Abstract
Cyanobacteria from underexplored and extreme habitats are attracting increasing attention in the search for new bioactive substances. However, cyanobacterial communities from tropical and subtropical regions are still largely unknown, especially with respect to metabolite production. Among the structurally diverse secondary metabolites produced by these organisms, peptides are by far the most frequently described structures. In this work, liquid chromatography/electrospray ionization coupled to high resolution quadrupole time-of-flight tandem mass spectrometry with positive ion detection was applied to study the peptide profile of a group of cyanobacteria isolated from the Southeastern Brazilian coastal forest. A total of 38 peptides belonging to three different families (anabaenopeptins, aeruginosins, and cyanopeptolins) were detected in the extracts. Of the 38 peptides, 37 were detected here for the first time. New structural features were proposed based on mass accuracy data and isotopic patterns derived from full scan and MS/MS spectra. Interestingly, of the 40 surveyed strains only nine were confirmed to be peptide producers; all of these strains belonged to the order Nostocales (three Nostoc sp., two Desmonostoc sp. and four Brasilonema sp.).
1. Introduction
Cyanobacteria represent a rich source of bioactive compounds, many of which remain unexplored. To date, antibacterial, antiviral, antifungal, anticancer, immunosuppressive, protease inhibitory, and other pharmacological properties have been described for the secondary metabolites isolated from these microorganisms [1,2,3,4,5,6,7]. Structurally, these compounds are primarily of a peptide nature [1,3,8] and are mainly synthesized via non-ribosomal peptide synthetases (NRPS) or a combination of NRPS and the polyketide synthetase (NRPS/PKS) [3]. Most of these cyanopeptides can be grouped into seven families based on conserved substructures [3]: anabaenopeptins, cyanopeptolins, cyclamides, microcystins, microginins, aeruginosins, and microviridins. Particular interest arises from cyanobacteria growing in extreme environments, since these underexplored habitats may harbor a microbial diversity able to synthesize a unique variety of secondary metabolites [9].
The Atlantic Forest is a continental biome that extends primarily along the Atlantic coast of Brazil and also includes small portions of Argentina and Paraguay [10,11]. Considered one of the richest regions in the world in terms of biodiversity and endemism, this forest is also one of the most threatened regions [12]. With respect to the microorganisms that inhabit this region, little is known [13]. Particularly, referring to cyanobacteria, the available information is typically focused on diversity and descriptions of species [14,15,16,17] and scarce information about secondary metabolite production is available [18]. Cyanobacteria inhabiting the phyllosphere (leaf surface) are exposed to hostile conditions such as low availability of nutrients, large temperature range, osmotic stress, and high incidence of ultraviolet rays [19], representing a little explored community of extremophilic organisms with potential for production of novel bioactive compounds.
Due to the development in instrumentation and technology, mass spectrometry (MS) has rapidly become a fundamental tool for characterizing large biomolecules such as proteins and peptides [20,21]. The development of electrospray ionization (ESI) and matrix-assisted laser desorption/ionization (MALDI) in conjunction with the introduction of multi-stage and hybrid analyzers was decisive for the success of MS analysis of biomolecules [20]. Time-of-flight (TOF) is one type of mass analyzer that finds wide application in this field and can provide high-resolution spectra for protein and peptides. Particularly when combined with other types of mass analyzer, such as the quadrupole mass analyzer forming the hybrid quadrupole time-of-flight (QTOF), valuable peptide or amino acid sequence information can also be obtained. Liquid chromatography (LC) is a typical inlet method for MS analysis and provides an extra dimension of separation. LC can be coupled with ESI (online/offline) or MALDI (offline). Matrix-assisted laser desorption ionization-time-of-flight-mass spectrometry (MALDI-TOF-MS) has been employed in the identification of many known and new cyanopeptides directly from cyanobacteria [22,23,24,25,26]. Applications of liquid chromatography (LC) coupled to quadrupole time-of-flight mass spectrometry (QTOF-MS) have also been reported [27,28,29].
In this study, the peptide profiles of 40 strains of cyanobacteria isolated from the phyllosphere of four native species of plants in the coastal forest of Southeastern Brazil [30] (Table 1) were investigated using LC-ESI-Q-TOF-MS with positive ion detection and revealed a high production potential in the heterocytous strains. Accurate masses and isotope patterns for both precursor and product ions were used to propose the planar structures of the observed peptides.

Table 1.
Cyanobacteria included in the present study.
| Strain | Order | Family | Genus a | Source b |
|---|---|---|---|---|
| CENA353 | Chroococcales | Xenococcaceae | Chroococcidiopsis sp. | Mn-SV |
| CENA367 | Chroococcidiopsis sp. | Ee-SV | ||
| CENA351 | Hydrococcacceae | Pleurocapsa sp. | Mn-SV | |
| CENA350 | Pseudanabaenales | Pseudanabaenaceae | Leptolyngbya sp. | Mn-SV |
| CENA355 | Leptolyngbya sp. | Mn-SV | ||
| CENA359 | - | Gg-Pi | ||
| CENA364 | Leptolyngbya sp. | Gg-Pi | ||
| CENA370 | Oculatella sp. | Ee-SV | ||
| CENA374 | Leptolyngbya sp. | Ee-Pi | ||
| CENA372 | Leptolyngbya sp. | Ee-Pi | ||
| CENA375 | Leptolyngbya sp. | Ee-Pi | ||
| CENA377 | Leptolyngbya sp. | Ee-Pi | ||
| CENA378 | Leptolyngbya sp. | Ee-Pi | ||
| CENA384 | - | Go-Pi | ||
| CENA385 | - | Go-Pi | ||
| CENA387 | Leptolyngbya sp. | Go-Pi | ||
| CENA354 | Nostocales | Microchaetaceae | - | Mn-SV |
| CENA352 | Nostocaceae | Nostoc sp. | Mn-SV | |
| CENA356 | Nostoc sp. | Mn-SV | ||
| CENA357 | Nostoc sp. | Gg-Pi | ||
| CENA358 | Nostoc sp. | Gg-Pi | ||
| CENA362 | Desmonostoc sp. | Gg-Pi | ||
| CENA363 | Desmonostoc sp. | Gg-Pi | ||
| CENA365 | Desmonostoc sp. | Gg-Pi | ||
| CENA368 | Nostoc sp. | Ee-SV | ||
| CENA369 | Nostoc sp. | Ee-Pi | ||
| CENA371 | Desmonostoc sp. | Ee-SV | ||
| CENA373 | Nostoc sp. | Ee-SV | ||
| CENA376 | Nostoc sp. | Ee-Pi | ||
| CENA379 | Nostoc sp. | Ee-SV | ||
| CENA380 | Desmonostoc sp. | Ee-SV | ||
| CENA383 | Desmonostoc sp. | Go-Pi | ||
| CENA386 | Desmonostoc sp. | Go-Pi | ||
| CENA388 | Nostoc sp. | Go-Pi | ||
| CENA389 | Nostoc sp. | Go-Pi | ||
| CENA360 | Nostocales | Scytonemataceae | Brasilonema sp. | Gg-Pi |
| CENA361 | Brasilonema sp. | Gg-Pi | ||
| CENA366 | Brasilonema sp. | Gg-Pi | ||
| CENA381 | Brasilonema sp. | Ee-Pi | ||
| CENA382 | Brasilonema sp. | Ee-Pi |
a Identity criteria for the determination of genus was established in ≥95% when comparing with 16S rRNA genes between the surveyed cyanobacteria and the sequences deposited in the GenBank (NBCI). b Plant species: Ee: Euterpe edualis; Mn: Merostachys neesii; Gg: Garcinia gardneriana; Go: Guapira opposita. Localization Pi: Picinguaba; SV: Santa Virginia sites, in Parque Estadual da Serra do Mar, São Paulo, Brazil.
Table 2.
LC-QTOF data of the peptides from the hydromethanolic extracts of cyanobacteria isolated from the Brazilian Atlantic Forest.
| No | Compound a | RT (min) | [M + H]+ m/z | Molecular Formula b | Error (ppm) | mSigma | Detected in Strains CENA | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nostoc sp | Brasilonema sp | Desmonostoc sp | ||||||||||||||
| aeruginosins | ||||||||||||||||
| 1 | Hpla-Leu-(Hex)-OHChoi-Agma | 17.2 | 753.4042 | C35H55N6O12 | −1.7 | 13.2 | 352 | 360 | 381 | 382 | this study | |||||
| 2 | Hpla-Leu-(Hex)Choi-Agma | 18.0 | 767.3828 | C35H55N6O13 | −1.6 | 2.1 | 352 | 358 | 360 | 381 | 382 | this study | ||||
| 3 | Hpla-Leu-(Gluc)-OHChoi-Agma | 18.2 | 737.4044 | C35H55N6O11 | −1.2 | 2.2 | 352 | 358 | 360 | 381 | 382 | this study | ||||
| 4 | Hpla-Leu-(Hex,But)-OHChoi-Agma | 20.0 | 823.4470 | C39H63N6O13 | −2.7 | 1.0 | 352 | 358 | 360 | 381 | this study | |||||
| 5 | Hpla-Leu-(Gluc,But)-OHChoi-Agma | 20.6 | 837.4253 | C39H61N6O14 | 1.0 | 13.0 | 352 | 358 | 360 | this study | ||||||
| 6 | Hpla-Leu-(Hex,Hexan)-OHChoi-Agma | 22.5 | 851.4780 | C41H67N6O13 | 2.3 | 2.0 | 352 | 358 | 369 | 360 | 361 | 381 | 382 | this study | ||
| 7 | Hpla-Leu-(Gluc,Hexan)-OHChoi-Agma | 23.1 | 865.4560 | C41H65N6O14 | −0.8 | 2.8 | 352 | 358 | 369 | 360 | 361 | 381 | 382 | [37] | ||
| 8 | Hpla-NMeLeu-(Gluc,Hexan)-OHChoi-Agma | 23.8 | 879.4717 | C42H67N6O14 | −0.9 | 18.9 | 352 | 360 | this study | |||||||
| 9 | Hpla-Leu-(Gluc,Hep)-OHChoi-Agma | 25.6 | 879.5078 | C43H71N6O13 | −0.5 | 10.7 | 358 | 360 | 381 | this study | ||||||
| 10 | Pla-dehydroLeu-(Gluc,Hexan)-OHChoi-Agma | 25.1 | 849.4596 | C41H65N6O13 | 0.9 | 9.1 | 352 | 358 | 360 | 381 | this study | |||||
| 11 | Hpla-Leu-(Gluc,Oct)-OHChoi-Agma | 26.1 | 893.4872 | C43H69N6O14 | −0.6 | 0.8 | 352 | 358 | 360 | 381 | 382 | this study | ||||
| anabaenopeptins | ||||||||||||||||
| 12 | Lys-CO[Lys-Ile-Hph-NMeAsn-Phe] | 24.0 | 850.4818 | C43H64N9O9 | 0.4 | 7.2 | 386 | this study | ||||||||
| 13 | Arg-CO[Lys-Ile-Hph-NMeAsn-Phe] | 24.4 | 878.4864 | C43H64N11O9 | 2.2 | 11.0 | 386 | this study | ||||||||
| 14 | Lys-CO[Lys-Ile-MeHph-NMeAsn-Phe] | 24.8 | 864.4972 | C44H66N9O9 | 0.7 | 1.8 | 386 | this study | ||||||||
| 15 | Arg-CO[Lys-Ile-MeHph-NMeAsn-Phe] | 25.2 | 892.5023 | C44H66N11O9 | 1.8 | 4.0 | 386 | this study | ||||||||
| 16 | Lys-CO[Lys-Ile-EtHph-NMeAsn-Phe] | 26.0 | 878.5122 | C45H68N9O9 | 1.4 | 2.0 | 386 | this study | ||||||||
| 17 | Arg-CO[Lys-Ile-EtHph-NMeAsn-Phe] | 26.3 | 906.5196 | C45H68N11O9 | −0.4 | 5.0 | 386 | this study | ||||||||
| 18 | Phe-CO[Lys-Val-Hty-MeAla-Hty] | 31.1 | 858.4405 | C45H60N7O10 | −1.1 | 2.5 | 352 | this study | ||||||||
| 19 | Phe-CO[Lys-Ile-Hty-MeAla-Hty] | 31.8 | 872.4544 | C46H62N7O10 | 1.0 | 5.9 | 352 | this study | ||||||||
| 20 | Val-CO[Lys-Ile-Trp-MeAla-Phe] | 34.1 | 803.4425 | C42H59N8O8 | 3.2 | 5.2 | 360 | 382 | this study | |||||||
| 21 | Val-CO[Lys-Ile-Trp-MeAla-Phe] | 34.6 | 803.4417 | C42H59N8O8 | 4.1 | 9.6 | 360 | 382 | this study | |||||||
| 22 | Leu-CO[Lys-Ile-MeHph-NMeAsn-Phe] | 34.8 | 849.4872 | C44H65N8O9 | −0.4 | 3.5 | 386 | this study | ||||||||
| 23 | Phe-CO[Lys-Val-Hph-MeAla-Hty] | 35.0 | 842.4416 | C45H60N7O9 | −3.1 | 1.9 | 352 | this study | ||||||||
| 24 | Phe-CO[Lys-Ile-Hph-MeAla-Hty] | 35.8 | 856.4561 | C46H62N7O9 | 5.0 | 1.6 | 352 | this study | ||||||||
| 25 | Phe-CO[Lys-Ile-MeHph-NMeAsn-Phe] | 35.4 | 883.4695 | C47H63N8O9 | 2.0 | 12.6 | 358 | this study | ||||||||
| 26 | Leu-CO[Lys-Ile-EtHph-NMeAsn-Phe] | 36.1 | 863.5028 | C45H67N8O9 | 0.3 | 4.0 | 386 | this study | ||||||||
| 27 | Phe-CO[Lys-Ile-EtHph-NMeAsn-Phe] | 36.7 | 897.4858 | C48H65N8O9 | 1.2 | 3.3 | 358 | this study | ||||||||
| cyanopeptolins | ||||||||||||||||
| 28 | Mdhp-Gln[Thr-Leu-Ahp-Leu-NMe-Cl-Tyr-Leu] | 27.7 | 1002.5031 | C48H73ClN9O12 | 3.0 | 12.0 | 371 | 386 | this study | |||||||
| 29 | AcPro-Gln[Thr-Leu-Ahp-Val-NMe-OMe-Tyr-Val] | 28.9 | 984.5381 | C48H74N9O13 | 2.0 | 13.5 | 386 | this study | ||||||||
| 30 | AcPro-Gln[Thr-Leu-Ahp-Val-NMe-Cl-Tyr-Val] | 29.1 | 1004.4831 | C47H71ClN9O13 | 2.3 | 5.9 | 371 | 386 | this study | |||||||
| 31 | AcPro-Gln[Thr-Leu-Ahp-Leu-NMe-Tyr-Leu] | 29.9 | 998.5531 | C49H76N9O13 | 2.7 | 9.9 | 386 | this study | ||||||||
| 32 | AcPro-Gln[Thr-Leu-Ahp-Val-NMe-Cl-Tyr-Leu] | 30.5 | 1018.5011 | C48H73ClN9O13 | 0.0 | 4.8 | 371 | 386 | this study | |||||||
| 33 | AcPro-Gln[Thr-Leu-Ahp-Val-NMe-OMe-Tyr-Val] | 31.0 | 984.5398 | C48H74N9O13 | 0.3 | 18.4 | 371 | 386 | this study | |||||||
| 34 | AcPro-Gln[Thr-Leu-Ahp-Leu-NMe-Cl-Tyr-Leu] | 31.6 | 1032.5170 | C49H75ClN9O13 | 0.3 | 4.9 | 371 | 386 | this study | |||||||
| 35 | AcPro-Gln[Thr-Leu-Ahp-Val-NMe-OMe-Tyr-Leu] | 32.1 | 998.5549 | C49H76N9O13 | 0.8 | 15.8 | 371 | 386 | this study | |||||||
| 36 | AcPro-Gln[Thr-Leu-Ahp-Leu-NMe-OMe-Tyr-Val] | 32.7 | 998.5564 | C49H76N9O13 | −0.7 | 2.6 | 371 | 386 | this study | |||||||
| 37 | AcPro-Gln[Thr-Leu-Ahp-Leu-NMe-OMe-Tyr-Leu] | 33.9 | 1012.5737 | C50H78N9O13 | 2.4 | 8.0 | 371 | 386 | this study | |||||||
| 38 | PrPro-Gln[Thr-Leu-Ahp-Leu-NMe-OMe-Tyr-Leu] | 35.2 | 1026.5858 | C51H80N9O13 | 1.1 | 26.7 | 371 | 386 | this study | |||||||
a Hpla: hydroxy-phenyl lactic acid; Pla: phenyl lactic acid; OH: hydroxy; Hex: hexose; Gluc: glucuronic acid; But: butanoic acid; Hexan: hexanoic acid; Hep: heptanoic acid; Oct: octanoic acid; Me: methyl; Et: ethyl; [] cyclo; Ac: acetyl; Pr: propanoyl; Mdhp: methyl-dehydroproline; NMe-Cl-Tyr: N-methyl-3-chloro-tyrosine; NMe-OMe-Tyr: N-methyl-O-methyl-tyrosine.
2. Result and Discussion
The peptide profiles of the 40 strains isolated from the Atlantic Forest (CENA350-389) were investigated using LC/DAD/ESI/QTOF/MS/MS. As an example, Figure 1 shows the chromatographic profiles of two strains: one Brasilonema sp. and one Desmonostoc sp. The retention times (RT), protonated molecules ([M + H]+), molecular formula provided for the experimental m/z, and error and millisigma (mSigma) values for the major peaks are summarized in Table 2. Mass accuracy was below 5 ppm for all the detected compounds. Peak identification was performed based on the data presented in Table 2, Table 3, Table 4, Table 5, Table 6, Table 7, Table 8, Table 9, Table 10, Table 11, Table 12, Table 13 and Table 14 and based on previously published data [23,24,26,31,32,33,34,35,36,37,38,39,40]. This approach allowed the elucidation of the planar structures of 38 peptides, including 10 new aeruginosins, 16 new anabaenopeptins, and 11 new cyanopeptolins. Among the surveyed strains the heterocytous strains Nostoc sp. CENA352, CENA358, and CENA369, Brasilonema sp. CENA360, CENA361, CENA381, and CENA382 and Desmonostoc sp. CENA386 and CENA371 were identified as producers of cyanopeptides. Cyanopeptides were not detected in the extracts of the remaining 31 cyanobacterial strains under our experimental conditions.
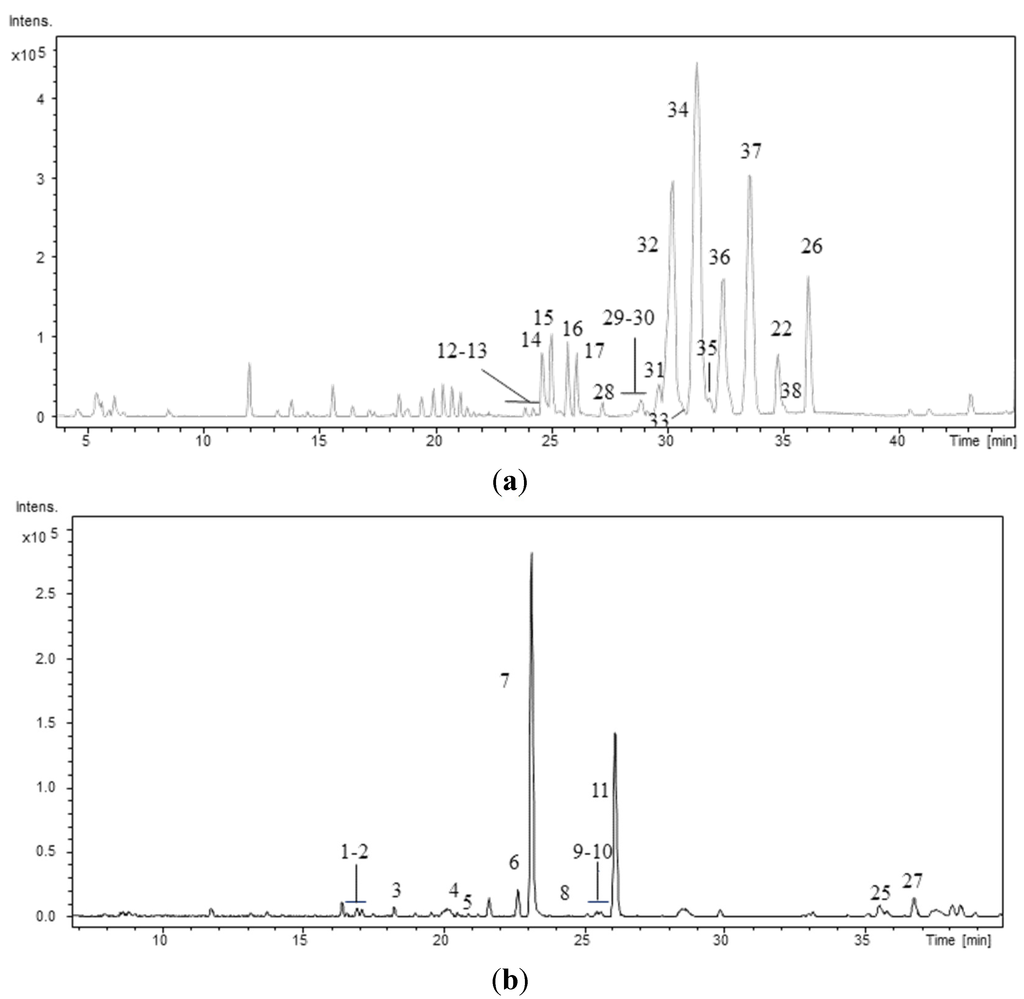
Figure 1.
Base Peak Chromatogram of the hydromethanolic extract of the strains (a) Desmonostoc sp. CENA386 and (b) Brasilonema sp. CENA358. Conditions as described in experimental section.
Figure 1.
Base Peak Chromatogram of the hydromethanolic extract of the strains (a) Desmonostoc sp. CENA386 and (b) Brasilonema sp. CENA358. Conditions as described in experimental section.
2.1. Aeruginosins
Seven strains belonging to the orders Nostocales (Nostoc sp. CENA352, CENA358, and CENA369; and Brasilonema sp. CENA360, CENA361, CENA381, and CENA382) were found to produce aeruginosins (1–11). Aeruginosins are linear tetrapeptides that contain the unusual amino acid 2-carboxy-6-hydroxyoctahydroindol (Choi) in the central position and typically contain an arginine derivative at the C-terminus [3]. The N-terminal position is occupied by a 3-(4-hydroxyphenyl) lactic or phenyl lactic acid, which can be acetylated, brominated, chlorinated, or sulfonated [41,42,43,44]. A small hydrophobic amino acid (leucine or isoleucine) is typically present in position 2. Additionally, hydroxylation, sulfation, and chlorination of the Choi moiety have also been observed [45]. These peptides are potent inhibitors of serine proteases, and this bioactivity is largely related to C-terminal modifications [46]. This family of compounds has been reported to be produced by cyanobacteria of the genera Microcystis [41,42,43,44,47], Planktothrix [24], Nodularia [48], and Nostoc [37].
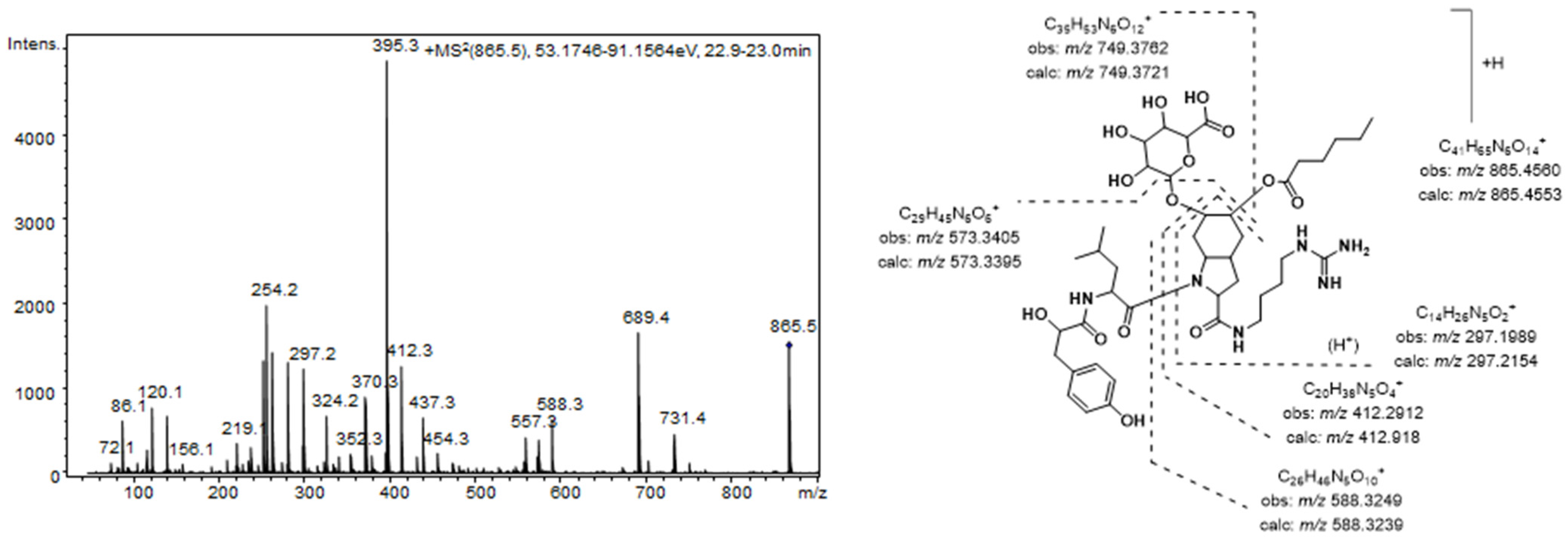
The aeruginosins found in these extracts were characterized by closely related structures, most of which were common to both Nostoc and Brasilonema producer species. The most prominent peak detected in the MS chromatogram of these extracts (7) was assigned to the aeruginosin 865 (m/z 865.4565 [M + H]+) [37]. This compound, which was recently isolated from a terrestrial cyanobacterium belonging to Nostoc sp., was structurally characterized as containing both a fatty acid and a carbohydrate attached to the Choi moiety [37]. Figure 2 shows the product ion spectra of this aeruginosin. A collision energy of 70 eV was necessary to obtain a spectrum with abundant and intense product ions. Consistent with the existence of an agmatine (Agma) residue in the molecule, the C-terminal ions (m/z 588.3249, 412.2912, and 297.1989) and the corresponding satellite ions, which were produced via ammonia or water loss (m/z 571.2985, 395.2657, 279.1820, and 261.1717) dominated the spectrum. Additionally, the ions generated by the cleavage of the glycosidic acid and/or the ester bond established the sugar and the lipid acids as glucuronic acid and hexanoic acid, respectively. The presence of ions at m/z 18 u higher (m/z 156.1035 and 138.0866) than the diagnostic ions that are typically generated from the Choi residue (m/z 140 and 122) indicated the dihydroxylation of the indole ring in this amino acid and were key fragments for the detection of other aeruginosin congeners.
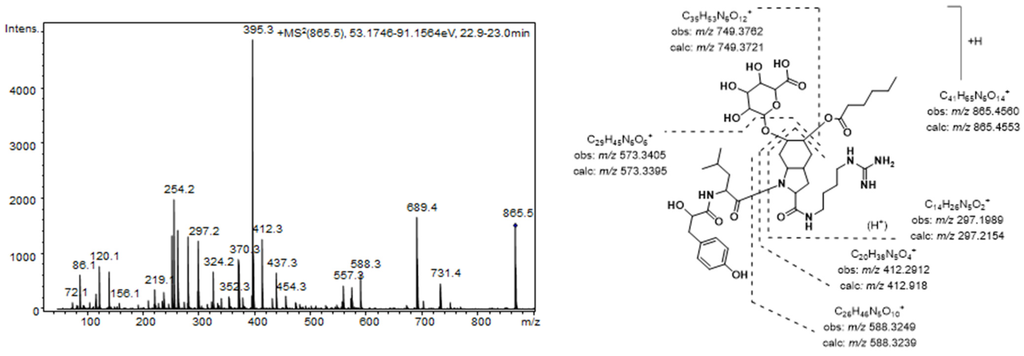
Figure 2.
Product ion spectrum for [M + H]+ of compound 7 and its predicted fragmentation pattern. Conditions as mentioned in the experimental section.
Figure 2.
Product ion spectrum for [M + H]+ of compound 7 and its predicted fragmentation pattern. Conditions as mentioned in the experimental section.
A total of 10 additional aeruginosins that shared conserved substructures (1–6, 8–11) and yielded [M + H]+ at m/z 753.4042 (1), 767.3828 (2), 737.4044 (3), 823.4470 (4), 837.4253 (5), 851.4780 (6), 879.5078 (8), 879.5078 (9), 849.4596 (10), and 893.4872 (11) were identified as new congeners. The product ion spectra of these aeruginosins were similar to that of compound 7, allowing us to elucidate their structures based on a comparison of spectra (Table 3, Table 4 and Table 5).
Table 3.
Product ion spectra data for compounds 4–7, 9, and 11.
| Product Ion Assignment a | 4 (m/z) | 5 (m/z) | 6 (m/z) | 7 (m/z) | 9 (m/z) | 11 (m/z) |
|---|---|---|---|---|---|---|
| Leu immonium | 86.0967 | 86.0947 | 86.0907 | 86.0945 | 86.0906 | 86.0932 |
| OH-Choi immonium ion − H2O | 138.0791 | 138.0866 | 138.0910 | 138.0912 | 138.0938 | 138.0903 |
| OH-Choi immonium ion | 156.1035 | 156.1035 | 156.1033 | 156.1026 | - | 156.1031 |
| OH-Choi-Agma − NH3 − H2O + H | 261.1693 | 261.1707 | 261.1799 | 261.1717 | 261.1791 | 261.1709 |
| OH-Choi-Agma − H2O + H | 279.1802 | 279.1801 | 279.1799 | 279.1820 | 279.1796 | 279.1828 |
| OH-Choi-Agma + H | 297.1925 | 297.1885 | 297.1882 | 297.1989 | 297.1880 | 297.1878 |
| (R1)-O-Choi-Agma + H − NH3 | 367.2348 | 367.2342 | 395.2672 | 395.2657 | 409.2803 | 423.2969 |
| (R1)-O-Choi-Agma + H | 384.2618 | 384.2617 | 412.2803 | 412.2912 | 426.3113 | 440.3226 |
| (R1, R2)-OH-Choi-Agma+ H − NH3 | - b | 543.2599 | - | 571.2985 | - | 599.3216 |
| (R1, R2)-OH-Choi-Agma + H | 546.3176 | 560.2818 | 574.3454 | 588.3249 | - | 616.3618 |
| Hpla-Leu-OHChoi-Agma + H − H2O | 573.3423 | 573.3422 | 573.3417 | 573.3405 | - | 573.3409 |
| Hpla-Leu-(R1)OH-Choi-Agma | 661.3949 | 661.3909 | 689.4266 | 689.4250 | 703.4423 | 717.4543 |
| Hpla-Leu-(R2)OH-Choi-Agma | 735.3952 | 749.3722 | 735.3945 | 749.3762 | - | - |
| Hpla-Leu-(R1, R2)-OH-Choi-Agma | 823.4470 | 837.4253 | 851.4780 | 865.4560 | 879.5078 | 893.4872 |
a R1 aliphatic lipid acid linked to hydroxyChoi in position 5: butanoic acid for compounds 4 and 5; hexanoic acid for 6 and 7; heptanoic acid for 9 and octanoic acid for 11. R2 sugar linked to hydroxyChoi in position 6: hexose for compounds 4 and 6; and glucuronoic acid for 5 and 7; b - not detected.
Table 4.
Product ion spectra data for compounds 1–3.
| Product Ion Assignment a | 1 (m/z) | 3 (m/z) | 2 * (m/z) |
|---|---|---|---|
| Leu immonium | 86.0908 | 86.0908 | 86.0967 |
| OH-Choi immonium ion − H2O | 138.0867 | 138.0941 | 122.1066 * |
| OH-Choi immonium ion | 156.0957 | 156.1036 | 140.1034 * |
| OH-Choi-Agma − NH3 − H2O | 261.1592 | - b | - |
| OH-Choi-Agma − NH3 | 297.1920 | 297.1928 | 263.1868 * |
| OH-Choi-Agma | 314.2143 | 314.2255 | 281.1914 * |
| R2-OH-Choi-Agma − NH3 | - | 473.2107 | 443.2511 * |
| R2-OH-Choi-Agma | 476.2717 | 490.2454 | 460.2740 * |
| Hpla-Leu-OHChoi-Agma | 591.3360 | 591.3204 | 575.3841 * |
| Hpla-Leu-(R2)-OH-Choi-Agma | 753.4042 | 767.3828 | 737.4044 * |
a R2 sugar linked to hydroxyChoi in position 6: hexose for compounds 1 and 2; and glucuronoic acid for 3; b - not detected; * structure lacking the 5-hydroxylation in Choi moiety.
Table 5.
Product ion spectra data for compounds 8 and 10.
| Product Ion Assignment a | 8 (m/z) | 10 (m/z) |
|---|---|---|
| X immonium | 100.1148 | 86.0906 |
| OH-Choi immonium ion − H2O | 138.1248 | 138.0863 |
| OH-Choi immonium ion | - b | 156.0952 |
| OH-Choi-Agma − NH3 − H2O | 261.1689 | 261.1687 |
| OH-Choi-Agma − H2O | 279.1797 | 279.1796 |
| OH-Choi-Agma − NH3 | 297.1771 | - |
| OH-Choi-Agma + H | - | - |
| (R1)-OH-Choi-Agma − NH3 | 395.2544 | 395.2659 |
| (R1)-OH-Choi-Agma + H | 412.2930 | 412.2928 |
| (R1, R2)-OH-Choi-Agma − NH3 | 571.3340 | 571.2881 |
| (R1, R2)-OH-Choi-Agma + H | 588.3205 | 588.3238 |
| Z-X-(R1)OH-Choi-Agma + H | - | 673.4238 |
| Z-X-(R1)-OH-Choi-Agma + H | 703.4398 | - |
| Z-X-(R1, R2)-OH-Choi-Agma + H | 879.4717 | 849.4596 |
a R1: butanoic acid linked to hydroxyChoi in position 5; R2: glucuronoic acid linked to hydroxyChoi in position 6. X aminoacid in second position: methyl-leucine for compound 8 and leucine for 10; Z phenyl alkanoic acid in position 1: hydroxylphenyl lactic acid for compound 8 and phenyllactic acid for 10; b - not detected.
In this sense, a structure similar to that of aeruginosin 865 was proposed for compounds 5, 9, and 11 except for the fatty acid esterifying position 5 of the Choi moiety. In this position, butanoic, heptanoic, and octanoic acid were proposed for each compound, respectively. These changes were clearly evidenced by the sequence of ions containing the aforementioned fatty acids (m/z 384.2617, 543.2599, 560.2818, and 661.3909 for compound 5; m/z 409.2803 and 703.4423 for compound 9; and m/z 423.2969, 599.3216, 616.3618, and 717.4543 for compound 11). On the other hand, structural differences between the pairs of compounds 4, 5 and 6, 7 were attributed to Choi-glycosylation.A similar product ion spectra, which differed only in the ions generated by the cleavage of the glycosidic bond (m/z 546.3176, 574.3454, and 735.3952/735.3945) suggested that the glucuronic acid in compounds 5 and 7 was replaced by a hexose in compounds 4 and 6.
Compounds 1 and 3, which also showed structures similar to compounds 4 and 6 and 5, 7, 9, and 11, respectively, were distinguished by the lack of fatty acids in their structures (Table 4). These peptides could be biosynthetic intermediates of the respective fatty acid-containing aeruginosins. Along with these compounds, an oxygen-deficient variant of compound 1 (2) was also detected; this structural difference was likely due to the absence of the Choi 5-hydroxylation.
Finally, two other structural variants of compound 7 were also detected (8 and 10). For compound 8, methylation of the amino acid in the second position was suggested based on the mostly conserved product ion spectrum and the presence of a fragment ion at m/z 100.1148 (NMeLeu immonium). Similarly, for compound 10, a phenyl lactic acid was proposed for the N-terminus instead of a hydroxyl-phenyl lactic acid (Table 5).
From a biomedical point of view, the pharmacological potential of these new aeruginosin variants must be evaluated. As mentioned above, aeruginosins typically exhibit antithrombotic activity, making these compounds interesting candidates for the development of anticoagulant drugs [46]. Additionally, all of these compounds are structurally similar to aeruginosin-865, which has exhibited remarkable anti-inflammatory activity [37]. Evaluations of the bioactivity of these compounds could provide insights into the structure-activity relationship of this class of aeruginosins.
2.2. Anabaenopeptins
Fifteen compounds produced by Nostoc sp. CENA352, Brasilonema sp. CENA360, and Desmonostoc sp. CENA386 were characterized as new anabaenopeptin analogs (12–27). Protonated molecules in the mass range of anabaenopeptins and an important loss of the amino acid in the side-chain position were the diagnostic criteria used to classify these peptides [23]. These new anabaenopeptins were characterized based on empirical formulae, product ion spectra, and previously described sequences [23,24,26,34,36].
Anabaenopeptins are hexapeptides that contain a ring of five amino acids. Position 2 is always occupied by d-Lys, which both closes the ring with the amino acid at position 6 and establishes a ureido link with the amino acid in position 1, giving rise to a side chain. Positions 4 and 5 are typically occupied by aromatic and methylated amino acids, respectively. Position 3 has been reported to be occupied primarily by valine or isoleucine/leucine and less frequently by methionine [3]. Various biological activities have been described for these structures, including inhibition of protein phosphatase [49], carboxypeptidases A [50,51] and U [52], and other protease inhibitory activity [53]. To date, 30 anabaenopeptins have been isolated from many different cyanobacteria genera [8,54] (Anabaena [55,56], Aphanizomenon [51], Lyngbya [57], Microcystis [58], Oscillatoria [53,59], Planktothrix [60], and Schizothrix [61]) and also from marine sponges [62,63].
The detected anabaenopeptins could be grouped according to their structural features. Among these compounds, 10 anabaenopeptins that shared similar structural characteristics were detected in Desmonostoc sp. CENA360 and Brasilonema sp. CENA386 (12–17, 22, 25–27). The representative fragmentation pathways and spectra of these compounds are described in Figure 3 and Figure 4, and the assignments of the principal ions are shown in Table 6, Table 7, Table 8 and Table 9. Since the nature of the exocyclic amino acid varies among them, two different fragmentation patterns were observed for these compounds depending on the nature of the amino acid side chain. Extensive fragmentation was observed due to the absence of polar and basic residues in this chain. These anabaenopeptins commonly incorporate the amino acid N-methyl asparagine (N-MeAsn) in position 5, and on several occasions, either methylation or ethylation was postulated for the homovariant amino acid in position 4.
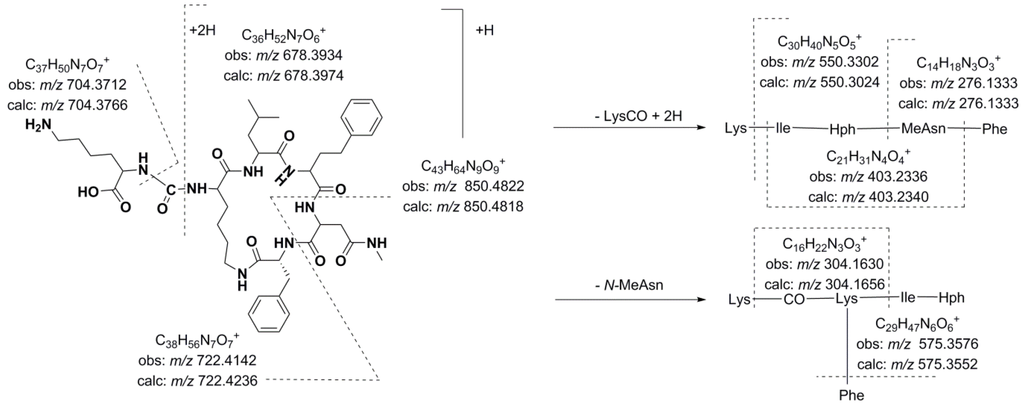
Figure 3.
Predicted fragmentation pattern for compound 12.
Figure 3.
Predicted fragmentation pattern for compound 12.
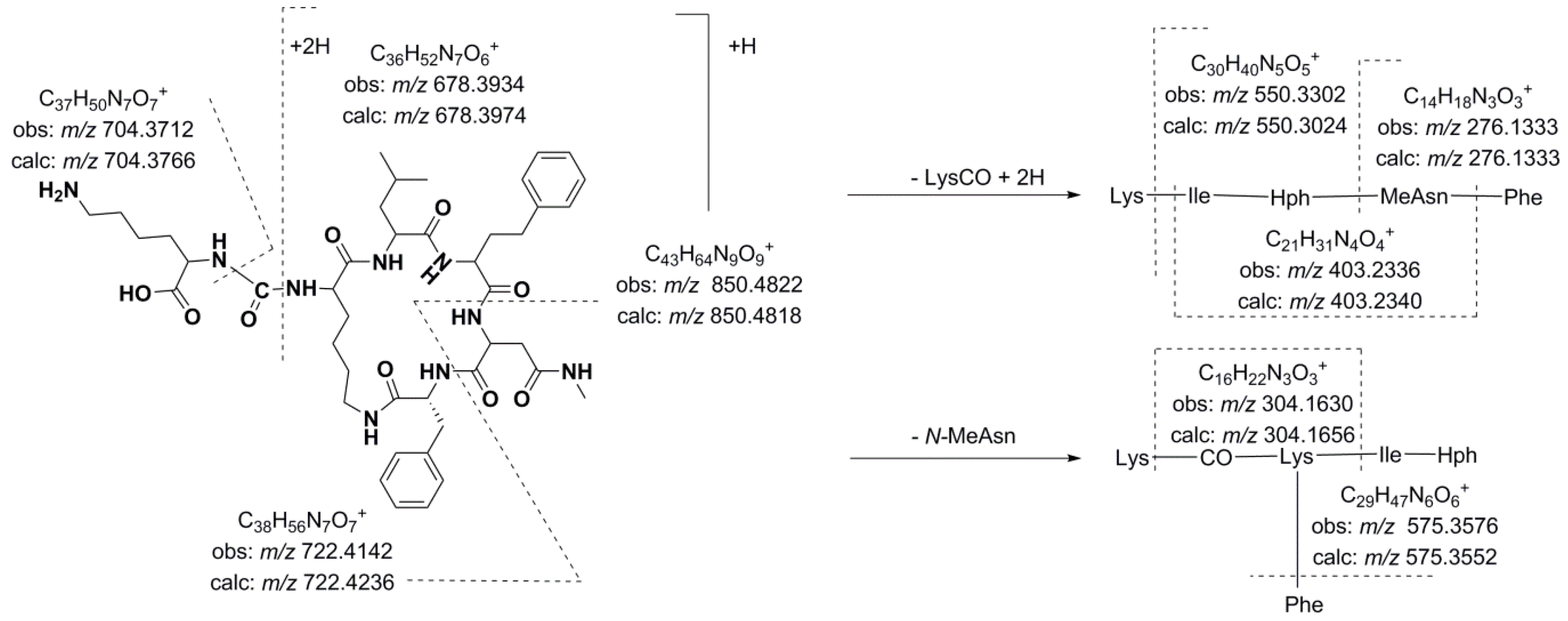
Figure 4.
Product ion spectra for [M + H]+ of compound 14 at (a) CE: 35 eV; (b) CE: 50 eV; and (c) CE: 70 eV; and (d) compound 21 at CE 35 eV.
Figure 4.
Product ion spectra for [M + H]+ of compound 14 at (a) CE: 35 eV; (b) CE: 50 eV; and (c) CE: 70 eV; and (d) compound 21 at CE 35 eV.
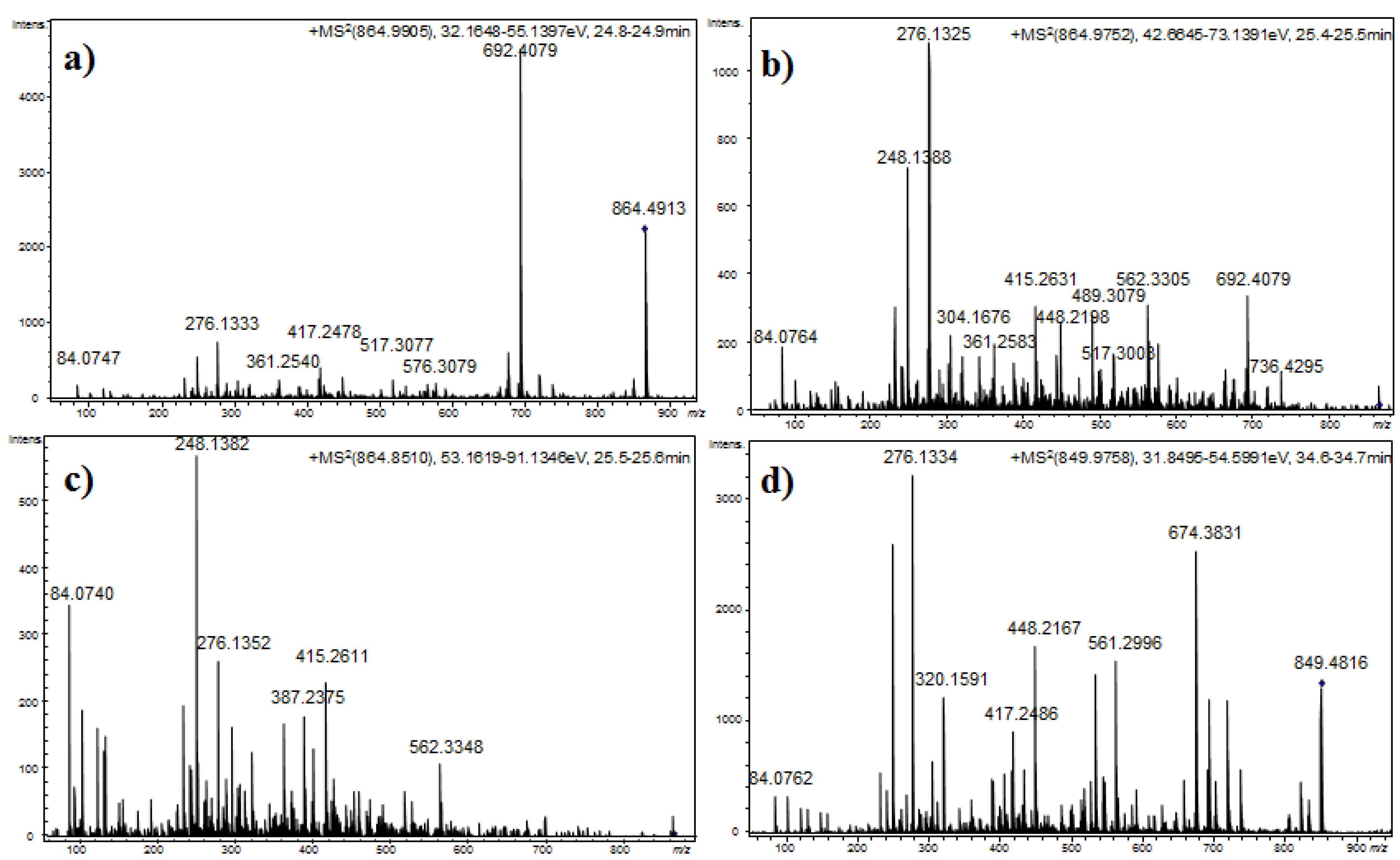
Table 6.
Product ion spectra data for compounds 12, 14, and 16.
| Product Ion Assignment a | 12 (m/z) | 14 (m/z) | 16 (m/z) |
|---|---|---|---|
| Lys fragment | 84.0768 | 84.0748 | 84.0760 |
| MeAsn immonium ion | 101.0702 | 101.0670 | 101.0649 |
| Phe immonium ion | 120.0738 | 120.0780 | 120.0793 |
| Z immonium ion | - b | 148.1125 | 162.1259 |
| MeAsn-Phe − CO + H | 248.1406 | 248.1400 | 248.1355 |
| MeAsn-Phe + H | 276.1333 | 276.1350 | 276.1320 |
| CO-Lys-Phe + H | 304.1630 | 304.1642 | - |
| Z-MeAsn + H | - | - | 318.1750 |
| Phe-Lys-Ile + H | - | 387.2375 | 387.2269 |
| Lys-Ile-Z + H | - | - | 429.2842 |
| Z-Ile-Lys + H | 401.2661 | 415.2674 | 429.2808 |
| Ile-Z-MeAsn + H | 403.2336 | 417.2517 | 431.2620 |
| Phe-Lys-Ile-Z + H | - | 562.3374 | 576.3330 |
| Ile-Z-MeAsn-Phe + H | 550.3302 | 564.3783 | 578.3556 |
| Lys-CO-Lys-(Ile-Z) + H | 575.3576 | 589.3531 | 603.3697 |
| Lys-Ile-Z-MeAsn-Phe − NH3 + 2H | 661.3682 | 675.3851 | 689.4008 |
| Lys-Ile-Z-MeAsn-Phe + 2H | 678.3934 | 692.4075 | 706.4232 |
| CO-[Lys-Ile-Z-MeAsn-Phe] + H | 704.3712 | 718.3935 | 732.4006 |
| Lys-CO-Lys-(Phe)-(Ile-Z) + H | 722.4142 | 736.4312 | 750.4436 |
| Lys-CO-[Lys-Ile-Z-MeAsn-Phe] + H | 850.4818 | 864.4972 | 878.5122 |
a Z amino acid in the fourth position: Hph for compound 12, MeHph for 14, EtHph for 16; b - not detected.
Table 7.
Product ion spectra data for compounds 13, 15, and 17.
| Product Ion Assignment a | 13 (m/z) | 15 (m/z) | 17 (m/z) |
|---|---|---|---|
| Lys related fragment | 70.0544 | 70.0619 | 70.0552 |
| Lys fragment | 84.0706 | 84.0731 | 84.0350 |
| MeAsn immonium ion | 101.0617 | 101.068 | - |
| Phe immonium ion | 120.0679 | 120.0748 | - |
| Arg + H | 175.1205 | 175.1184 | 175.1171 |
| CO-Arg | 201.0964 | 201.0966 | 201.0970 |
| MeAsn-Phe − CO + H | -b | 248.1435 | 248.1286 |
| MeAsn-Phe + H | - | 276.1298 | 276.1337 |
| Phe-Lys-Ile + H | - | 387.2358 | 387.2356 |
| Ile-Z-MeAsn + H | 403.7866 | 417.2410 | - |
| Ile-Z-MeAsn-Phe + 2H | - | 564.3384 | - |
| Lys-Ile-Z-MeAsn-Phe − NH3 + 2H | - | - | 689.3691 |
| Lys-Ile-Z-MeAsn-Phe + 2H | 678.4045 | 692.4084 | 706.4222 |
| Arg-CO-Lys-(Phe)-(Ile-Z) + H | - | - | 778.4472 |
| Arg-CO-[Lys-Ile-Z-MeAsn-Phe] + H | 878.4864 | 892.5023 | 906.5196 |
a Z amino acid in the fourth position: Hph for compound 13, MeHph for 15, EtHph for 17; b - not detected.
Table 8.
Product ion spectra data for compounds 22 and 26.
| Product Ion Assignment a | 22 (m/z) | 26 (m/z) |
|---|---|---|
| Lys fragment | 84.0753 | 84.0768 |
| Leu immonium ion | 86.0924 | 86.0924 |
| MeAsn immonium ion | 101.0695 | 101.0677 |
| Phe immonium ion | 120.0810 | 120.0724 |
| Z immonium ion | 148.1124 | 162.1273 |
| MeAsn-Phe − CO + H | 248.1381 | 248.1369 |
| MeAsn-Phe + H | 276.1335 | 276.1326 |
| CO-Lys-Phe + H | 304.1667 | - b |
| Phe-Lys-Ile | 387.2406 | 387.2370 |
| Ile-Z-MeAsn + H | 417.2486 | 431.2661 |
| Z-Ile-Lys + H | 415.2558 | - |
| Ile-Z-MeAsn-Phe + 2H | 564.3281 | - |
| Leu-CO-Lys-Phe | 433.2435 | - |
| CO-Lys-Phe-NMeAsn | 431.1872 | - |
| Leu-CO-Lys-NMeAsn-Phe − CO + H | 533.2997 | 533.2996 |
| Leu-CO-Lys-NMeAsn-Phe | 561.2995 | 561.2967 |
| Leu-CO-Lys-(Ile)-(NMeAsn-Phe) − NH3+ H | - | 657.3648 |
| Leu-CO-Lys-(Ile)-(NMeAsn-Phe) | 674.3823 | 674.3810 |
| Leu-CO-Lys-Z-NMeAsn-Phe | 736.3966 | 750.4096 |
| [Lys-Ile-Z-MeAsn-Phe] + 2H | 692.4077 | 706.4201 |
| Leu-CO[Lys-Ile-Z-NMeAsn-Phe] | 849.4872 | 863.5028 |
a Z amino acid in the fourth position: MeHph for compounds 22 and EtHph for 26; b - not detected.
Table 9.
Product ion spectra data for compounds 25 and 27.
| Product Ion Assignment a | 25 (m/z) | 27 (m/z) |
|---|---|---|
| Lys fragment | 84.0714 | 84.0714 |
| MeAsn immonium ion | 101.0633 | 101.057 |
| Phe immonium ion | 120.0701 | 120.077 |
| Z immonium ion | 148.1074 | 162.1336 |
| MeAsn-Phe − CO + H | 248.1425 | 248.1362 |
| MeAsn-Phe + H | 276.1338 | 276.1333 |
| CO-Lys-Phe + H | 304.1570 | - b |
| Phe-CO-Lys + H | 320.1601 | 320.1534 |
| Phe-Lys-Ile + H | - | 387.2388 |
| Ile-Z-MeAsn + H | 417.2582 | - |
| Z-Ile-Lys + H | 415.2709 | - |
| Phe-CO-Lys-Phe + H | 467.2262 | - |
| Phe-CO-Lys-Phe-MeAsn + H | 595.2844 | 595.2842 |
| Phe-CO-Lys-Phe-MeAsn-Z + H | 770.3866 | - |
| Phe-CO-Lys-(Ile-Z)-(MeAsn) + H | 736.3827 | - |
| Phe-CO-Lys-Ile-Z-MeAsn + H | 736.3827 | - |
| Lys-Ile-Z-MeAsn-Phe | - | 706.4299 |
| Lys-Ile-Z-MeAsn-Phe + 2H | 708.3705 | 708.3720 |
| CO-[Lys-Ile-Z-MeAsn-Phe] + H | 718.3895 | 732.4082 |
| Phe-CO-(Lys-Ile-Z)-(Phe) + H | 755.3925 | 767.3730 |
| Phe-CO-[Lys-Ile-Z-MeAsn-Phe] + H | 883.4695 | 897.4858 |
a Z amino acid in the fourth position: MeHph for 25 and EtHph for 27; b - not detected.
Thus, compound 12 (m/z 850.4818 [M + H]+) was characterized as Lys-CO[Lys-Ile-Hph-MeAsn-Phe]. As expected, the presence of Lys in the side chain generated a simple fragmentation pattern. Consequently, just two low intensity series of fragment ions that were assigned to two preferential primary cleavages were observed in the spectrum (Figure 4). However, enough information was generated to allow a tentative interpretation of the spectrum. The preferential fragmentation observed was attributed to the cleavage of the ureido linkage and the opening of the ring and indicated the presence of lysine in the side chain (m/z 678.3934). Further amino acid residue losses from this lineal ion (i.e., acylium or similar) yielded a series of ions at m/z 550.3302, 403.2336, 304.1630, and 276.1333 (loss of Lys, Phe, Ile/Leu, and Hph, respectively). Another series of less abundant ions that were generated by the loss of N-methyl asparagine and phenylalanine was also observed at m/z 722.4142 (loss of MeAsn) and 575.3576 (loss of Phe). The dipeptide fragments at m/z 304.1630 and 276.1333 determined the partial sequence Hph-MeAsn-Phe and established the isoleucine/leucine at position 3 (m/z 403.2336) and the lysine that closed the ring (m/z 678.3934). Ions that are typically observed in the low m/z region were also present (Lys at m/z 84.0768, MeAsn at m/z 101.0702, Phe at m/z 120.0738).
Compounds 14 and 16, which exhibited molecular mass increases of 14 and 28 Da, respectively, when compared to the compound at m/z 850.4818 [M + H]+, exhibited similar product ion spectra. Differences that were observed in the fragments containing the amino acid at position 4 suggested that this residue was either methylated (m/z 417.2517, 564.3783, 692.4075, and 736.4312) or ethylated (m/z 431.2620, 576.3330, 689.4008, and 750.4436). The existence of ions at m/z 148.1125 and 162.1213 that were attributed to the N-methylhomophenylalanine (N-MeHph) and N-ethylhomophenylalanine (N-EtHph) immonium ions, respectively, also supported these assignments.
Using similar reasoning, the remaining anabaenopeptins were subsequently characterized. Losses of 157 u that were observed for compounds 22 and 26, losses of 191 u that were observed for compounds 25 and 27, and losses of 200 u that were observed for compounds 13, 15, and 17 were used to identify the amino acid side chain as leucine, phenylalanine, or arginine, respectively. A fragmentation pattern similar to that described in the preceding paragraphs characterized the cyclic structures. As mentioned above, compounds with [M + H]+ at m/z 878.5122, 892.5023, and 906.5196 exhibited low efficiency fragmentations, which are characteristic of oligopeptides containing strongly basic residues. When the collision energy used for fragmentation was increased to improve efficiency, the abundance of the fragment ions was compromised (Figure 4).
Four additional anabaenopeptins (18, 19, 23, 24) were found in Nostoc sp. CENA352. Accurate mass measurement, isotopic profiles, and MS/MS spectra were conclusive to distinguish these compounds from previously described anabaenopeptins and characterize these compounds as new variants [36,53]. Structurally, all of these anabaenopeptins incorporated phenylalanine in the side chain position, as evidenced by the loss of a 191-u fragment from the protonated molecules (Table 10). Positions 5 and 6 of these four peptides were also conserved and were occupied by N-methyl-alanine and homotyrosine respectively. For the homovariant amino acids in position 4, either homophenylalanine (18, 19) or homotyrosine (23, 24) was proposed; for position 3, the commonly reported valine (18 and 23) or leucine/isoleucine (19 and 24) was proposed. For example, diagnostic low mass fragment ions that were observed in the product ion spectrum of the compound at m/z 842.4416 [M + H]+ (23) provided information about the amino acid residues, indicating the presence of lysine (m/z 84.0744), N-methyl-alanine (m/z 114.0515), phenylalanine (m/z 120.0736), homophenylalanine (m/z 134.0921), and homotyrosine (150.0887), while higher product ions suggested their sequence. Fragment ions at m/z 247.1440 and 263.1399 indicated the attachment of N-methyl-alanine to both homotyrosine and homophenylalanine, and subsequently established the sequence Val-Hph-MeAla for m/z 346.2124 and Lys-Val-Hph for m/z 387.2384. Taking into account the reported anabaenopeptins structures, the Phe-CO-[Lys-Val-Hph-MeAla-Hty] sequence was reasonably postulated. The intense fragment ion generated by the loss of a homophenylalanine residue at m/z 681.3591 and the less intense series of b-ions (m/z 596.3041, 497.2381, 320.1602, and 419.2283) were thus comprehensively attributed (Table 10).
Table 10.
Product ion spectra data for compounds 18, 19, 23, and 24.
| Product Ion Assignment a | 18 (m/z) | 19 (m/z) | 23 (m/z) | 24 (m/z) |
|---|---|---|---|---|
| Lys fragment | 84.0734 | 84.0757 | 84.0744 | 84.0749 |
| MeAla + H − CO | 114.0499 | 114.0557 | 114.0515 | 114.0514 |
| Phe immonium ion | - b | 120.0736 | 120.0736 | 120.0742 |
| Hph immonium | - | - | 134.0921 | 134.0929 |
| Htyr immonium | 150.0907 | 150.0907 | 150.0887 | 150.0873 |
| Hph-MeAla +H | - | - | 247.1440 | 247.1485 |
| HTyr-MeAla + H | 263.1381 | 263.1394 | 263.1399 | 263.1391 |
| Y-Z-MeAla + H | 362.2138 | 376.2236 | 346.2124 | 360.2270 |
| Lys-Y-Z + H | 403.2366 | 417.2447 | 387.2384 | 401.2544 |
| MeAla-Hty-Lys-Y + H | 490.2942 | - | 490.3023 | 514.3045 |
| Phe-CO-Lys-Y + H | - | - | 419.2283 | - |
| Phe-CO-Lys + 2H | - | - | 320.1602 | 320.1605 |
| Phe-CO-Lys-Hty + 2H | 497.2336 | 497.2298 | 497.2381 | 497.2387 |
| Phe-CO-Lys-Y-Z + H | - | 610.3218 | 596.3041 | 610.3266 |
| Phe-CO-Lys-Hty-MeAla + 2H | 582.2993 | 582.2962 | 582.2918 | 582.2915 |
| Lys-Y-Z-MeAla-Hty | 665.3655 | 679.3793 | 649.3720 | 663.3819 |
| Lys-Y-Z-MeAla-Hty + 2H | 667.3786 | 681.3953 | 651.3873 | 665.4013 |
| Phe-CO-Lys-(Y)-(Hty-MeAla) + 2H | 681.3604 | 695.3756 | 681.3597 | 695.3756 |
| CO-[Lys-Y-Z-MeAla-Hty] + H | 693.3629 | - | - | - |
| Phe-CO-[Lys-Y-Z-MeAla-Hty] + H − CO | 830.4384 | 844.4575 | 814.4502 | 828.4634 |
| Phe-CO-[Lys-Y-Z-MeAla-Hty] + H − H2O | 840.4385 | 854.4465 | 824.4377 | 838.4398 |
| Phe-CO-[Lys-Y-Z-MeAla-Hty] + H | 858.4405 | 872.4544 | 842.4416 | 856.4561 |
a Y amino acid in the third position: Val for compounds 18 and 23 and Ile for 19 and 24; Z amino acid in the fourth position: Hty for 18 and 19 and Hph for 23 and 24; b - not detected.
In addition, a pair of unusual tryptophan-containing anabaenopeptins (20–21) was also detected at m/z 803.4417 [M + H]+ and 803.4425 [M + H]+, exclusively in the genus Brasilonema sp. (CENA360 and CENA382). As this pair of compounds exhibited similar protonated molecules and product ion spectra but differed in retention times, these compounds were classified as diastereoisomers. The structures of these compounds were postulated in accordance with the fragment ions listed in Table 11. According to our literature search, prior to our work, the occurrence of tryptophan-containing anabaenopeptins in cyanobacteria was limited to the genus Tychonema sp. [64]. The structurally related compounds isolated from this genus, the brunsvicamides A–C, inhibit tyrosine phosphatase B of Mycobacterium tuberculosis (MptpB) [64] and are highly selectivity inhibitors for human leukocyte elastase (HLE) [65]. However, those peptides and the compounds reported here differ in their amino acid sequences. Unlike the brunsvicamides, the postulated anabaenopeptins contain tryptophan in position 4 and N-methyl-alanine in the methylated amino acid position. The structures of the compounds described in this study are more closely related to a synthetic brunsvicamide analog described by Walther et al. [66], which was found to be an inhibitor of carboxypeptidase A. Thus, further biological tests of these compounds are warranted.
Table 11.
Product ion spectra data for compounds 20 and 21.
| Product Ion Assignment | 20 (m/z) | 21 (m/z) |
|---|---|---|
| Lys fragment | 84.0730 | 84.0721 |
| MeAla + H − CO | 114.0519 | 114.0504 |
| Phe immonium ion | 120.0833 | 120.0747 |
| Trp fragment | 130.0645 | 130.0607 |
| Lys-Ile + H | 240.1738 | 240.1752 |
| Trp-MeAla + H | 272.1384 | 272.1429 |
| Ile-Trp-MeAla + H | 385.2229 | 385.2238 |
| CO-Lys-Phe + H | 320.1599 | 320.1602 |
| Val-CO-Lys-Ile + H | 385.2229 | 385.2211 |
| CO-Lys-Phe-MeAla + H | 405.2128 | 405.2084 |
| Val-CO-Lys-Ile + H | 419.2214 | 419.2240 |
| Val-CO-Lys-Phe-MeAla + H | 504.2829 | 504.2806 |
| Val-CO-Lys-Phe-Ile + H | 532.3069 | 532.3204 |
| Val-CO-Lys-(Ile)-(Phe-MeAla) + H | 617.3706 | 617.3686 |
| Lys-Ile-Trp-MeAla-Phe + H | 658.3680 | 658.3493 |
| Lys-Ile-Trp-MeAla-Phe + 2H | 660.3879 | 660.3833 |
| CO-Lys-Ile-Trp-MeAla-Phe + H | 686.3690 | 686.3681 |
| Val-CO-Lys-(Ile-Trp)-(Phe) + H–CO2 | 674.3749 | 674.3970 |
| Val-CO-[Lys-Ile-Trp-MeAla-Phe] + H − CO2 | 759.4538 | 759.4521 |
| Val-CO-[Lys-Ile-Trp-MeAla-Phe] + H − CO | 775.4563 | 775.4595 |
| Val-CO-[Lys-Ile-Trp-MeAla-Phe] + H − H2O | 785.4408 | 785.4278 |
| Val-CO-[Lys-Ile-Trp-MeAla-Phe] + H | 803.4417 | 803.4425 |
2.3. Cyanopeptolins
Eleven new cyanopeptolins with protonated molecules at m/z 984.5381 (29), 984.5398 (33), 998.5531 (31), 998.5549 (35), 998.5564 (36), 1002.5031 (28), 1004.4831 (30), 1012.5737 (37), 1018.5011 (32), 1026.5858 (38), and 1032.5170 (34) were present in the extracts from Desmonostoc strains CENA371 and CENA386. Although no single diagnostic fragment ion can be used to identify this family of peptides, series of fragments related to the conserved position 4 (3-amino-6-hydroxy-2-piperidone amino acid (Ahp) could be used to identify these compounds [23]. Under our experimental conditions, doubly charged ions ([M − H2O + 2H]2+) with an abundance comparable to that of protonated molecules were also observed in the mass spectra of all detected cyanopeptolins. MS and MS/MS analyses suggested closely related structures for all of these cyanopeptides (Table 12, Table 13 and Table 14). As the most common structural feature, these cyanopeptolins contained N-acetyl-proline-glutamine as side chain, with the fifth position occupied by either a dimethylated tyrosine or chlorinated-methylated tyrosine. Valine and leucine alternated in positions 4 and 6.
Table 12.
Product ion spectra data for compounds 30, 32, 34, and 37.
| Product Ion Assignment a | 30 (m/z) | 32 (m/z) | 34 (m/z) | 37 (m/z) |
|---|---|---|---|---|
| Pro immonium | 70.0542 | 70.0595 | 70.0571 | 70.0541 |
| Val immonium | -b | 72.0692 | - | - |
| Leu immonium | - | - | 86.0900 | 86.0935 |
| NAcPro immonium | 112.0695 | 112.0732 | 112.0679 | 112.0679 |
| NAcPro + H | 140.0623 | 140.0773 | 140.0678 | 140.0706 |
| NMe-R1-Tyr immonium | 184.0572 | 184.0486 | 184.0523 | 164.1052 |
| Leu-Ahp − H2O − CO + H | - | 181.1263 | 181.1290 | 181.1300 |
| Leu-Ahp − H2O + H | 209.1239 | - | 209.1257 | 209.1286 |
| Ac-Pro-Gln + H | 268.1207 | 268.1276 | 268.1270 | 268.1271 |
| Ac-Pro-Gln-Thr − H2O + H | 351.1656 | 351.1645 | 351.1638 | 351.1633 |
| NMe-R1-Tyr-X-Ahp − H2O + H | 406.1509 | 406.1515 | 420.1648 | 400.2181 |
| AcPro-Gln-Thr-Leu − H2O + H | 464.2465 | 464.2447 | 464.2467 | 464.2468 |
| AcPro-Gln-Thr-Leu-Ahp-X − 2H2O + H | - | - | - | 672.3922 |
| AcPro-Gln-Thr-Leu-Ahp-X − H2O + H | - | - | - | 690.3777 |
| Thr-Leu-Ahp-X-NMe-R1-Tyr-Z − H2O + 2H | 727.0780 | 733.3705 | 747.3811 | 727.4318 |
| AcPro-Gln-Thr-(Z-NMe-R1-Tyr)-(Leu) + H | - | - | 806.3871 | 786.4305 |
| AcPro-Gln-Thr-Leu-Ahp-Z-NMe-R1Tyr − H2O + H | - | - | 901.4207 | 881.4576 |
| AcPro-Gln-[Thr-Leu-Ahp-X-NMe-R1-Tyr-Z] − H2O +H | 986.4693 | 1000.4815 | 1014.5018 | 994.5553 |
| AcPro-Gln-[Thr-Leu-Ahp-X-NMe-R1-Tyr-Z] + H | 1004.4831 | 1018.5011 | 1032.5170 | 1012.5737 |
a X amino acid in the fourth position: Val for compounds 30 and 32 and Leu for 34, 36-37; Z amino acid in the sixth position: Val for 30 and 36 and Leu for 32, 34, and 37; R1: O-methyl for compounds 36 and 37; Cl for compounds 30, 32, and 34; b - not detected.
Table 13.
Product ion spectra data for compounds 29, 31, 33, and 35–36.
| Product Ion Assignment a | 29 (m/z) | 31 (m/z) | 33 (m/z) | 35 (m/z) | 36 (m/z) |
|---|---|---|---|---|---|
| Pro immonium | -b | - | - | - | 70.0563 |
| Val immonium | - | - | - | - | - |
| Leu immonium | 86.0924 | - | - | - | 86.0876 |
| NAcPro immonium | 112.0755 | 112.0755 | 112.0755 | 112.0688 | 112.0679 |
| NAcPro + H | 140.0618 | 140.0693 | 140.0693 | 140.0693 | 140.0696 |
| NMe-R1-Tyr immonium | 164.1024 | 150.0907 | 164.1024 | 164.1024 | 164.1052 |
| Leu-Ahp − H2O − CO + H | 181.1353 | 181.1267 | - | - | 181.1283 |
| Leu-Ahp − H2O + H | 209.1243 | 209.1335 | - | 209.1244 | 209.1331 |
| Ac-Pro-Gln + H | 268.1258 | 268.1258 | - | 268.1258 | 268.1207 |
| Ac-Pro-Gln-Thr − H2O + H | 351.1645 | 351.1637 | 351.1638 | 351.1658 | 351.1642 |
| NMe-R1-Tyr-X-Ahp − H2O + H | 386.2025 | 386.2008 | 386.2064 | 386.2034 | 400.2250 |
| AcPro-Gln-Thr-Leu − H2O + H | 464.2439 | 464.2464 | 464.2493 | 464.2495 | 464.2549 |
| AcPro-Gln-Thr-Leu-Ahp-X − 2H2O + H | - | - | - | 658.3446 | - |
| AcPro-Gln-Thr-Leu-Ahp-X − H2O + H | - | - | - | - | - |
| Thr-Leu-Ahp-X-NMe-R1-Tyr-Z − H2O + 2H | 699.4064 | 713.4158 | 699.4065 | 713.4195 | 713.3971 |
| AcPro-Gln-Thr-(Z-NMe-R1-Tyr)-(Leu) + H | - | 772.3942 | - | 786.4170 | - |
| AcPro-Gln-Thr-Leu-Ahp-X-NMe-R1Tyr − H2O + H | - | - | - | 867.4460 | 867.4647 |
| AcPro-Gln-[Thr-Leu-Ahp-X-NMe-R1-Tyr-Z] − H2O +H | 966.5061 | 980.5489 | 966.5144 | 980.5472 | 980.5382 |
| Ac-Pro-Gln-[Thr-Leu-Ahp-X-NMe-R1-Tyr-Z] + H | 984.5381 | 998.5531 | 984.5398 | 998.5550 | 998.5564 |
a X amino acid in the fourth position: Val for compounds 29, 33 and 35 and Leu for 31 and 36; Z amino acid in the sixth position: Val for 29, 33 and 36 and Leu for 31, 35 and 37; R1: O-methyl for compounds 29, 31, 33, 35 and 36; b - not detected.
Table 14.
Product ion spectra data for compound 28 and 38.
| Product Ion Assignment a | 28 (m/z) | 38 (m/z) |
|---|---|---|
| Pro immonium | - b | - |
| Leu immonium | 86.08763 | - |
| Y immonium | - | 126.0845 |
| Y + H | - | 154.0743 |
| NMe-R1-Tyr immonium | 184.0486 | 164.1060 |
| Leu-Ahp − H2O − CO + H | - | - |
| Leu-Ahp − H2O + H | 209.1239 | - |
| Y-Gln + H | 268.1207 | 288.1510 |
| Ac-Pro-Gln-Thr − H2O + H | 321.1521 | 365.1662 |
| NMe-R1-Tyr-X-Ahp − H2O + H | 420.1585 | 400.2211 |
| Y-Gln-Thr-Leu − H2O + H | 434.2416 | 478.2652 |
| Y-Gln-Thr-Leu-Ahp-X − 2H2O + H | - | 686.5626 |
| Y-Gln-Thr-Leu-Ahp-X − H2O + H | - | - |
| Thr-Leu-Ahp-X-NMe-R1-Tyr-Z − H2O + 2H | - | 727.4402 |
| Y-Gln-Thr-(Z-NMe-R1-Tyr)-(Leu) + H | - | 800.4147 |
| Y-Gln-Thr-Leu-Ahp-X-NMe-R1Tyr − H2O + H | - | - |
| Y-Gln-[Thr-Leu-Ahp-X-NMe-R1-Tyr-Z] − H2O +H | 984.4914 | 1008.5481 |
| Y-Gln-[Thr-Leu-Ahp-X-NMe-R1-Tyr-Z] + H | 1002.5031 | 1026.5858 |
a Y exocyclic amino acid in position 1: methyl-dehydroproline (Mdhp) for compound 28 and N-propanoyl-proline for compound 38; X amino acid in the fourth position: Leu; Z amino acid in the sixth position Leu; R1: Cl for compound 28 and O-methyl for compound 38; b - not detected.
This family of cyclic peptides with high structural variability featured a ring formed by six amino acids and a side chain of different lengths and composition [3]. An ester bond between the hydroxyl group of the threonine in position one and the carboxyl group of the terminal amino acid cyclizes the ring. The threonine amino acid in position 1 is occasionally replaced by 3-hydroxy-4-methylproline [67,68]. The 3-amino-6-hydroxy-2-piperidone amino acid (Ahp) always occupied position 3, while a methylated aromatic amino acid and other neutral amino acids are found in positions 5 and 6, respectively. The highly variable side chain may contain an aliphatic fatty acid or a glyceric acid, which is attached either directly to the threonine in position 1 or through one or two amino acids [3]. This family of peptides is often described as protease inhibitors [69,70,71,72,73]. Cyanopeptolins have been isolated mostly from Microcystis [71,74,75] but also from other genera such as Lyngbya [76,77], Nostoc [67,78], Oscillatoria [79,80], Planktothrix [60,81], Scytonema [82], or Symploca [72].
Figure 5 shows the proposed depsipeptide cyclic structure for compound 37 (Ac-Pro-Gln[Thr-Leu-Ahp-Leu-NMe-OMe-Tyr-Leu]) and its predicted fragmentation pattern. The most abundant ions observed in the product ion spectrum of this compound were attributed to the loss of amino acid residues from the C-terminus (m/z 881.4576, 690.3777, 464.2468, 351.1633, and 268.1271) of the dehydrated protonated molecule at m/z 994.5553. This precursor ion was suggested to be generated by the cleavage of the ester linkage accompanied by the dehydration of the Thr [34]. Further loss of the acetyl-proline-glutamine side chain from this linear ion was also noted (m/z 727.4318). Simultaneously, evidence of dehydration at the Ahp position was observed at m/z 400.2184 and 786.4305. An inspection of the low m/z region of the spectrum also supported the proposed structure, revealing the presence of ions associated with the amino acids Ahp (m/z 209.1286 and 181.1300), NMeOMeTyr (m/z 164.1052), AcPro (m/z 112.0679 and 140.0703), Leu (m/z 86.0979), and Pro (m/z 70.0563). To our knowledge, the N-acetyl-proline-glutamine side chain has not been
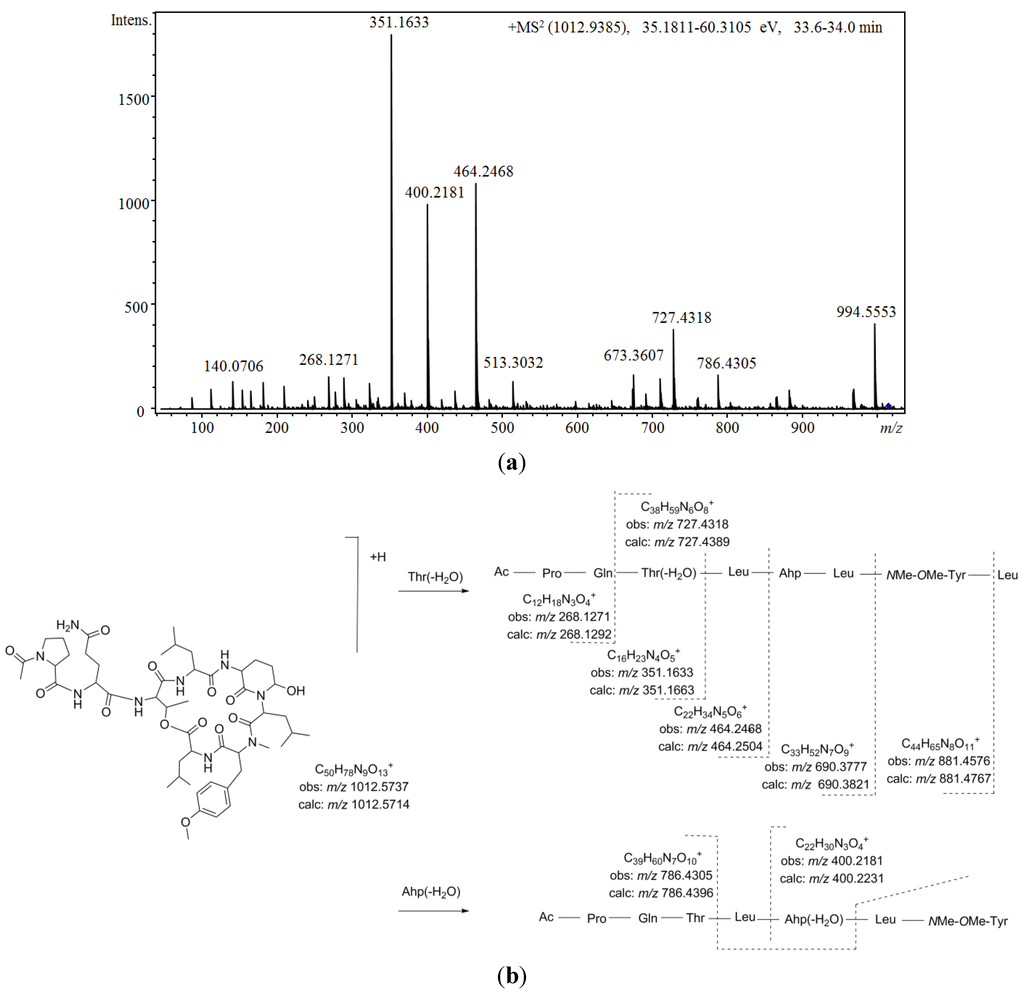
Figure 5.
(a) Product ion spectrum for [M + H]+ of compound 37 and (b) its predicted fragmentation pattern. Conditions as described in the experimental section.
Figure 5.
(a) Product ion spectrum for [M + H]+ of compound 37 and (b) its predicted fragmentation pattern. Conditions as described in the experimental section.
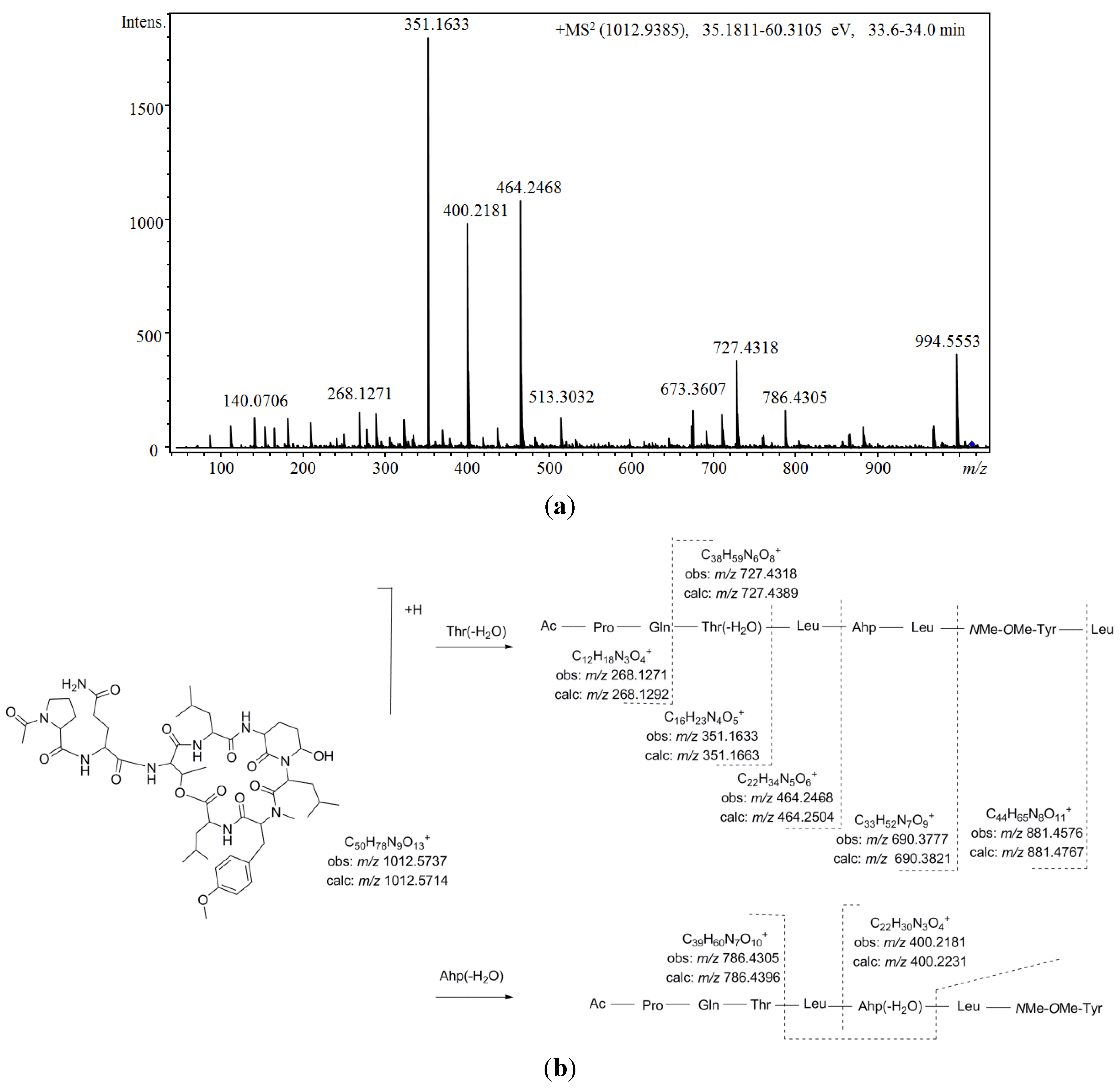
Comparing the aforementioned data with those obtained for compound 36, a highly similar structure was deduced. Differences of 14 u in the protonated molecules and a fragment ion at m/z 713.3971 in combination with the existence of product ions at m/z 464.2549 and 400.2250 indicated that the leucine attributed to position 6 in compound 37 was replaced by valine. In addition, two other isobaric compounds were observed at earlier retention times (31 at 30.0 min, and 35 at 31.1 min). Structural differences from compound 36 were proposed based on mass data (Table 13). Thus, monomethylated tyrosine was suggested for compound 31 (m/z 772.3942, 713.3942, and 150.0907) and a valine residue occupying the fourth position instead of the sixth position was suggested for 35 (m/z 867.4460, 786.4170, 713.4195, and 164.1024). Furthermore, two other minor compounds (29 and 33) with molecular masses 14 Da lower than the compounds mentioned above and with similar product ion spectra were also detected. Based on the mass results (m/z 966.5061/966.5144 and 699.4064/699.4065), the incorporation of valine at positions 4 and 6 was postulated for this pair of diastereoisomers. Simultaneously with these compounds, another congener was observed at m/z 1026.5858 [M + H]+ (38). Propanoylation of the proline residue instead of the more commonly detected acetylation was suggested for this compound on the basis of its product ion spectrum, which differed in product ions containing the side chain (m/z 365.1662, 478.2652, and 686.5626) in combination with the existence of product ions at m/z 126.0845 and 154.0743 (attributed to N-propanoyl-proline).
Similarly, four additional compounds (28, 30, 32, and 34) exhibited fragmentation patterns highly similar to that of compound 37. While most of the mass spectrum remained conserved, fragment ions containing the original dimethylated tyrosine amino acid exhibited m/z shifts when compared to that of the model compound. Additionally, the isotopic pattern of these fragment ions in conjunction with that of the protonated molecule revealed the presence of a chlorine atom in their structures. For compound 34, these fragment ions shifted 20 u (m/z 901.4207, 806.3871, 747.3811, 420.1648, and 184.0523), leading us to propose the chlorination of a methylated tyrosine at position 5. For compound 32, shifts of 20 and 14 u were observed (m/z 733.3705, 406.2447, and 184.0486), suggesting that in addition to the modification mentioned above, a substitution of leucine with valine at position 4 also occurred. Based on the same logic, a structure similar to compound 34, just with the leucine residue at position 6 replaced by valine, was proposed for compound 30. Finally, for compound 28, differences of 30 u in product ions containing the side chain were observed in comparison to compound 34 (m/z 321.1521 and m/z 434.2416). Thus, the same cyclic peptide was proposed for compound 28 with the side chain tentatively attributed to methylated-dehydroproline-glutamine (Mdhp-Gln) or other analogs. The chlorinated and methylated tyrosine amino acid proposed for position 5 was quite unusual and has only been observed in a small number of cyanopeptolins [71,80].
The structural similarity of these compounds to other cyanopeptolins with observed protease inhibitory activity warrants further bioactivity assays. Trypsin inhibitory selectivity was suggested to be related to the existence of basic residues adjacent to Ahp, while chymotrypsin selectivity was proposed to be related to hydrophobic residues. Additionally, residues in other positions as side chains or in the fifth position appear to influence this activity [5,72,83]. These assays will be able to establish the influence of the particular properties of these compounds on the selectivity and potency of these and other activities.
3. Experimental Section
3.1. Strains of Cyanobacteria and Cultivation Conditions
The 40 surveyed cyanobacterial strains belong to the culture collection of the Center for Nuclear Energy in Agriculture Collection/University of São Paulo (CENA/USP), Brazil. All strains were isolated from the leaves of four plant species—Euterpe edulis, Guapira opposita, Garcinia gardneriana, and Merostachys neesii—that were collected in two regions of the Parque Estadual Serra do Mar (Southeastern Brazil). The isolates (three Choococcales, 13 Pseudanabaenales, and 24 Nostocales) were previously identified using both morphological analysis and phylogenies based on the 16S rRNA gene [30] (Table 1). The cultures of cyanobacteria were maintained in liquid BG11 medium under white fluorescent light (30 mmol photons·m−2·s−1) with a 14:10 h light/dark cycle at 25 ± 1 °C under constant agitation (150 rpm) for 21 days. The cells were then concentrated by centrifugation (7000× g, 5 min), washed three times in saline solution (NaCl 0.8%), and re-inoculated into 500-mL flasks containing 200 mL of medium and cultured for a further 21 days.
3.2. Extraction of Peptides
The cell suspensions were concentrated by centrifugation at 7000× g for 5 min and lyophilized (SNL216V, Thermo Electron Corporation). The lyophilized cells (10 mg) were extracted three times with 70% methanol via probe sonication (amplitude of 30%, 2 min, Soni Omni Disruptor), centrifuged (9000× g, 4 °C, 10 min) and concentrated under a stream of nitrogen (TE-concentrator, Techal). The residue was redissolved in 1 mL of methanol:water (50/50, v/v) for LC/MS analysis.
3.3. LC/MS Analyses
Analyses were carried out on a Shimadzu Prominence Liquid Chromatography system coupled to a quadrupole time-of-flight mass spectrometer (Micro TOF-QII; Bruker Daltonics, MA, USA) with an ESI interphase. Separations were achieved using a Luna C18 (2) column (250 mm × inner diameter 3:00, 5 μm) (Phenomenex, Torrance, CA, USA) protected with a guard column of the same material. Samples (5 μL) were eluted using a mobile phase A (water, 0.1% formic acid, and 5 mM ammonia formate) and a mobile phase B (acetonitrile). The gradient increased linearly from 5% to 90% B over 50 min at a flow rate of 0.2 mL/min. The ionization source conditions were as follows: positive ionization, capillary potential of 3500 V, temperature and flow of drying gas (nitrogen) of 5 mL/min and 300 °C, respectively, nebulizer pressure of 35 psi. Mass spectra were acquired using electrospray ionization in the positive mode over the range of m/z from 50 to 3000. The Q/TOF instrument was operated in scan and AutoMS/MS mode, performing MS/MS experiments on the three most intense ions from each MS survey scan. Three collision-induced dissociation CID experiments were performed by varying the collision energies from 30 to 70 to produce many fragment ions of high abundance. The collision energies for fragmentation were as follows. Experiment 1: m/z 0–500, (a) 35 eV (single charged precursor ions) and (b) 25 eV (doubly charged charge); m/z 500–1000, (a) 50 eV and (b) 40 eV; and m/z 1000–2000, (a) 70 eV and (b) 50 eV. Experiment 2: m/z 0–500 (a) 50 eV and (b) 40 eV; m/z 500–1000, (a) 65 eV and (b) 55 eV; and m/z 1000–2000; (a) 85 eV and (b) 65 eV. Experiment 3: m/z 0–500, (a) 35 eV and (b) 25 eV; m/z 500–1000, (a) 65 eV and (b) 55 eV; and m/z 1000–2000, (a) 70 eV and (b) 50 eV with a 75–150% collision energy sweep. The mass spectrometer was calibrated externally with a 10 mM sodium formate cluster solution consisting of 10 mM sodium hydroxide and 0.1% formic acid in water-isopropanol 1:1 (v:v). The accurate mass data were processed using Data Analysis 4.0 software (Bruker Daltonics, Bremen, Germany) which provided a ranking of possible elemental formulae (EF) by using the SmartFormulaEditorTM. For each EF, error (deviations between the measured and theoretical mass of a given sum formula) and sigma value (comparisons of the theoretical and the measured isotope pattern of a given formula) are calculated [65]. The confirmation of the elemental formula was based on the widely accepted thresholds of 5 ppm and 20 m Sigma. Every experiment was run in triplicate.
3.4. Chemical
HPLC grade methanol and acetonitrile from J.T. Baker (USA). Ammonium formate and formic acid, both for mass analyses, were obtained from Fluka (Germany).
4. Conclusions
In the present study, liquid chromatography coupled to a quadrupole time-of-flight mass spectrometer and equipped with an ESI interface was successfully applied to study in depth the cyanopeptide composition of 40 cyanobacterial strains from the Brazilian Atlantic Forest. This approach allowed us to tentatively identify 38 peptides, of which 37 had not been previously described in literature, including aeruginosins, anabaenopeptins, and cyanopeptolins. Based on the mass accuracy data in scan and product ion spectra in combination with the isotopic pattern of the deprotonated and product ions, a planar structure was postulated for each of the detected peptides. In addition to the recently reported aeruginosin 865, 10 novel structural variants were described here. Either hexose or glucuronic acid and butanoic, hexanoic, heptanoic, or octanoic acid O-linked to a Choi motif were observed. With respect to anabaenopeptins, this study led to the characterization of 16 anabaenopeptins. Among those anabaenopeptins, two tryptophan-containing anabaenopeptins and 10 additional anabaenopeptins that incorporated the amino acid N-methyl asparagine were identified. Furthermore, on several occasions, ethylation was postulated for the homovariant amino acid in position four. With respect to cyanopeptolins, 11 new variants were characterized. An N-acetyl proline-glutamine side chain mostly featured these compounds. Additionally, four of these compounds contained the unusual chlorinated N-methylated tyrosine. These results highlight the potential of LC-ESI-QTOF-MS for peptide characterization purposes in complex mixtures from small quantities of material. However, the combination of MS to other techniques such as X-ray crystallography or nuclear magnetic resonance (NMR) applied to the pure compounds is necessary for a complete spectroscopic characterization of the proposed peptide structures including the stereochemistry determination.
Among the surveyed strains of cyanobacteria, only nine strains were observed to produce cyanopeptides (three Nostoc sp., two Desmonostoc sp., and four Brasilonema sp.). However, a highly diverse array of new peptide variants was revealed in the producer strains, which emphasizes the potential of underexplored environments as a source of bioactive compounds.
Acknowledgments
This work was supported by the São Paulo State Research Foundation (FAPESP) through a postdoctoral scholarship (Process 2012/25272-0) to Miriam Sanz and by a PhD scholarship (Process 2009/15402-1) to Ana Paula Dini Andreote. This work was also funded by the process FAPESP 2011/51950-3 and 2013/09192-0 to Ernani Pinto and Marli Fatima Fiore, respectively. Marli Fatima Fiore would also like to thank CNPq for a research fellowship (306607/2012-3).
Author Contributions
Miriam Sanz, Ana Paula Dini Andreote, Felipe Augusto Dörr, Marli Fatima Fiore, and Ernani Pinto designed the experiment; Miriam Sanz and Ana Paula Dini Andreote performed the experiments, assisted by Marli Fatima Fiore and Ernani Pinto. LC-MS analyses were assisted by Felipe Augusto Dörr; Miriam Sanz worked in structural elucidation; and all the authors contributed in writing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Grant Burgess, J.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Singh, S.; Kate, B.N.; Banerjee, U. Bioactive compounds from cyanobacteria and microalgae: An overview. Crit. Rev. Biotechnol. 2005, 25, 73–95. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; von Döhren, H. Cyanobacterial peptides–nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar] [CrossRef] [PubMed]
- Gademann, K.; Portmann, C. Secondary metabolites from cyanobacteria: Complex structures and powerful bioactivities. Curr. Org. Chem. 2008, 12, 326–341. [Google Scholar] [CrossRef]
- Villa, F.A.; Gerwick, L. Marine natural product drug discovery: Leads for treatment of inflammation, cancer, infections, and neurological disorders. Immunopharm. Immunot. 2010, 32, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Chlipala, G.E.; Mo, S.; Orjala, J. Chemodiversity in freshwater and terrestrial cyanobacteria—A source for drug discovery. Curr. Drug Targets 2011, 12, 1654–1673. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Rath, J. Extremophilic Cyanobacteria For Novel Drug Development; Springer: Zürich, Switzerland, 2015. [Google Scholar]
- Galindo-Leal, C.; Câmara, I.D.G. Status do hotspot Mata Atlântica: Uma síntese. In Mata Atlântica: Biodiversidade, Ameaças e Perspectivas; Galindo-Leal, C., Câmara, I.G., Eds.; Fundação SOS Mata Atlântica, Conservação Internacional, Centro de Ciências Aplicadas à Biodiversidade: Belo Horizonte, Brazil, 2005. [Google Scholar]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Cons. 2009, 142, 1141–1153. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.; Martinez, I.B.; Neto, O.M.; Vicente, A.C.P.; Kruger, R.H.; Thompson, F.L. Bacterial community diversity in the Brazilian Atlantic forest soils. Microb. Ecol. 2010, 60, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.F.; Sant’Anna, C.L.; Azevedo, M.T.D.P.; Komárek, J.; Kaštovský, J.; Sulek, J. The cyanobacterial genus Brasilonema, gen. nov., a molecular and phenotypic evaluation. J. Phycol. 2007, 43, 789–798. [Google Scholar] [CrossRef]
- Sant’Anna, C.L.; Azevedo, M.; Kastovsky, J.; Komárek, J. Two form-genera of aerophytic heterocytous cyanobacteria from Brazilian rainy forest,“Mata Atlântica”. Fottea 2010, 10, 217–228. [Google Scholar]
- Sant’Anna, C.; Azevedo, M.; Fiore, M.; Lorenzi, A.; Kaštovský, J.; Komárek, J. Subgeneric diversity of Brasilonema (Cyanobacteria, Scytonemataceae). Braz. J. Bot. 2011, 34, 51–62. [Google Scholar]
- Gama, W.A., Jr.; Laughinghouse, H.D.; st Anna, C.L. How diverse are coccoid cyanobacteria? A case study of terrestrial habitats from the Atlantic Rainforest (São Paulo, Brazil). Phytotaxa 2014, 178, 61–97. [Google Scholar] [CrossRef]
- Silva-Stenico, M.E.; Silva, C.S.P.; Lorenzi, A.S.; Shishido, T.K.; Etchegaray, A.; Lira, S.P.; Moraes, L.A.B.; Fiore, M.F. Non-ribosomal peptides produced by Brazilian cyanobacterial isolates with antimicrobial activity. Microbiol. Res. 2011, 166, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Whipps, J.; Hand, P.; Pink, D.; Bending, G.D. Phyllosphere microbiology with special reference to diversity and plant genotype. J. Appl. Microbiol. 2008, 105, 1744–1755. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Aslanian, A.; Yates, J.R. Mass spectrometry for proteomics. Curr. Opin. Chem. Biol. 2008, 12, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Chernushevich, I.; Wilm, M.; Mann, M. De novo peptide sequencing by nanoelectrospray tandem mass spectrometry using triple quadrupole and quadrupole/time-of-flight instruments. In Mass Spectrometry of Proteins and Peptides; Springer: Totowa, NJ, USA, 2000; pp. 1–16. [Google Scholar]
- Welker, M.; Erhard, M. Consistency between chemotyping of single filaments of Planktothrix rubescens (Cyanobacteria) by MALDI-TOF and the peptide patterns of strains determined by HPLC-MS. J. Mass Spectrom. 2007, 42, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Maršálek, B.; Šejnohová, L.; von Doehren, H. Detection and identification of oligopeptides in Microcystis (Cyanobacteria) colonies: Toward an understanding of metabolic diversity. Peptides 2006, 27, 2090–2103. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Christiansen, G.; von Döhren, H. Diversity of coexisting Planktothrix (Cyanobacteria) chemotypes deduced by mass spectral analysis of microystins and other oligopeptides. Arch. Microbiol. 2004, 182, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Erhard, M.; von Döhren, H. Determination of oligopeptide diversity within a natural population of Microcystis spp.(Cyanobacteria) by typing single colonies by matrix-assisted laser desorption ionization–time of flight mass spectrometry. Appl. Environ. Microbiol 2001, 67, 5069–5076. [Google Scholar] [CrossRef] [PubMed]
- Erhard, M.; von Döhren, H.; Jungblut, P.R. Rapid identification of the new anabaenopeptin G from Planktothrix agardhii HUB 011 using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 1999, 13, 337–343. [Google Scholar] [CrossRef]
- Ferranti, P.; Nasi, A.; Bruno, M.; Basile, A.; Serpe, L.; Gallo, P. A peptidomic approach for monitoring and characterising peptide cyanotoxins produced in Italian lakes by matrix-assisted laser desorption/ionisation and quadrupole time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Marzec, H.; Meriluoto, J.; Pliński, M.; Szafranek, J. Characterization of nodularin variants in Nodularia spumigena from the Baltic Sea using liquid chromatography/mass spectrometry/mass spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, R.L.; Dörr, F.A.; Dörr, F.; Bortoli, S.; Delherbe, N.; Vásquez, M.; Pinto, E. Co-occurrence of microcystin and microginin congeners in Brazilian strains of Microcystis sp. FEMS Microbiol. Ecol. 2012, 82, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Andreote, A.P.D.; Fiore, M.D.F. Polyphasic assessment of culturable cyanobacteria from the phyllosphere of Atlantic Forest. J. Phycol. 2015. under review. [Google Scholar]
- Welker, M.; Brunke, M.; Preussel, K.; Lippert, I.; von Döhren, H. Diversity and distribution of Microcystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry. Microbiology 2004, 150, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Mayumi, T.; Noguchi, K.; Kashiwagi, T.; Akashi, S.; Sivonen, K.; Hirayama, K.; Harada, K.-I. Mass spectrometric studies of peptides from cyanobacteria under FAB MS/MS conditions. J. Mass Spectrom. Soc. Jpn. 2000, 48, 56–64. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R. MALDI-TOF mass spectrometry of cyanobacteria: A global approach to the discovery of novel secondary metabolites. Chem. N. Zeal. 2008, 72, 68–71. [Google Scholar]
- Mayumi, T.; Kato, H.; Kawasaki, Y.; Harada, K.I. Formation of diagnostic product ions from cyanobacterial cyclic peptides by the two-bond fission mechanism using ion trap liquid chromatography/multi-stage mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Tooming-Klunderud, A.; Rohrlack, T.; Shalchian-Tabrizi, K.; Kristensen, T.; Jakobsen, K.S. Structural analysis of a non-ribosomal halogenated cyclic peptide and its putative operon from Microcystis: Implications for evolution of cyanopeptolins. Microbiology 2007, 153, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Marzec, H.; Kaczkowska, M.J.; Blaszczyk, A.; Akcaalan, R.; Spoof, L.; Meriluoto, J. Diversity of peptides produced by Nodularia spumigena from various geographical regions. Mar. Drugs 2012, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kapuścik, A.; Hrouzek, P.; Kuzma, M.; Bártová, S.; Novák, P.; Jokela, J.; Pflüger, M.; Eger, A.; Hundsberger, H.; Kopecký, J. Novel aeruginosin-865 from Nostoc sp. as a potent anti-inflammatory agent. Chem. Biochem. 2013, 14, 2329–2337. [Google Scholar]
- Qi, Y.; Rosso, L.; Sedan, D.; Giannuzzi, L.; Andrinolo, D.; Volmer, D.A. Seven new microcystin variants discovered from a native Microcystis aeruginosa strain—Unambiguous assignment of product ions by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2015, 29, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Dörr, F.A.; Oliveira-Silva, D.; Lopes, N.P.; Iglesias, J.; Volmer, D.A.; Pinto, E. Dissociation of deprotonated microcystin variants by collision-induced dissociation following electrospray ionization. Rapid Commun. Mass Spectrom. 2011, 25, 1981–1992. [Google Scholar] [CrossRef] [PubMed]
- Dahlmann, J.; Budakowski, W.R.; Luckas, B. Liquid chromatography–electrospray ionisation–mass spectrometry based method for the simultaneous determination of algal and cyanobacterial toxins in phytoplankton from marine waters and lakes followed by tentative structural elucidation of microcystins. J. Chromatogr. A 2003, 994, 45–57. [Google Scholar] [CrossRef]
- Murakami, M.; Okita, Y.; Matsuda, H.; Okino, T.; Yamaguchi, K. Aeruginosin 298-A, a thrombin and trypsin inhibitor from the blue-green alga Microcystis aeruginosa (NIES-298). Tetrahedron Lett. 1994, 35, 3129–3132. [Google Scholar] [CrossRef]
- Murakami, M.; Ishida, K.; Okino, T.; Okita, Y.; Matsuda, H.; Yamaguchi, K. Aeruginosins 98-A and B, trypsin inhibitors from the blue-green alga Microcystis aeruginosa (NIES-98). Tetrahedron Lett. 1995, 36, 2785–2788. [Google Scholar] [CrossRef]
- Ishida, K.; Okita, Y.; Matsuda, H.; Okino, T.; Murakami, M. Aeruginosins, protease inhibitors from the cyanobacterium Microcystis aeruginosa. Tetrahedron 1999, 55, 10971–10988. [Google Scholar] [CrossRef]
- Elkobi-Peer, S.; Faigenbaum, R.; Carmeli, S. Bromine-and chlorine-containing aeruginosins from Microcystis aeruginosa bloom material collected in Kibbutz Geva, Israel. J. Nat. Prod. 2012, 75, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Aeruginosins 205A and-B, serine protease inhibitory glycopeptides from the cyanobacterium Oscillatoria agardhii (NIES-205). J. Org. Chem. 1997, 62, 1810–1813. [Google Scholar] [CrossRef]
- Ersmark, K.; del Valle, J.R.; Hanessian, S. Chemistry and biology of the aeruginosin family of serine protease inhibitors. Angew. Chem. Int. Ed. 2008, 47, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Vegman, M.; Carmeli, S. Three aeruginosins and a microviridin from a bloom assembly of Microcystis spp. collected from a fishpond near Kibbutz Lehavot HaBashan, Israel. Tetrahedron 2014, 70, 6817–6824. [Google Scholar] [CrossRef]
- Fewer, D.P.; Jokela, J.; Paukku, E.; Österholm, J.; Wahlsten, M.; Permi, P.; Aitio, O.; Rouhiainen, L.; Gomez-Saez, G.V.; Sivonen, K. New structural variants of aeruginosin produced by the toxic bloom forming cyanobacterium Nodularia spumigena. PLoS ONE 2013, 8, e73618. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Usui, T.; Ueda, K.; Osada, H.; Kaya, K. Isolation of new protein phosphatase inhibitors from two cyanobacteria species, Planktothrix spp. J. Nat. Prod. 2001, 64, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Itou, Y.; Suzuki, S.; Ishida, K.; Murakami, M. Anabaenopeptins G and H, potent carboxypeptidase A inhibitors from the cyanobacterium Oscillatoria agardhii (NIES-595). Bioorg. Med. Chem. Lett. 1999, 9, 1243–1246. [Google Scholar] [CrossRef]
- Murakami, M.; Suzuki, S.; Itou, Y.; Kodani, S.; Ishida, K. New anabaenopeptins, potent carboxypeptidase-A inhibitors from the cyanobacterium Aphanizomenon flos-aquae. J. Nat. Prod. 2000, 63, 1280–1282. [Google Scholar] [CrossRef] [PubMed]
- Bjorquist, P.; Buchanan, M.; Campitelli, M.; Carroll, A.; Hyde, E.; Neve, J.; Polla, M.; Quinn, R. Use of Cyclic Anabaenopeptin-Type Peptides for the Treatment of a Condition Wherein Inhibition of Carboxypeptidase U is Beneficial, Novel Anabaenopeptin Derivatives and Intermediates Thereof. U.S. Patent WO2005039617A1, 28 October 2004. [Google Scholar]
- Sano, T.; Kaya, K. Oscillamide Y, a chymotrypsin inhibitor from toxic Oscillatoria agardhii. Tetrahedron Lett. 1995, 36, 5933–5936. [Google Scholar] [CrossRef]
- Adiv, S.; Carmeli, S. Protease Inhibitors from Microcystis aeruginosa Bloom Material Collected from the Dalton Reservoir, Israel. J. Nat. Prod. 2013, 76, 2307–2315. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Sivonen, K.; Nakano, T.; Harada, K.-I. Structural elucidation of cyanobacterial peptides encoded by peptide synthetase gene in Anabaena species. Tetrahedron 2002, 58, 6863–6871. [Google Scholar] [CrossRef]
- Grach-Pogrebinsky, O.; Carmeli, S. Three novel anabaenopeptins from the cyanobacterium Anabaena sp. Tetrahedron 2008, 64, 10233–10238. [Google Scholar] [CrossRef]
- Zi, J.; Lantvit, D.D.; Swanson, S.M.; Orjala, J. Lyngbyaureidamides A and B, two anabaenopeptins from the cultured freshwater cyanobacterium Lyngbya sp. (SAG 36.91). Phytochemistry 2012, 74, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Gesner-Apter, S.; Carmeli, S. Protease inhibitors from a water bloom of the cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2009, 72, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Shin, H.J.; Matsuda, H.; Ishida, K.; Yamaguchi, K. A cyclic peptide, anabaenopeptin B, from the cyanobacterium Oscillatoria agardhii. Phytochemistry 1997, 44, 449–452. [Google Scholar] [CrossRef]
- Okumura, H.S.; Philmus, B.; Portmann, C.; Hemscheidt, T.K. Homotyrosine-containing cyanopeptolins 880 and 960 and anabaenopeptins 908 and 915 from Planktothrix agardhii CYA 126/8. J. Nat. Prod. 2008, 72, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Reshef, V.; Carmeli, S. Schizopeptin 791, a new anabeanopeptin-like cyclic peptide from the cyanobacterium Schizothrix sp. J. Nat. Prod. 2002, 65, 1187–1189. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.I.; Sato, M.; Murayama, T.; Ishibashi, M.; Wälchi, M.R.; Kanai, M.; Shoji, J.; Ohizumi, Y. Konbamide, a novel peptide with calmoduiin antagonistic activity from the Okinawan marine sponge Theonella sp. J. Chem. Soc. Chem. Commun. 1991, 1050–1052. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Harper, M.K.; Faulkner, D.J. Mozamides A and B, cyclic peptides from a theonellid sponge from Mozambique. J. Nat. Prod. 1997, 60, 779–782. [Google Scholar] [CrossRef]
- Müller, D.; Krick, A.; Kehraus, S.; Mehner, C.; Hart, M.; Küpper, F.C.; Saxena, K.; Prinz, H.; Schwalbe, H.; Janning, P. Brunsvicamides A-C: Sponge-related cyanobacterial peptides with Mycobacterium tuberculosis protein tyrosine phosphatase inhibitory activity. J. Med. Chem. 2006, 49, 4871–4878. [Google Scholar] [CrossRef] [PubMed]
- Sisay, M.T.; Hautmann, S.; Mehner, C.; König, G.M.; Bajorath, J.; Gütschow, M. Inhibition of human leukocyte elastase by brunsvicamides A–C: Cyanobacterial cyclic peptides. ChemMedChem 2009, 4, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.; Renner, S.; Waldmann, H.; Arndt, H.D. Synthesis and structure–activity correlation of a brunsvicamide-inspired cyclopeptide collection. ChemBioChem 2009, 10, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Okino, T.; Qi, S.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Nostopeptins A and B, elastase inhibitors from the cyanobacterium Nostoc minutum. J. Nat. Prod. 1997, 60, 158–161. [Google Scholar] [CrossRef]
- Mehner, C.; Müller, D.; Kehraus, S.; Hautmann, S.; Gütschow, M.; König, G.M. New peptolides from the cyanobacterium Nostoc insulare as selective and potent inhibitors of human leukocyte elastase. ChemBioChem 2008, 9, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Jakobi, C.; Oberer, L.; Quiquerez, C.; König, W.A.; Weckesser, J. Cyanopeptolin S, a sulfate-containing depsipeptide from a water bloom of Microcystis sp. FEMS Microbiol. Lett. 1995, 129, 129–133. [Google Scholar] [PubMed]
- Bister, B.; Keller, S.; Baumann, H.I.; Nicholson, G.; Weist, S.; Jung, G.; Süssmuth, R.D.; Jüttner, F. Cyanopeptolin 963A, a chymotrypsin inhibitor of Microcystis PCC 7806. J. Nat. Prod. 2004, 67, 1755–1757. [Google Scholar] [CrossRef] [PubMed]
- Von Elert, E.; Oberer, L.; Merkel, P.; Huhn, T.; Blom, J.F. Cyanopeptolin 954, a chlorine-containing chymotrypsin inhibitor of Microcystis aeruginosa NIVA Cya 43. J. Nat. Prod. 2005, 68, 1324–1327. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; McPhail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2007, 71, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Gademann, K.; Portmann, C.; Blom, J.F.; Zeder, M.; Jüttner, F. Multiple toxin production in the cyanobacterium Microcystis: Isolation of the toxic protease inhibitor cyanopeptolin 1020. J. Nat. Prod. 2010, 73, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.-I.; Mayumi, T.; Shimada, T.; Suzuki, M.; Kondo, F.; Watanabe, M.F. Occurrence of four depsipeptides, aeruginopeptins, together with microcystins from toxic cyanobacteria. Tetrahedron Lett. 1993, 34, 6091–6094. [Google Scholar] [CrossRef]
- Okano, T.; Sano, T.; Kaya, K. Micropeptin T-20, a novel phosphate-containing cyclic depsipeptide from the cyanobacterium Microcystis aeruginosa. Tetrahedron Lett. 1999, 40, 2379–2382. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatin 4, a dolastatin 13 analogue with elastase and chymotrypsin inhibitory activity from the marine cyanobacterium Lyngbya confervoides. J. Nat. Prod. 2007, 70, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Taori, K.; Matthew, S.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatins 5–7, potent elastase inhibitors from Floridian marine cyanobacteria, Lyngbya spp. J. Nat. Prod. 2007, 70, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Kaya, K.; Sano, T.; Beattie, K.A.; Codd, G.A. Nostocyclin, a novel 3-amino-6-hydroxy-2-piperidone-containing cyclic depsipeptide from the cyanobacterium Nostoc sp. Tetrahedron Lett. 1996, 37, 6725–6728. [Google Scholar] [CrossRef]
- Itou, Y.; Ishida, K.; Shin, H.J.; Murakami, M. Oscillapeptins A to F, serine protease inhibitors from the three strains of Oscillatoria agardhii. Tetrahedron 1999, 55, 6871–6882. [Google Scholar] [CrossRef]
- Plaza, A.; Bewley, C.A. Largamides A–H, unusual cyclic peptides from the marine cyanobacterium Oscillatoria sp. J. Org. Chem. 2006, 71, 6898–6907. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.F.; Bister, B.; Bischoff, D.; Nicholson, G.; Jung, G.; Süssmuth, R.D.; Jüttner, F. Oscillapeptin J, a new grazer toxin of the freshwater Cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2003, 66, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Matern, U.; Oberer, L.; Erhard, M.; Herdman, M.; Weckesser, J. Hofmannolin, a cyanopeptolin from Scytonema hofmanni PCC 7110. Phytochemistry 2003, 64, 1061–1067. [Google Scholar] [CrossRef]
- Nagarajan, M.; Maruthanayagam, V.; Sundararaman, M. SAR analysis and bioactive potentials of freshwater and terrestrial cyanobacterial compounds: A review. J. Appl. Toxicol. 2013, 33, 313–349. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).