J. The ladder-shaped polyether toxin gambierol anchors the gating machinery of Kv3.1 channels in the resting state. J. Gen. Physiol. 2013, 141, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Cui, Y.; Busse, E.; Mehrotra, S.; Rainier, J.D.; Murray, T.F. Gambierol inhibition of voltage-gated potassium channels augments spontaneous Ca2+ oscillations in cerebrocortical neurons. J. Pharmacol. Exp. Ther. 2014, 350, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Pérez, S.; Vale, C.; Alonso, E.; Fuwa, H.; Sasaki, M.; Konno, Y.; Goto, T.; Suga, Y.; Vieytes, M.R.; Botana, L.M. Effect of Gambierol and Its Tetracyclic and Heptacyclic Analogues in Cultured Cerebellar Neurons: A Structure–Activity Relationships Study. Chem. Res. Toxicol. 2012, 25, 1929–1937. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.; Vale, C.; Sasaki, M.; Fuwa, H.; Konno, Y.; Perez, S.; Vieytes, M.R.; Botana, L.M. Calcium oscillations induced by gambierol in cerebellar granule cells. J. Cell. Biochem. 2010, 110, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Huang, J.; Yuan, X.; Peng, B.; Liu, W.; Han, S.; He, X. Toxins Targeting the Kv1.3 Channel: Potential Immunomodulators for Autoimmune Diseases. Toxins 2015, 7, 1749–1764. [Google Scholar] [CrossRef] [PubMed]
- Rubiolo, J.A.; Vale, C.; Martín, V.; Fuwa, H.; Sasaki, M.; Botana, L.M. Potassium currents inhibition by gambierol analogs prevents human T lymphocyte activation. Arch. Toxicol. 2015, 89, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Beeton, C.; Wulff, H.; Standifer, N.E.; Azam, P.; Mullen, K.M.; Pennington, M.W.; Kolski-Andreaco, A.; Wei, E.; Grino, A.; Counts, D.R.; et al. Kv1.3 channels are a therapeutic target for T cell-mediated autoimmune diseases. Proc. Natl. Acad. Sci. USA 2006, 103, 17414–17419. [Google Scholar] [CrossRef] [PubMed]
- LePage, K.T.; Rainier, J.D.; Johnson, H.W.B.; Baden, D.G.; Murray, T.F. Gambierol acts as a functional antagonist of neurotoxin site 5 on voltage-gated sodium channels in cerebellar granule neurons. J. Pharmacol. Exp. Ther. 2007, 323, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Baden, D.G. Brevetoxins: Unique polyether dinoflagellate toxins. FASEB J. 1989, 3, 1807–1817. [Google Scholar] [PubMed]
- Plakas, S.M.; Dickey, R.W. Advances in monitoring and toxicity assessment of brevetoxins in molluscan shellfish. Toxicon 2010, 56, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Xu, J.; Tsang, T.Y.; Au, D.W.T. Toxicity comparison between Chattonella marina and Karenia brevis using marine medaka (Oryzias melastigma): Evidence against the suspected ichthyotoxins of Chattonella marina. Chemosphere 2010, 80, 585–591. [Google Scholar] [CrossRef] [PubMed]
- De Boer, M.K.; Tyl, M.R.; Fu, M.; Kulk, G.; Liebezeit, G.; Tomas, C.R.; Lenzi, A.; Naar, J.; Vrieling, E.G.; van Rijssel, M. Haemolytic activity within the species Fibrocapsa japonica (Raphidophyceae). Harmful Algae 2009, 8, 699–705. [Google Scholar] [CrossRef]
- Khan, S.; Arakawa, O.; Onoue, Y. Neurotoxins in a toxic red tide of Heterosigma akashiwo (Raphidophyceae) in Kagoshima Bay, Japan. Aquac. Res. 1997, 28, 9–14. [Google Scholar] [CrossRef]
- Baden, D.G.; Bourdelais, A.J.; Jacocks, H.; Michelliza, S.; Naar, J. Natural and derivative brevetoxins: Historical background, multiplicity, and effects. Environ. Health Perspect. 2005, 113, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Michelliza, S.; Abraham, W.M.; Jacocks, H.M.; Schuster, T.; Baden, D.G. Synthesis, modeling, and biological evaluation of analogues of the semisynthetic brevetoxin antagonist beta-naphthoyl-brevetoxin. Chembiochem 2007, 8, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Benson, J.; Hahn, F.; March, T.; McDonald, J.; Sopori, M.; Seagrave, J.; Gomez, A.; Bourdelais, A.; Naar, J.; Zaias, J.; et al. Inhalation toxicity of brevetoxin 3 in rats exposed for 5 days. J. Toxicol. Environ. Health A 2004, 67, 1443–1456. [Google Scholar] [CrossRef] [PubMed]
- Poli, M.A.; Mende, T.J.; Baden, D.G. Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptosomes. Mol. Pharmacol. 1986, 30, 129–135. [Google Scholar] [PubMed]
- Catterall, W.A.; Gainer, M. Interaction of brevetoxin A with a new receptor site on the sodium channel. Toxicon 1985, 23, 497–504. [Google Scholar] [CrossRef]
- LePage, K.T.; Baden, D.G.; Murray, T.F. Brevetoxin derivatives act as partial agonists at neurotoxin site 5 on the voltage-gated Na+ channel. Brain Res. 2003, 959, 120–127. [Google Scholar] [CrossRef]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Jeglitsch, G.; Rein, K.; Baden, D.G.; Adams, D.J. Brevetoxin-3 (PbTx-3) and its derivatives modulate single tetrodotoxin-sensitive sodium channels in rat sensory neurons. J. Pharmacol. Exp. Ther. 1998, 284, 516–525. [Google Scholar] [PubMed]
- Gawley, R.E.; Rein, K.S.; Jeglitsch, G.; Adams, D.J.; Theodorakis, E.A.; Tiebes, J.; Nicolaou, K.C.; Baden, D.G. The relationship of brevetoxin “length” and A-ring functionality to binding and activity in neuronal sodium channels. Chem. Biol. 1995, 2, 533–541. [Google Scholar] [CrossRef]
- Joseph, A. Oceans: Abode of Nutraceuticals, Pharmaceuticals, and Biotoxins. In Investigating Seafloors and Oceans; Joseph, A., Ed.; Candice Janco: Goa, India, 2016; pp. 493–554. ISBN 9780128093573. [Google Scholar]
- Murrell, R.N.; Gibson, J.E. Brevetoxins 2, 3, 6, and 9 show variability in potency and cause significant induction of DNA damage and apoptosis in Jurkat E6-1 cells. Arch. Toxicol. 2009, 83, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, T.; Krzanowski, J.; Nelson, R.; Martin, D.F.; Polson, J.; Duncan, R.; Lockey, R. In vitro red tide toxin effects on human bronchial smooth muscle. J. Allergy Clin. Immunol. 1988, 81, 1187–1191. [Google Scholar] [CrossRef]
- Abraham, W.M.; Bourdelais, A.J.; Sabater, J.R.; Ahmed, A.; Lee, T.A.; Serebriakov, I.; Baden, D.G. Airway responses to aerosolized brevetoxins in an animal model of asthma. Am. J. Respir. Crit. Care Med. 2005, 171, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Hilderbrand, S.C.; Murrell, R.N.; Gibson, J.E.; Brown, J.M. Marine brevetoxin induces ige-independent mast cell activation. Arch. Toxicol. 2011, 85, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Sas, K.M.; Baatz, J.E. Brevetoxin-2 induces an inflammatory response in an alveolar macrophage cell line. Int. J. Hyg. Environ. Health 2010, 213, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.J.; Leggett, S.R.; Strohbehn, K.; Pierce, R.H.; Sleasman, J.W. Effects of in vitro brevetoxin exposure on apoptosis and cellular metabolism in a leukemic T cell line (Jurkat). Mar. Drugs 2008, 6, 291–307. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Baden, D.G.; Gerwick, W.H.; Murray, T.F. Bidirectional in fl uence of sodium channel activation on NMDA receptor–dependent cerebrocortical neuron structural plasticity. Proc. Natl. Acad. Sci. USA 2012, 109, 19840–19845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xu, X.; Li, T.; Liu, Z. Shellfish Toxins Targeting Voltage-Gated Sodium Channels. Mar. Drugs 2013, 11, 4698–4723. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U.; Elbrächter, M.; Krock, B.; John, U.; Cembella, A. Azadinium spinosum gen. et sp. nov. (Dinophyceae) identified as a primary producer of azaspiracid toxins. Eur. J. Phycol. 2009, 44, 63–79. [Google Scholar] [CrossRef]
- Percopo, I.; Siano, R.; Rossi, R.; Soprano, V.; Sarno, D.; Zingone, A. A new potentially toxic Azadinium species (Dinophyceae) from the Mediterranean Sea, A. dexteroporum sp. nov. J. Phycol. 2013, 49, 950–966. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.; Tillmann, U.; Potvin, É.; Jeong, H.J.; Drebing, W.; Kilcoyne, J.; Al-Jorani, A.; Twiner, M.J.; Gothel, Q.; Kock, M. Structure elucidation and in vitro toxicity of new azaspiracids isolated from the marine dinoflagellate Azadinium poporum. Mar. Drugs 2015, 13, 6687–6702. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; Mccarron, P.; Krock, B.; Miles, C.O. Azaspiracids: Chemistry, Biosynthesis, Metabolism, and Detection. In Seafood and Freshwater Toxins; Botana, L.M., Ed.; CRC Press: London, UK, 2014; pp. 799–822. [Google Scholar]
- Busch, J.A.; Andree, K.B.; Diogène, J.; Fernández-Tejedor, M.; Toebe, K.; John, U.; Krock, B.; Tillmann, U.; Cembella, A.D. Toxigenic algae and associated phycotoxins in two coastal embayments in the Ebro Delta (NW Mediterranean). Harmful Algae 2016, 55, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Ofuji, K.; Naoki, H.; James, K.J.; Furey, A.; McMahon, T.; Silke, J.; Yasumoto, T. Azaspiracid, a New Marine Toxin Having Unique Spiro Ring Assemblies, Isolated from Irish Mussels, Mytilus edulis. J. Am. Chem. Soc. 1998, 120, 9967–9968. [Google Scholar] [CrossRef]
- Vilariño, N. Marine toxins and the cytoskeleton: Azaspiracids. FEBS J. 2008, 275, 6075–6081. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Hess, P.; Bottein Dechraoui, M.Y.; McMahon, T.; Samons, M.S.; Satake, M.; Yasumoto, T.; Ramsdell, J.S.; Doucette, G.J. Cytotoxic and cytoskeletal effects of azaspiracid-1 on mammalian cell lines. Toxicon 2005, 45, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Colman, J.R.; Twiner, M.J.; Hess, P.; McMahon, T.; Satake, M.; Yasumoto, T.; Doucette, G.J.; Ramsdell, J.S. Teratogenic effects of azaspiracid-1 identified by microinjection of Japanese medaka (Oryzias latipes) embryos. Toxicon 2005, 45, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; LePage, K.T.; Frederick, M.O.; Nicolaou, K.C.; Murray, T.F. Involvement of Caspase Activation in Azaspiracid-Induced Neurotoxicity in Neocortical Neurons. Toxicol. Sci. 2010, 114, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Vale, C.; Nicolaou, K.C.; Frederick, M.O.; Vieytes, M.R.; Botana, L.M. Cell volume decrease as a link between azaspiracid-induced cytotoxicity and c-Jun-.-terminal kinase activation in cultured neurons. Toxicol. Sci. 2009, 113, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Román, Y.; Alfonso, A.; Vieytes, M.R.; Ofuji, K.; Satake, M.; Yasumoto, T.; Botana, L.M. Effects of Azaspiracids 2 and 3 on Intracellular cAMP, [Ca2+], and pH. Chem. Res. Toxicol. 2004, 17, 1338–1349. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Hanagriff, J.C.; Butler, S.; Madhkoor, A.K.; Doucette, G.J. Induction of Apoptosis Pathways in Several Cell Lines following Exposure to the Marine Algal Toxin Azaspiracid. Chem. Res. Toxicol. 2012, 25, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Ryan, J.C.; Morey, J.S.; Smith, K.J.; Hammad, S.M.; Van Dolah, F.M.; Hess, P.; McMahon, T.; Satake, M.; Yasumoto, T.; et al. Transcriptional profiling and inhibition of cholesterol biosynthesis in human T lymphocyte cells by the marine toxin azaspiracid. Genomics 2008, 91, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Doucette, G.J.; Rasky, A.; Huang, X.-P.; Roth, B.L.; Sanguinetti, M.C. Marine Algal Toxin Azaspiracid Is an Open-State Blocker of hERG Potassium Channels. Chem. Res. Toxicol. 2012, 25, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Shoji, M.; Oshima, Y.; Naoki, H.; Fujita, T.; Yasumoto, T. Gymnocin-A, a cytotoxic polyether from the notorious red tide dinoflagellate, Gymnodinium mikimotoi. Tetrahedron Lett. 2002, 43, 5829–5832. [Google Scholar] [CrossRef]
- Tsukano, C.; Sasaki, M. Structure-activity relationship studies of gymnocin-A. Tetrahedron Lett. 2006, 47, 6803–6807. [Google Scholar] [CrossRef]
- Houdai, T.; Matsuoka, S.; Matsumori, N.; Murata, M. Membrane-permeabilizing activities of amphidinol 3, polyene-polyhydroxy antifungal from a marine dinoflagellate. Biochim. Biophys. Acta-Biomembr. 2004, 1667, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.L.; Hill, R.T.; Place, A.R.; Hamann, M.T. The expanding role of marine microbes in pharmaceutical. Curr. Opin. Biotechnol. 2011, 21, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Adolf, J.E.; Bachvaroff, T.R.; Deeds, J.R.; Place, A.R. Ichthyotoxic Karlodinium veneficum (Ballantine) J Larsen in the Upper Swan River Estuary (Western Australia): Ecological conditions leading to a fish kill. Harmful Algae 2015, 48, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Bachvaroff, T.R.; Adolf, J.E.; Squier, A.H.; Harvey, H.R.; Place, A.R. Characterization and quantification of karlotoxins by liquid chromatography-mass spectrometry. Harmful Algae 2008, 7, 473–484. [Google Scholar] [CrossRef]
- Waters, A.L.; Oh, J.; Place, A.R.; Hamann, M.T. Stereochemical Studies of the Karlotoxin Class Using NMR Spectroscopy and DP4 Chemical-Shift Analysis: Insights into their Mechanism of Action. Angew. Chem. Int. Ed. 2015, 54, 15705–15710. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, R.M.; Deeds, J.R.; Tatters, A.O.; Place, A.R.; Tomas, C.R.; Wright, J.L.C. Structure and relative potency of several karlotoxins from Karlodinium veneficum. J. Nat. Prod. 2010, 73, 1360–1365. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; He, S.; Zhou, C.; Place, A.R.; Haq, S.; Ding, L.; Chen, H.; Jiang, Y.; Guo, C.; Xu, Y.; et al. Two new karlotoxins found in Karlodinium veneficum (strain GM2) from the East China Sea. Harmful Algae 2016, 58, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Place, A.R.; Yoshida, W.; Anklin, C.; Hamann, M.T. Structure and absolute configuration of Karlotoxin-2, an ichthyotoxin from the marine dinoflagellate Karlodinium veneficum. J. Am. Chem. Soc. 2010, 132, 3277–3279. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, R.M.; Deeds, J.R.; Satake, M.; Ribeiro, A.A.; Place, A.R.; Wright, J.L.C. Isolation and characterization of karlotoxin 1, a new amphipathic toxin from Karlodinium veneficum. Tetrahedron Lett. 2008, 49, 6457–6461. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Matsuoka, S.; Matsumori, N.; Paul, G.K.; Tachibana, K. Absolute configuration of amphidinol 3, the first complete structure determination from amphidinol homologues: Application of a new configuration analysis based on carbon-hydrogen spin-coupling constants. J. Am. Chem. Soc. 1999, 121, 870–871. [Google Scholar] [CrossRef]
- Place, A.R.; Bowers, H.A.; Bachvaroff, T.R.; Adolf, J.E.; Deeds, J.R.; Sheng, J. Karlodinium veneficum—The little dinoflagellate with a big bite. Harmful Algae 2012, 14, 179–195. [Google Scholar] [CrossRef]
- Deeds, J.R.; Reimschuessel, R.; Place, A.R. Histopathological Effects in Fish Exposed to the Toxins from Karlodinium micrum. J. Aquat. Anim. Health 2006, 18, 136–148. [Google Scholar] [CrossRef]
- Deeds, J.R.; Hoesch, R.E.; Place, A.R.; Kao, J.P.Y. The cytotoxic mechanism of karlotoxin 2 (KmTx 2) from Karlodinium veneficum (Dinophyceae). Aquat. Toxicol. 2014, 159, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Swasono, R.T.; Mouri, R.; Morsy, N.; Matsumori, N.; Oishi, T.; Murata, M. Sterol effect on interaction between amphidinol 3 and liposomal membrane as evidenced by surface plasmon resonance. Bioorg. Med. Chem. Lett. 2010, 20, 2215–2218. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.; Place, A. Sterol-specific membrane interactions with the toxins from Karlodinium micrum (Dinophyceae)—A strategy for self-protection? Afr. J. Mar. Sci. 2006, 28, 421–425. [Google Scholar] [CrossRef]
- Sieg, R.D.; Poulson-Ellestad, K.L.; Kubanek, J. Chemical ecology of the marine plankton. Nat. Prod. Rep. 2011, 28, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.D.; Lewis, N.I.; Quilliam, M.A. The marine dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the causative organism of spirolide shellfish toxins. Phycologia 2000, 39, 67–74. [Google Scholar] [CrossRef]
- Touzet, N.; Franco, J.M.; Raine, R. Morphogenetic diversity and biotoxin composition of Alexandrium (Dinophyceae) in Irish coastal waters. Harmful Algae 2008, 7, 782–797. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Stirling, D.J.; MacKenzie, A.L. Gymnodimine C, an Isomer of Gymnodimine B, from Karenia selliformis. J. Agric. Food Chem. 2003, 51, 4838–4840. [Google Scholar] [CrossRef] [PubMed]
- Davidson, K.; Baker, C.; Higgins, C.; Higman, W.; Swan, S.; Veszelovszki, A.; Turner, A.D. Potential threats posed by new or emerging marine biotoxins in UK waters and examination of detection methodologies used for their Control: Cyclic imines. Mar. Drugs 2015, 13, 7087–7112. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Curtis, J.M.; Walter, J.A.; Wright, J.L.C. Characterization of biologically inactive spirolides E and F: Identification of the spirolide pharmacophore. Tetrahedron Lett. 1996, 37, 7671–7674. [Google Scholar] [CrossRef]
- Guéret, S.M.; Brimble, M.A. Spiroimine shellfish poisoning (SSP) and the spirolide family of shellfish toxins: Isolation, structure, biological activity and synthesis. Nat. Prod. Rep. 2010, 27, 1350–1366. [Google Scholar] [CrossRef] [PubMed]
- Couesnon, A.; Aráoz, R.; Iorga, B.I.; Benoit, E.; Reynaud, M.; Servent, D.; Molgó, J. The Dinoflagellate Toxin 20-Methyl Spirolide-G Potently Blocks Skeletal Muscle and Neuronal Nicotinic Acetylcholine Receptors. Toxins 2016, 8, 249. [Google Scholar] [CrossRef] [PubMed]
- Roach, J.S.; LeBlanc, P.; Lewis, N.I.; Munday, R.; Quilliam, M.A.; MacKinnon, S.L. Characterization of a Dispiroketal Spirolide Subclass from Alexandrium ostenfeldii. J. Nat. Prod. 2009, 72, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Murphy, M.; Clausen, J.; Richard, D.; Quilliam, M.; MacKinnon, S.; LaBlanc, P.; Mueller, R.; Pulido, O. Neural injury biomarkers of novel shellfish toxins, spirolides: A pilot study using immunochemical and transcriptional analysis. Neurotoxicology 2003, 24, 593–604. [Google Scholar] [CrossRef]
- Seki, T.; Satake, M.; Mackenzie, L.; Kaspar, H.F.; Yasumoto, T. Gymnodimine, a new marine toxin of unprecedented structure isolated from New Zealand oysters and the dinoflagellate, Gymnodinium sp. Tetrahedron Lett. 1995, 36, 7093–7096. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Misner, I.; Tomas, C.R.; Wright, J.L.C. Occurrence of 12-methylgymnodimine in a spirolide-producing dinoflagellate Alexandrium peruvianum and the biogenetic implications. Tetrahedron Lett. 2011, 52, 4243–4246. [Google Scholar] [CrossRef]
- Harju, K.; Koskela, H.; Kremp, A.; Suikkanen, S.; De La Iglesia, P.; Miles, C.O.; Krock, B.; Vanninen, P. Identification of gymnodimine D and presence of gymnodimine variants in the dinoflagellate Alexandrium ostenfeldii from the Baltic Sea. Toxicon 2016, 112, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Kharrat, R.; Servent, D.; Girard, E.; Ouanounou, G.; Amar, M.; Marrouchi, R.; Benoit, E.; Molgó, J. The marine phycotoxin gymnodimine targets muscular and neuronal nicotinic acetylcholine receptor subtypes with high affinity. J. Neurochem. 2008, 107, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Hauser, T.A.; Hepler, C.D.; Kombo, D.C.; Grinevich, V.P.; Kiser, M.N.; Hooker, D.N.; Zhang, J.; Mountfort, D.; Selwood, A.; Akireddy, S.R.; et al. Comparison of acetylcholine receptor interactions of the marine toxins, 13-desmethylspirolide C and gymnodimine. Neuropharmacology 2012, 62, 2238–2249. [Google Scholar] [CrossRef] [PubMed]
- Dragunow, M.; Trzoss, M.; Brimble, M.A.; Cameron, R.; Beuzenberg, V.; Holland, P.; Mountfort, D. Investigations into the cellular actions of the shellfish toxin gymnodimine and analogues. Environ. Toxicol. Pharmacol. 2005, 20, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.; Vale, C.; Vieytes, M.R.; Laferla, F.M.; Giménez-Llort, L.; Botana, L.M. The Cholinergic Antagonist Gymnodimine Improves Aβ and Tau Neuropathology in an in Vitro Model of Alzheimer Disease. Cell. Physiol. Biochem. 2011, 27, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Naila, I.B.; Hamza, A.; Gdoura, R.; Diogène, J.; de la Iglesia, P. Prevalence and persistence of gymnodimines in clams from the Gulf of Gabes (Tunisia) studied by mouse bioassay and LC–MS/MS. Harmful Algae 2012, 18, 56–64. [Google Scholar] [CrossRef]
- Nagai, H.; Murata, M.; Torigoe, K.; Satake, M.; Yasumoto, T. Gambieric acids, new potent antifungal substances with unprecedented polyether structures from a marine dinoflagellate Gambierdiscus toxicus. J. Org. Chem. 1992, 57, 5448–5453. [Google Scholar] [CrossRef]
- Morohashi, A.; Satake, M.; Nagai, H.; Oshima, Y.; Yasumoto, T. The Absolute Configuration of Gambieric Acids A–D, Potent Antifungal Polyethers, Isolated from the Marine Dinoflagellate Gambierdiscus toxicus. Tetrahedron 2000, 56, 8995–9001. [Google Scholar] [CrossRef]
- Nagai, H.; Mirakami, Y.; Yazawa, K.; Gonoi, T.; Yasumoto, T. Biological activities of novel polyether antifungals, gamberic acids A and B from a marine dinogflagellate Gambierdiscus toxicus. J. Antibiot. 1993, 46, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Fuwa, H. Total synthesis and complete structural assignment of gambieric acid A, a large polycyclic ether marine natural product. Chem. Rec. 2014, 14, 678–703. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Hirama, M.; Satake, M.; Sugiyama, K.; Yasumoto, T. Inhibition of brevetoxin binding to the voltage-gated sodium channel by gambierol and gambieric acid-A. Toxicon 2003, 41, 469–474. [Google Scholar] [CrossRef]
- Murakami, M.; Makabe, K.; Yamaguchi, K.; Konosu, S.; Wälchli, M.R. Goniodomin a, a novel polyether macrolide from the dinoflagellate goniodoma pseudogoniaulax. Tetrahedron Lett. 1988, 29, 1149–1152. [Google Scholar] [CrossRef]
- Hsia, M.H.; Morton, S.L.; Smith, L.L.; Beauchesne, K.R.; Huncik, K.M.; Moeller, P.D.R. Production of goniodomin A by the planktonic, chain-forming dinoflagellate Alexandrium monilatum (Howell) Balech isolated from the Gulf Coast of the United States. Harmful Algae 2006, 5, 290–299. [Google Scholar] [CrossRef]
- Triki, H.Z.; Laabir, M.; Moeller, P.; Chomerat, N.; Daly-Yahia, K.O. First report of goniodomin A production by the dinoflagellate Alexandrium pseudogonyaulax developing in southern Mediterranean (Bizerte Lagoon, Tunisia). Toxicon 2016, 111, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Shi, J.; Oikawa, M.; Sasaki, M. Assignment of the Absolute Configuration of Goniodomin A by NMR Spectroscopy and Synthesis of Model Compounds. Org. Lett. 2008, 10, 1013–1016. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Inoue, D.; Matsunaga, K.; Ohizumi, Y.; Ueda, H.; Asano, T.; Murakami, M.; Sato, Y. Goniodomin A, an antifungal polyether macrolide, exhibits antiangiogenic activities via inhibition of actin reorganization in endothelial cells. J. Cell. Physiol. 2002, 190, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Nakahata, N.; Ito, E.; Murakami, M.; Yamaguchi, K.; Ohizumi, Y. Goniodomin A, an antifungal polyether macrolide, increases the filamentous actin content of 1321N1 human astrocytoma cells. J. Pharm. Pharmacol. 1998, 50, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Espiña, B.; Cagide, E.; Louzao, M.C.; Vilariño, N.; Vieytes, M.R.; Takeda, Y.; Sasaki, M.; Botana, L.M. Cytotoxicity of goniodomin A and B in non contractile cells. Toxicol. Lett. 2016, 250–251, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Tsuda, M. Amphidinolides, bioactive macrolides from symbiotic marine dinoflagellates. Nat. Prod. Rep. 2004, 21, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J. Search for New Bioactive Marine Natural Products. Chem. Pharm. Bull. 2016, 64, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, T.K.; Das, S. Chemistry of Potent Anti-Cancer Compounds, Amphidinolides. Curr. Med. Chem. Anticancer Agents 2001, 1, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J. Amphidinolides and its related macrolides from marine dinoflagellates. J. Antibiot. 2008, 61, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Usui, T.; Kazami, S.; Dohmae, N.; Mashimo, Y.; Kondo, H.; Tsuda, M.; Terasaki, A.G.; Ohashi, K.; Kobayashi, J.; Osada, H. Amphidinolide H, a Potent Cytotoxic Macrolide, Covalently Binds on Actin Subdomain 4 and Stabilizes Actin Filament. Chem. Biol. 2004, 11, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Echigoya, R.; Rhodes, L.; Oshima, Y.; Satake, M. The structures of five new antifungal and hemolytic amphidinol analogs from Amphidinium carterae collected in New Zealand. Harmful Algae 2005, 4, 383–389. [Google Scholar] [CrossRef]
- Nuzzo, G.; Cutignano, A.; Sardo, A.; Fontana, A. Antifungal amphidinol 18 and its 7-sulfate derivative from the marine dinoflagellate Amphidinium carterae. J. Nat. Prod. 2014, 77, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Cutignano, A.; Nuzzo, G.; Sardo, A.; Fontana, A. The missing piece in biosynthesis of amphidinols: First evidence of glycolate as a starter unit in New polyketides from Amphidinium carterae. Mar. Drugs 2017, 15, 157. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Cornelio, K.; Hanashima, S.; Malabed, R.; Murata, M.; Matsumori, N.; Zhang, H.; Hayashi, F.; Mori, S.; Kim, J.S.; et al. Structures of the Largest Amphidinol Homologues from the Dinoflagellate Amphidinium carterae and Structure–Activity Relationships. J. Nat. Prod. 2017, 80, 2883–2888. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Murata, M.; Yasumoto, T.; Fujita, T.; Naoki, H. Amphidinol, a polyhydroxy-polyene antifungal agent with an unprecedented structure, from a marine dinoflagellate, Amphidinium klebsii. J. Am. Chem. Soc. 1991, 113, 9859–9861. [Google Scholar] [CrossRef]
- Manabe, Y.; Ebine, M.; Matsumori, N.; Murata, M.; Oishi, T. Confirmation of the Absolute Configuration at C45 of Amphidinol 3. J. Nat. Prod. 2012, 75, 2003–2006. [Google Scholar] [CrossRef] [PubMed]
- Houdai, T.; Matsuoka, S.; Morsy, N.; Matsumori, N.; Satake, M.; Murata, M. Hairpin conformation of amphidinols possibly accounting for potent membrane permeabilizing activities. Tetrahedron 2005, 61, 2795–2802. [Google Scholar] [CrossRef]
- Espiritu, R.A.; Matsumori, N.; Tsuda, M.; Murata, M. Direct and stereospecific interaction of amphidinol 3 with sterol in lipid bilayers. Biochemistry 2014, 53, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Akakabe, M.; Kumagai, K.; Tsuda, M.; Konishi, Y.; Tominaga, A.; Tsuda, M.; Fukushi, E.; Kawabata, J. Amphirionin-5, a novel linear polyketide from a cultured marine dinoflagellate Amphidinium species with a potent cell proliferation-promoting activity. Tetrahedron Lett. 2014, 55, 3491–3494. [Google Scholar] [CrossRef]
- Minamida, M.; Kumagai, K.; Ulanova, D.; Akakabe, M.; Konishi, Y.; Tominaga, A.; Tanaka, H.; Tsuda, M.; Fukushi, E.; Kawabata, J.; et al. Amphirionin-4 with potent proliferation-promoting activity on bone marrow stromal cells from a marine dinoflagellate Amphidinium species. Org. Lett. 2014, 16, 4858–4861. [Google Scholar] [CrossRef] [PubMed]
- Akakabe, M.; Kumagai, K.; Konishi, Y.; Tominaga, A.; Tsuda, M.; Fukushi, E.; Kawabata, J.; Tsuda, M. Iriomoteolide-13a, a cytotoxic 22-membered macrolide from a marine dinoflagellate Amphidinium species. Tetrahedron 2014, 70, 2962–2965. [Google Scholar] [CrossRef]
- Bensoussan, C.; Rival, N.; Hanquet, G.; Colobert, F.; Reymond, S.; Cossy, J. Isolation, structural determination and synthetic approaches toward amphidinol 3. Nat. Prod. Rep. 2014, 31, 468. [Google Scholar] [CrossRef] [PubMed]
- Lahlou, M. The Success of Natural Products in Drug Discovery. Pharmacol. Pharm. 2013, 4, 17–31. [Google Scholar] [CrossRef]
- Shimizu, Y. Microalgae as a Drug Source. In Drugs from the Sea; Fusetani, N., Ed.; Karger Medical and Scientific Publishers: Basel, Switzerland, 2000; pp. 30–45. ISBN 978-3-318-00599-8. [Google Scholar]
- Mendes, A.; Reis, A.; Vasconcelos, R.; Guerra, P.; Lopes Da Silva, T. Crypthecodinium cohnii with emphasis on DHA production: A review. J. Appl. Phycol. 2009, 21, 199–214. [Google Scholar] [CrossRef]
- Beuzenberg, V.; Mountfort, D.; Holland, P.; Shi, F.; Mackenzie, L. Optimization of growth and production of toxins by three dinoflagellates in photobioreactor cultures. J. Appl. Phycol. 2012, 24, 1023–1033. [Google Scholar] [CrossRef]
- Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; García-Camacho, F.; Cerón-García, M.C.; Belarbi, E.H.; Chisti, Y.; Molina-Grima, E. Causes of shear sensitivity of the toxic dinoflagellate Protoceratium reticulatum. Biotechnol. Prog. 2009, 25, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; García-Camacho, F.; Cerón-García, M.; Belarbi, E.H.; Molina-Grima, E. Culture of dinoflagellates in a fed-batch and continuous stirred-tank photobioreactors: Growth, oxidative stress and toxin production. Process Biochem. 2010, 45, 660–666. [Google Scholar] [CrossRef]
- Ten Lohuis, M.R.; Miller, D.J. Genetic transformation of dinoflagellates (Amphidinium and Symbiodinium): Expression of GUS in microalgae using heterologous promoter constructs. Plant J. 1998, 13, 427–435. [Google Scholar] [CrossRef]
- Radakovits, R.; Jinkerson, R.E.; Darzins, A.; Posewitz, M.C. Genetic engineering of algae for enhanced biofuel production. Eukaryot. Cell 2010, 9, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Beer, L.L.; Boyd, E.S.; Peters, J.W.; Posewitz, M.C. Engineering algae for biohydrogen and biofuel production. Curr. Opin. Biotechnol. 2009, 20, 264–271. [Google Scholar] [CrossRef] [PubMed]
- McEwan, M.; Humayun, R.; Slamovotis, C.H.; Keeling, P.J. Nuclear Genome Sequence Survey of the Dinoflagellate Heterocapsa triquetra. J. Eukaryot. Microbiol. 2008, 55, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Wisecaver, J.H.; Hackett, J.D. Dinoflagellate Genome Evolution. Annu. Rev. Microbiol. 2011, 65, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glöckner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative Genomic and Transcriptomic Characterization of the Toxigenic Marine Dinoflagellate Alexandrium ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef] [PubMed]
- Lin, S. Genomic understanding of dinoflagellates. Res. Microbiol. 2011, 162, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; Zhang, S.F.; Zhang, Y.; Lin, L. Paralytic shellfish toxin biosynthesis in cyanobacteria and dinoflagellates: A molecular overview. J. Proteom. 2016, 135, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Rutjes, F.P.J.T.; Theodorakis, E.A.; Tiebes, J.; Sato, M.; Untersteller, E. Total Synthesis of Brevetoxin B. 3. Final Strategy and Completion. J. Am. Chem. Soc. 1995, 117, 10252–10263. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Gunzner, J.L.; Shi, G.; Agrios, K.A.; Gärtner, P.; Yang, Z. Total Synthesis of Brevetoxin A: Part 4: Final Stages and Completion. Chem. Eur. J. 1999, 5, 646–658. [Google Scholar] [CrossRef]
- Fuwa, H.; Kainuma, N.; Tachibana, K.; Sasaki, M. Total Synthesis of (−)-Gambierol. J. Am. Chem. Soc. 2002, 124, 14983–14992. [Google Scholar] [CrossRef] [PubMed]
- Tsukano, C.; Ebine, M.; Sasaki, M. Convergent Total Synthesis of Gymnocin-A and Evaluation of Synthetic Analogues. J. Am. Chem. Soc. 2005, 127, 4326–4335. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Koftis, T.V.; Vyskocil, S.; Petrovic, G.; Tang, W.; Frederick, M.O.; Chen, D.Y.; Li, Y.; Ling, T.; Yamada, Y.M. Total Synthesis and Structural Elucidation of Azaspiracid-1. Final Assignment and Total Synthesis of the Correct Structure of Azaspiracid-1. J. Am. Chem. Soc. 2006, 128, 2244–2257. [Google Scholar] [CrossRef] [PubMed]
- Fuwa, H.; Ishigai, K.; Hashizume, K.; Sasaki, M. Total Synthesis and Complete Stereostructure of Gambieric Acid A. J. Am. Chem. Soc. 2012, 134, 11984–11987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rainier, J.D. Synthesis of the ABCDEF and FGHI ring system of yessotoxin and adriatoxin. J. Antibiot. 2016, 69, 259–272. [Google Scholar] [CrossRef] [PubMed]
- García-Camacho, F.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Cerón-García, M.C.; Belarbi, E.H.; Molina-Grima, E. Determination of shear stress thresholds in toxic dinoflagellates cultured in shaken flasks. Implications in bioprocess engineering. Process Biochem. 2007, 42, 1506–1515. [Google Scholar] [CrossRef]
- López-Rosales, L.; García-Camacho, F.; Sánchez-Mirón, A.; Beato, E.M.; Chisti, Y. Pilot-scale bubble column photobioreactor culture of a marine dinoflagellate microalga illuminated with light emission diodes. Bioresour. Technol. 2016, 216, 845–855. [Google Scholar] [CrossRef] [PubMed]
- López-Rosales, L.; Sánchez-Mirón, A.; Contreras-Gomes, A.; García-Camacho, F.; Molina-Grima, E. An optimisation approach for culturing shear-sensitive dinoflagellate microalgae in bench-scale bubble column photobioreactors. Bioresour. Technol. 2015, 197, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Grünewald, C.; Bayliss, C.; Fonlut, F.; Chapuli, E. Long-term dinoflagellate culture performance in a commercial photobioreactor: Amphidinium carterae case. Bioresour. Technol. 2016, 218, 533–540. [Google Scholar] [CrossRef] [PubMed]
- García-Camacho, F.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Belarbi, E.H.; Chisti, Y.; Molina-Grima, E. Photobioreactor scale-up for a shear-sensitive dinoflagellate microalga. Process Biochem. 2011, 46, 936–944. [Google Scholar] [CrossRef]
- Hyka, P.; Lickova, S.; Přibyl, P.; Melzoch, K.; Kovar, K. Flow cytometry for the development of biotechnological processes with microalgae. Biotechnol. Adv. 2013, 31, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Grima, E.M.; Acie, F.G.; Medina, A.R.; Chisti, Y. Recovery of microalgal biomass and metabolites: Process options and economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef]
- Grima, E.; Belarbi, E.-H.; Acién Fernández, F.; Chisti, Y. Tubular photobioreactor design for algal cultures. J. Biotechnol. 2001, 92, 113–131. [Google Scholar]
- Acién, F.F.G.; Fernández, S.J.M.; Sánchez, P.J.A.; Molina, G.E.; Chisti, Y. Airlift-driven external-loop tubular photobioreactors for outdoor production of microalgae: Assessment of design and performance. Chem. Eng. Sci. 2001, 56, 2721–2732. [Google Scholar] [CrossRef]
- Wong, J.T.Y.; Kwok, A.C.M. Proliferation of dinoflagellates: Blooming or bleaching. BioEssays 2005, 27, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Fensome, R.A.; MacRae, R.A.; Williams, G.L. Dinoflagellate evolution and diversity through time. Mar. Micropaleontol. 1995, 4, 1–12. [Google Scholar] [CrossRef]
- Doblin, M.A.; Thompson, P.A.; Revill, A.T.; Butler, E.C.V.; Blackburn, S.I.; Hallegraeff, G.M. Vertical migration of the toxic dinoflagellate Gymnodinium catenatum under different concentrations of nutrients and humic substances in culture. Harmful Algae 2006, 5, 665–677. [Google Scholar] [CrossRef]
- Juhl, A.R.; Trainer, V.L.; Latz, M.I. Effect of fluid shear and irradiance on population growth and cellular toxin content of the dinoflagellate Alexandrium fundyense. Limnol. Oceanogr. 2001, 46, 758–764. [Google Scholar] [CrossRef]
- Laabir, M.; Collos, Y.; Masseret, E.; Grzebyk, D.; Abadie, E.; Savart, V.; Sibat, M.; Amzil, Z. Influence of environmental factors on the paralytic shellfish toxin content and profile of Alexandrium catenella (Dinophyceae) isolated from the Mediterranean Sea. Mar. Drugs 2013, 11, 1583–1601. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.M.; Muñoz, M.G.; Contreras, A.M. Temperature as a factor regulating growth and toxin content in the dinoflagellate Alexandrium catenella. Harmful Algae 2006, 5, 762–769. [Google Scholar] [CrossRef]
- Scalco, E.; Brunet, C.; Marino, F.; Rossi, R.; Soprano, V.; Zingone, A.; Montresor, M. Growth and toxicity responses of Mediterranean Ostreopsis cf. ovata to seasonal irradiance and temperature conditions. Harmful Algae 2012, 17, 25–34. [Google Scholar] [CrossRef]
- López-Rosales, L.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Cerón-García, M.d.C.; Belarbi, E.H.; García-Camacho, F.; Molina-Grima, E. Simultaneous effect of temperature and irradiance on growth and okadaic acid production from the marine dinoflagellate Prorocentrum belizeanum. Toxins 2013, 6, 229–253. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Chanley, M.H., Smith, W.L., Eds.; Springer: Boston, MA, USA, 1975; pp. 29–60. ISBN 0306308045. [Google Scholar]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar] [CrossRef]
- Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Cerón-García, M.C.; Belarbi, E.H.; García-Camacho, F.; Chisti, Y.; Molina-Grima, E. Macronutrients requirements of the dinoflagellate Protoceratium reticulatum. Harmful Algae 2009, 8, 239–246. [Google Scholar] [CrossRef]
- Hardison, D.R.; Sunda, W.G.; Shea, D.; Litaker, R.W. Increased toxicity of Karenia brevis during phosphate limited growth: Ecological and evolutionary implications. PLoS ONE 2013, 8, e58545. [Google Scholar] [CrossRef] [PubMed]
- Frangópulos, M.; Guisande, C.; deBlas, E.; Maneiro, I. Toxin production and competitive abilities under phosphorus limitation of Alexandrium species. Harmful Algae 2004, 3, 131–139. [Google Scholar] [CrossRef]
- John, E.H.; Flynn, K.J. Growth dynamics and toxicity of Alexandrium fundyense (Dinophyceae): The effect of changing N:P supply ratios on internal toxin and nutrient levels. Eur. J. Phycol. 2000, 35, 11–23. [Google Scholar] [CrossRef]
- Vanucci, S.; Pezzolesi, L.; Pistocchi, R.; Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Tartaglione, L.; Guerrini, F. Nitrogen and phosphorus limitation effects on cell growth, biovolume, and toxin production in Ostreopsis cf. ovata. Harmful Algae 2012, 15, 78–90. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Fernández Amandi, M.; McKenzie, L.; Furey, A.; James, K.J. Effects of selenium, iron and cobalt addition to growth and yessotoxin production of the toxic marine dinoflagellate Protoceratium reticulatum in culture. J. Exp. Mar. Biol. Ecol. 2004, 313, 337–351. [Google Scholar] [CrossRef]
- Cruz-López, R.; Maske, H. The vitamin B1 and B12 required by the marine dinoflagellate Lingulodinium polyedrum can be provided by its associated bacterial community in culture. Front. Microbiol. 2016, 7, 560. [Google Scholar] [CrossRef] [PubMed]
- García-Camacho, F.; López-Rosales, L.; Sánchez-Mirón, A.; Belarbi, E.H.; Chisti, Y.; Molina-Grima, E. Artificial neural network modeling for predicting the growth of the microalga Karlodinium veneficum. Algal Res. 2016, 14, 58–64. [Google Scholar] [CrossRef]
- López-Rosales, L.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Contreras-Gómez, A.; García-Camacho, F.; Molina-Grima, E. Modelling of multi-nutrient interactions in growth of the dinoflagellate microalga Protoceratium reticulatum using artificial neural networks. Bioresour. Technol. 2013, 146, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Weuster-Botz, D. Experimental design for fermentation media development: Statistical design or global random search? J. Biosci. Bioeng. 2000, 90, 473–483. [Google Scholar] [CrossRef]
- García-Camacho, F.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Chisti, Y.; Molina-Grima, E. Genetic algorithm-based medium optimization for a toxic dinoflagellate microalga. Harmful Algae 2011, 10, 697–701. [Google Scholar] [CrossRef]
- López-Rosales, L.; García-Camacho, F.; Sánchez-Mirón, A.; Chisti, Y. An optimal culture medium for growing Karlodinium veneficum: Progress towards a microalgal dinoflagellate-based bioprocess. Algal Res. 2015, 10, 177–182. [Google Scholar] [CrossRef]
- Schulze, P.S.C.; Barreira, L.A.; Pereira, H.G.C.; Perales, J.A.; Varela, J.C.S. Light emitting diodes (LEDs) applied to microalgal production. Trends Biotechnol. 2014, 32, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.K.; Oh, S.J.; Yang, H.-S.; Kim, D.-M.; Kang, I.J.; Oshima, Y. Laboratory Study for the Phytoremediation of Eutrophic Coastal Sediment Using Benthic Microalgae and Light Emitting Diode (LED). J. Fac. Agric. Kyushu Univ. 2013, 58, 417–425. [Google Scholar]
- Wang, S.; Chen, J.; Li, Z.; Wang, Y.; Fu, B.; Han, X.; Zheng, L. Cultivation of the benthic microalga Prorocentrum lima for the production of diarrhetic shellfish poisoning toxins in a vertical flat photobioreactor. Bioresour. Technol. 2015, 179, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ghosh, S.; Fixler, D.; Dubinsky, Z.; Iluz, D. Flashing light in microalgae biotechnology. Bioresour. Technol. 2016, 203, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Tomori, Y.; Tanimoto, Y.; Oku, O.; Adachi, M. Evaluation of the effects of light intensity on growth of the benthic dinoflagellate Ostreopsis sp. 1 using a newly developed photoirradiation-culture system and a novel regression analytical method. Harmful Algae 2014, 39, 48–54. [Google Scholar] [CrossRef]
- Etheridge, S.M.; Roesler, C.S. Effects of temperature, irradiance, and salinity on photosynthesis, growth rates, total toxicity, and toxin composition for Alexandrium fundyense isolates from the Gulf of Maine and Bay of Fundy. Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2491–2500. [Google Scholar] [CrossRef]
- Lee, T.C.H.; Fong, F.L.Y.; Ho, K.C.; Lee, F.W.F. The mechanism of diarrhetic shellfish poisoning toxin production in Prorocentrum spp.: Physiological and molecular perspectives. Toxins 2016, 8, 272. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Cembella, A.D.; Quilliam, M.A. Cell cycle and toxin production in the benthic dinoflagellate Prorocentrum lima. Mar. Biol. 1999, 134, 541–549. [Google Scholar] [CrossRef]
- Taroncher-Oldenburg, G.; Kulis, D.M.; Anderson, D.M. Toxin variability during the cell cycle of the dinoflagellate Alexandrium fundyense. Limnol. Oceanogr. 1997, 42, 1178–1188. [Google Scholar] [CrossRef]
- John, U.; Quilliam, M.A.; Medlin, L.; Cembella, A.D. Spirolide production and photoperiod-dependent growth of the marine dinoflagellate Alexandrium ostenfeldii. In Harmful Algal Blooms 2000; Hallegraeff, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; International Oceanographic Commission (UNESCO): Paris, France, 2001; pp. 299–302. [Google Scholar]
- Loader, J.I.; Hawkes, A.D.; Beuzenberg, V.; Jensen, D.J.; Cooney, J.M.; Wilkins, A.L.; Fitzgerald, J.M.; Briggs, L.R.; Miles, C.O. Convenient Large-Scale Purification of Yessotoxin from Protoceratium reticulatum Culture and Isolation of a Novel Furanoyessotoxin. J. Agric. Food Chem. 2007, 55, 11093–11100. [Google Scholar] [CrossRef] [PubMed]
- Jauffrais, T.; Sechet, V.; Herrenknecht, C.; Krock, B.; Amzil, Z.; Hess, P. Growth and toxin production of Azadinium spinosum in batch and continuous culture. In Proceedings of the 14th International Conference on Harmful Algae, Hersonissos, Crete, Greece, 1–5 November 2010; Volume 3, pp. 18–20. [Google Scholar]
- Jauffrais, T.; Kilcoyne, J.; Séchet, V.; Herrenknecht, C.; Truquet, P.; Hervé, F.; Bérard, J.B.; Nulty, C.; Taylor, S.; Tillmann, U.; et al. Production and isolation of azaspiracid-1 and -2 from Azadinium spinosum culture in pilot scale photobioreactors. Mar. Drugs 2012, 10, 1360–1382. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Shi, Y.; Cong, W. Improvement in growth and toxin production of Alexandrium tamarense by two-step culture method. J. Appl. Phycol. 2006, 18, 119–126. [Google Scholar] [CrossRef]
- Wang, D.Z.; Ho, A.Y.T.; Hsieh, D.P.H. Production of C2 toxin by Alexandrium tamarense CI01 using different culture methods. J. Appl. Phycol. 2002, 14, 461–468. [Google Scholar] [CrossRef]
- Wang, D.; Hsieh, D.P.H. Dynamics of C2 toxin and chlorophyll-a formation in the dinoflagellate Alexandrium tamarense during large scale cultivation. Toxicon 2001, 39, 1533–1536. [Google Scholar] [CrossRef]
- Gallardo-Rodríguez, J.J.; García-Cerón, M.C.; García-Camacho, F.; Sánchez-Mirón, A.; Belarbi, E.H.; Molina-Grima, E. New culture approaches for yessotoxin production from the dinoflagellate Protoceratium reticulatum. Biotechnol. Prog. 2007, 23, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Parker, N.S.; Negri, A.P.; Frampton, D.M.F.; Rodolfi, L.; Tredici, M.R.; Blackburn, S.I. Growth of the toxic dinoflagellate Alexandrium minutum (Dinophyceae) using high biomass culture systems. J. Appl. Phycol. 2002, 14, 313–324. [Google Scholar] [CrossRef]
- Medhioub, W.; Sechet, V.; Truquet, P.; Bardouil, M.; Amzil, Z.; Lassus, P.; Soudant, P. Alexandrium ostenfeldii growth and spirolide production in batch culture and photobioreactor. Harmful Algae 2011, 10, 794–803. [Google Scholar] [CrossRef]
- Han, M.; Lee, H.; Anderson, D.M.; Kim, B. Paralytic shellfish toxin production by the dinoflagellate Alexandrium pacificum (Chinhae Bay, Korea) in axenic, nutrient-limited chemostat cultures and nutrient-enriched batch cultures. Mar. Pollut. Bull. 2016, 104, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, K.W.; Ko, J.Y.; Shah, M.M.R.; Lee, J.H.; Kang, M.C.; O-Nam, K.; Lee, J.B.; Jeon, Y.J. In vitro studies of anti-inflammatory and anticancer activities of organic solvent extracts from cultured marine microalgae. Algae 2013, 28, 111–119. [Google Scholar] [CrossRef]
- Fuentes-Grünewald, C.; Garcés, E.; Alacid, E.; Rossi, S.; Camp, J. Biomass and Lipid Production of Dinoflagellates and Raphidophytes in Indoor and Outdoor Photobioreactors. Mar. Biotechnol. 2013, 15, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Benstein, R.M.; Çebi, Z.; Podola, B.; Melkonian, M. Immobilized Growth of the Peridinin-Producing Marine Dinoflagellate Symbiodinium in a Simple Biofilm Photobioreactor. Mar. Biotechnol. 2014, 16, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Rahman Sha, M.M.; Samarakoon, K.W.; An, S.-J.; Jeon, Y.-J.; Lee, J.-B. Growth Characteristics of Three Benthic Dinoflagellates in Mass Culture and Their Antioxidant Properties. J. Fish. Aquat. Sci. 2016, 11, 268–277. [Google Scholar] [CrossRef]
- Tilney, C.L.; Hoadley, K.D.; Warner, M.E. Comparing the diel vertical migration of Karlodinium veneficum (dinophyceae) and Chattonella subsalsa (Raphidophyceae): PSII photochemistry, circadian control, and carbon assimilation. J. Photochem. Photobiol. B Biol. 2015, 143, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Zittelli, G.C.; Rodolfi, L.; Bassi, N.; Biondi, N.; Tredici, M.R. Photobioreactors for Microalgal Biofuel Production. In Algae for Biofuels and Energy; Borowitzka, M.A., Moheimani, N., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 115–131. [Google Scholar]



















{kind=link}
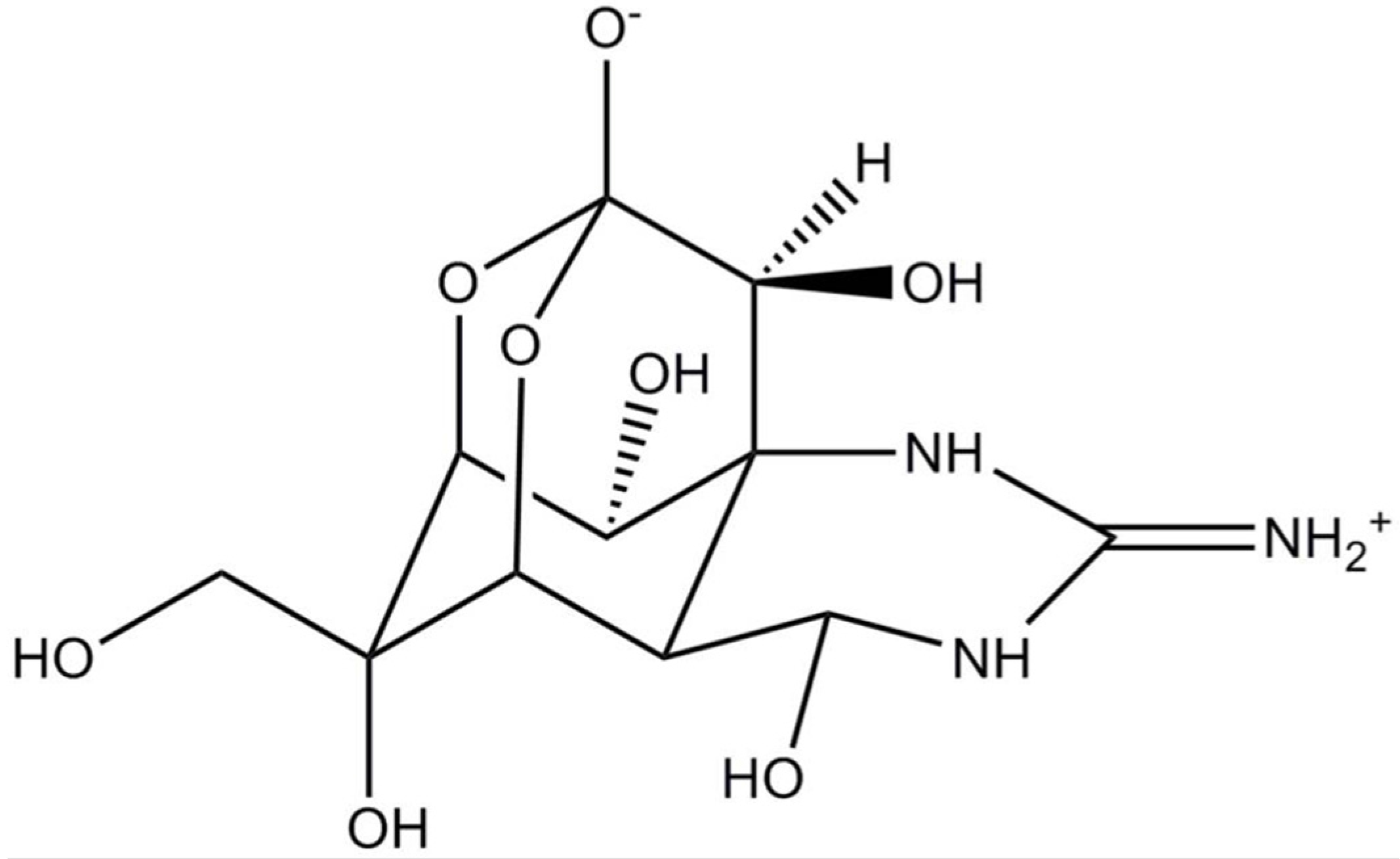{kind=link}
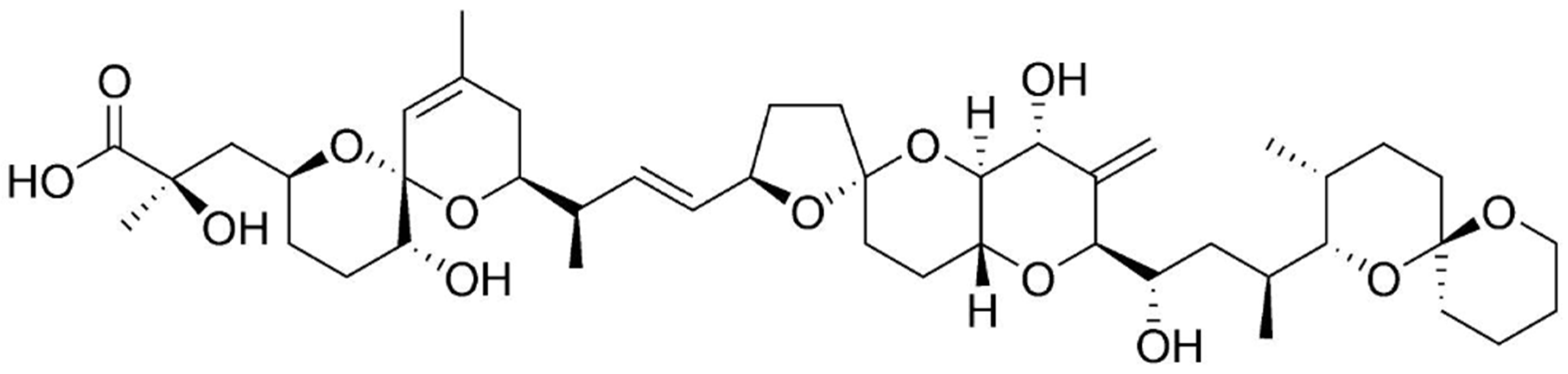{kind=link}
{kind=link}
{kind=link}
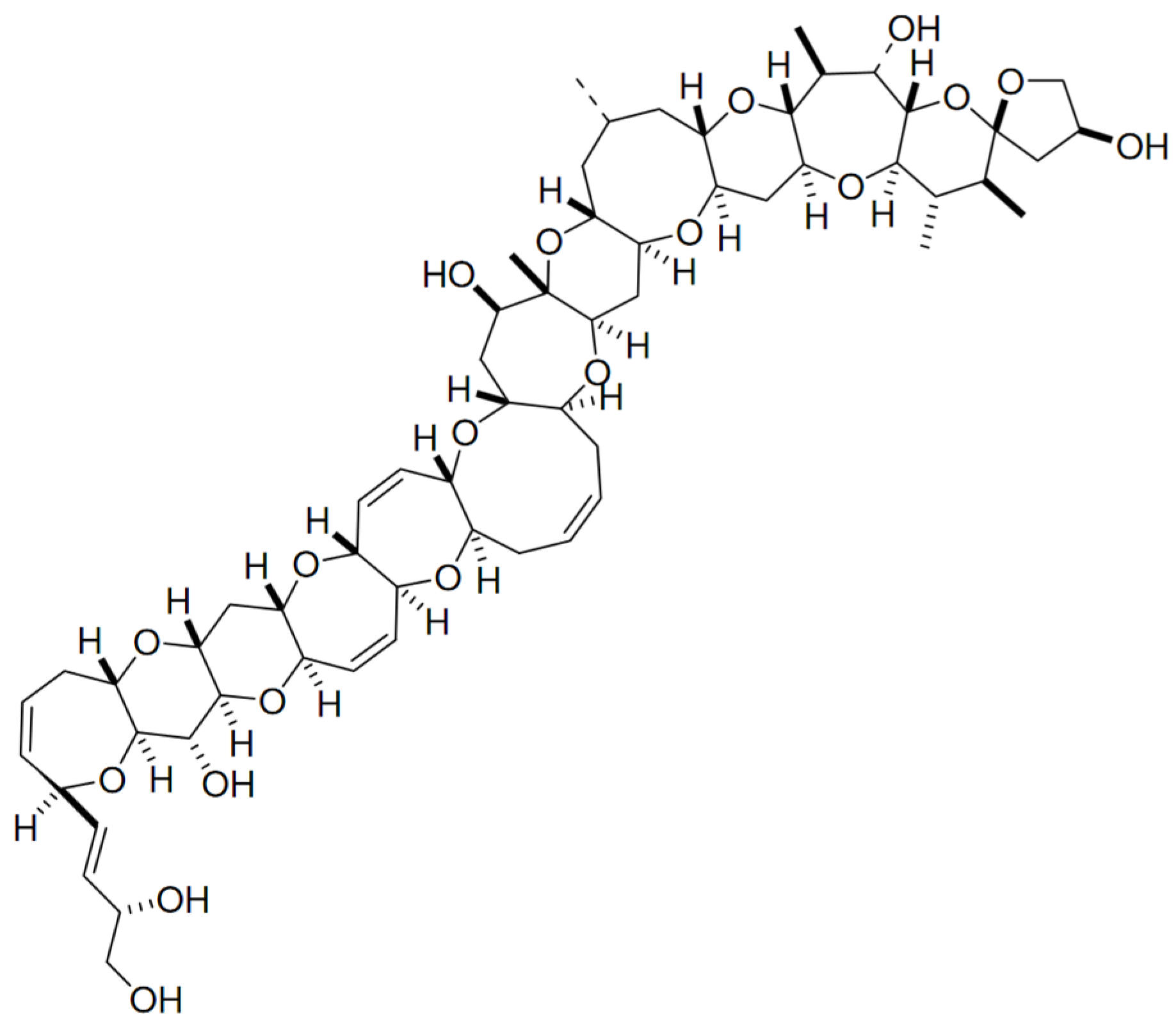{kind=link}
{kind=link}
{kind=link}
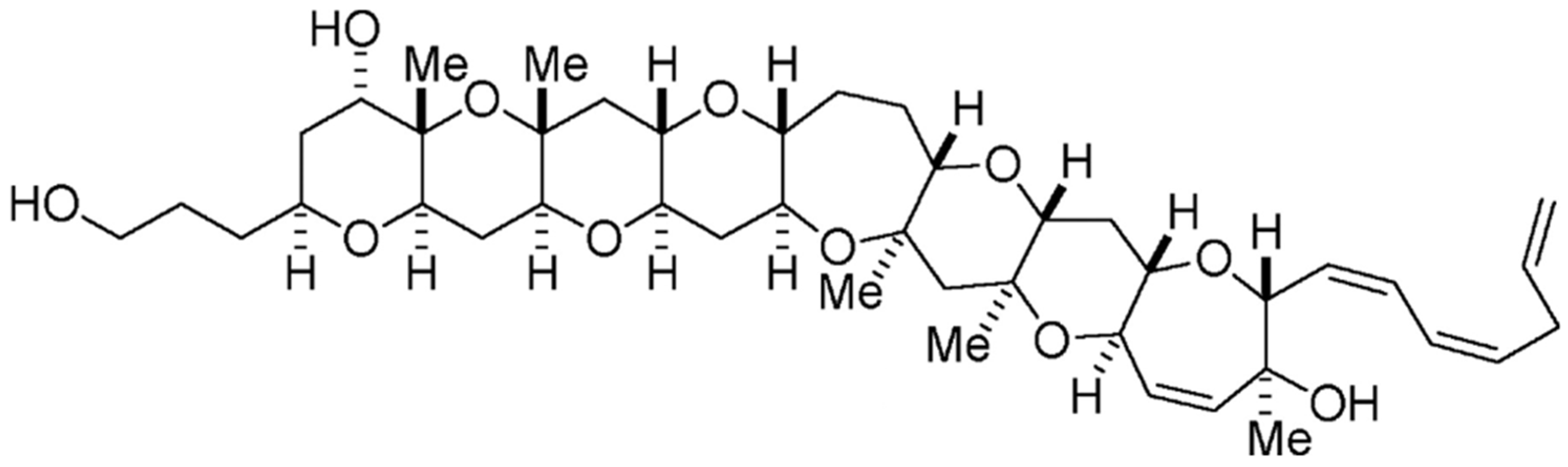{kind=link}
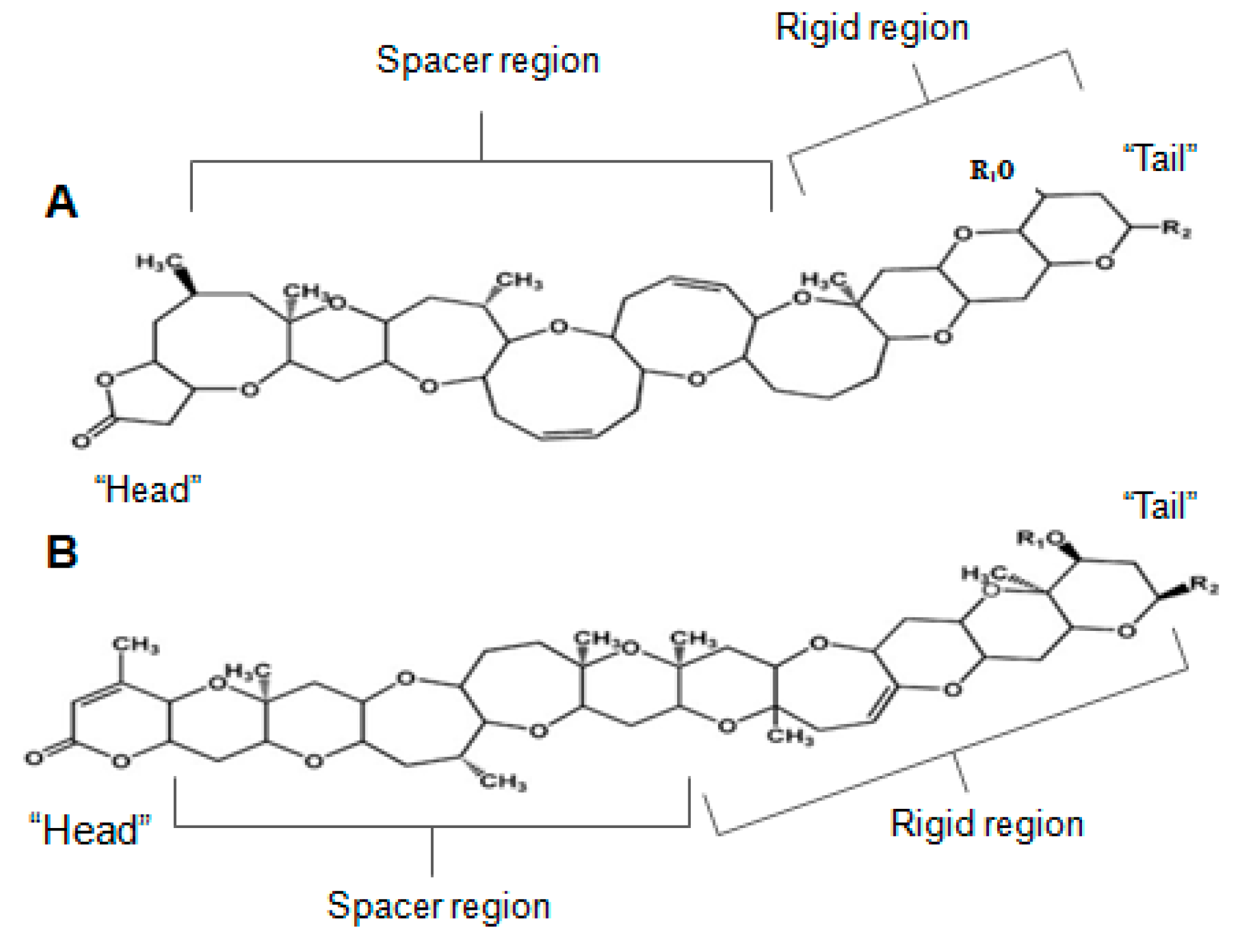{kind=link}
{kind=link}
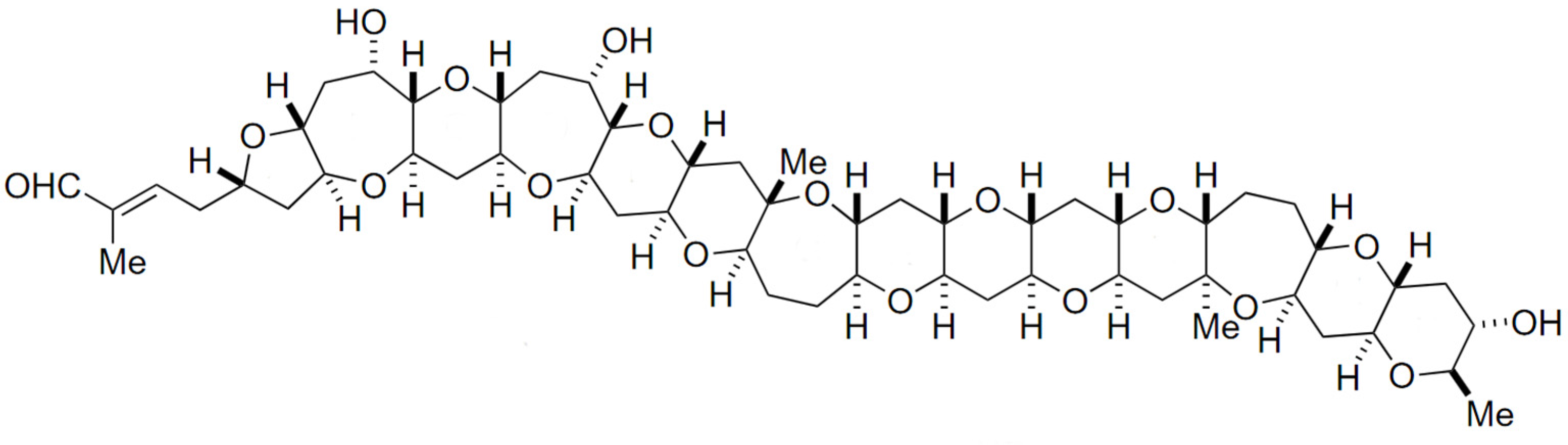{kind=link}
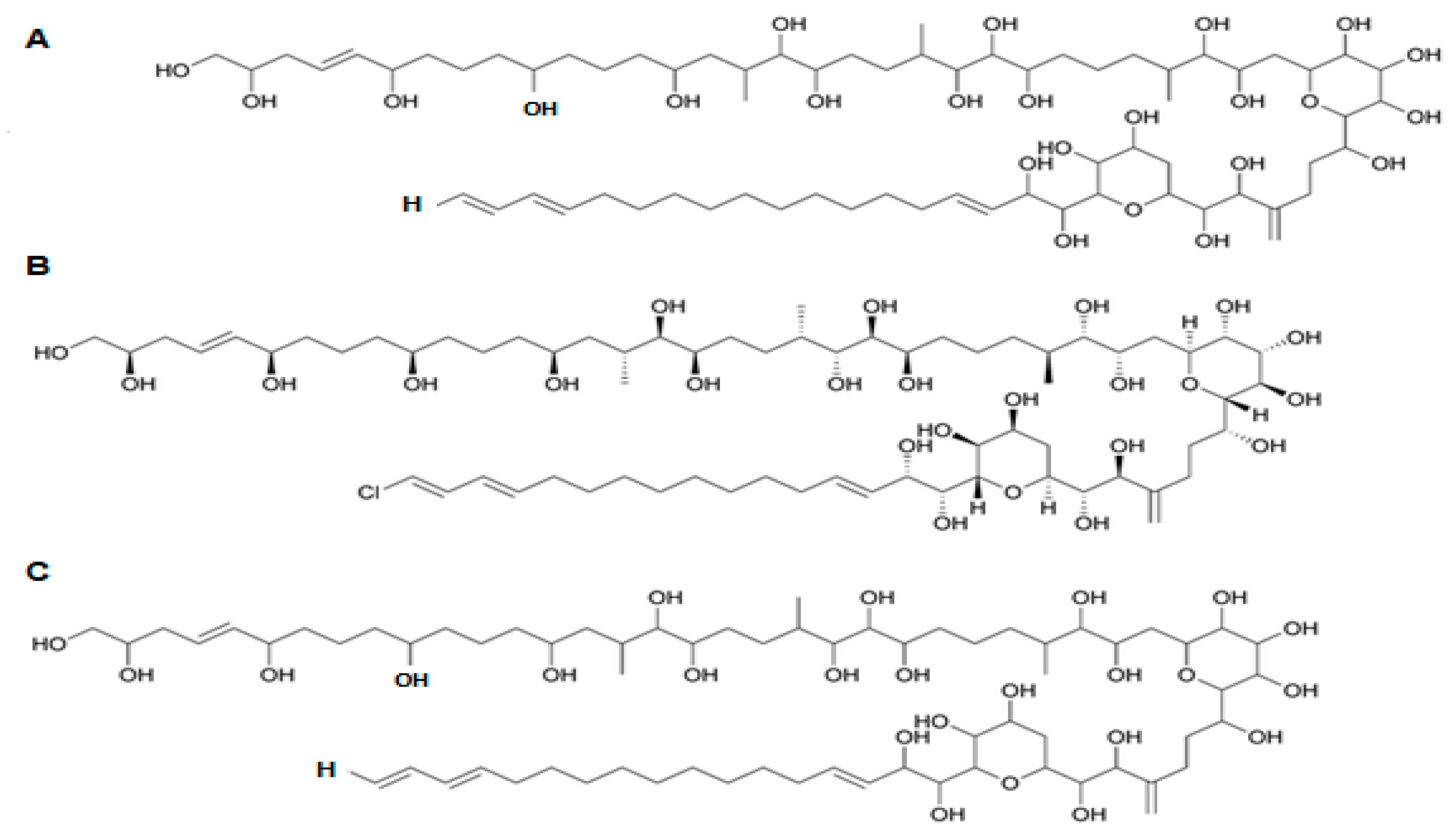{kind=link}
{kind=link}
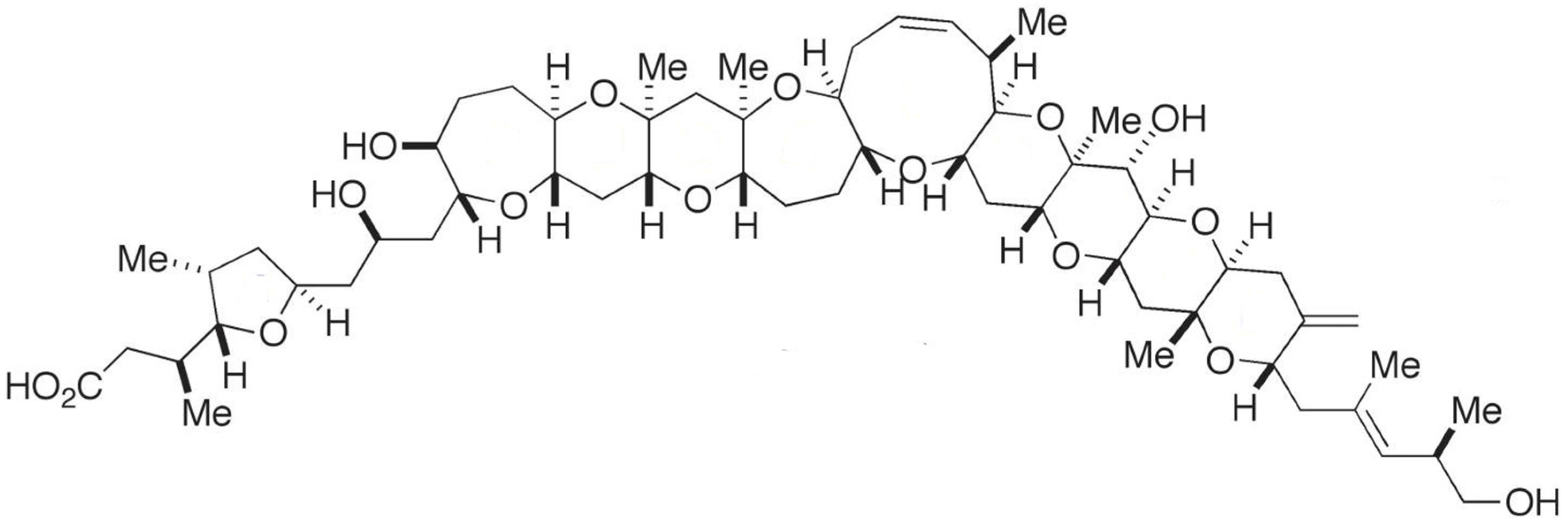{kind=link}
{kind=link}
{kind=link}
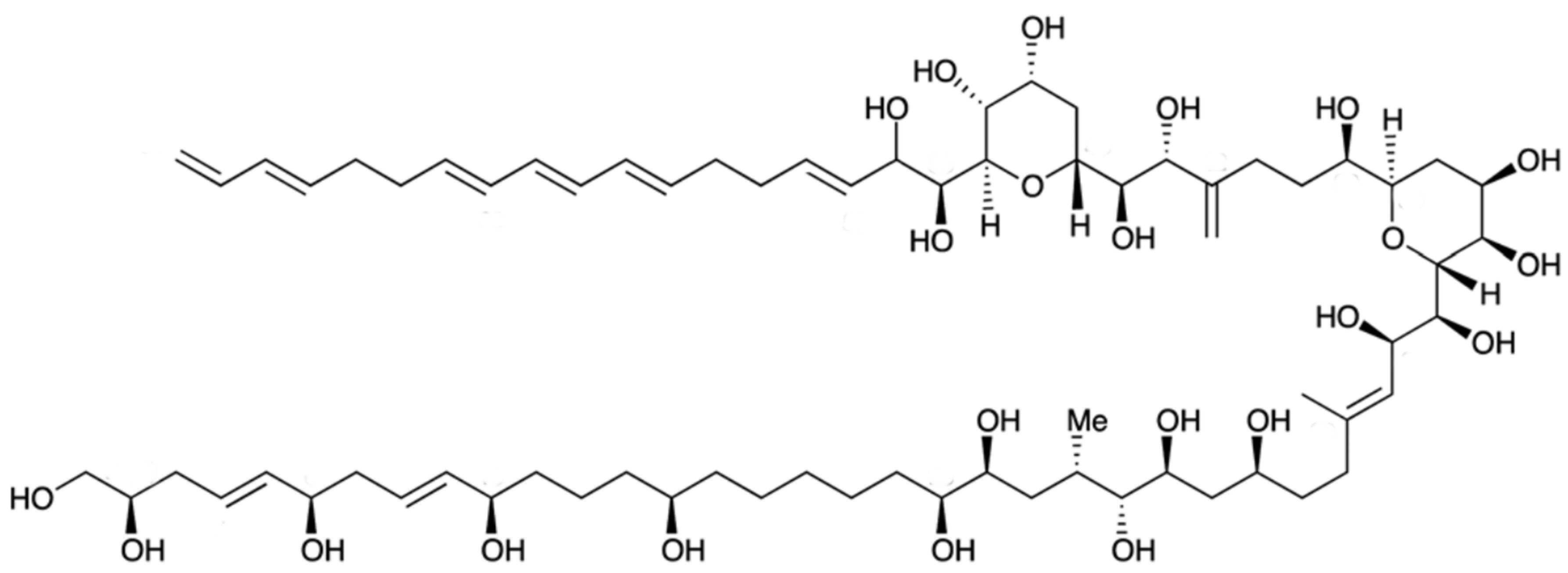{kind=link}
| Patent Name/Application | Biotoxin Used | Year | Reference |
|---|---|---|---|
| 10′,11′-modified saxitoxins useful for treatment of pain | Modified saxitoxin | 2017 | US20170029431 |
| Use of sodium channel blockers for treatment of neuropathic pain developing as consequence of chemotherapy | Tetrodotoxin, saxitoxin (analogues and derivatives) | 2017 | US20170000797 |
| Ladder-frame polyether conjugates | Brevetoxin, maitotoxin, yessotoxin, gambierol | 2016 | US20160128321 |
| Sodium channel blocker for treatment of loss of superficial sensitivity | Gonyautoxin | 2016 | US20160000793 |
| Palytoxyn, medical use and process for its isolation | Palytoxin | 2014 | EP3087172 |
| Neosaxitoxin combination formulations for prolonged local anesthesia | Neosaxitoxin | 2014 | WO2014145580 |
| Treatment of loss of sense of touch with saxitoxin derivatives | Saxitoxin | 2014 | EP2533785 |
| Using yessotoxin and its derivatives for treatment of gliomas | Yessotoxin and derivatives | 2013 | ES2393696 |
| Use of yessotoxin and analogues and derivatives thereof for treating and/or preventing neurodegenerative diseases linked to tau and beta amyloid | Yessotoxin (analogues and derivatives) | 2013 | US20130035302 |
| Use of gambierol for treating and/or preventing neurodegenerative diseases related to tau and beta-amyloid | Gambierol | 2012 | US20120283321 |
| Use of yessotoxins and derivatives thereof for treatment and/or prevention of metabolic diseases | Yessotoxin and derivatives | 2012 | WO2012140298 |
| Use of gymnodimine, analogues and derivatives for treatment and/or prevention of neurodegenerative diseases associated with tau and beta-amyloid | Gymnodimine (analogues and derivatives) | 2012 | US20120245223 |
| Therapeutic use of yessotoxin as human tumor cell growth inhibitor | Yessotoxin | 2011 | EP1875906 |
| Methods and compositions for studying, imaging, and treating pain | Saxitoxin, gonyautoxin and other analogues | 2011 | WO2010129864 |
| Use of sodium ion channel blocker in treating biology drug resistance of antibiotic | Tetrodotoxin, saxitoxin | 2009 | CN101450056 |
| Polyether brevetoxin derivatives as treatment for cystic fibrosis, mucociliary dysfunction, and pulmonary diseases | Brevetoxins and derivatives | 2008 | US7399782 |
| Use of sodium channel blockers for treatment of preterm labor | Tetrodotoxin, saxitoxin (analogues and derivatives) | 2007 | WO2007096170 |
| Polyether brevetoxin derivatives as treatment for neurotoxic shellfish poisoning and ciguatera fish poisoning | Brevetoxin and derivatives | 2005 | WO2005027903 |
| Methods of treating wounds with gonyautoxins | Gonyautoxin | 2005 | WO2005110275 |
| Dinoflagellate karlotoxins, methods of isolation and uses thereof | Karlotoxin | 2005 | US2005/0209104 |
| Use of yessotoxin in treatment of allergic and asthmatic processes | Yessotoxin | 2003 | EP1875907 |
| Toxin | Supplier | Source | Price Range (€ mg−1) |
|---|---|---|---|
| Okadaic acid | SGA GEN LCL MER SL TC BP SCB | Prorocentrum concavum unkown unkown Prorocentrum sp. unkown unkown unkown Prorocentrum sp. | 1430 3510 561 2538 3479 5000 1210 1850 |
| Okadaic acid, ammonium salt | SGA LCL | Prorocentrum concavum unkown | 16,620 561 |
| Okadaic acid, sodium salt | SGA GEN LCL SL | Prorocentrum concavum unkown unkown unkown | 9730 2320 561 2657 |
| Okadaic acid, potassium salt | SGA GEN LCL | Prorocentrum concavum unkown unkown | 8050 2320 561 |
| Okadaic acid, solution | MER | Prorocentrum sp. | 4480 |
| Tetrodotoxin | GEN TC | unkown unkown | 451 195 |
| Saxitoxin, diacetate | GEN | unkown | 9920 |
| Brevetoxin 2 | GEN | unkown | 17,960 |
| MER | Karenia brevis | 4390 | |
| SCB | Karenia brevis | 5970 | |
| Brevetoxin 3 | GEN | unkown | 17,530 |
| MER | Karenia brevis | 3880 | |
| Brevetoxin 9 | GEN | unkown | 19,380 |
| Yessotoxin, antibody | GEN | unkown | 1580 |
| Maitotoxin | LCL | unkown | unkown |
| Azaspiracid-1 | SCB | Marine mussel | 357,000 |
| Azaspiracid-2 | SCB | Marine mussel | 359,000 |
| Azaspiracid-3 | SCB | Marine mussel | 540,000 |
| Species and Strain | Type of Reactor | Reactor Operation | Reactor Size | Type of Aeration/Agitation | T (°C) | Irradiance Culture System | Light Intensity (µmol m−2 s−1) | Light Regime | Biotoxin Produced | Specific r-Emsarks | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Prorocentrum lima spp. | Carboy | Batch | 36 L | Stirring (discontinuous) | 18 ± 1 | - | 90 ± 5 | Continuous dark/14 h:10 h | Okadaic acid Dinophysistoxin | - | [326] |
| Protoceratium reticulatum CAWD129 | Carboy | Batch | 226 L | - | - | - | - | 12 h/12 h | Yessotoxin Furanoyessotoxin | Serial bulk culture (size ~14 L each) | [329] |
| Azadinium spinosum 3D9 | Chemostat in series | Continuous | 100 L | Stirring (not specified) | 18 | - | 200 | 16 h/8 h | Azaspiracid | - | [330] |
| Azadinium spinosum 3D9 | Tubular PBRs in series | Continuous | 100 L | Stirring (Rushton turbine) | 18 | Neon tube lamps | 200 | 16 h/8 h | Azaspiracid | Culture collected in an aerated harvesting tank (300 L) | [331] |
| Alexandrium tamarense ATHK | Airlift PBR | Batch | 2.5 L | Airlift | 22 | Cool-white fluorescent lamps | 60 | Continuous | Saxitoxin | Two-step batch culture method | [332] |
| Alexandrium tamarense ATCI01 | Glass rectangular tank | Batch/Semi-continuous | 70 L 40 L | Air bubbling (continuous) | 23 | Cool-white fluorescent lamps | 108 | 16 h/8 h | C2 toxin | - | [333] [334] |
| Protoceratium reticulatum GG1AM | Stirred-PBR | Batch/Fed-batch /Semi-continuous | 15 L | Stirring (impeller)/gas sparging | 19 ± 1 | Cool-white fluorescent lamps | 242–766 | - | Yessotoxin | - | [290] |
| Protoceratium reticulatum GG1AM and VGO764 | Stirred-glass fermenter | Semi-continuous Fed-batch Continuous | 2 L | Stirring (impeller) | - | Cool-white fluorescent lamps | (e.g., 34, 44, 54, 66, 81, 99, 100) | - | Yessotoxin | - | [335] [270] |
| Amphidinium carterae ACRN03 | Airlift bubble column PBR | Semi-continuous | 540 L 320 L 48 L | Compressed air (continuous) | 23.3 ± 2.2 | - | 158 ± 22 (indoor) ~464 (outdoor) | 18 h/ 6h | N/A | Production of biomass | [289] |
| Karenia selliformes CAWD79 Alexandrium ostenfeldii CAWD135 Karenia Brevisulcata CAWD82 | Column PBRs Carboy | Batch/ Continuous | 5.4 L 52 L | Magnetic stirring/Air bubbling (low) | - | Cool-white fluorescent lamps | 38 | 12 h/12 h | Gymnodimine Spirolide Brevisulcatic acid | - | [268] |
| Karlodinium veneficum K10 | Bubble column PBR | Batch (sequential) | 80 L | Gas Sparging | 21 ± 1 | LEDs | 220 1500 (>7th day) | 12 h/12 h | N/A | - | [287] |
| Amphidinium sp. | Flat-bottom flask | Batch | 3 L | - | 25 | - | 108 | 16 h/8 h | Amphidinolide | - | [248] |
| Alexandrium minutum AMAD06 and AMAD16 | Alveolar panel PBR | Batch/Semi-continuous | 4 L | Alveoli | - | Cool-white fluorescent lamps | 100 | 12 h/12 h | Gonyautoxin/fraction of saxitoxin, neosaxitoxin and C-toxin | - | [336] |
| Alexandrium ostenfeldii CCMP1773 | Flat-bottom vessel | Batch | 8 L | N/A | 16 | Cool-white fluorescent lamps | 155 | 12 h/12 h | Spirolide | - | [337] |
| Alexandrium ostenfeldii CCMP1773 | Column PBR | Continuous | 100 L | Stirring (paddle impeller turbines) | 18 | Cool-white fluorescent lamps | 190 | 16 h/8 h | Spirolide | - | [337] |
| Alexandrium pacificum HYM9704 | Chemostat system | Batch Semi-continuous | 2 L | Air bubbling | 15 | Cool-white fluorescent lamps | 150 | 12/12 | C-toxin Gonyatoxin Saxitoxin | - | [338] |
| Amphidinium carterae (Hulbert) | Plastic cylinder | Batch | 40 L | Airlift | 22 | Cool-white fluorescent lamps | 40 | 12 h/12 h | N/A | Identification of pharmacological activity in vitro using algal extracts | [339] |
| Prorocentrum lima CCMP 2579 | Vertical flat PBR | Batch | 100 L | Air bubbling | 20 | Cool-white fluorescent lamps | 100 | 12 h/12 h | Okadaic acid Dinophysistoxins | - | [321] |
| Karlodinium veneficum ICMB 252 Alexandrium. minutum AMP4 | Column PBR | Batch (indoors) Semi-continuous (outdoors) | 350 L | Air bubbling | 20 ± 1 (indoors) Variable (outdoors) | Cool-white fluorescent lamps | 110 (indoors) 202–4020 (outdoors) | 12 h/12 h | N/A | Each PBR column size ~35 L each Production of lipids | [340] |
| Symbiodinium voratum | Twin Layer PBR (Biofilm-immobilization) | Continuous | 414 cm2 | Air tube (continuous) | 23 ± 1 25 ± 3 | Cool-white fluorescent and Sodium lamps | 26 ± 2 73.5 ± 17.5 | 14 h/10 h | N/A | Production of peridin | [341] |
| Amphidinium carterae (JHWAC) Symbiodinium sp. (JHLSD1) Prorocentrum rathymum (JHWPMX1) | Vertical column PBR system (12× column) | Batch | 700 L | Air bubbling | 20 ± 1 | Cool-white fluorescent lamps | 40–50 | 12 h/12 h | N/A | Each PBR column size ~60 L each Screening antioxidant properties | [342] |
| Karlodinium veneficum (CCMP 2936) | Vertical column PBR | Batch | 31 L | N/A | 24 | LEDs | 50 | 14 h/10 h | N/A | Study on vertical migration | [343] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assunção, J.; Guedes, A.C.; Malcata, F.X. Biotechnological and Pharmacological Applications of Biotoxins and Other Bioactive Molecules from Dinoflagellates. Mar. Drugs 2017, 15, 393. https://doi.org/10.3390/md15120393
Assunção J, Guedes AC, Malcata FX. Biotechnological and Pharmacological Applications of Biotoxins and Other Bioactive Molecules from Dinoflagellates. Marine Drugs. 2017; 15(12):393. https://doi.org/10.3390/md15120393
Chicago/Turabian StyleAssunção, Joana, A. Catarina Guedes, and F. Xavier Malcata. 2017. "Biotechnological and Pharmacological Applications of Biotoxins and Other Bioactive Molecules from Dinoflagellates" Marine Drugs 15, no. 12: 393. https://doi.org/10.3390/md15120393
APA StyleAssunção, J., Guedes, A. C., & Malcata, F. X. (2017). Biotechnological and Pharmacological Applications of Biotoxins and Other Bioactive Molecules from Dinoflagellates. Marine Drugs, 15(12), 393. https://doi.org/10.3390/md15120393