
5-Episinuleptolide Decreases the Expression of the Extracellular Matrix in Early Biofilm Formation of Multi-Drug Resistant Acinetobacter baumannii


 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Efficacy of 5-Episinuleptolide against A. baumannii Biofilm Formation Rather Than Eradication of Biofilm
2.2. Antimicrobial Susceptibility Testing of Multi-Drug Resistant A. baumannii Isolates
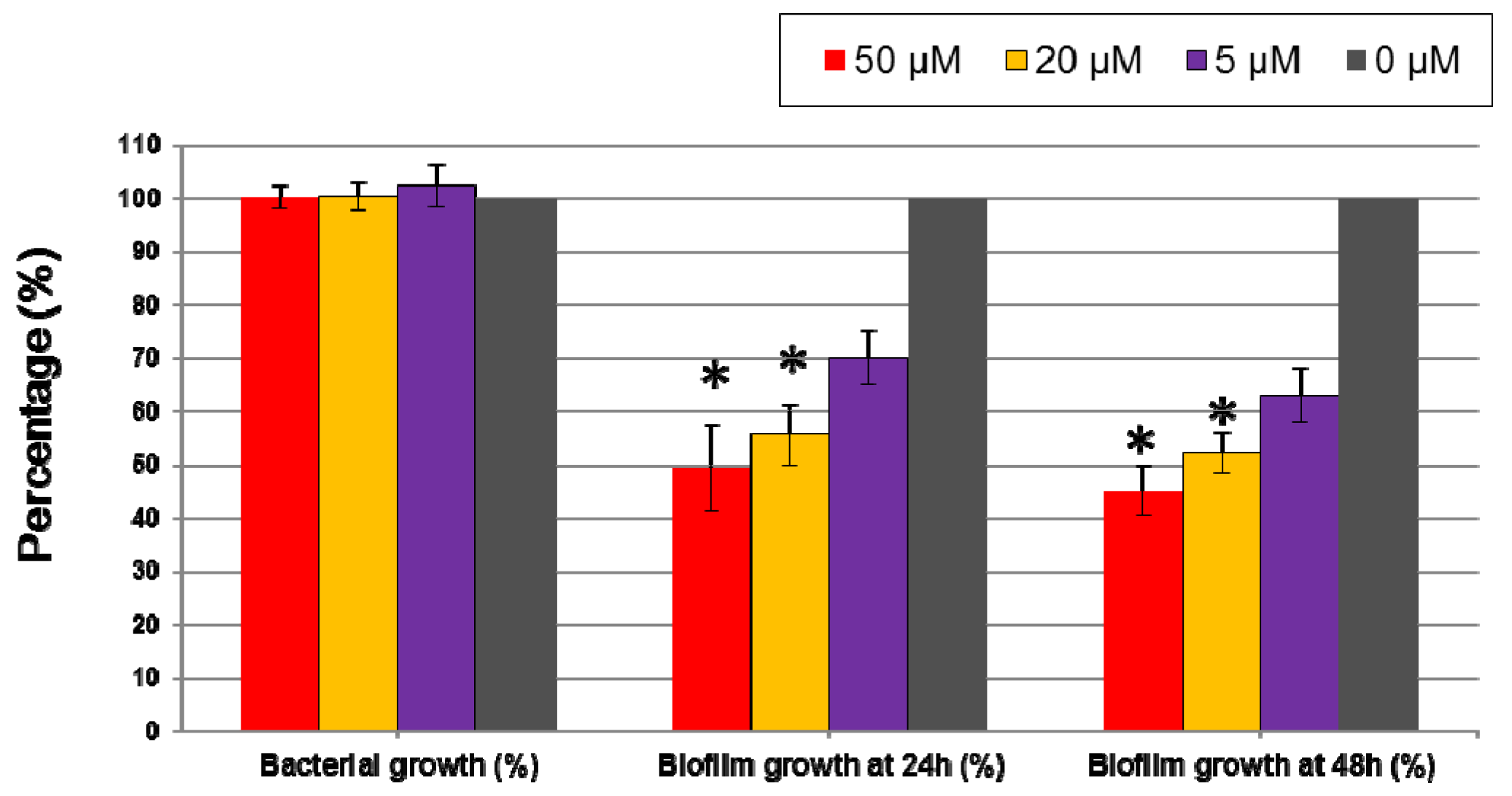
2.3. Inhibition of Biofilm Formation by 5-Episinuleptolide in Multi-Drug Resistant A. baumannii Clinical Isolates and Reference Strains
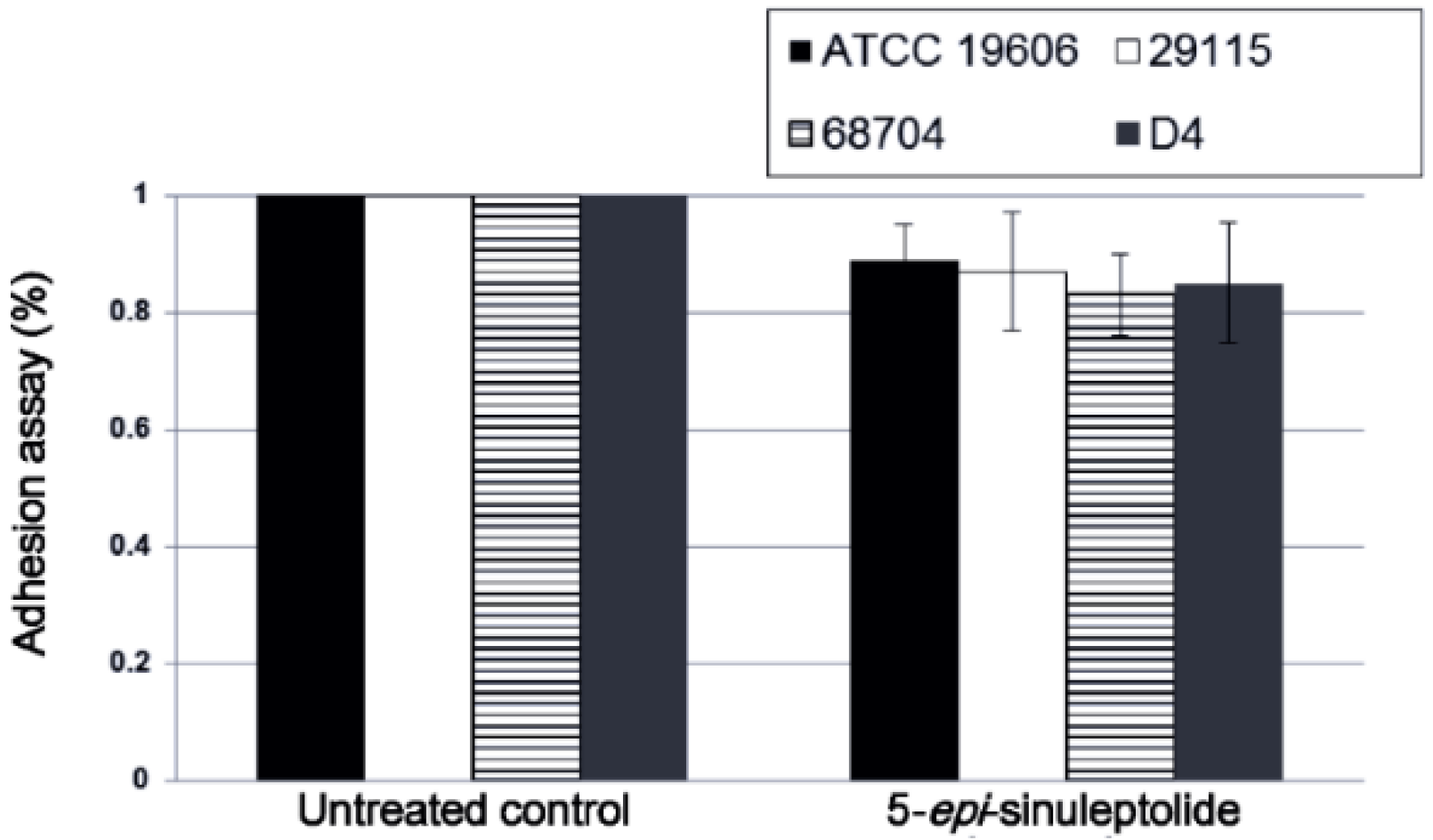
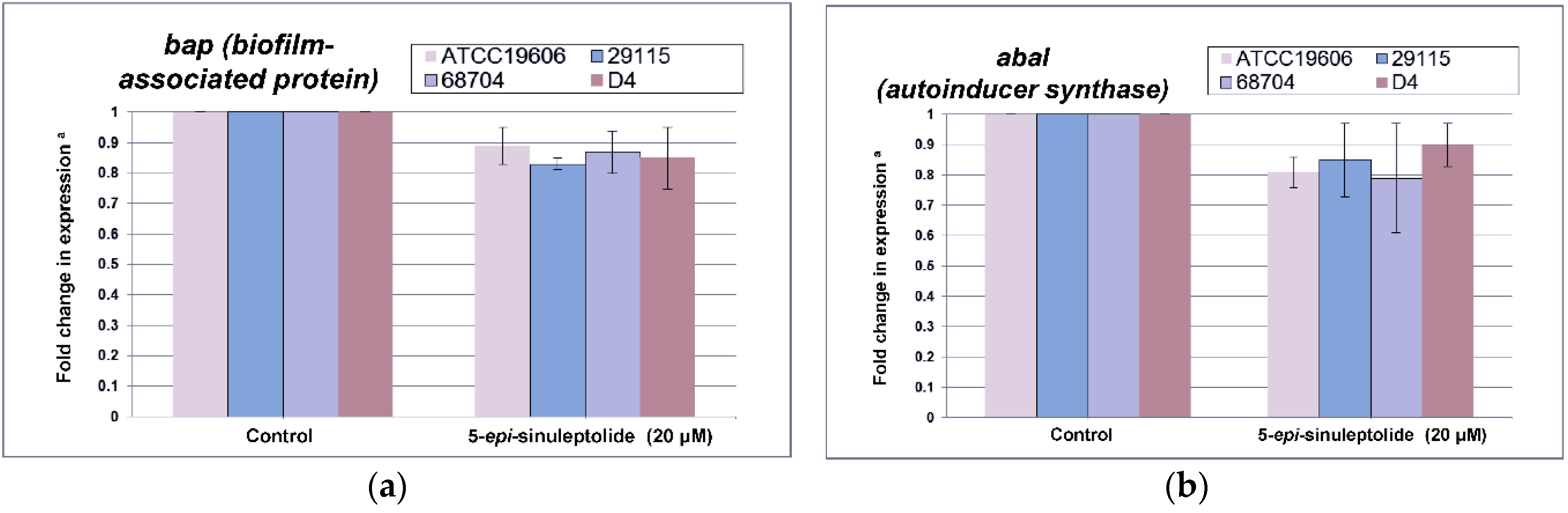
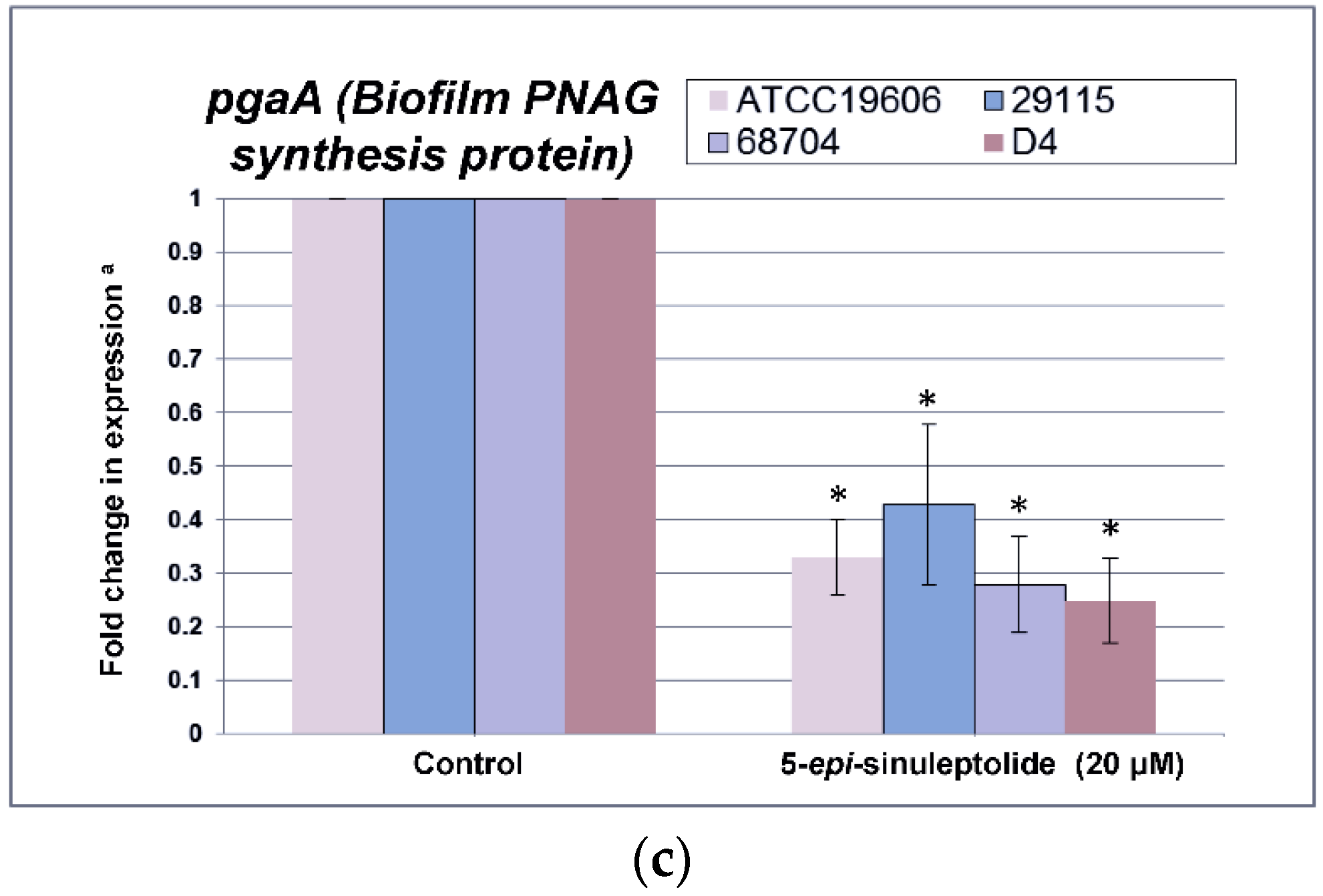
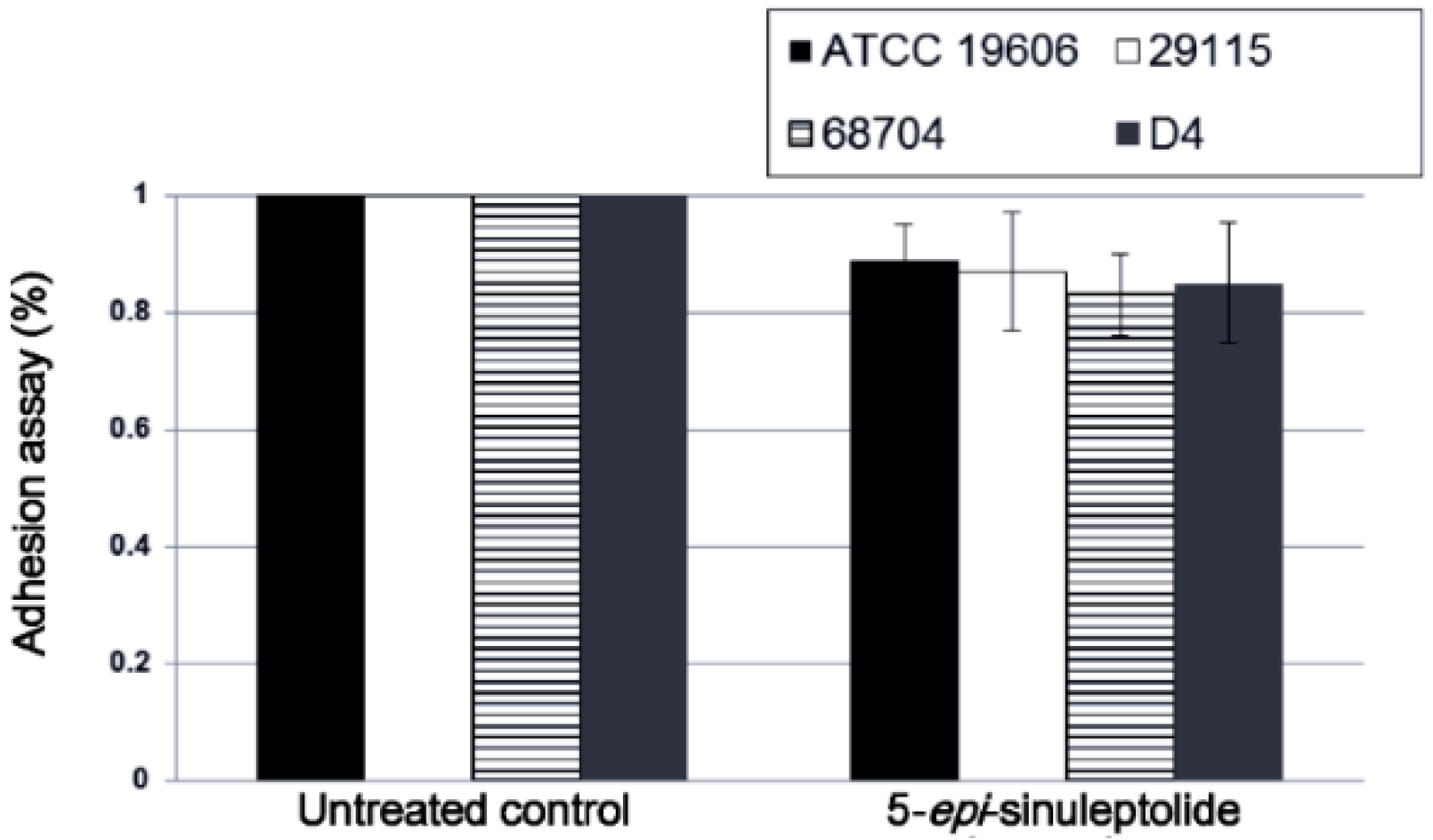
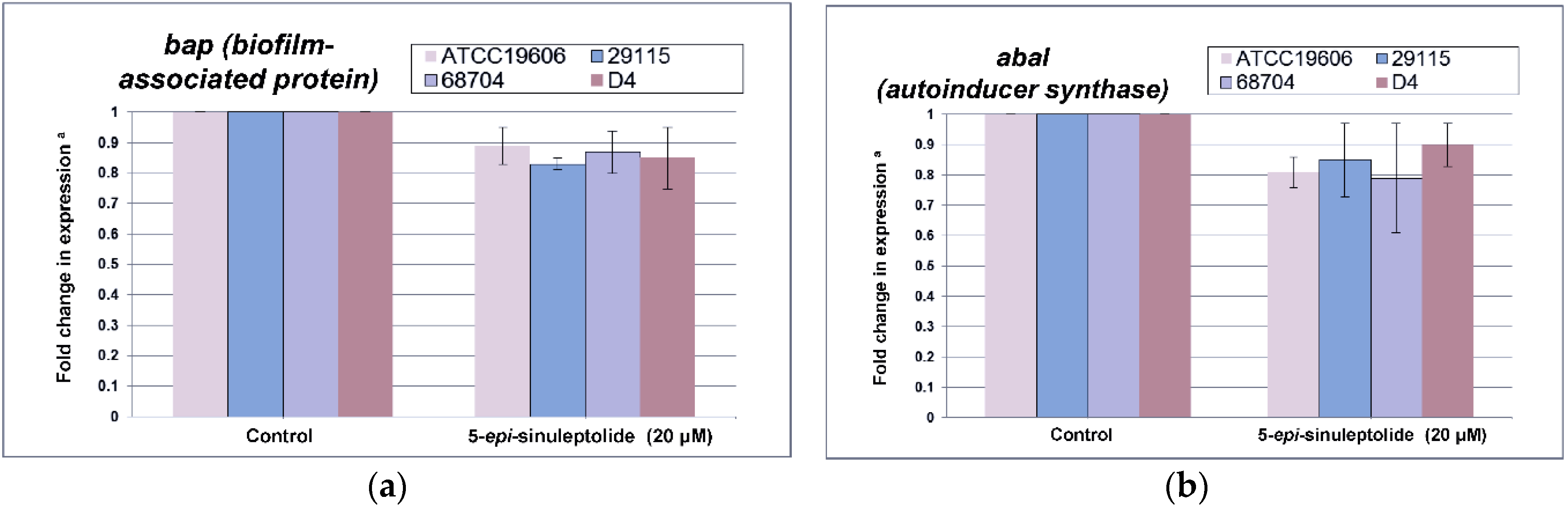
2.4. Investigation of the Biofilm Inhibitory Mechanism of 5-Episinuleptolide
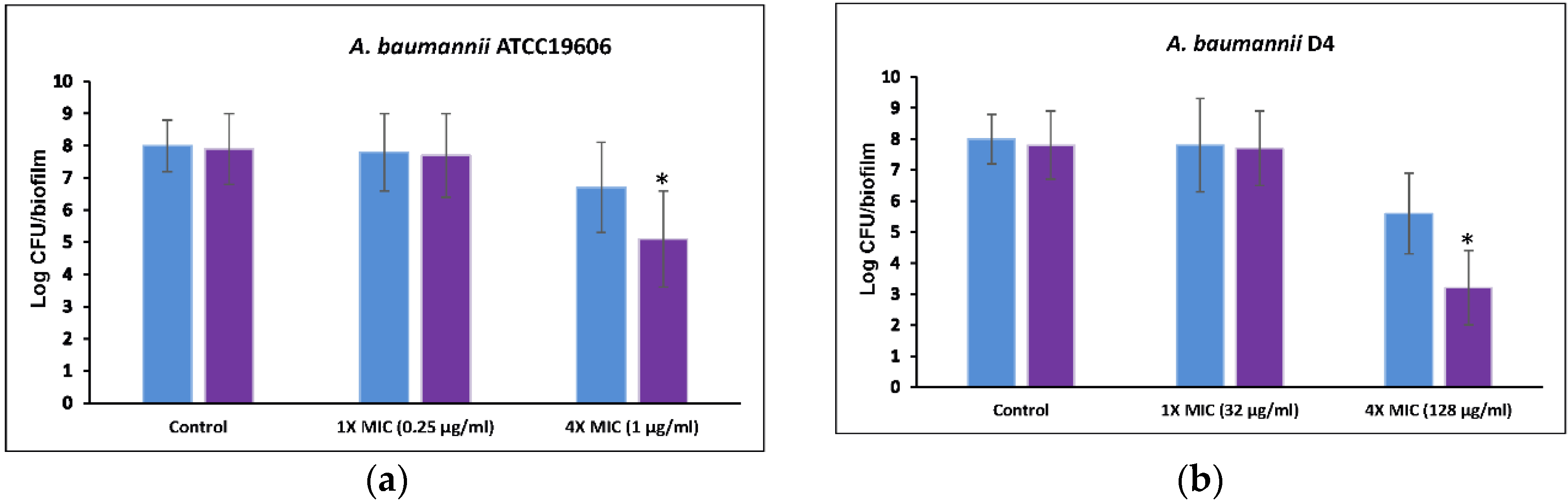
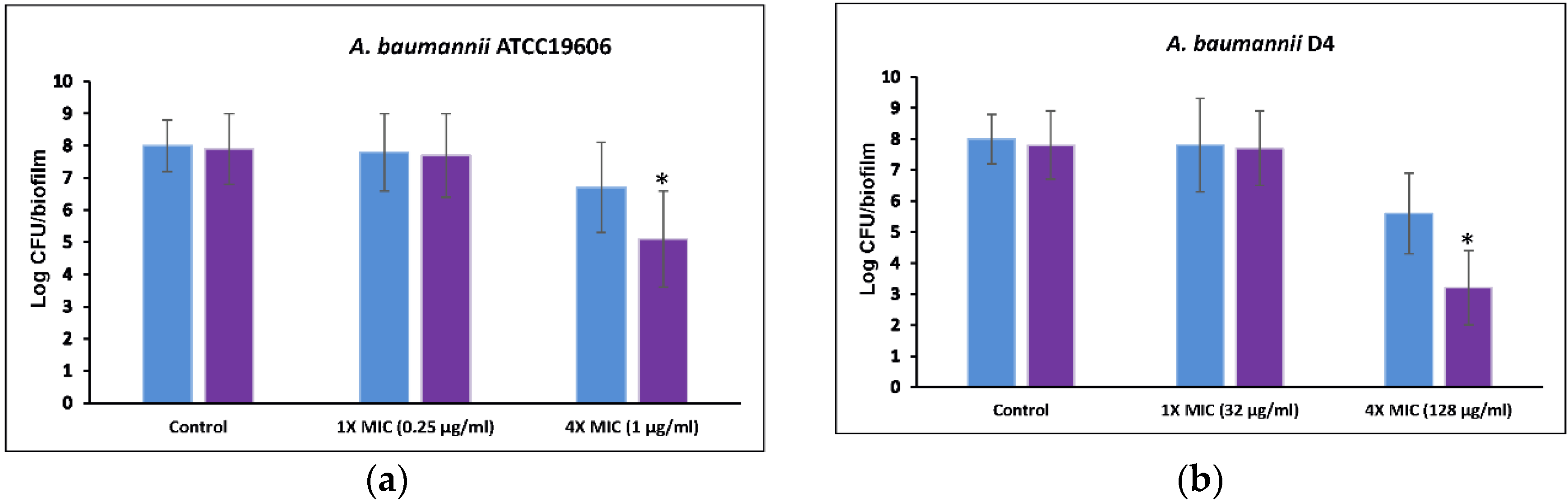
2.5. Anti-Biofilm Efficacy of a Combination of 5-Episinuleptolide with Levofloxacin
3. Discussion
4. Materials and Methods
4.1. Marine Natural Products
4.2. Bacterial Isolates
4.3. Ethics Statement
4.4. Antimicrobial Susceptibility Testing
4.5. Measurement of Bacterial Growth
4.6. Adhesion and Biofilm Formation Assays
4.7. Biofilm Eradication Assay
4.8. RNA Extraction and Synthesis of cDNA
4.9. Quantitative Reverse Transcription-PCR (qRT-PCR) for Biofilm-Related Genes
4.10. Analysis of Biofilm Formation by Scanning Electron Microscopy
4.11. Anti-Biofilm Efficacy of 5-Episinuleptolide in Combination with Levofloxacin
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Price, L.S.; Arheart, K.; Nordmann, P.; Boulanger, A.E.; Cleary, T.; Alvarez, R.; Pizano, L.; Namias, N.; Kett, D.H.; Poirel, L. Eighteen years of experience with Acinetobacter baumannii in a tertiary care hospital. Crit. Care Med. 2013, 41, 2733–2742. [Google Scholar] [CrossRef] [PubMed]
- Karageorgopoulos, D.E.; Falagas, M.E. Current control and treatment of multidrug-resistant Acinetobacter baumannii infections. Lancet Infect. Dis. 2008, 8, 751–762. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, F.R.; Keiser, P.; Craft, D.W.; Gage, F.; Robson, M.; Brown, T.S.; Petersen, K.; Sincock, S.; Kasper, M.; Hawksworth, J.; et al. The majority of us combat casualty soft-tissue wounds are not infected or colonized upon arrival or during treatment at a continental us military medical facility. Am. J. Surg. 2010, 200, 489–495. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Nunez, C.; Reffuveille, F.; Fernandez, L.; Hancock, R.E. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.H.; Slamti, L.; Avci, F.Y.; Pier, G.B.; Maira-Litran, T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-β-1-6-N-acetylglucosamine, which is critical for biofilm formation. J. Bacteriol. 2009, 191, 5953–5963. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.; Fischer, W.; Krokotsch, A.; Leopold, K.; Hartmann, R.; Egge, H.; Laufs, R. The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a linear β-1,6-linked glucosaminoglycan: Purification and structural analysis. J. Bacteriol. 1996, 178, 175–183. [Google Scholar] [PubMed]
- Maira-Litran, T.; Kropec, A.; Abeygunawardana, C.; Joyce, J.; Mark, G., 3rd; Goldmann, D.A.; Pier, G.B. Immunochemical properties of the Staphylococcal poly-N-acetylglucosamine surface polysaccharide. Infect. Immun. 2002, 70, 4433–4440. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Preston, J.F., 3rd; Romeo, T. The pgaABCD locus of Escherichia coli promotes the synthesis of a polysaccharide adhesin required for biofilm formation. J. Bacteriol. 2004, 186, 2724–2734. [Google Scholar] [CrossRef] [PubMed]
- Vuong, C.; Voyich, J.M.; Fischer, E.R.; Braughton, K.R.; Whitney, A.R.; DeLeo, F.R.; Otto, M. Polysaccharide intercellular adhesin (PIA) protects Staphylococcus epidermidis against major components of the human innate immune system. Cell. Microbiol. 2004, 6, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Koh, Y.M.; Kim, J.; Lee, J.C.; Lee, Y.C.; Seol, S.Y.; Cho, D.T.; Kim, J. Capacity of multidrug-resistant clinical isolates of Acinetobacter baumannii to form biofilm and adhere to epithelial cell surfaces. Clin. Microbiol. Infect. 2008, 14, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, I.; Penades, J.R. Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef] [PubMed]
- Hinsa, S.M.; Espinosa-Urgel, M.; Ramos, J.L.; O′Toole, G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 2003, 49, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Latasa, C.; Roux, A.; Toledo-Arana, A.; Ghigo, J.M.; Gamazo, C.; Penades, J.R.; Lasa, I. BapA, a large secreted protein required for biofilm formation and host colonization of Salmonella enterica serovar enteritidis. Mol. Microbiol. 2005, 58, 1322–1339. [Google Scholar] [CrossRef] [PubMed]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect. Immun. 2012, 80, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Loehfelm, T.W.; Luke, N.R.; Campagnari, A.A. Identification and characterization of an Acinetobacter baumannii biofilm-associated protein. J. Bacteriol. 2008, 190, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Lebeaux, D.; Ghigo, J.M.; Beloin, C. Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Penesyan, A.; Gillings, M.; Paulsen, I.T. Antibiotic discovery: Combatting bacterial resistance in cells and in biofilm communities. Molecules 2015, 20, 5286–5298. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Yamashita, A. Pharmacological studies on the root bark of paeonia moutan. I. Central effects of paeonol. Yakugaku Zasshi 1969, 89, 1205–1211. [Google Scholar] [PubMed]
- Piao, X.; Piao, X.L.; Kim, H.Y.; Cho, E.J. Antioxidative activity of geranium (Pelargonium inquinans ait) and its active component, 1,2,3,4,6-Penta-O-galloyl-β-d-glucopyranose. Phytother. Res. 2008, 22, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.H.; Chang, F.R.; Hua, M.Y.; Wu, Y.C.; Liu, S.T. Inhibitory effects of 1,2,3,4,6-Penta-O-galloyl-β-d-glucopyranose on biofilm formation by Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Pinkston, K.L.; Singh, K.V.; Gao, P.; Wilganowski, N.; Robinson, H.; Ghosh, S.; Azhdarinia, A.; Sevick-Muraca, E.M.; Murray, B.E.; Harvey, B.R. Targeting pili in enterococcal pathogenesis. Infect. Immun. 2014, 82, 1540–1547. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Zhang, Y.; Yang, Y.; Wang, L. Isolation of the autoinducer-quenching strain that inhibits LasR in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2014, 15, 6328–6342. [Google Scholar] [CrossRef] [PubMed]
- Chabane, Y.N.; Mlouka, M.B.; Alexandre, S.; Nicol, M.; Marti, S.; Pestel-Caron, M.; Vila, J.; Jouenne, T.; de, E. Virstatin inhibits biofilm formation and motility of Acinetobacter baumannii. BMC Microbiol. 2014, 14, 62. [Google Scholar] [CrossRef] [PubMed]
- Sawadogo, W.R.; Boly, R.; Cerella, C.; Teiten, M.H.; Dicato, M.; Diederich, M. A survey of marine natural compounds and their derivatives with anti-cancer activity reported in 2012. Molecules 2015, 20, 7097–7142. [Google Scholar] [CrossRef] [PubMed]
- Rinehart, K.L., Jr.; Gloer, J.B.; Hughes, R.G., Jr.; Renis, H.E.; McGovren, J.P.; Swynenberg, E.B.; Stringfellow, D.A.; Kuentzel, S.L.; Li, L.H. Didemnins: Antiviral and antitumor depsipeptides from a caribbean tunicate. Science 1981, 212, 933–935. [Google Scholar] [CrossRef] [PubMed]
- Liaw, C.C.; Chen, P.C.; Shih, C.J.; Tseng, S.P.; Lai, Y.M.; Hsu, C.H.; Dorrestein, P.C.; Yang, Y.L. Vitroprocines, new antibiotics against Acinetobacter baumannii, discovered from marine Vibrio sp. QWI-o6 using mass-spectrometry-based metabolomics approach. Sci. Rep. 2015, 5, 12856. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.C.; Sung, P.J.; Duh, C.Y.; Chen, B.W.; Sheu, J.H.; Yang, N.S. Anti-inflammatory activities of natural products isolated from soft corals of Taiwan between 2008 and 2012. Mar. Drugs 2013, 11, 4083–4126. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.H.; Ahmed, A.F.; Shiue, R.T.; Dai, C.F.; Kuo, Y.H. Scabrolides A–D, four new norditerpenoids isolated from the soft coral Sinularia scabra. J. Nat. Prod. 2002, 65, 1904–1908. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.F.; Shiue, R.-T.; Wang, G.-H.; Dai, C.-F.; Kuo, Y.-H.; Sheu, J.-H. Five novel norcembranoids from Sinularia leptoclados and S. parva. Tetrahedron 2003, 59, 7337–7344. [Google Scholar] [CrossRef]
- Tseng, Y.J.; Ahmed, A.F.; Dai, C.F.; Chiang, M.Y.; Sheu, J.H. Sinulochmodins A–C, three novel terpenoids from the soft coral Sinularia lochmodes. Org. Lett. 2005, 7, 3813–3816. [Google Scholar] [CrossRef] [PubMed]
- Takaki, H.; Koganemaru, R.; Iwakawa, Y.; Higuchi, R.; Miyamoto, T. Inhibitory effect of norditerpenes on LPS-induced TNF-α production from the okinawan soft coral, Sinularia sp. Biol. Pharm. Bull. 2003, 26, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, N.; Sharma, P.; Capalash, N. Quorum sensing in Acinetobacter: An emerging pathogen. Crit. Rev. Microbiol. 2010, 36, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Gaddy, J.A.; Actis, L.A. Regulation of Acinetobacter baumannii biofilm formation. Future Microbiol. 2009, 4, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Khodaverdian, V.; Pesho, M.; Truitt, B.; Bollinger, L.; Patel, P.; Nithianantham, S.; Yu, G.; Delaney, E.; Jankowsky, E.; Shoham, M. Discovery of antivirulence agents against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 3645–3652. [Google Scholar] [CrossRef] [PubMed]
- Monnappa, A.K.; Dwidar, M.; Seo, J.K.; Hur, J.H.; Mitchell, R.J. Bdellovibrio bacteriovorus inhibits Staphylococcus aureus biofilm formation and invasion into human epithelial cells. Sci. Rep. 2014, 4, 3811. [Google Scholar] [CrossRef] [PubMed]
- Peach, K.C.; Cheng, A.T.; Oliver, A.G.; Yildiz, F.H.; Linington, R.G. Discovery and biological characterization of the auromomycin chromophore as an inhibitor of biofilm formation in Vibrio cholerae. Chembiochem 2013, 14, 2209–2215. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, M.; Riedel, K.; Rasmussen, T.B.; Heydorn, A.; Andersen, J.B.; Parsek, M.R.; Rice, S.A.; Eberl, L.; Molin, S.; Hoiby, N.; et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 2002, 148, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.B.; Manefield, M.; Andersen, J.B.; Eberl, L.; Anthoni, U.; Christophersen, C.; Steinberg, P.; Kjelleberg, S.; Givskov, M. How Delisea pulchra furanones affect quorum sensing and swarming motility in Serratia liquefaciens MG1. Microbiology 2000, 146, 3237–3244. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Bedzyk, L.A.; Setlow, P.; England, D.F.; Kjelleberg, S.; Thomas, S.M.; Ye, R.W.; Wood, T.K. Differential gene expression to investigate the effect of (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone on Bacillus subtilis. Appl. Environ. Microbiol. 2004, 70, 4941–4949. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.H.; Wang, G.H.; Chou, T.H.; Wang, S.H.; Lin, R.J.; Chan, L.P.; So, E.C.; Sheu, J.H. 5-epi-sinuleptolide induces cell cycle arrest and apoptosis through tumor necrosis factor/mitochondria-mediated caspase signaling pathway in human skin cancer cells. Biochim. Biophys. Acta 2012, 1820, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Fey, P.D.; Olson, M.E. Current concepts in biofilm formation of Staphylococcus epidermidis. Future Microbiol. 2010, 5, 917–933. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Clemmer, K.M.; Bonomo, R.A.; Rather, P.N. Isolation and characterization of an autoinducer synthase from Acinetobacter baumannii. J. Bacteriol. 2008, 190, 3386–3392. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Cheong, H.J.; Noh, J.Y.; Kim, W.J. In vitro comparison of anti-biofilm effects against carbapenem-resistant Acinetobacter baumannii: Imipenem, colistin, tigecycline, rifampicin and combinations. Infect. Chemother. 2015, 47, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.P.; Lin, Y.T.; Tsai, J.C.; Hung, W.C.; Chen, H.J.; Chen, P.F.; Hsueh, P.R.; Teng, L.J. Genotypes and phenotypes of Staphylococcus lugdunensis isolates recovered from bacteremia. J. Microbiol. Immunol. Infect. 2015, 48, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H.J.; Coenye, T. In vitro activity of ceftazidime, ciprofloxacin, meropenem, minocycline, tobramycin and trimethoprim/sulfamethoxazole against planktonic and sessile Burkholderia cepacia complex bacteria. J. Antimicrob. Chemother. 2009, 64, 801–809. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics a | A. baumannii Isolates | ||||
|---|---|---|---|---|---|
| ATCC 19606 | BAA747 | 29115 | 68704 | D4 | |
| SXT | >8/152 (R) | <1/19 (S) | >8/152 (R) | >8/152 (R) | >8/152 (R) |
| DOX | <2 (S) | <2 (S) | 64 (R) | 64 (R) | 64 (R) |
| TIM | 128/2 (R) | 16/2 (S) | 32/2 (I) | 32/2 (I) | 256/2 (R) |
| CT | <1 (S) | <1 (S) | <1 (S) | <1 (S) | <1 (S) |
| CAZ | 16 (I) | 16 (I) | >128 (R) | >128 (R) | >128 (R) |
| MEM | <2 (S) | <2 (S) | 32 (R) | 64 (R) | >128 (R) |
| GM | 32 (R) | <2 (S) | >128 (R) | >128 (R) | >128 (R) |
| LEV | <1 (S) | <1 (S) | 16 (R) | 32 (R) | 32 (R) |
| TIC | 128 (R) | 16 (S) | 32 (I) | 32 (I) | 128 (R) |
| Strain | Bacterial Growth (%) | Biofilm Growth (%) | ||||
|---|---|---|---|---|---|---|
| 50 μM | 20 μM | 5 μM | 50 μM | 20 μM | 5 μM | |
| A. baumannii | ||||||
| ATCC 19606 | 100.2 ± 2.1 | 100.4 ± 2.7 | 102.4 ± 4.1 | 49.38 ± 8.1 * | 55.58 ± 5.6 * | 70.02 ± 5.0 |
| 29115 | 96.2 ± 2.3 | 100.3 ± 4.4 | 103.1 ± 4.8 | 59.90 ± 6.1 * | 61.07 ± 4.2 * | 76.81 ± 8.7 |
| 68704 | 105.8 ± 7.2 | 98.4 ± 6.2 | 103.9 ± 4.9 | 45.83 ± 2.2 * | 87.13 ± 5.4 | 96.33 ± 6.0 |
| D4 | 102.4 ± 2.9 | 98.6 ± 2.3 | 97.7 ± 5.5 | 43.90 ± 3.8 * | 60.23 ± 4.6 * | 78.75 ± 6.7 |
| E. coli | ||||||
| ATCC 25922 | 99.5 ± 6.2 | 98.5 ± 3.7 | 100.4 ± 8.2 | 45.48 ± 7.7 * | 50.18 ± 9.4 * | 82.33 ± 7.1 |
| P. aeruginosa | ||||||
| ATCC 27853 | 100.5 ± 7.6 | 108.9 ± 6.1 | 102.8 ± 5.7 | 53.01 ± 3.8 * | 77.82 ± 5.5 | 95.48 ± 9.2 |
| S. aureus | ||||||
| ATCC 29213 | 103.3 ± 9.3 | 99.7 ± 5.6 | 105.3 ± 9.2 | 90.25 ± 7.4 | 99.57 ± 6.8 | 103.82 ± 5.6 |
| S. epidermidis | ||||||
| RP62A | 99.7 ± 9.5 | 99.4 ± 2.8 | 98.7 ± 7.4 | 88.98 ± 7.9 | 113.06 ± 10.1 | 116.19 ± 8.1 |
| Strain | Biofilm Growth Without 0.5× MIC (%) | Biofilm Growth With 0.5× MIC (%) | ||||
|---|---|---|---|---|---|---|
| 50 μM | 20 μM | 5 μM | 50 μM | 20 μM | 5 μM | |
| A. baumannii | ||||||
| ATCC 19606 | 49.38 ± 8.1 * | 55.58 ± 5.6 * | 70.02 ± 5.0 | 25.62 ± 7.3 * | 31.83 ± 4.4 * | 67.92 ± 8.4 |
| 29115 | 59.90 ± 6.1 * | 61.07 ± 4.2 * | 76.81 ± 8.7 | 33.46 ± 7.7 * | 40.58 ± 8.7 * | 69.15 ± 5.7 |
| 68704 | 45.83 ± 2.2 * | 87.13 ± 5.4 | 96.33 ± 6.0 | 27.47 ± 6.8 * | 47.46 ± 3.6 * | 79.57 ± 8.6 |
| D4 | 43.90 ± 3.8 * | 60.2 3± 4.6 * | 78.75 ± 6.7 | 26.35 ± 8.4 * | 35.19 ± 6.3 * | 71.73 ± 7.7 |
| Target | Primer Name | Sequence (5′ to 3′) | Sizes (bp) | Reference |
|---|---|---|---|---|
| 16SrDNA | 16S-F | TCCTCCAGTTTGTCACTGGC | 116 | This study |
| 16S-R | GTCAGCTCGTGTCGTGAGAT | |||
| bap | BAP-F | CCTTGGTAACCACAGAGGGA | 114 | This study |
| BAP-R | TGACTGCATTGGTACCCTCC | |||
| pgaA | PGA-F | GCTGAAGCTCAAGATGTGGC | 91 | This study |
| PGA-R | ATGCAACCCGTACCAACTGA | |||
| abaI | abaI-F abaI-R | GTACAGTCGACGTATTTGTTGAATATTTGGG CGTACGTCTAGAGTAATGAGTTGTTTTGCGCC | 382 | [44] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, S.-P.; Hung, W.-C.; Huang, C.-Y.; Lin, Y.-S.; Chan, M.-Y.; Lu, P.-L.; Lin, L.; Sheu, J.-H. 5-Episinuleptolide Decreases the Expression of the Extracellular Matrix in Early Biofilm Formation of Multi-Drug Resistant Acinetobacter baumannii. Mar. Drugs 2016, 14, 143. https://doi.org/10.3390/md14080143
Tseng S-P, Hung W-C, Huang C-Y, Lin Y-S, Chan M-Y, Lu P-L, Lin L, Sheu J-H. 5-Episinuleptolide Decreases the Expression of the Extracellular Matrix in Early Biofilm Formation of Multi-Drug Resistant Acinetobacter baumannii. Marine Drugs. 2016; 14(8):143. https://doi.org/10.3390/md14080143
Chicago/Turabian StyleTseng, Sung-Pin, Wei-Chun Hung, Chiung-Yao Huang, Yin-Shiou Lin, Min-Yu Chan, Po-Liang Lu, Lin Lin, and Jyh-Horng Sheu. 2016. "5-Episinuleptolide Decreases the Expression of the Extracellular Matrix in Early Biofilm Formation of Multi-Drug Resistant Acinetobacter baumannii" Marine Drugs 14, no. 8: 143. https://doi.org/10.3390/md14080143
APA StyleTseng, S.-P., Hung, W.-C., Huang, C.-Y., Lin, Y.-S., Chan, M.-Y., Lu, P.-L., Lin, L., & Sheu, J.-H. (2016). 5-Episinuleptolide Decreases the Expression of the Extracellular Matrix in Early Biofilm Formation of Multi-Drug Resistant Acinetobacter baumannii. Marine Drugs, 14(8), 143. https://doi.org/10.3390/md14080143