
Ecklonia cava Polyphenol Has a Protective Effect against Ethanol-Induced Liver Injury in a Cyclic AMP-Dependent Manner


Abstract
:1. Introduction
2. Results
2.1. The Effects of ECP on the Cell Viability of Ethanol-Treated Hepatocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Cell Viability (% of Control) |
|---|---|
| Control | 100.0 ± 6.7 |
| 100 mM ethanol | 82.5 ± 4.8 ** |
| 100 mM ethanol + 3.1 μg/mL ECP | 84.8 ± 5.3 ** |
| 100 mM ethanol + 6.25 μg/mL ECP | 97.2 ± 5.3 |
| 100 mM ethanol + 12.5 μg/mL ECP | 97.2 ± 8.2 |
| 100 mM ethanol + 25 μg/mL ECP | 95.7 ± 5.5 |
| 100 mM ethanol + 50 μg/mL ECP | 89.6 ± 4.7 |
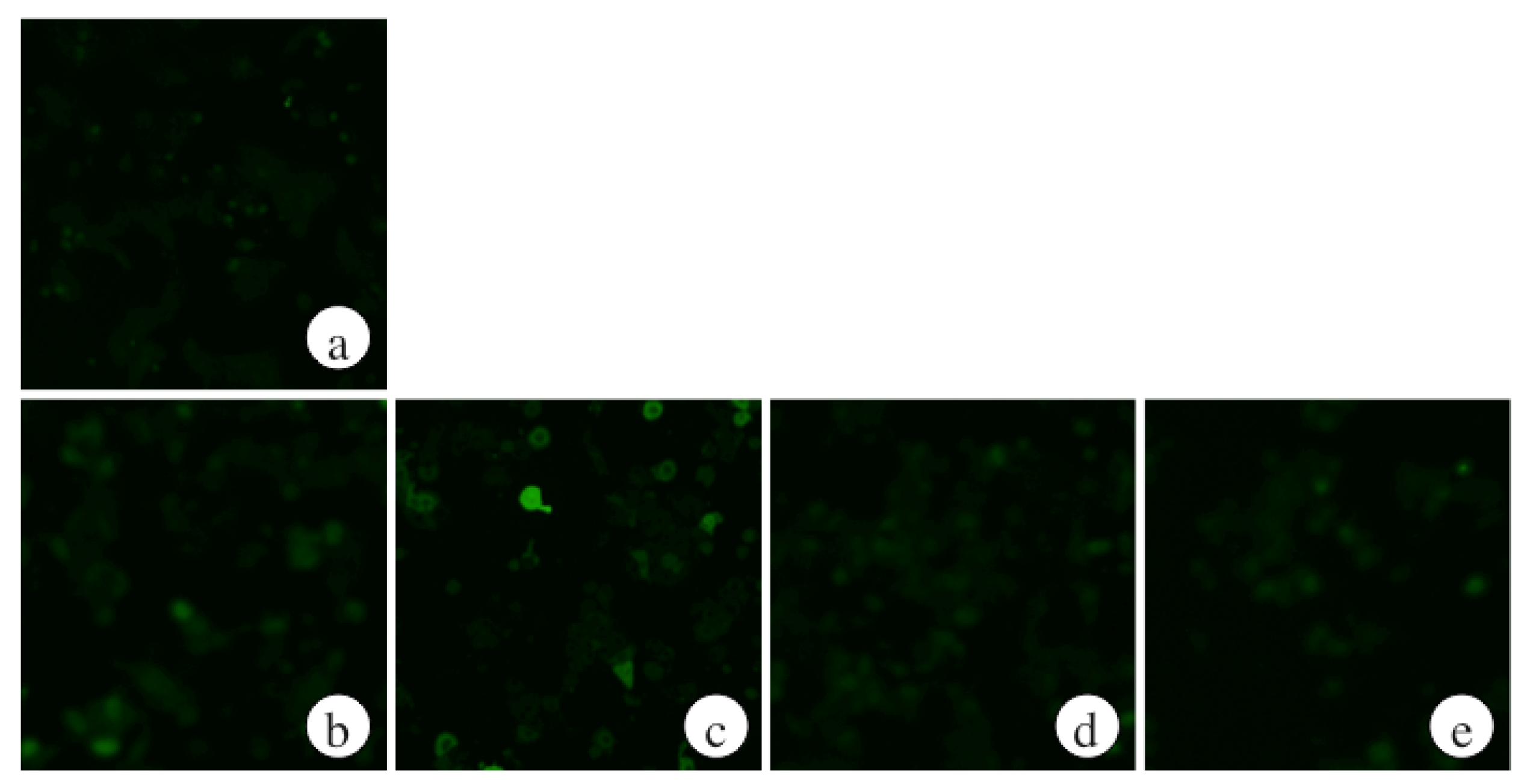
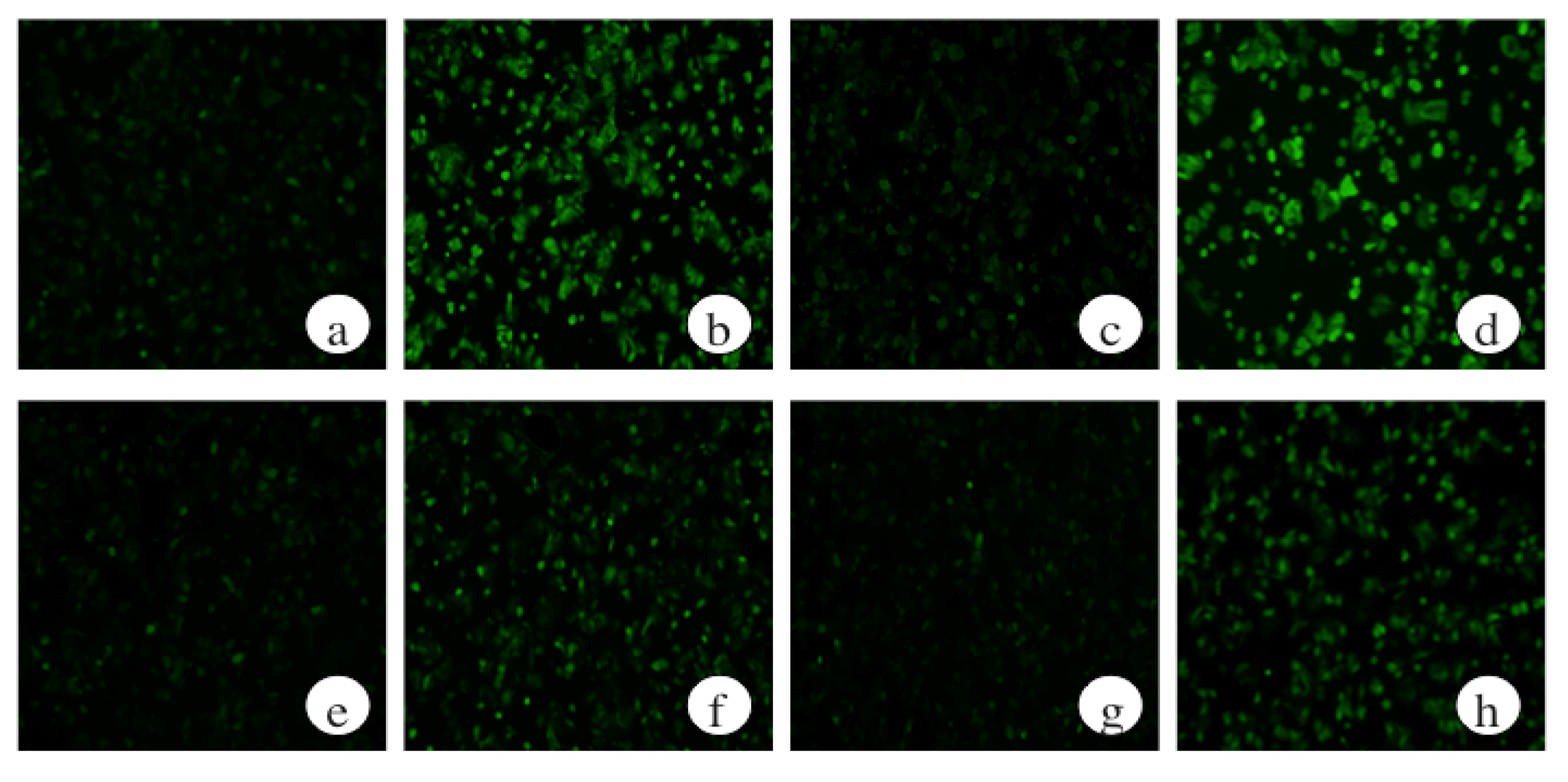
2.2. The Effects of ECP on ROS Levels of Ethanol-Treated Hepatocytes
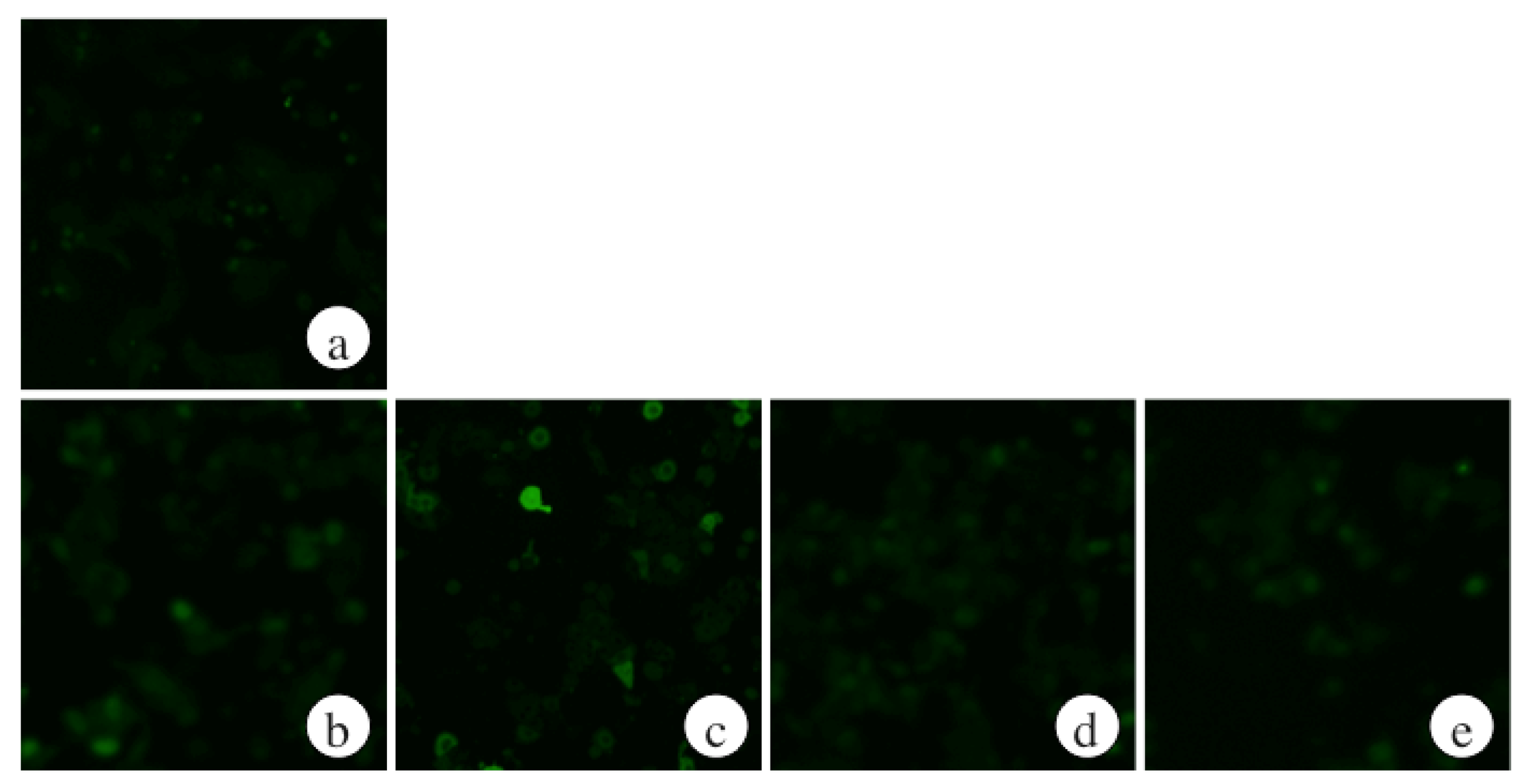
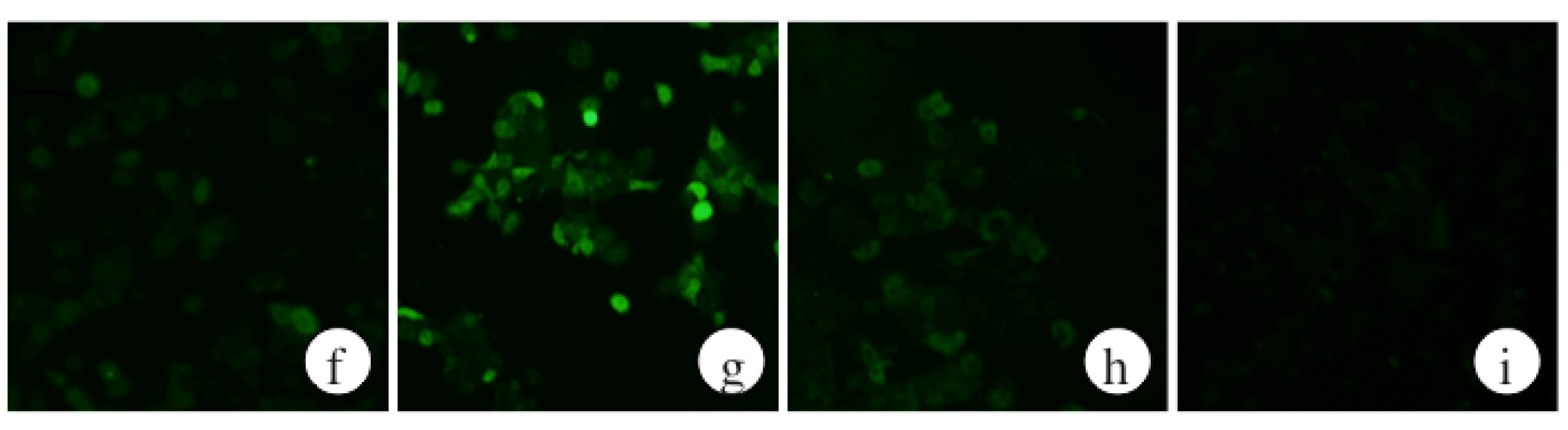
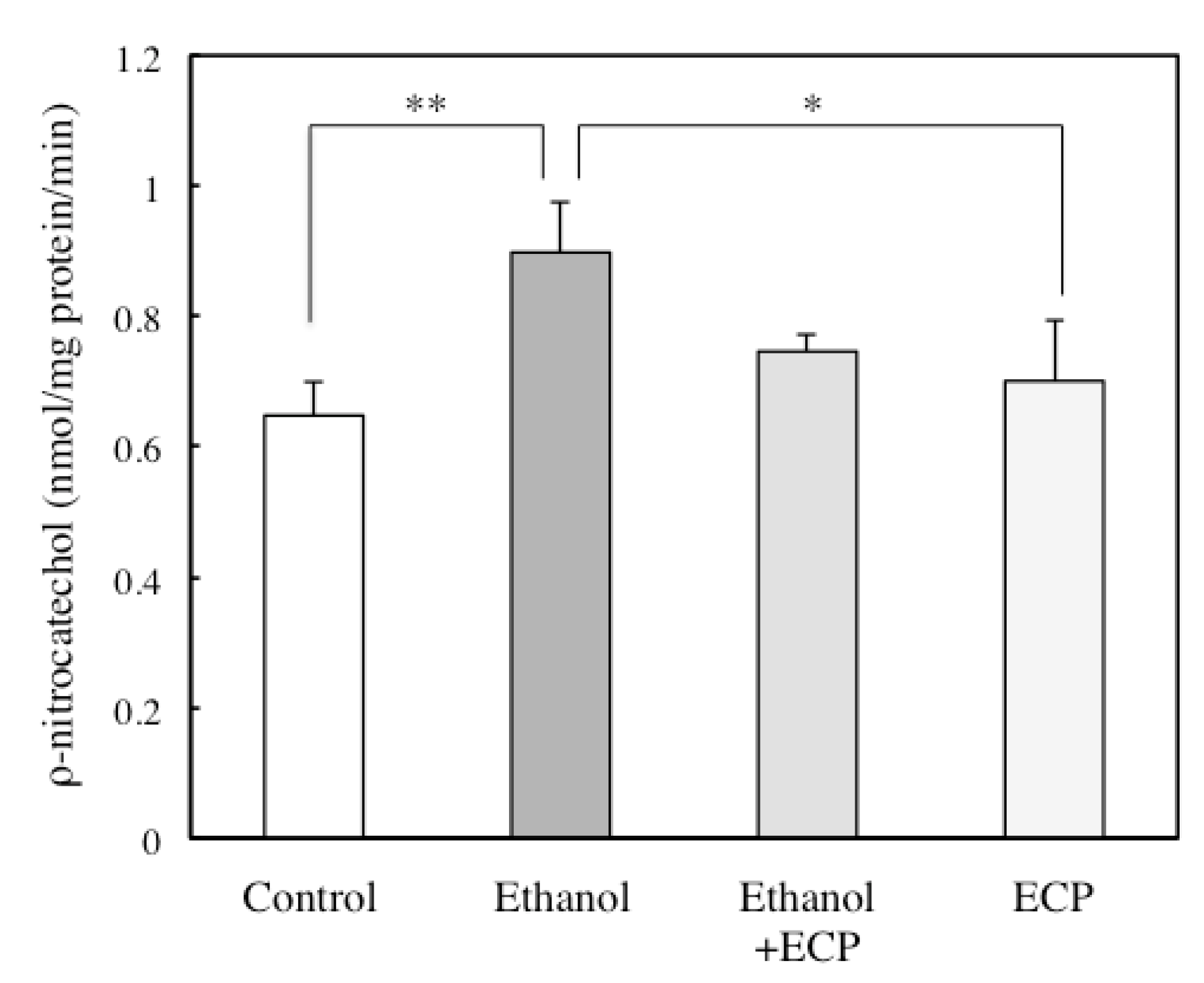
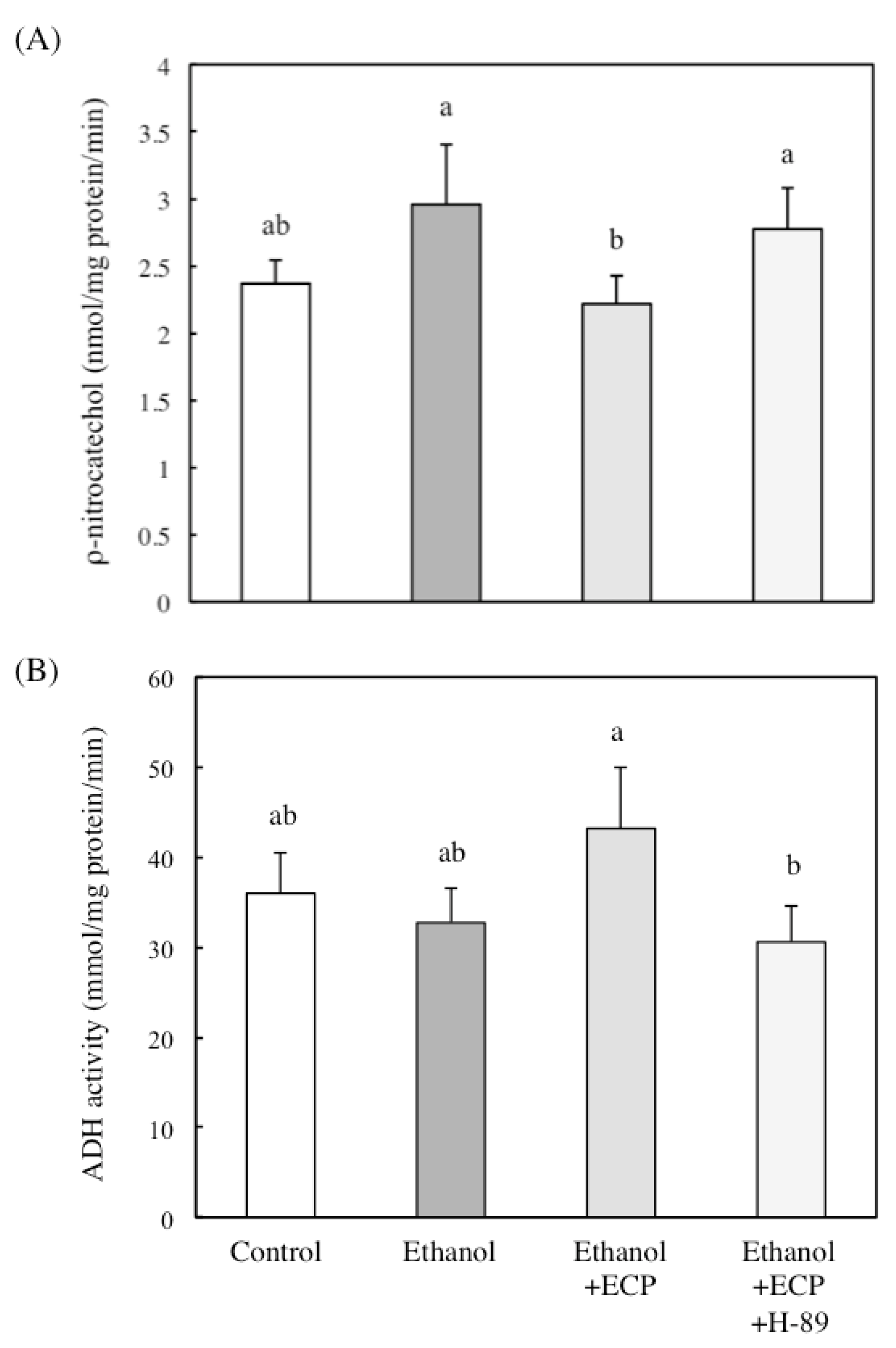
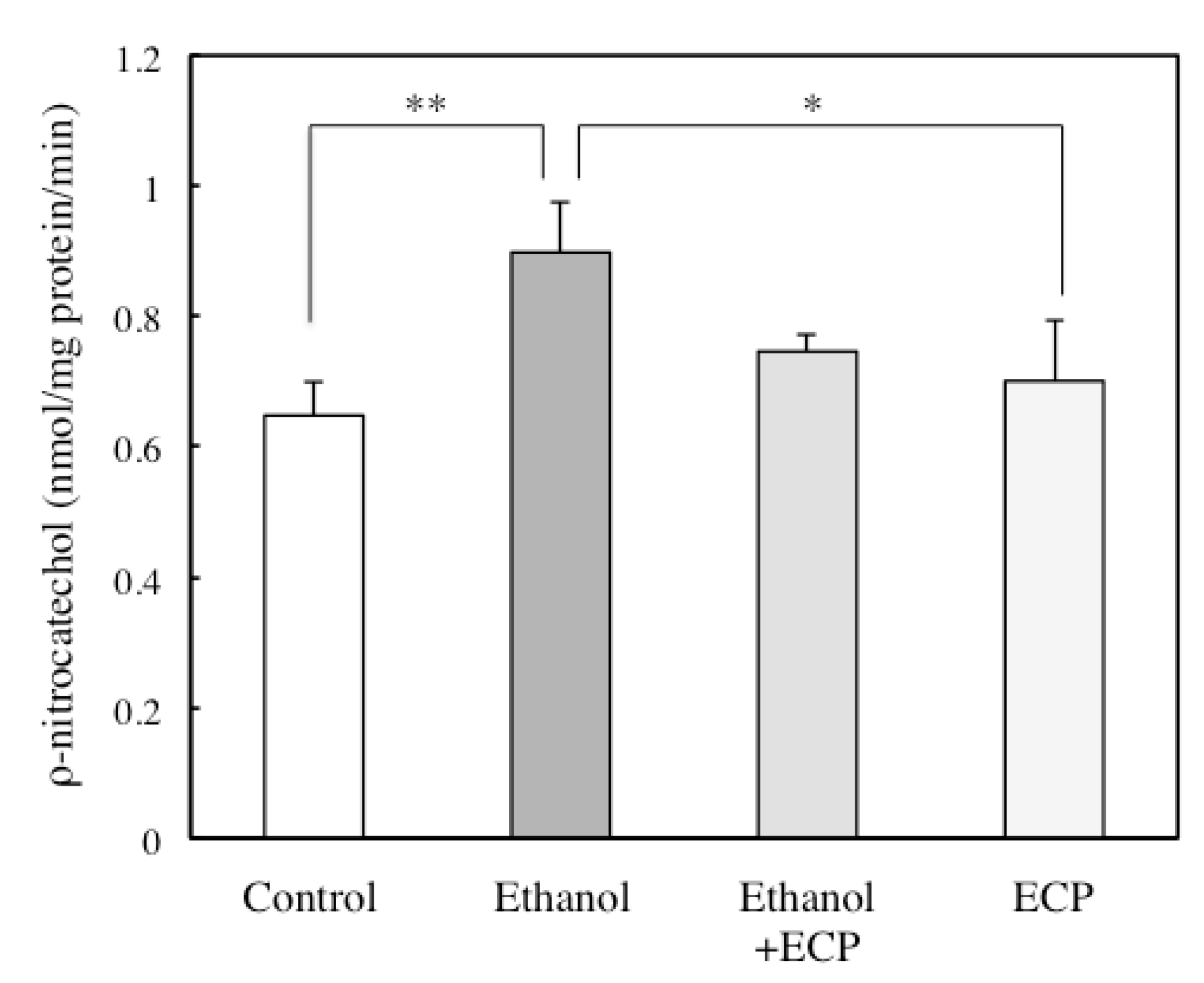
2.3. The Effects of ECP on CYP2E1 Activities of Ethanol-Treated Hepatocytes
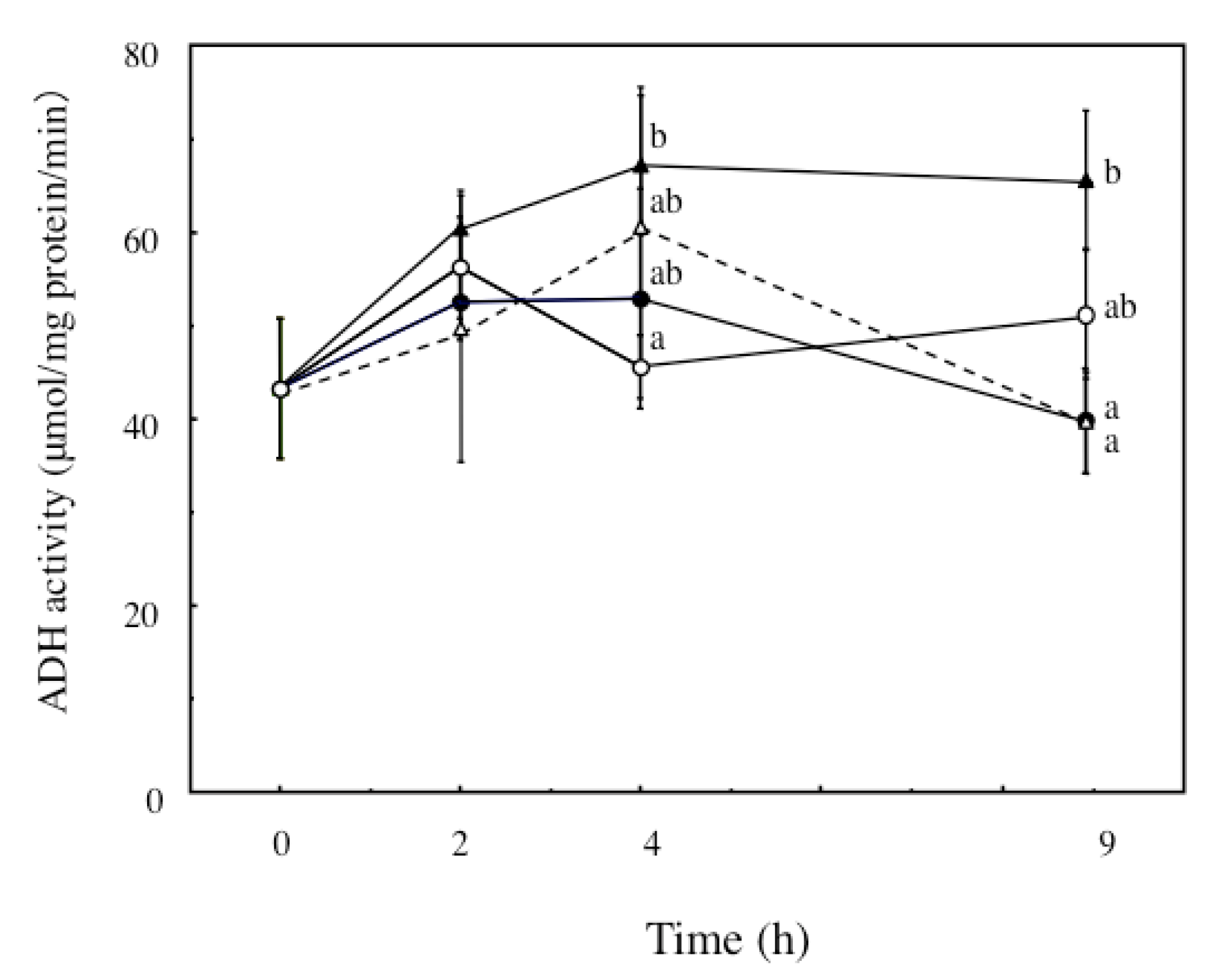
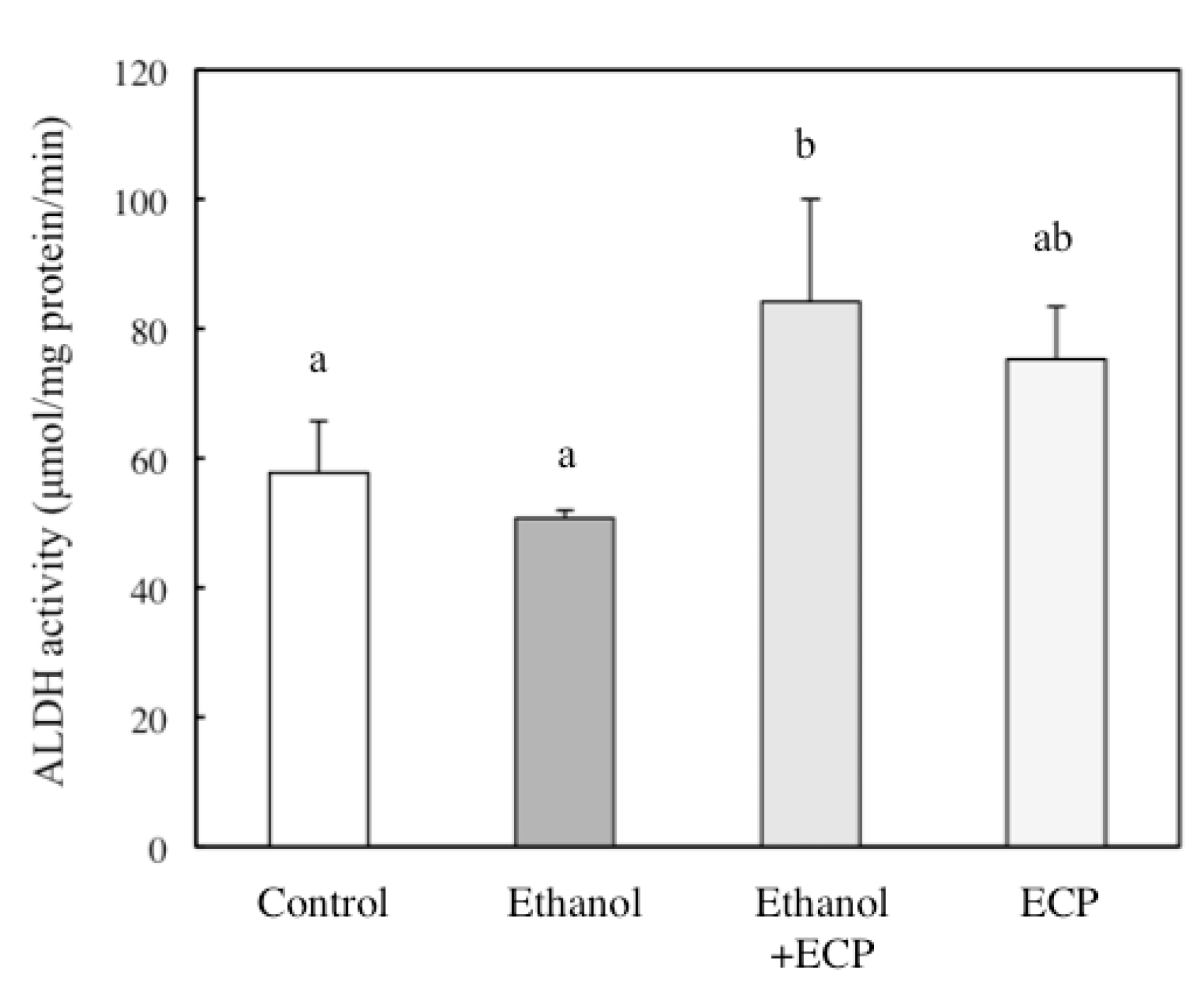
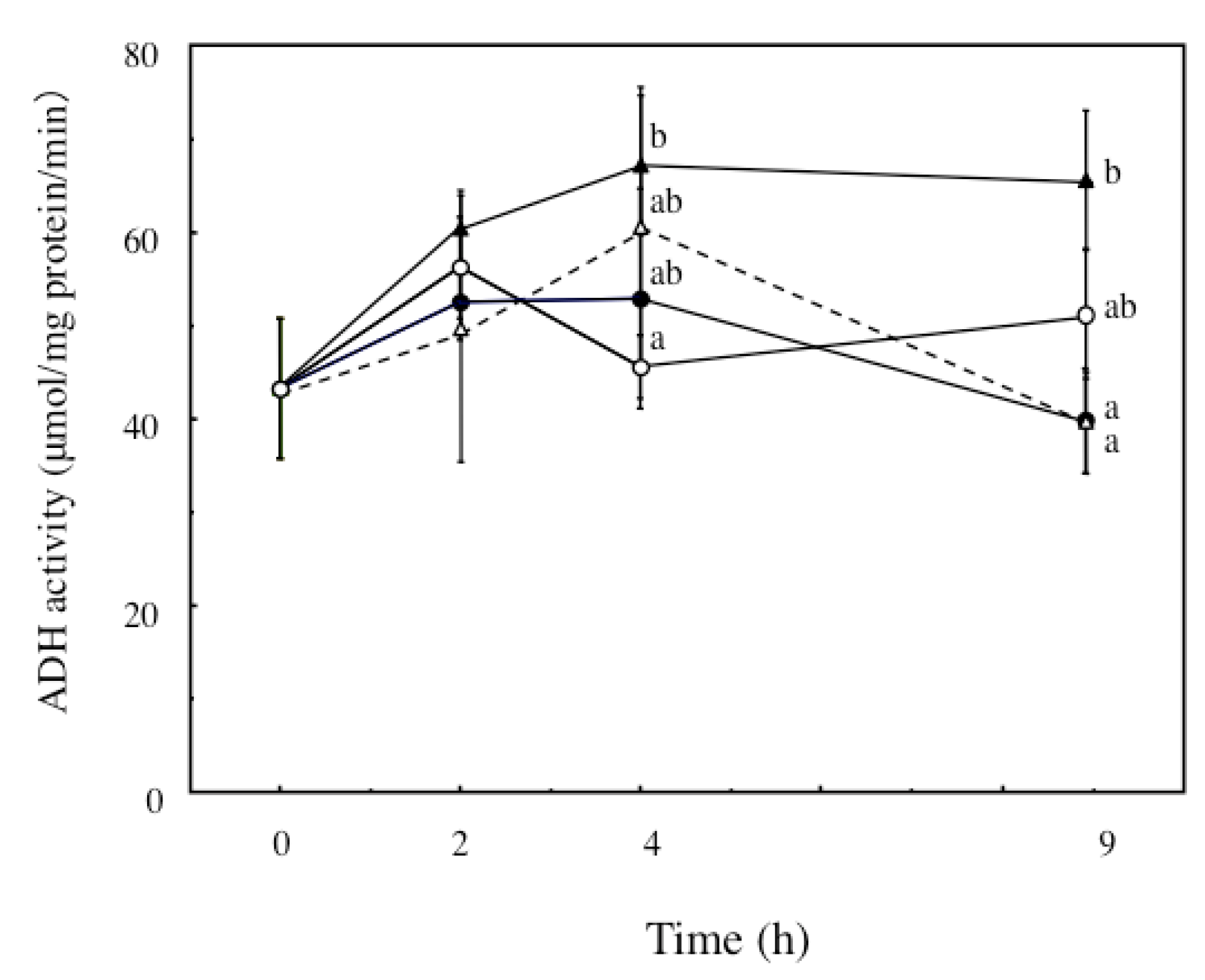
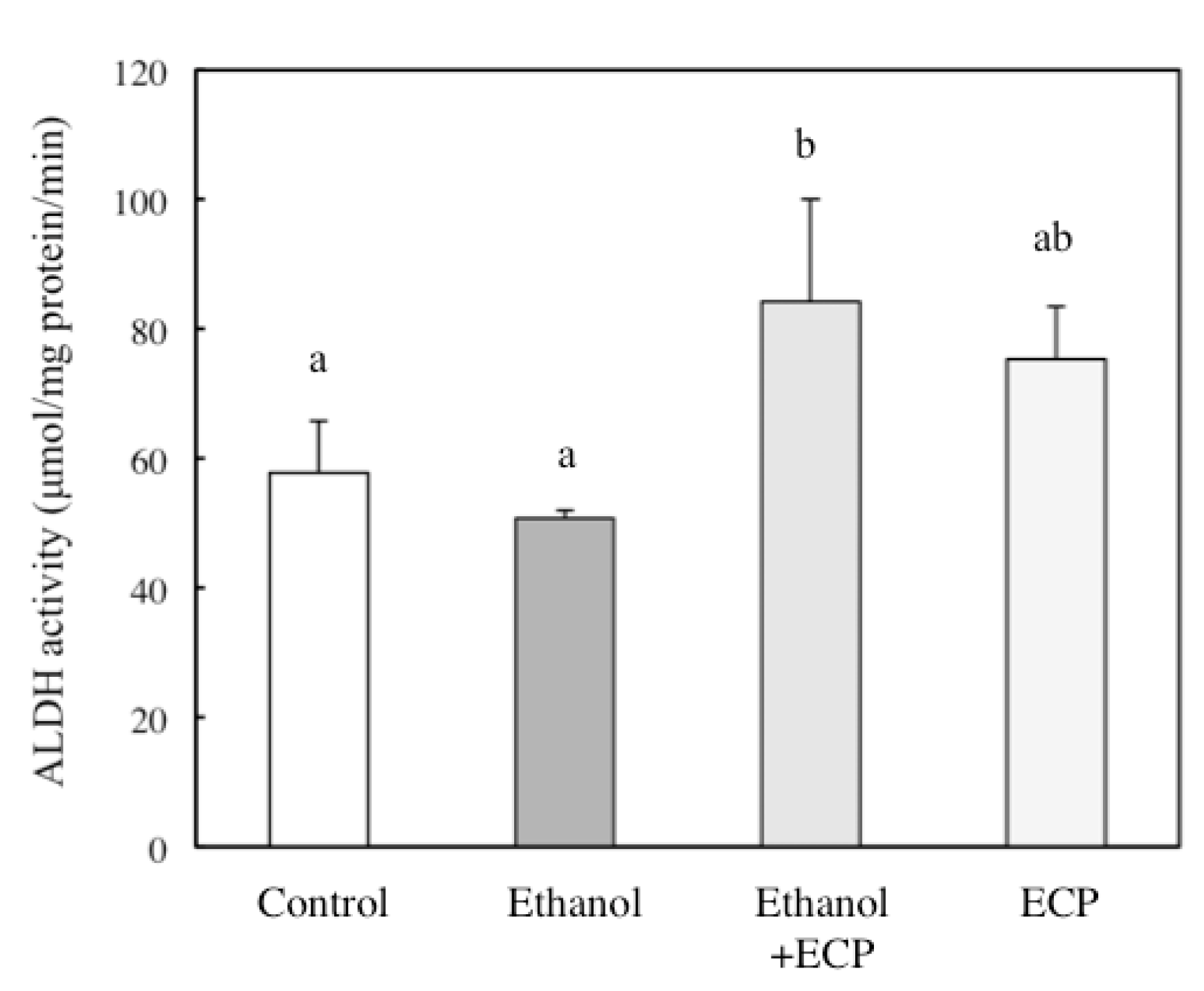
2.4. The Effects of ECP on ADH and ALDH Activities of Ethanol-Treated Hepatocytes
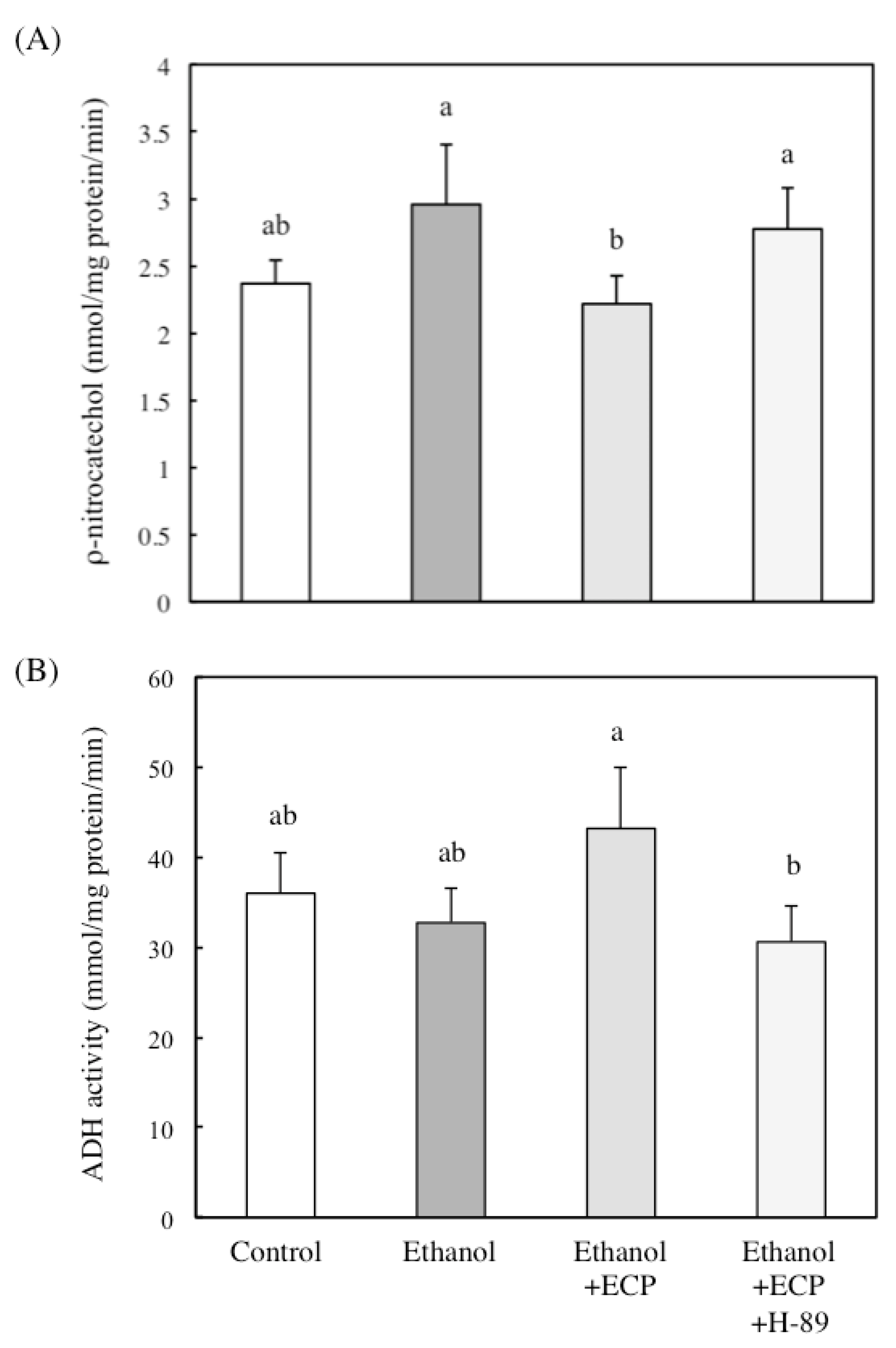
2.5. The Effects of Protein Kinase A Inhibition on CYP2E1 and ADH Activities of Ethanol and ECP-Treated Hepatocytes
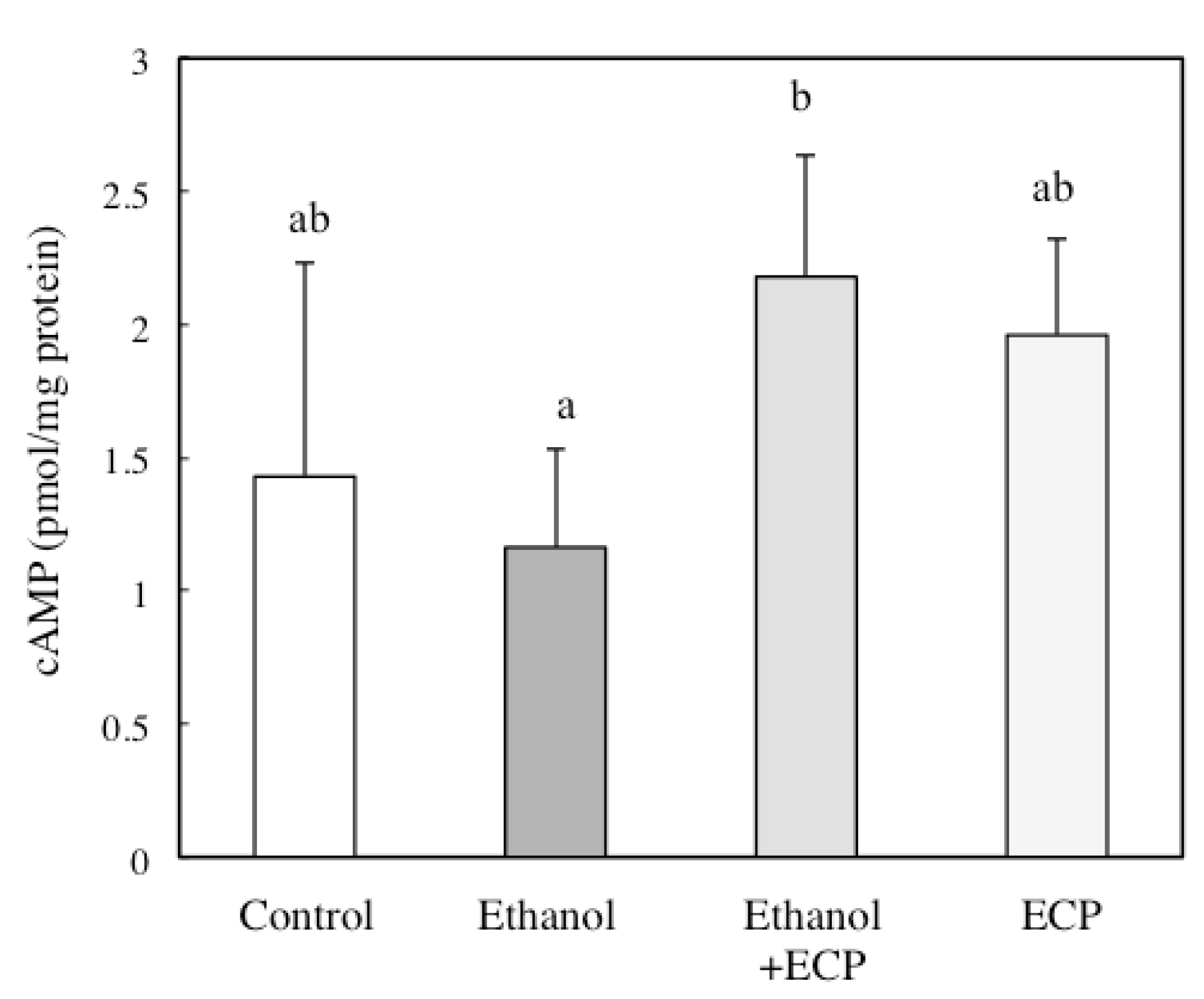
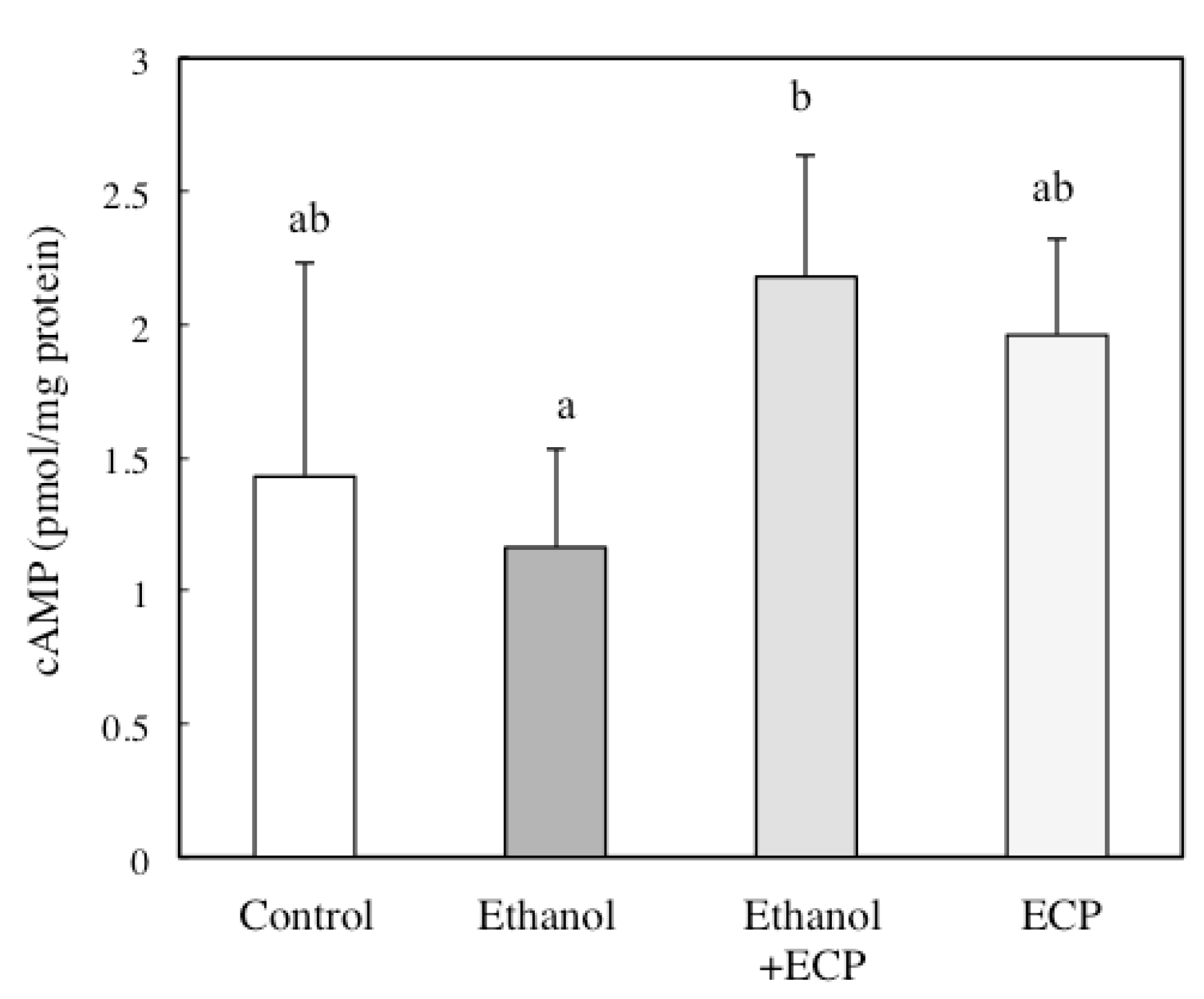
2.6. The Effects of ECP on the cAMP Concentration of Ethanol-Treated Hepatocytes
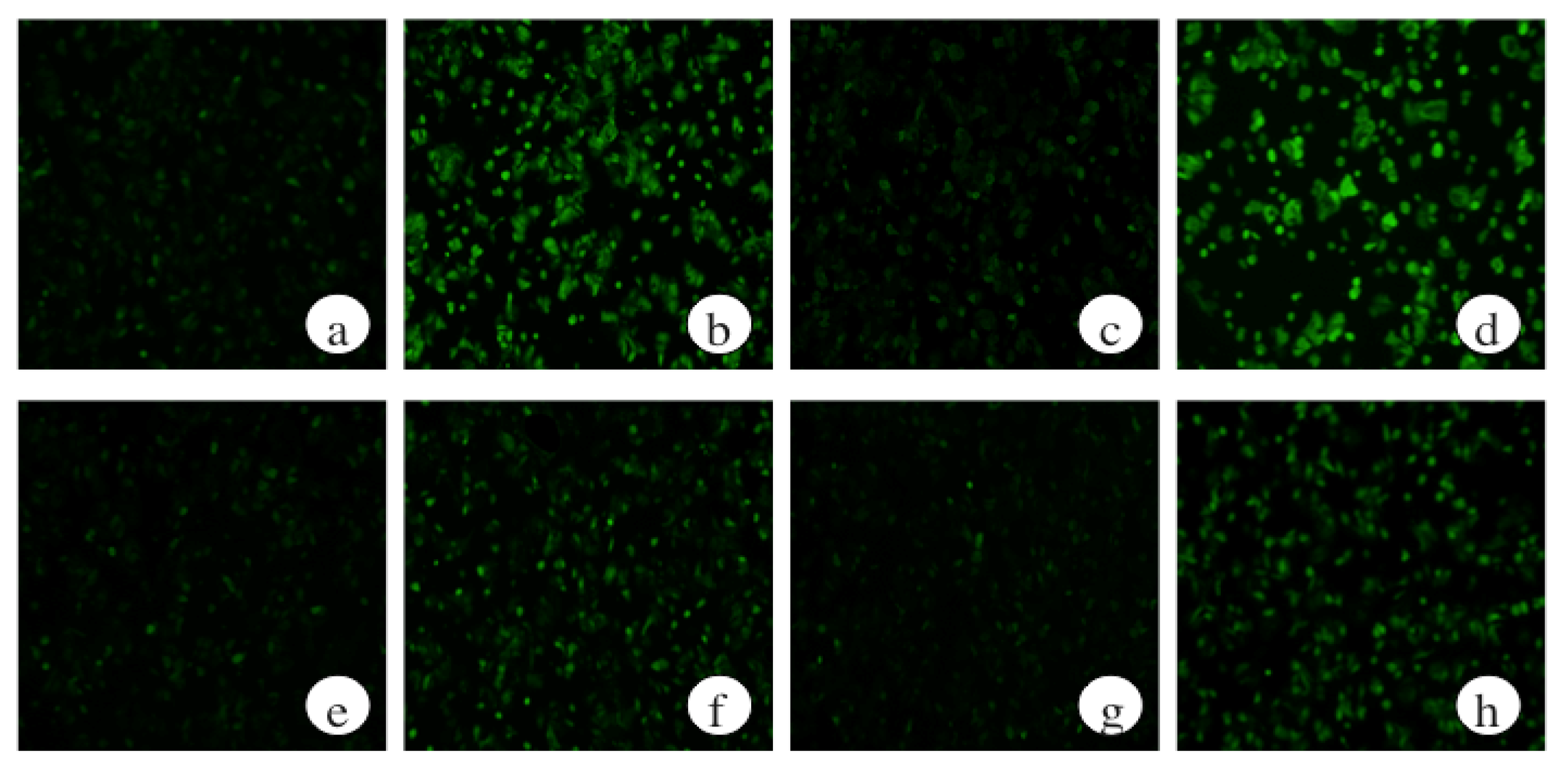
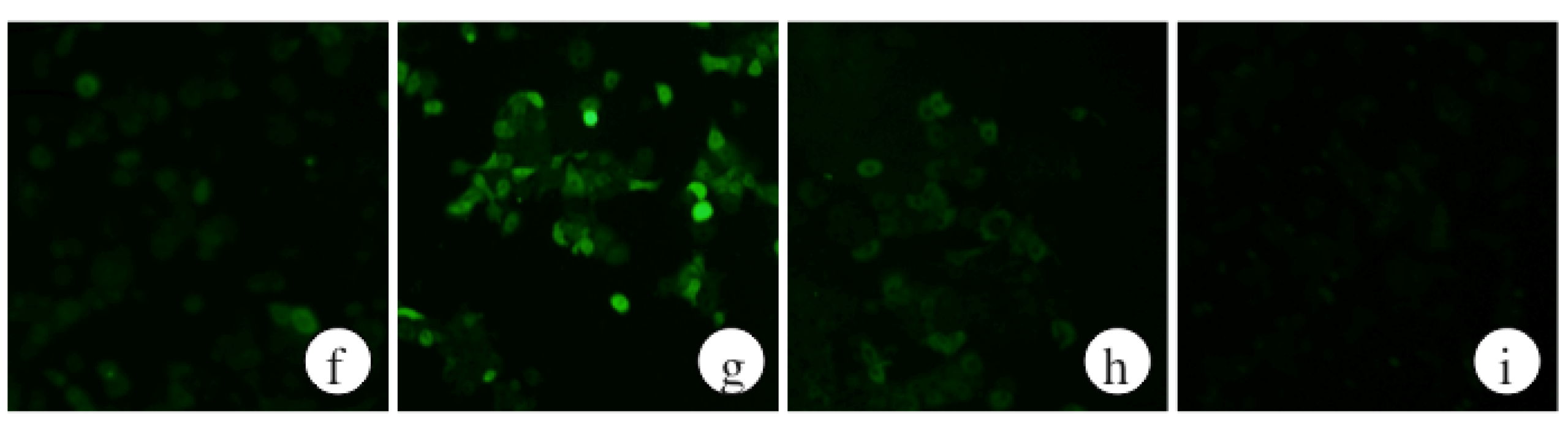
2.7. The Effects of a Protein Kinase A Inhibitor on the ROS Levels of Ethanol-Treated Hepatocytes
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Preparation of ECP
4.3. Animals
4.4. Hepatocyte Preparation and Culture
4.5. Cell Viability Assay
4.6. Western Blot Analysis of CYP2E1
4.7. Intracellular ROS Formation
4.8. CYP2E1 Activity Assay
4.9. Assay of ADH and ALDH Activities
4.10. Assay of Intracellular cAMP Levels
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lieber, C.S. Ethanol metabolism, cirrhosis and alcoholism. Clin. Chim. Acta 1997, 257, 59–84. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Lu, S.C. Current concepts in the pathogenesis of alcoholic liver injury. FASEB J. 2001, 15, 1335–1349. [Google Scholar] [CrossRef] [PubMed]
- Lucey, M.R.; Mathurin, P.; Morgan, T.R. Alcoholic hepatitis. N. Engl. J. Med. 2009, 360, 2758–2769. [Google Scholar] [CrossRef] [PubMed]
- Zima, T.; Fialová, L.; Mestek, O.; Janebová, M.; Crkovská, J.; Malbohan, I.; Stípek, S.; Mikulíková, L.; Popov, P. Oxidative stress, metabolism of ethanol and alcohol-related disease. J. Biomed. Sci. 2001, 8, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Albano, E. Alcohol, oxidative stress and free radical damage. Proc. Nutr. Soc. 2006, 65, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Regan, M.A.; Glombitza, K.W. Phlorotannins, brown algae polyphenols. In Progress Phycological Research; Round, F.E., Chapman, D.J., Eds.; Biopress Ltd.: Bristol, UK, 1986; Volume 4, pp. 129–252. [Google Scholar]
- Ahn, G.-N.; Kim, K.-N.; Cha, S.-H.; Song, C.-B.; Lee, J.; Heo, M.-S.; Yeo, I.-K.; Lee, N.-H.; Jee, Y.-H.; Kim, J.-S.; et al. Antioxidant activities of phlorotannins purified from Ecklonia cava on free radical scavenging using ESR and H2O2-mediated DNA damage. Eur. Food Res. Technol. 2007, 226, 71–76. [Google Scholar] [CrossRef]
- Kang, H.S.; Chung, H.Y.; Kim, J.Y.; Son, B.W.; Jung, H.A.; Choi, J.S. Inhibitory phlorotannins from the edible brown alga Ecklonia stolonifera on total reactive oxygen speacies (ROS) generation. Arch. Pharm. Res. 2004, 27, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, Y.; Kodama, M.; Miura, I.; Kinzyo, Z.; Kido, M.; Mori, H.; Nakayama, Y.; Takahashi, M. Structure of an anti-plasmin inhibitor, eckol, isolated from the brown alga Ecklonia kurome Okamura and inhibitory activities of its derivatives on plasmin inhibitors. Chem. Pharm. Bull. 1989, 37, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, N.D.; Choi, J.S.; Kim, Y.J.; Moon, Y.H.; Lim, S.Y.; Park, K.Y. Inhibitory effects of the methanolic extract of an edible brown alga, Ecklonia stolonifera and its component, phlorglucinol on aflatoxin B1 mutagenicity in vitro (Ames test) and on benzo(a)pyrene or N-methyl N-nitrosourea clastogenicity in vivo (mouse micronucleus test). Nat. Prod. Sci. 1998, 4, 105–114. [Google Scholar]
- Nagayama, K.; Iwamura, Y.; Shibata, T.; Hirayama, I.; Nakamura, T. Bactericidal activity of phlorotannins from the brown alga Ecklonia kurome. J. Antimicrob. Chemother. 2002, 50, 889–893. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.J.; Yoon, K.D.; Min, S.Y.; Lee, J.S.; Kim, J.H.; Kim, T.G.; Kim, N.G.; Huh, H.; Kim, J. Inhibition of HIV-1 reverse transcriptase and proteases by phlorotannins from brown alga Eclonia cava. Biol. Pharm. Bull. 2004, 27, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Park, Y.; Hwang, H.J.; Kim, S.H.; Lee, J.G.; Shin, H.C. Antioxidative properties of brown algae polyphenols and their perspectives as chemopreventive agents against vascular risk factors. Arch. Pharm. Res. 2003, 26, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Yokogawa, K.; Matsui-Yuasa, I.; Tamura, A.; Terada, M.; Kojima-Yuasa, A. Inhibitory effect of Ecklonia cava extract on high glucose-induced hepatic stellate cell activation. Mar. Drugs 2011, 9, 2793–2808. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.-J.; Park, E.-J.; Lee, K.-W.; Jeon, Y.-J. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol. 2005, 96, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Satake, N.; Yamashita, H.; Tamura, A.; Sasaki, M.; Matsui-Yuasa, I.; Tabuchi, M.; Akahishi, Y.; Terada, M.; Kojima-Yuasa, A. Ecklonia cava polyphenol protects the liver against ethanol-induced injury in rats. Biochim. Biophys. Acta 2012, 1820, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S.; DeCarli, L.M. Ethanol oxidation by hepatic microsomes—Adaptive increase after ethanol feeding. Science 1968, 162, 912–918. [Google Scholar] [CrossRef]
- Lieber, C.S. Metabolic effect of acetaldehyde. Biochem. Soc. Trans. 1988, 16, 241–247. [Google Scholar] [PubMed]
- Potter, J.J.; MacDougald, O.A.; Mezey, E. Regulation of rat alcohol dehydrogenase by cyclic AMP in primary hepatocyte culture. Arch. Biochem. Biophys. 1995, 321, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Oesch-Bartlomowicz, B.; Padma, P.R.; Becker, R.; Richter, B.; Hengstler, J.G.; Freeman, J.E.; Wolf, C.R.; Oesch, F. Differential modulation of CYP2E1 activity by cAMP-dependent protein kinase upon Ser129 replacement. Exp. Cell Res. 1998, 242, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Hock, J.B.; Pastorino, J.G. Ethanol, oxidative stress, and cytokine-induced liver cell injury. Alcohol 2002, 27, 63–68. [Google Scholar] [CrossRef]
- Johansson, I.; Eliasson, E.; Ingelman-Sundberg, M. Hormone controlled phosphorylation and degradation of CYP2B1 and CYP2E1 in isolated rat hepatocytes. Biochem. Biophys. Res. Commun. 1991, 174, 37–42. [Google Scholar] [CrossRef]
- Cederbaum, A.I.; Wu, D.; Mari, M.; Bal, J. CYP2E1-dependent toxicity and oxidative stress in HepG2 cells. Free Radic. Biol. Med. 2001, 31, 1539–1543. [Google Scholar] [CrossRef]
- Kessova, I.; Cederbaum, A.I. CYP2E1: Biochemistry, toxicology, regulation and function in ethanol-induced liver injury. Curr. Mol. Med. 2003, 3, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.; Leclercq, I.; Farrell, G.C. Nonalcholic steatosis and sreahepatitis. II. Cytochrome P-450 enzymes and oxidative stress. Am. J. Physiol. 2001, 281, G1135–G1139. [Google Scholar]
- Lu, Y.; Wu, D.; Wang, X.; Ward, S.C.; Cederbaum, A.I. Chronic alcohol-induced liver injury and oxidant stress is decreased in cytochrome P4502E1 knockout mice and restored in humanized cytochrome P4502E1 knockin mice. Free Radic. Biol. Med. 2010, 49, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.Q.; Wang, P.Y.; Wang, Y.; Kaneko, T.; Hoshi, K.; Sato, A. Voglibose potentiates the hepatotoxicity of carbon tetrachloride and acetaminophen by inducing CYP2E1 in rats. Hepatol. Res. 2005, 33, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Jia, Q.; Wang, Y.; Zhang, Y.; Xia, M. The anthocyanin cyaniding-3-O-β-glucoside, a flavonoid, increases hepatic glutathione synthesis and protects hepatocytes against reactive oxygen species during hyperglycemia: Involvement of a cAMP-PKA-dependent signaling pathway. Free Radic. Biol. Med. 2012, 52, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Ok, W.J.; Cho, H.J.; Kim, H.H.; Lee, D.H.; Kang, H.Y.; Kwon, H.W.; Rhee, M.H.; Kim, M.; Park, H.J. Epigallocatechin-3-Gallate Has an Anti-Platelet Effect in a Cyclic AMP-Dependent Manner. J. Atheroscler. Thromb. 2012, 19, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.E.; DeSilva, S.E.F. Ethanol increases receptor-dependent cyclic AMP production in cultured hepatocytes by decreasing Gi-mediated inhibition. Biochem. J. 1992, 286, 681–686. [Google Scholar] [PubMed]
- Moldéus, P.; Högber, J.; Orrenius, S. Isolation and use of liver cells. Methods Enzymol. 1978, 52, 60–71. [Google Scholar] [PubMed]
- Zhang, S.Z.; Lipsky, M.M.; Trump, B.F.; Hsu, I.C. Nuetral red (NR) assay for cell viability and xenobiotic-induced cytotoxicity in primary culture of human and rat hepatocytes. Cell Biol. Toxicol. 1990, 6, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Sasaki, M.; Yamashita, H.; Matsui-Yuasa, I.; Saku, T.; Hikima, T.; Tabuchi, M.; Munakata, H.; Kojima-Yuasa, A. Yerba-mate (Ilex paraguariensis) extract prevents ethanol-induced liver injury in rats. J. Funct. Foods 2013, 5, 1714–1723. [Google Scholar] [CrossRef]
- Sapone, A.; Affatato, A.; Canistro, D.; Broccoli, M.; Trespidi, S.; Pozzetti, L.; Biagi, G.L.; Cantelli-Forti, G.; Paolini, M. Induction and suppression of cytochrome P450 isoenzymes and generation of oxygen radicals by procymidone in liver, kidney, and lung of CD1 mice. Mutat. Res. 2003, 527, 67–80. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamashita, H.; Goto, M.; Matsui-Yuasa, I.; Kojima-Yuasa, A. Ecklonia cava Polyphenol Has a Protective Effect against Ethanol-Induced Liver Injury in a Cyclic AMP-Dependent Manner. Mar. Drugs 2015, 13, 3877-3891. https://doi.org/10.3390/md13063877
Yamashita H, Goto M, Matsui-Yuasa I, Kojima-Yuasa A. Ecklonia cava Polyphenol Has a Protective Effect against Ethanol-Induced Liver Injury in a Cyclic AMP-Dependent Manner. Marine Drugs. 2015; 13(6):3877-3891. https://doi.org/10.3390/md13063877
Chicago/Turabian StyleYamashita, Haruka, Mayu Goto, Isao Matsui-Yuasa, and Akiko Kojima-Yuasa. 2015. "Ecklonia cava Polyphenol Has a Protective Effect against Ethanol-Induced Liver Injury in a Cyclic AMP-Dependent Manner" Marine Drugs 13, no. 6: 3877-3891. https://doi.org/10.3390/md13063877
APA StyleYamashita, H., Goto, M., Matsui-Yuasa, I., & Kojima-Yuasa, A. (2015). Ecklonia cava Polyphenol Has a Protective Effect against Ethanol-Induced Liver Injury in a Cyclic AMP-Dependent Manner. Marine Drugs, 13(6), 3877-3891. https://doi.org/10.3390/md13063877