Marine Compound Catunaregin Inhibits Angiogenesis through the Modulation of Phosphorylation of Akt and eNOS in vivo and in vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
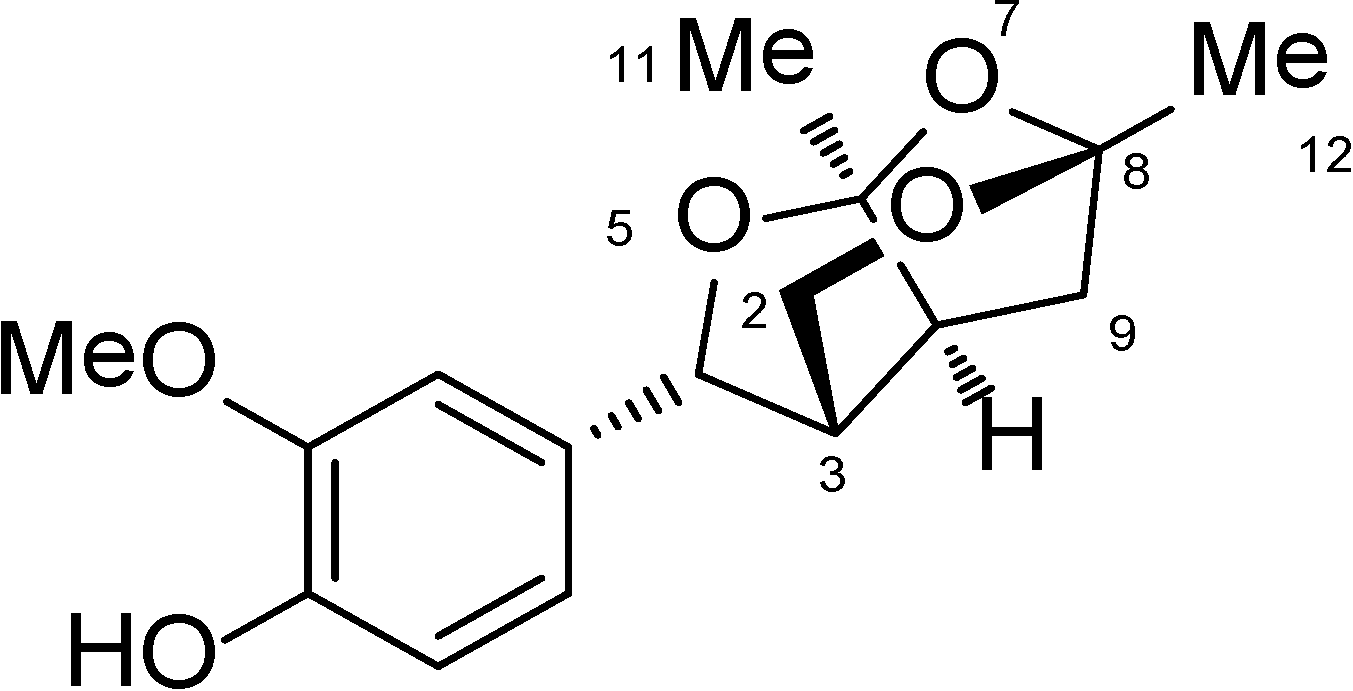
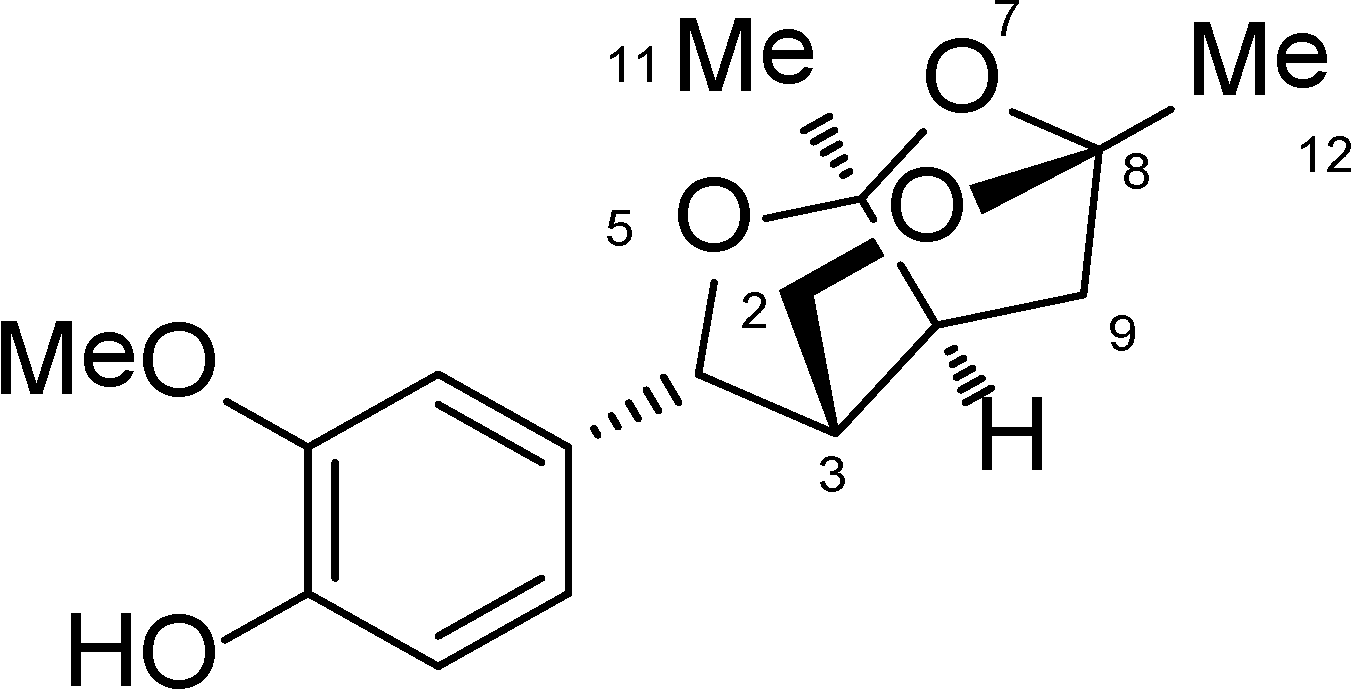
2.1. The Isolation and Preparation of Catunaregin
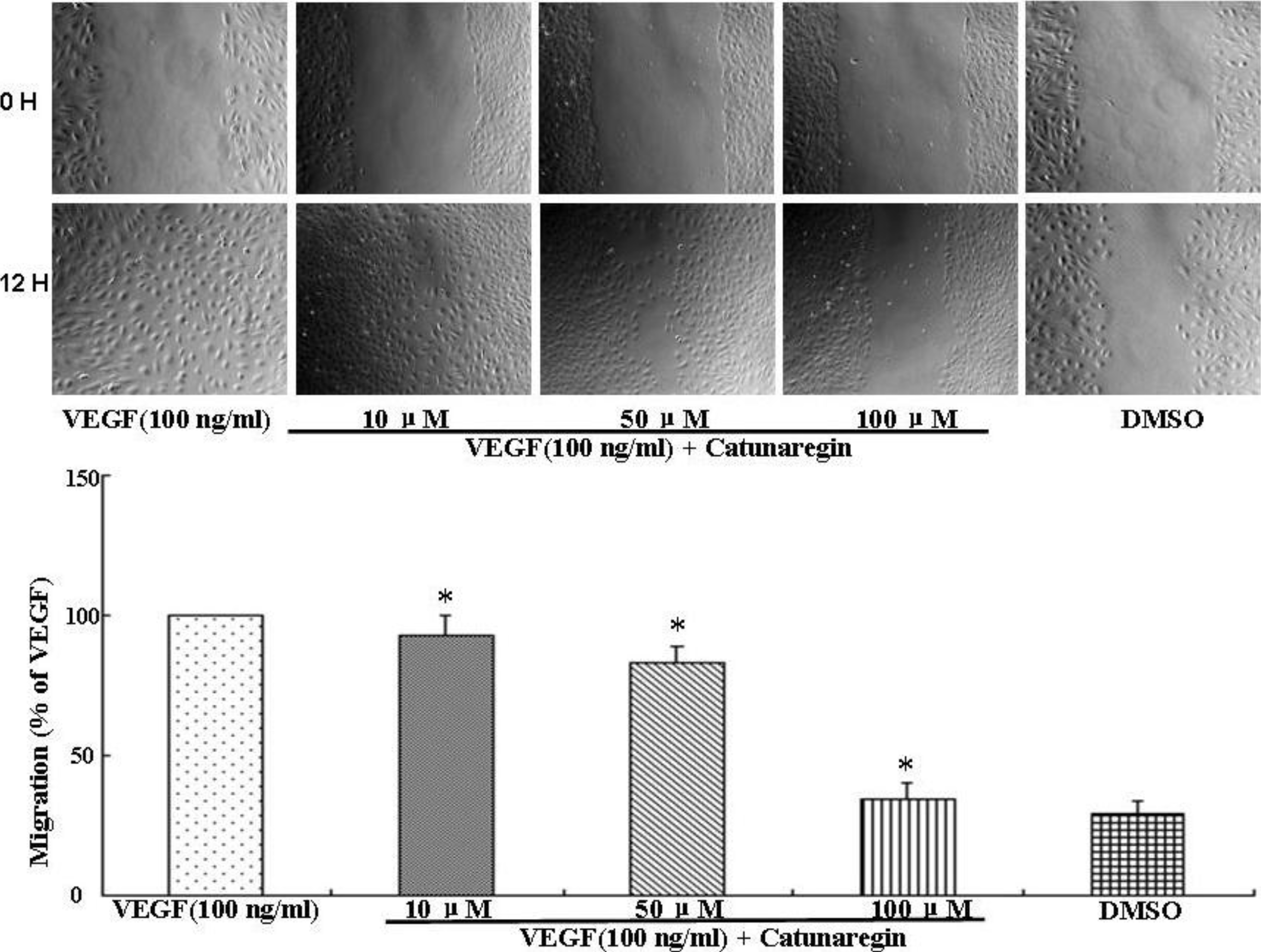
2.2. Inhibitory Effect of Catunaregin on VEGF-induced HUVEC Invasion, Migration, and Tube Formation
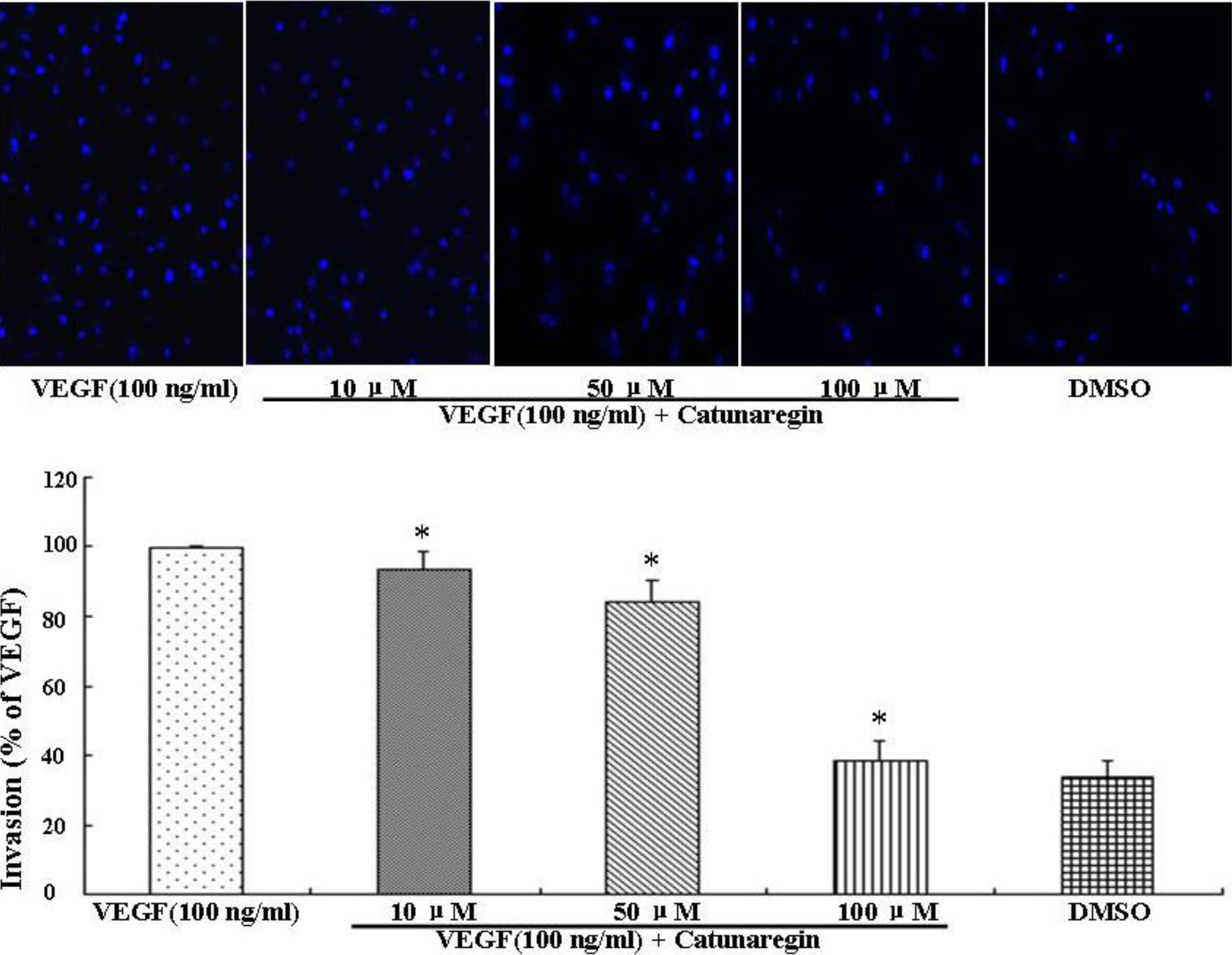
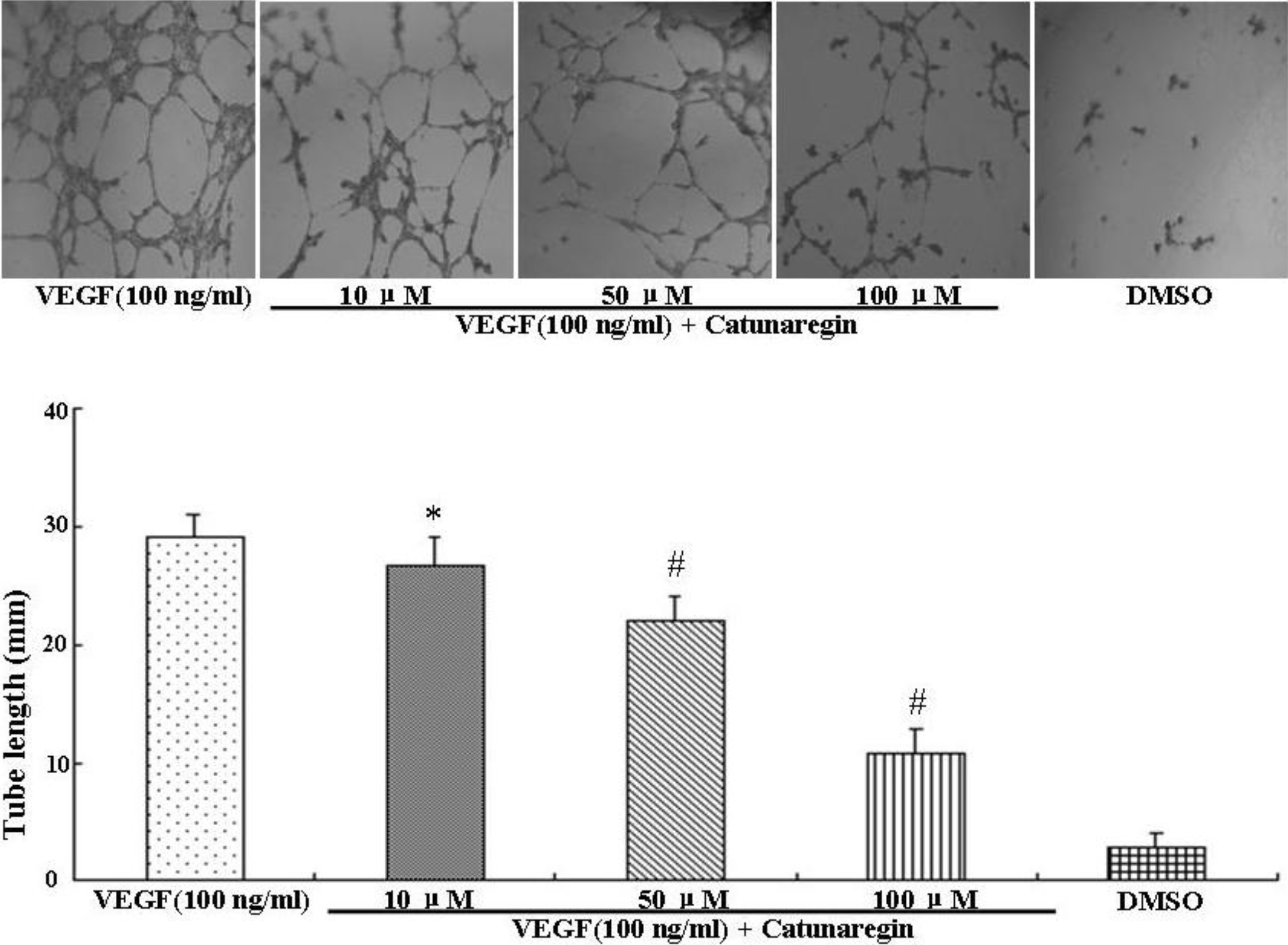
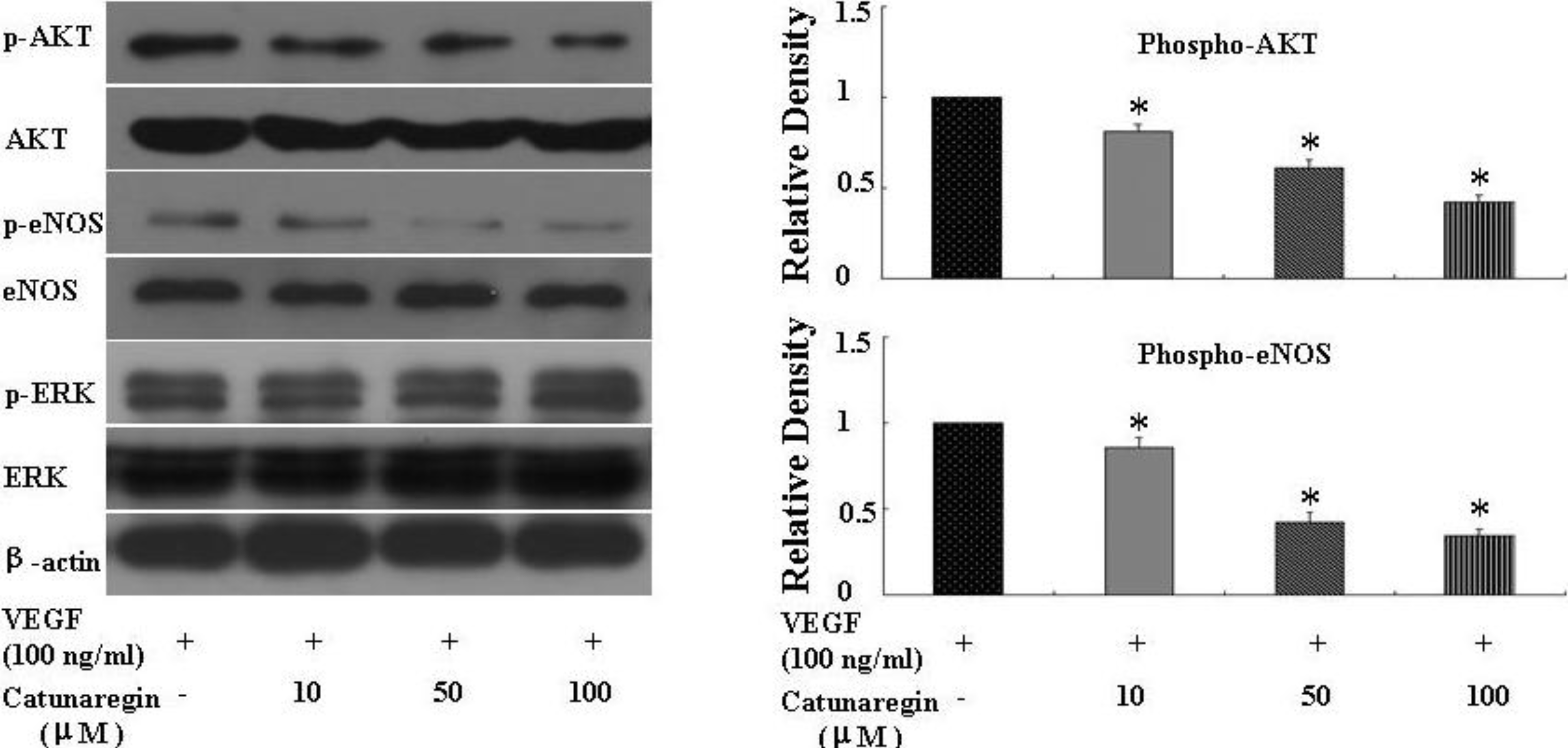
2.3. Catunaregin Inhibited Angiogenesis through Modulation of AKT and eNOS Signaling Pathways
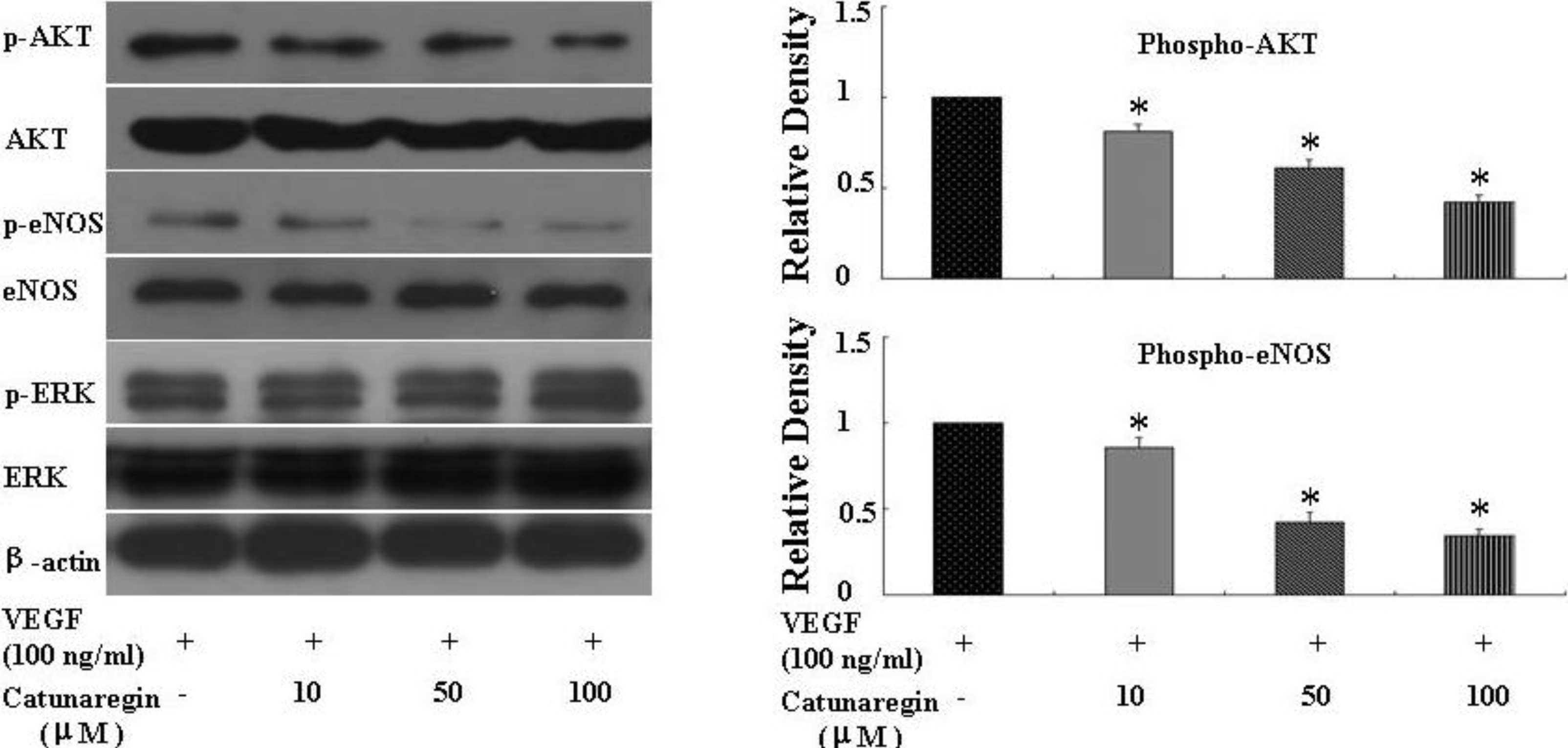
2.4. Catunaregin Inhibited Angiogenesis in Zebrafish Embryo and Caudal Fin Regeneration Assays
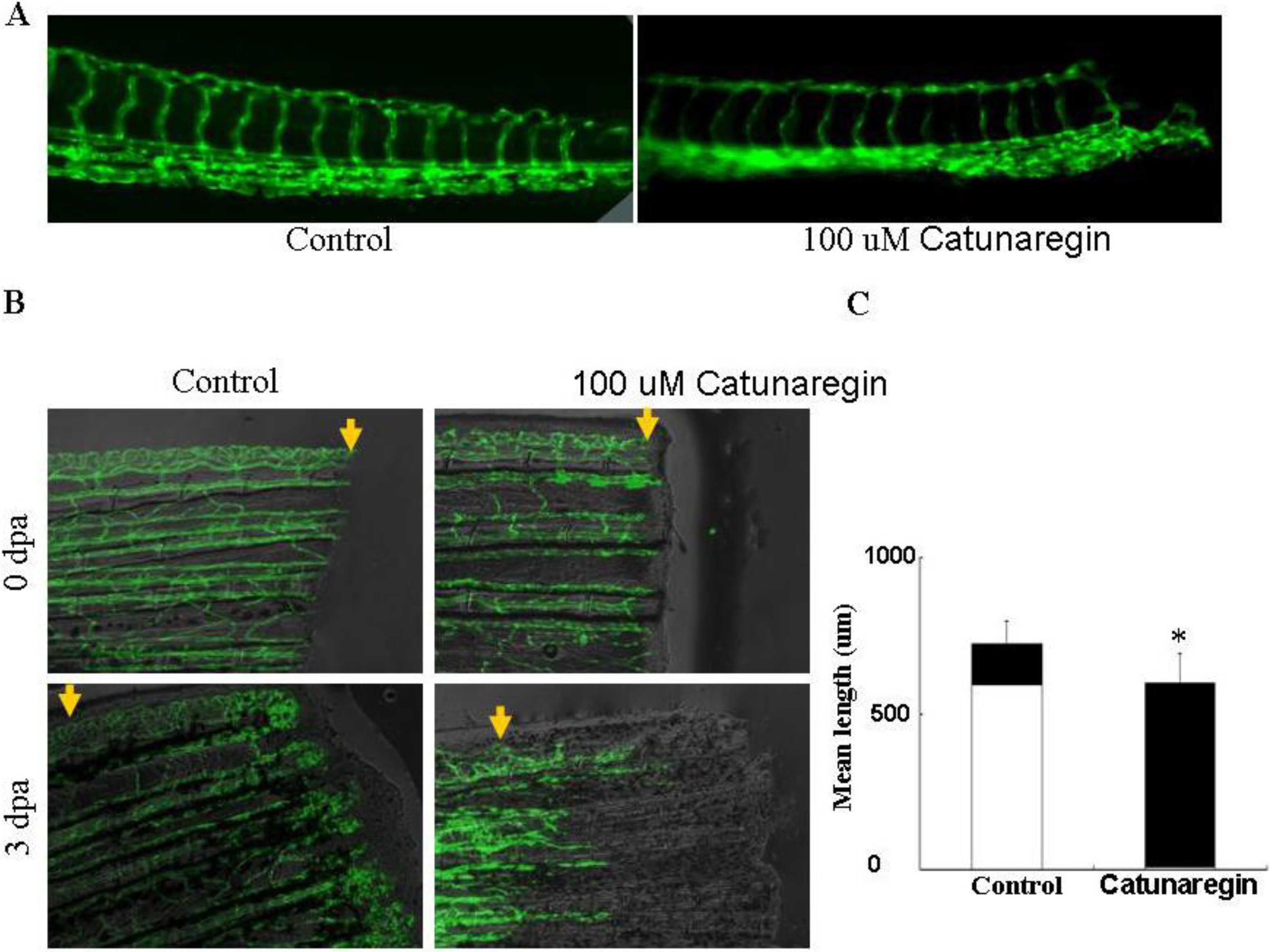
3. Experimental Section
3.1. Preparation of Catunaregin
3.2. The Isolation of Catunaregin
3.3. Preparation of Reagents and Cell Culture
3.4. Cell Invasion, Migration and Tube Formation
3.5. Western Immunoblot Analysis
3.6. Zebrafish Embryo Assay
3.7. Zebrafish Caudal Fin Regeneration Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Munoz-Chapuli, R.; Quesada, A.; Angel Medina, M. Angiogenesis and signal transduction in endothelial cells. Cell. Mol. Life. Sci. 2004, 61, 2224–2243. [Google Scholar]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Eng. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef]
- Jeong, S.; Koh, W.; Lee, E.; Lee, H.; Bae, H.; Lü, J.; Kim, S. Antiangiogenic phytochemicals and medicinal herbs. Phytother. Res. 2011, 25, 1–10. [Google Scholar] [CrossRef]
- Gao, G.; Luo, X.; Wei, X.; Qi, S.; Yin, H.; Xiao, Z.; Zhang, S. Catunaregin and epicatunaregin, two norneolignans possessing an unprecedented skeleton from Catunaregam spinosa. Helv. Chim. Acta. 2010, 93, 339–344. [Google Scholar] [CrossRef]
- Lu, X.; Xu, Z.; Yao, X.; Su, F.; Ye, C.; Li, J.; Lin, Y.; Wang, G.; Zeng, J.; Huang, R.; et al. Marine cyclotripeptide X-13 promotes angiogenesis in zebrafish and human endothelial cells via PI3K/Akt/eNOS signaling pathways. Mar. Drugs. 2012, 10, 1307–1320. [Google Scholar] [CrossRef]
- Bayliss, P.; Bellavance, K.; Whitehead, G.; Abrams, J.; Aegerter, S.; Robbins, H.; Cowan, D.; Keating, M.; O’Reilly, T.; Wood, J.; et al. Chemical modulation of receptor signaling inhibits regenerative angiogenesis in adult zebrafish. Nat. Chem. Biol. 2006, 2, 265–273. [Google Scholar]
- Lu, X.; Luo, D.; Yao, X.; Wang, G.; Liu, Z.; Li, Z.; Li, W.; Chang, F.; Wen, L.; Lee, S.; et al. dl-3n-butylphthalide promotes angiogenesis via the extracellular signal-regulated kinase 1/2 and phosphatidylinositol 3-kinase/Akt-endothelial nitric oxide synthase signaling pathways. J. Cardiovasc. Pharmacol. 2012, 59, 352–362. [Google Scholar] [CrossRef]
- Lam, H.W.; Lin, H.; Lao, S.; Gao, J.; Hong, S.; Leong, C.; Yue, P.; Kwan, Y.; Leung, A.; Wang, Y.; et al. The angiogenic effects of angelica sinensis extract on HUVEC in vitro and zebrafish in vivo. J. Cell. Biochem. 2008, 103, 195–211. [Google Scholar] [CrossRef]
- Raghunath, M.; Wong, Y.; Muhamma, F.; Ge, R. Pharmacologically induced angiogenesis in transgenic zebrafish. Biochem. Biophys. Res. Commun. 2009, 378, 766–771. [Google Scholar] [CrossRef]
- Abdel-Malak, N.; Mofarrahi, M.; Mayaki, D.; Khachigian, L.; Hussain, S. Early growth response-1 regulates angiopoietin-1-induced endothelial cell proliferation, migration, and differentiation. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 209–216. [Google Scholar] [CrossRef]
- Lam, K.H.; Alex, D.; Lam, I.K.; Tsui, S.K.; Yang, Z.F.; Lee, S.M. Nobiletin, a polymethoxylated flavonoid from citrus, shows anti-angiogenic activity in a zebrafish in vivo model and HUVEC in vitro model. J. Cell. Biochem. 2011, 112, 3313–3321. [Google Scholar] [CrossRef]
- Hong, S.; Wan, J.; Zhang, Y.; Hu, G.; Lin, H.; Seto, S.; Kwan, Y.; Lin, Z.; Wang, Y.; Lee, S. Angiogenic effect of saponin extract from Panax notoginseng on HUVECs in vitro and zebrafish in vivo. Phytother. Res. 2009, 23, 677–686. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, J.-X.; Luo, M.-Q.; Xia, M.; Wu, Q.; Long, S.-M.; Hu, Y.; Gao, G.-C.; Yao, X.-L.; He, M.; Su, H.; et al. Marine Compound Catunaregin Inhibits Angiogenesis through the Modulation of Phosphorylation of Akt and eNOS in vivo and in vitro. Mar. Drugs 2014, 12, 2790-2801. https://doi.org/10.3390/md12052790
Liu J-X, Luo M-Q, Xia M, Wu Q, Long S-M, Hu Y, Gao G-C, Yao X-L, He M, Su H, et al. Marine Compound Catunaregin Inhibits Angiogenesis through the Modulation of Phosphorylation of Akt and eNOS in vivo and in vitro. Marine Drugs. 2014; 12(5):2790-2801. https://doi.org/10.3390/md12052790
Chicago/Turabian StyleLiu, Jun-Xiu, Min-Qi Luo, Meng Xia, Qi Wu, Si-Mei Long, Yaohua Hu, Guang-Chun Gao, Xiao-Li Yao, Mian He, Huanxing Su, and et al. 2014. "Marine Compound Catunaregin Inhibits Angiogenesis through the Modulation of Phosphorylation of Akt and eNOS in vivo and in vitro" Marine Drugs 12, no. 5: 2790-2801. https://doi.org/10.3390/md12052790
APA StyleLiu, J.-X., Luo, M.-Q., Xia, M., Wu, Q., Long, S.-M., Hu, Y., Gao, G.-C., Yao, X.-L., He, M., Su, H., Luo, X.-M., & Yao, S.-Z. (2014). Marine Compound Catunaregin Inhibits Angiogenesis through the Modulation of Phosphorylation of Akt and eNOS in vivo and in vitro. Marine Drugs, 12(5), 2790-2801. https://doi.org/10.3390/md12052790