
_Kim.png)
The Isoforms of Ral Guanine Nucleotide Dissociation Stimulator (RalGDS) in LLC-PK1 Cells

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. RT-PCR, DNA Cloning, and Sequence Assembly
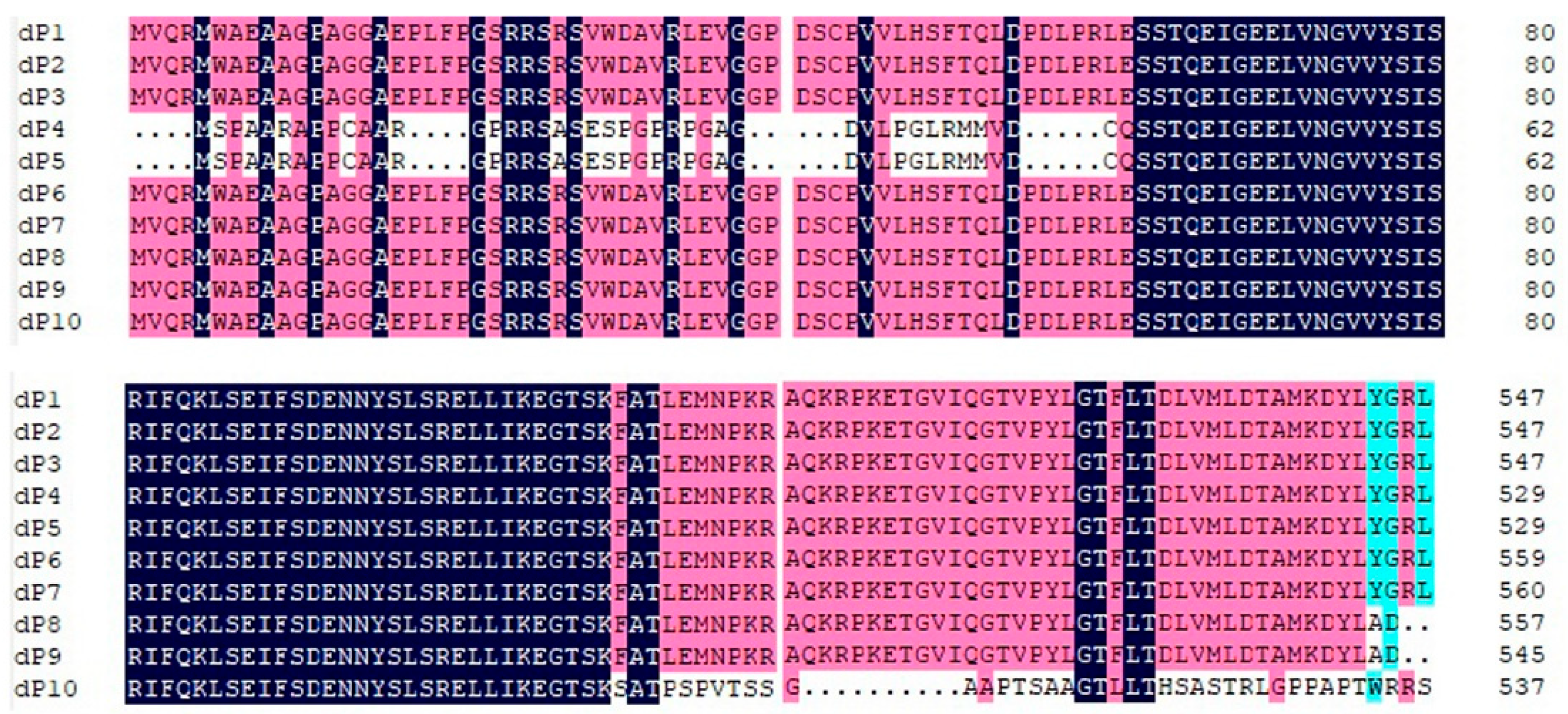
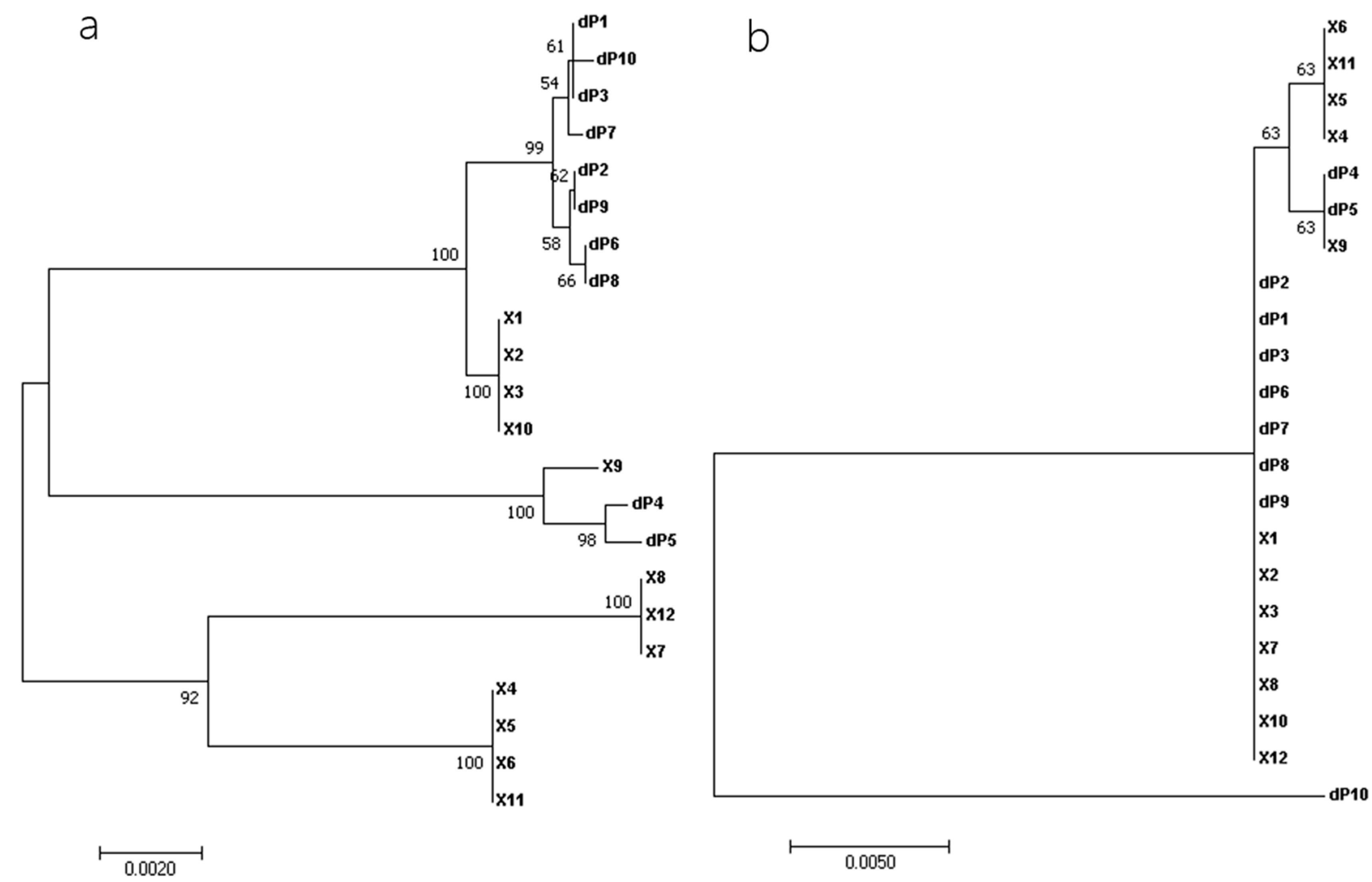
2.3. Multiple-Sequence Alignments and Phylogenetic Analyses
2.4. Physicochemical Analysis of RalGDS Proteins and Prediction of Global N6-Methyladenosine (m6A) Sites in RalGDS mRNA
3. Results
3.1. Complete Genomic Characterization of RalGDS
3.2. Phylogenetic Analysis
3.3. Physicochemical Properties, Secondary Structure Analysis, and m6A Sites’ Prediction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferro, E.; Trabalzini, L. RalGDS family members couple Ras to Ral signalling and that’s not all. Cell Signal. 2010, 22, 1804–1810. [Google Scholar] [CrossRef]
- Yoshizawa, R.; Umeki, N.; Yanagawa, M.; Murata, M.; Sako, Y. Single-molecule fluorescence imaging of RalGDS on cell surfaces during signal transduction from Ras to Ral. Biophys. Physicobiol. 2017, 14, 75–84. [Google Scholar] [CrossRef]
- Albright, C.F.; Giddings, B.W.; Liu, J.; Vito, M.; Weinberg, R.A. Characterization of a guanine nucleotide dissociation stimulator for a ras-related GTPase. EMBO J. 1993, 12, 339–347. [Google Scholar] [CrossRef]
- Luck, K.; Kim, D.K.; Lambourne, L.; Spirohn, K.; Begg, B.E.; Bian, W.; Brignall, R.; Cafarelli, T.; Campos-Laborie, F.J.; Charloteaux, B.; et al. A reference map of the human binary protein interactome. Nature 2020, 580, 402–408. [Google Scholar] [CrossRef]
- Zimin, A.V.; Delcher, A.L.; Florea, L.; Kelley, D.R.; Schatz, M.C.; Puiu, D.; Hanrahan, F.; Pertea, G.; Van Tassell, C.P.; Sonstegard, T.S.; et al. A whole-genome assembly of the domestic cow, Bos taurus. Genome Biol. 2009, 10, R42. [Google Scholar] [CrossRef] [PubMed]
- Postlethwait, J.H.; Farnsworth, D.R.; Miller, A.C. An intestinal cell type in zebrafish is the nexus for the SARS-CoV-2 receptor and the Renin-Angiotensin-Aldosterone System that contributes to COVID-19 comorbidities. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zheng, H.; Zhang, W.; Zhang, L.; Zhang, Z.; Li, J.; Lu, G.; Zhu, Y.; Wang, Y.; Huang, Y.; Liu, J.; et al. The genome of the hydatid tapeworm Echinococcus granulosus. Nat. Genet. 2013, 45, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Omholt, K.; Hansson, J. No evidence of RALGDS mutations in cutaneous melanoma. Melanoma Res. 2007, 17, 410–412. [Google Scholar] [CrossRef]
- Darlington, T.M.; Ehringer, M.A.; Larson, C.; Phang, T.L.; Radcliffe, R.A. Transcriptome analysis of Inbred Long Sleep and Inbred Short Sleep mice. Genes Brain Behav. 2013, 12, 263–274. [Google Scholar] [CrossRef]
- Naushad, S.M.; Hussain, T.; Al-Attas, O.S.; Prayaga, A.; Digumarti, R.R.; Gottumukkala, S.R.; Kutala, V.K. Molecular insights into the association of obesity with breast cancer risk: Relevance to xenobiotic metabolism and CpG island methylation of tumor suppressor genes. Mol. Cell. Biochem. 2014, 392, 273–280. [Google Scholar] [CrossRef]
- Mach, N.; Gao, Y.; Lemonnier, G.; Lecardonnel, J.; Oswald, I.P.; Estellé, J.; Rogel-Gaillard, C. The peripheral blood transcriptome reflects variations in immunity traits in swine: Towards the identification of biomarkers. BMC Genom. 2013, 14, 894. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Zhu, J.; Zhang, F.; Xiao, Q.; Xie, J.; He, K. Interaction of porcine circovirus-like virus P1 capsid protein with host proteins. BMC Vet. Res. 2021, 17, 227. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wen, L.; Sheng, S.; Wang, W.; Xiao, Q.; Qu, M.; Hu, Y.; Liu, C.; He, K. Porcine circovirus-like virus P1 inhibits Wnt signaling pathway in vivo and in vitro. Front. Microbiol. 2018, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xiao, Q.; Yin, L.; Zhang, F.; Wen, L.; Suolang, S.; He, K. Porcine circovirus-like virus P1 activates pancreatic secretion pathway by interacting with CHRM3 protein. Vet. Microbiol. 2022, 272, 109495. [Google Scholar] [CrossRef]
- Wen, L.; He, K. Genomic rearrangement and recombination of porcine circovirus type 2 and porcine circovirus-like virus P1 in China. Front. Vet. Sci. 2021, 8, 736366. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Yang, L.; Liu, N.; Yang, J.; Zhou, X.K.; Xia, Y.C.; He, Y.; He, Y.Q.; Gong, H.J.; Ma, D.F.; et al. Genome-wide identification, structure characterization, and expression pattern profiling of aquaporin gene family in cucumber. BMC Plant Biol. 2019, 19, 345. [Google Scholar] [CrossRef]
- González-García, A.; Pritchard, C.A.; Paterson, H.F.; Mavria, G.; Stamp, G.; Marshall, C.J. RalGDS is required for tumor formation in a model of skin carcinogenesis. Cancer Cell 2005, 7, 219–226. [Google Scholar] [CrossRef]
- Chan, K.C.; Lai, P.B.; Mok, T.S.; Chan, H.L.; Ding, C.; Yeung, S.W.; Lo, Y.M. Quantitative analysis of circulating methylated DNA as a biomarker for hepatocellular carcinoma. Clin. Chem. 2008, 54, 1528–1536. [Google Scholar] [CrossRef]
- Buhmeida, A.; Merdad, A.; Al-Maghrabi, J.; Al-Thobaiti, F.; Ata, M.; Bugis, A.; Syrjänen, K.; Abuzenadah, A.; Chaudhary, A.; Gari, M.; et al. RASSF1A methylation is predictive of poor prognosis in female breast cancer in a background of overall low methylation frequency. Anticancer Res. 2011, 31, 2975–2981. [Google Scholar]
- Miranda, E.; Bianchi, P.; Destro, A.; Morenghi, E.; Malesci, A.; Santoro, A.; Laghi, L.; Roncalli, M. Genetic and epigenetic alterations in primary colorectal cancers and related lymph node and liver metastases. Cancer 2013, 119, 266–276. [Google Scholar] [CrossRef]
- Liu, F.; Du, J.; Liu, J.; Wen, B. Identification of key target genes and pathways in laryngeal carcinoma. Oncol. Lett. 2016, 12, 1279–1286. [Google Scholar] [CrossRef]
- Kawai, M.; Kawashima, S.; Sakoda, T.; Toh, R.; Kikuchi, A.; Yamauchi-Takihara, K.; Kunisada, K.; Yokoyama, M. Ral GDP dissociation stimulator and Ral GTPase are involved in myocardial hypertrophy. Hypertension 2003, 41, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Scotland, R.L.; Allen, L.; Hennings, L.J.; Post, G.R.; Post, S.R. The ral exchange factor rgl2 promotes cardiomyocyte survival and inhibits cardiac fibrosis. PLoS ONE 2013, 8, e73599. [Google Scholar] [CrossRef]
- Rifki, O.F.; Bodemann, B.O.; Battiprolu, P.K.; White, M.A.; Hill, J.A. RalGDS-dependent cardiomyocyte autophagy is required for load-induced ventricular hypertrophy. J. Mol. Cell. Cardiol. 2013, 59, 128–138. [Google Scholar] [CrossRef]
- He, P.C.; He, C. m6A RNA methylation: From mechanisms to therapeutic potential. EMBO J. 2021, 40, e105977. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequences | Target Genes | PCR Product Size |
|---|---|---|---|
| 1 | F26-CCGTCCATGGTGCAGCGCATGTG; R721-GGTCGGAGCCGGGCATGTTGAGC | X1, X2, X3, X10 | 814 |
| 2 | F82-GCGTTCATGCTGGTGGTCC; R742-CTAGCTCTGGGGCTGGTTTC | X7, X8, X12 | 661 |
| 3 | F4341-ACCGGAAGAGGATGTGCAAC; R5220-CACTGGACTGGGTGCTAGAAG | X4, X5, X6, X11 | 880 |
| 4 | F234-CGGCTCAAACAATGGGATCAG; R1027-TCTAGCTCTGGGGCTGGTTT | X9 | 794 |
| 5 | F244-GAGTCGGCCCTGAACCTGTATGA; R1517-GGTACCCTGGATGACACCCGTCT | All | 1274 |
| 6 | F979-GGAGCTGGCTCTGTCGCAAAG; R2334-CGGATGATGCAGCAGTCGC | All | 1356 |
| 7 | F1833-TGGAGTGGCTCAGCGAGACTG; R3287-TGAGCGTGGTCTGCGAAGAG | All | 1455 |
| 8 | F136-CCGGTGGTGCTGCACAGCTTCA; R2065-TCTCCTGGCCATCGGGGGACTC | X1, X2, X3, X10 | 1951 |
| 9 | F193-TCCACACAGGAGATTGGCGAGG; R2429-CAGCTTGCGATCTTCCGAAATG | X9 | 2258 |
| Reference Gene (Length) | dP1 | dP2 | dP3 | dP4 | dP5 | dP6 | dP7 | dP8 | dP9 | dP10 |
|---|---|---|---|---|---|---|---|---|---|---|
| X1 (3554 bp) | 90.35 | 90.35 | 90.35 | 87.75 | 87.75 | 91.33 | 91.42 | 91.22 | 90.24 | 78.33 |
| X2 (3551 bp) | 90.43 | 90.43 | 90.43 | 87.83 | 87.83 | 91.41 | 91.33 | 91.30 | 90.31 | 78.40 |
| X3 (3514 bp) | 91.38 | 91.38 | 91.38 | 88.70 | 88.70 | 90.42 | 90.35 | 90.31 | 91.26 | 79.23 |
| X4 (7900 bp) | 39.36 | 39.36 | 39.36 | 39.42 | 39.42 | 39.80 | 39.84 | 39.75 | 39.31 | 33.95 |
| X5 (7897 bp) | 39.37 | 39.37 | 39.37 | 39.44 | 39.44 | 39.82 | 39.80 | 39.76 | 39.32 | 33.97 |
| X6 (7484 bp) | 41.54 | 41.54 | 41.54 | 41.61 | 41.61 | 41.33 | 41.32 | 41.28 | 41.49 | 35.84 |
| X7 (3460 bp) | 87.73 | 87.73 | 87.73 | 86.68 | 86.68 | 88.73 | 88.65 | 88.61 | 87.62 | 75.64 |
| X8 (3424 bp) | 88.64 | 88.64 | 88.64 | 87.57 | 87.57 | 87.71 | 87.63 | 87.59 | 88.52 | 76.42 |
| X9 (3744 bp) | 83.00 | 83.00 | 83.00 | 86.28 | 86.28 | 82.18 | 82.12 | 82.07 | 82.89 | 71.64 |
| X10 (3477 bp) | 88.08 | 88.08 | 88.08 | 85.56 | 85.56 | 89.06 | 89.15 | 88.95 | 87.96 | 77.65 |
| X11 (7783 bp) | 38.59 | 38.59 | 38.59 | 38.61 | 38.61 | 38.41 | 38.39 | 38.35 | 38.54 | 33.47 |
| X12 (3345 bp) | 86.38 | 86.38 | 86.38 | 85.32 | 85.32 | 85.47 | 85.39 | 85.35 | 86.26 | 75.84 |
| X1 (885 aa) | 98.53 | 98.53 | 98.53 | 92.54 | 92.54 | 99.89 | 100.00 | 62.71 | 61.24 | 56.95 |
| X2 (884 aa) | 98.64 | 98.64 | 98.64 | 93.21 | 93.21 | 100.00 | 99.77 | 62.78 | 61.43 | 57.01 |
| X3 (872 aa) | 100.00 | 100.00 | 100.00 | 94.50 | 94.50 | 98.64 | 98.42 | 61.43 | 62.27 | 57.80 |
| X4 (858 aa) | 92.09 | 92.09 | 92.09 | 94.46 | 94.46 | 94.01 | 93.67 | 56.84 | 54.92 | 51.07 |
| X5 (857 aa) | 92.76 | 92.76 | 92.76 | 94.57 | 94.57 | 94.12 | 93.45 | 56.90 | 55.09 | 51.13 |
| X6 (845 aa) | 94.04 | 94.04 | 94.04 | 95.90 | 95.90 | 92.76 | 92.09 | 55.54 | 55.85 | 51.83 |
| X7 (835 aa) | 91.97 | 91.97 | 91.97 | 93.88 | 93.88 | 93.33 | 93.11 | 56.11 | 54.75 | 50.34 |
| X8 (823 aa) | 93.23 | 93.23 | 93.23 | 95.20 | 95.20 | 91.97 | 91.75 | 54.75 | 55.50 | 51.03 |
| X9 (817 aa) | 93.00 | 93.00 | 93.00 | 95.67 | 95.67 | 91.74 | 91.53 | 54.52 | 55.28 | 50.80 |
| X10 (762 aa) | 78.85 | 78.85 | 78.85 | 74.12 | 74.12 | 80.20 | 80.43 | 72.74 | 71.04 | 66.06 |
| X11 (722 aa) | 73.74 | 73.74 | 73.74 | 76.23 | 76.23 | 72.75 | 72.55 | 64.44 | 64.93 | 60.27 |
| X12 (700 aa) | 73.97 | 73.97 | 73.97 | 75.53 | 75.53 | 72.41 | 72.22 | 63.52 | 64.53 | 59.33 |
| RalGDS Strain | Protein Length (AA) | MW(Da) | pI | Instability Index | Aliphatic Index | GRAVY | Secondary Structure Prediction (SOPMA) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Alpha Helix | Extended Strand | Beta Turn | Random Coil | |||||||
| dP1, dP2, dP3 | 872 | 96030.83 | 5.71 | 51.91 | 82.24 | −0.338 | 38.30 | 11.70 | 4.47 | 45.53 |
| dP4, dP5 | 854 | 93789.38 | 5.96 | 53.19 | 80.90 | −0.351 | 37.47 | 12.06 | 3.86 | 46.60 |
| dP6 | 884 | 97312.24 | 5.77 | 51.82 | 81.79 | −0.340 | 38.57 | 12.10 | 4.75 | 44.57 |
| dP7 | 885 | 97399.31 | 5.77 | 52.21 | 81.69 | −0.341 | 38.31 | 12.09 | 4.63 | 44.97 |
| dP8 | 557 | 61323.00 | 5.35 | 45.95 | 88.82 | −0.226 | 48.65 | 11.13 | 5.39 | 34.83 |
| dP9 | 545 | 60041.59 | 5.29 | 45.98 | 89.69 | −0.219 | 48.44 | 10.46 | 4.95 | 36.15 |
| dP10 | 538 | 58657.73 | 5.50 | 49.70 | 87.43 | −0.225 | 45.72 | 9.85 | 4.46 | 39.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Li, N.; Dong, X.; Xie, J.; Lai, H.; Zhu, H.; He, K.; Wen, L.; Suolang, S.; Xiao, Q. The Isoforms of Ral Guanine Nucleotide Dissociation Stimulator (RalGDS) in LLC-PK1 Cells. Curr. Issues Mol. Biol. 2025, 47, 566. https://doi.org/10.3390/cimb47070566
Song J, Li N, Dong X, Xie J, Lai H, Zhu H, He K, Wen L, Suolang S, Xiao Q. The Isoforms of Ral Guanine Nucleotide Dissociation Stimulator (RalGDS) in LLC-PK1 Cells. Current Issues in Molecular Biology. 2025; 47(7):566. https://doi.org/10.3390/cimb47070566
Chicago/Turabian StyleSong, Jingze, Na Li, Xinze Dong, Jianping Xie, Hongqiao Lai, Hengzhi Zhu, Kongwang He, Libin Wen, Sizhu Suolang, and Qi Xiao. 2025. "The Isoforms of Ral Guanine Nucleotide Dissociation Stimulator (RalGDS) in LLC-PK1 Cells" Current Issues in Molecular Biology 47, no. 7: 566. https://doi.org/10.3390/cimb47070566
APA StyleSong, J., Li, N., Dong, X., Xie, J., Lai, H., Zhu, H., He, K., Wen, L., Suolang, S., & Xiao, Q. (2025). The Isoforms of Ral Guanine Nucleotide Dissociation Stimulator (RalGDS) in LLC-PK1 Cells. Current Issues in Molecular Biology, 47(7), 566. https://doi.org/10.3390/cimb47070566