
Biological Characteristics and Genomic Analysis of Acinetobacter nosocomialis Lytic Phage XC1

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Strain and Sewage
2.1.2. Reagents and Instruments
2.2. Methods
2.2.1. Preliminary Isolation and Acquisition of XC1 Phage
2.2.2. Isolation and Purification of Phage and Determination of Titer
2.2.3. Determination of Optimal Phage Infection Multiplicity (MOI)
2.2.4. Determination of Phage One-Step Growth Curve
2.2.5. Determination of Phage Thermal Stability and pH Stability
- (1)
- Thermal Stability of Phage
- (2)
- pH Stability of Phage
2.2.6. Observation of Phage Morphology by Transmission Electron Microscopy
2.2.7. DNA Extraction and Whole-Genome Sequencing Analysis of XC1 Phage
2.2.8. XC1 Phage Gene Function Prediction and Annotation
2.2.9. Analysis of XC1 Phage Evolutionary Relationships
2.2.10. Comparative Genomics Analysis of the XC1 Phage
3. Results
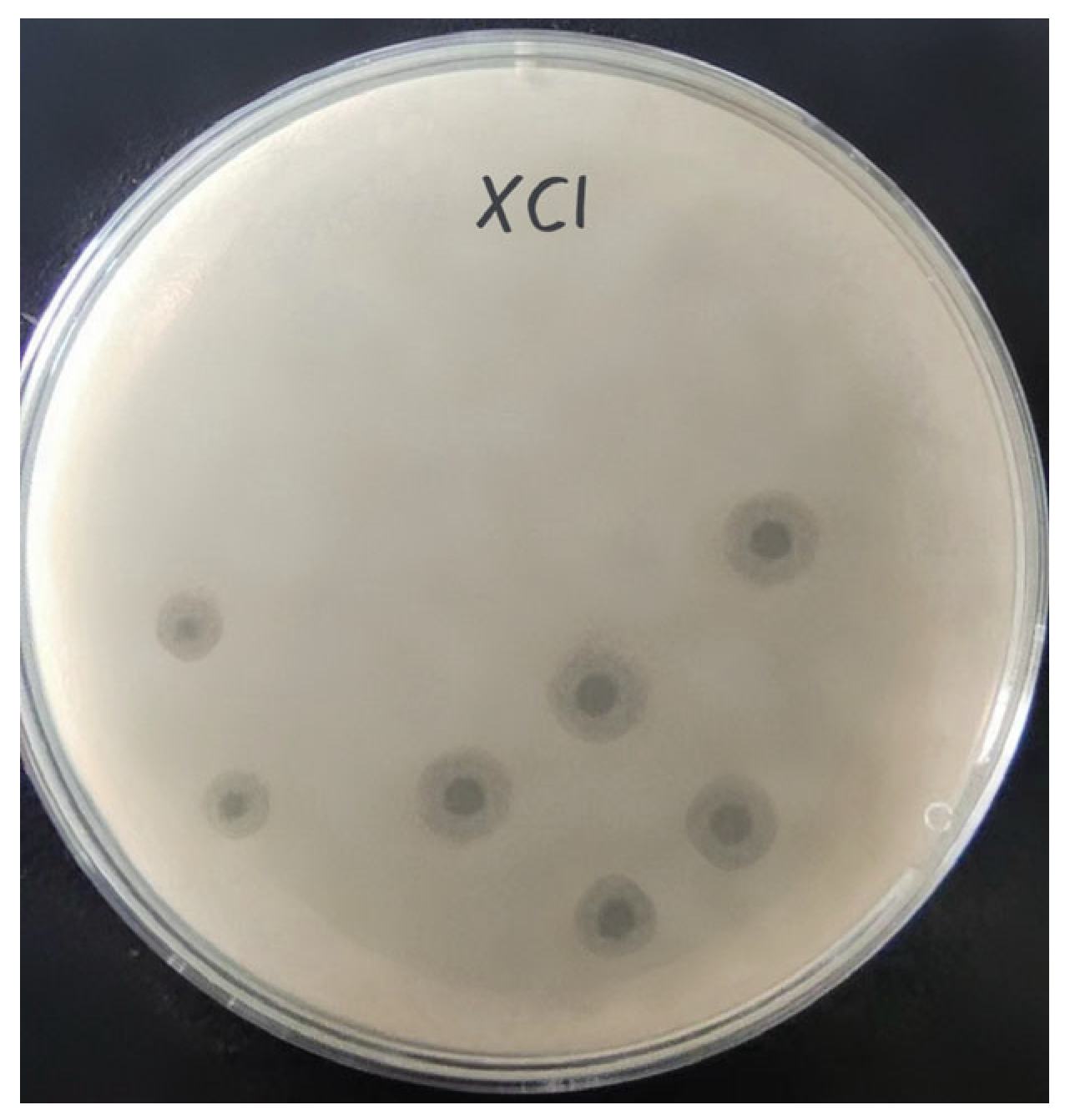
3.1. Isolation and Morphological Characterization of Phage XC1
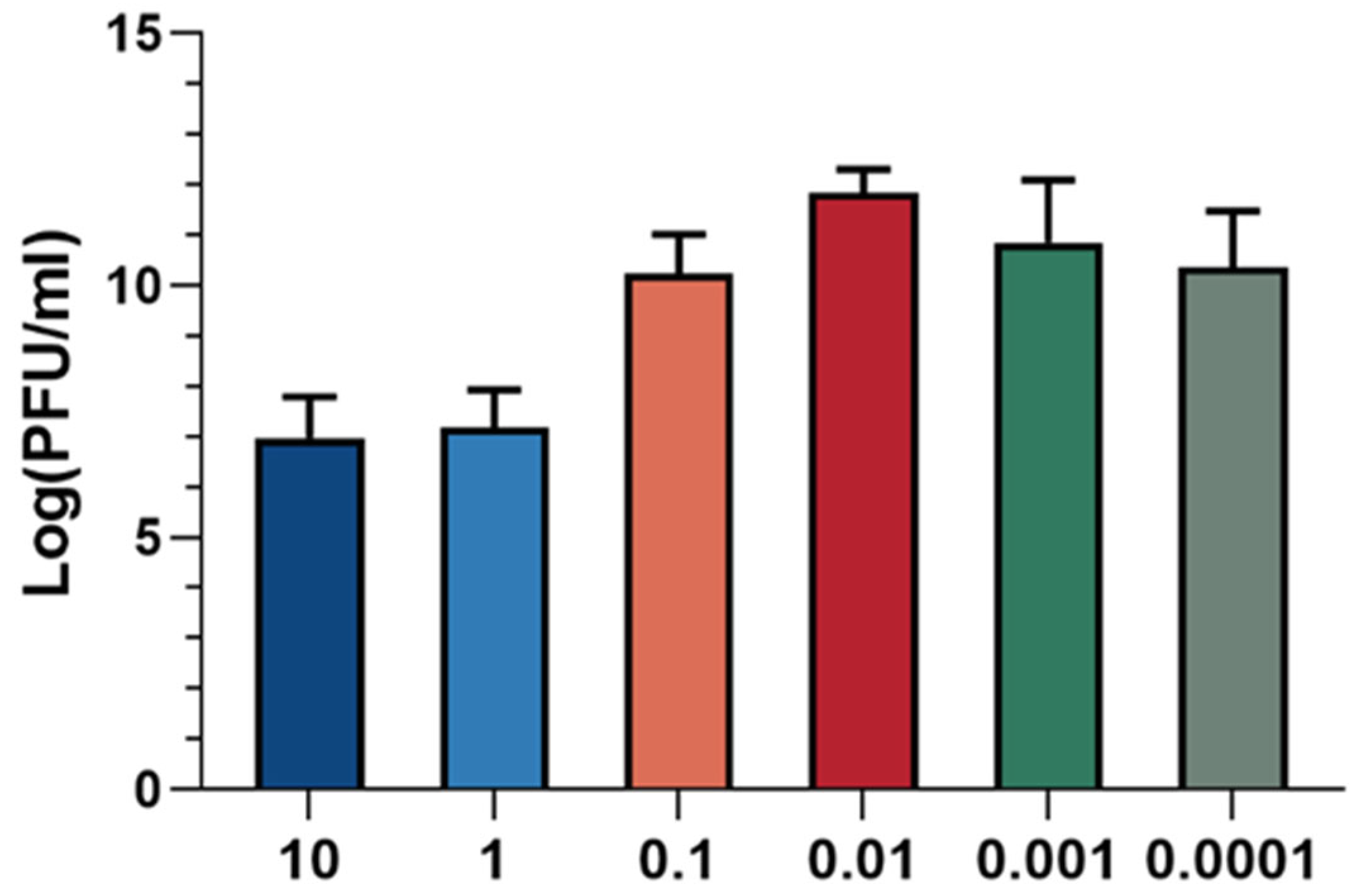
3.2. Optimal Multiplicity of Infection (MOI) Assay for Phage XC1
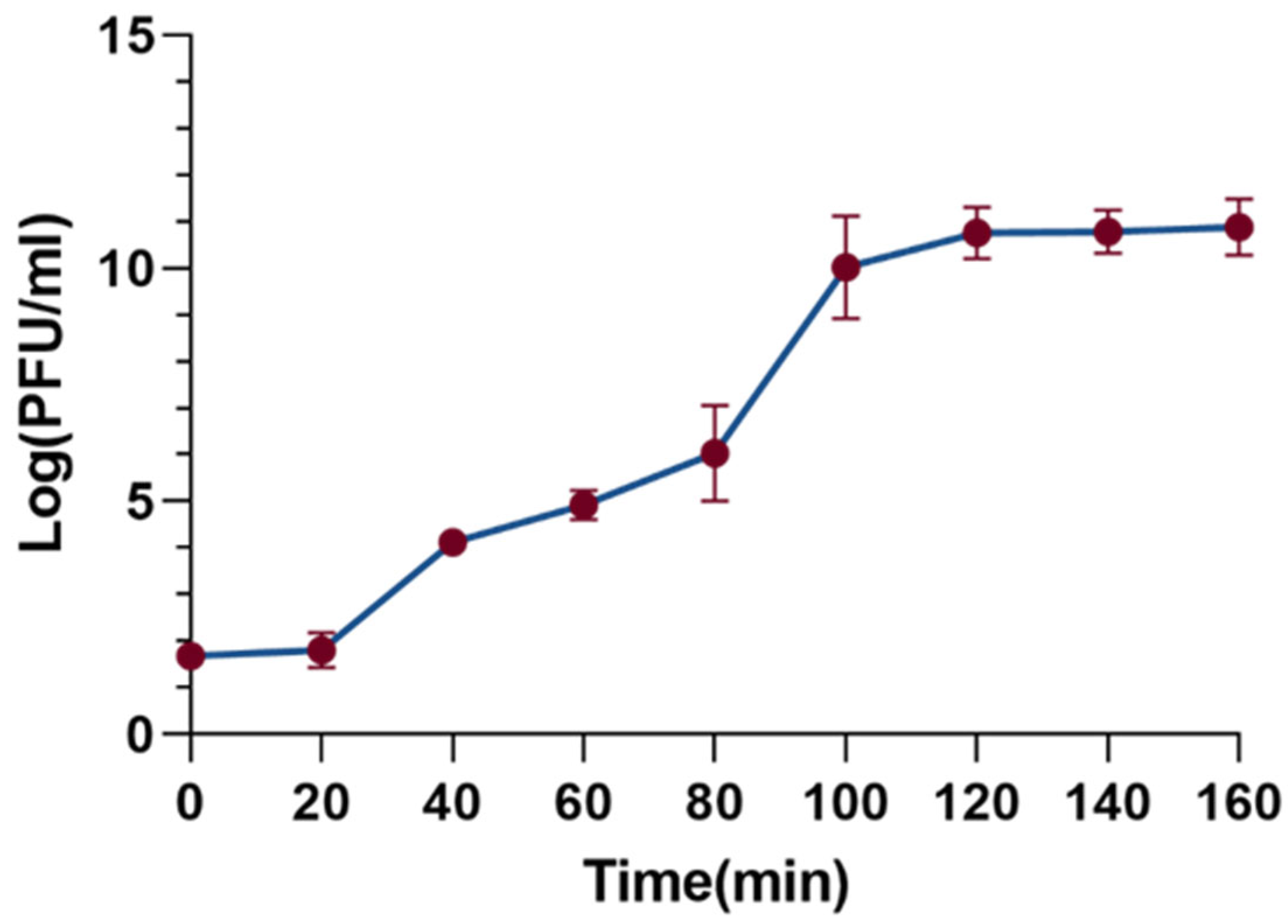
3.3. One-Step Growth Curve of Phage XC1
3.4. Determination of Phage Stability
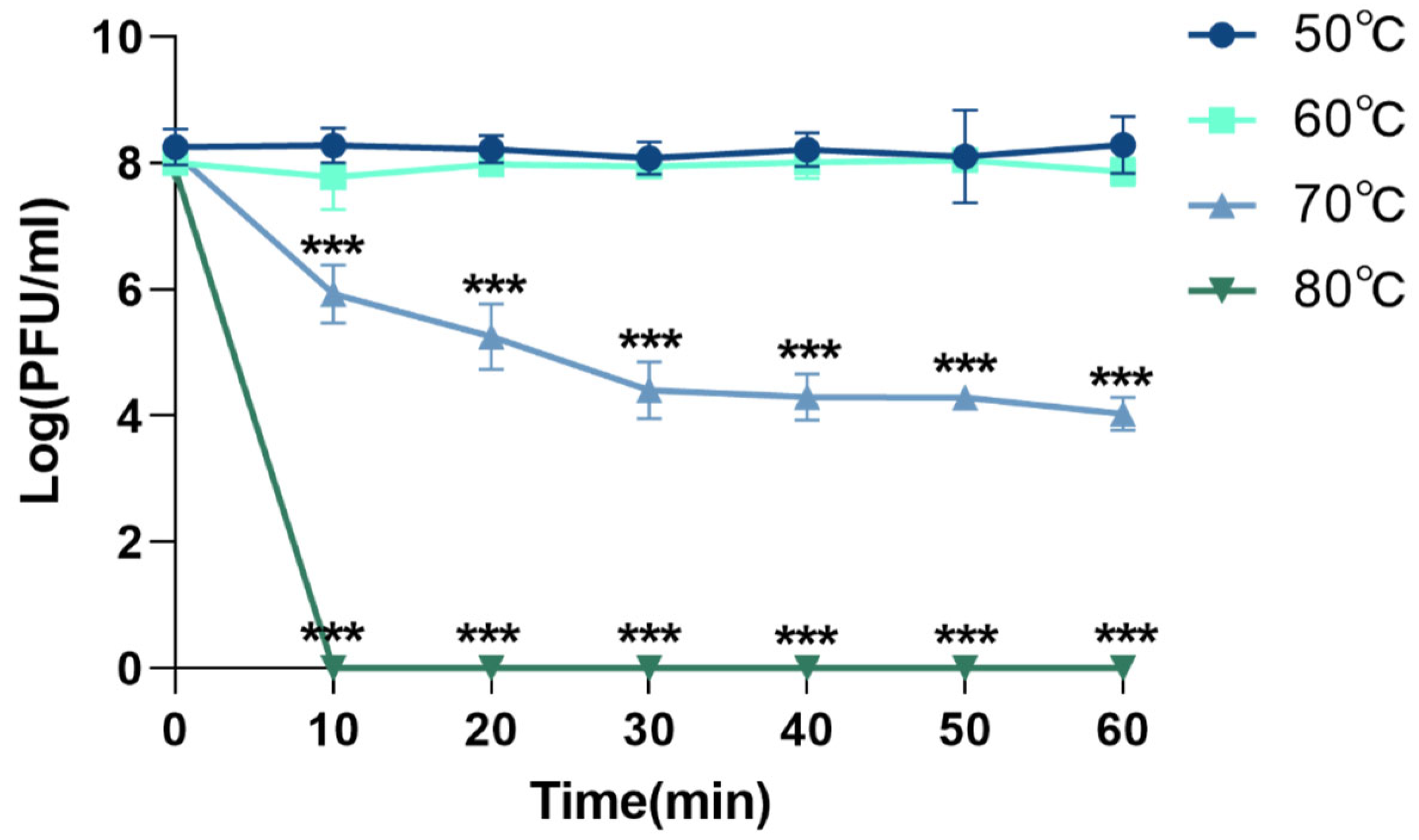
3.4.1. Thermal Stability of Phage XC1
3.4.2. pH Stability of Phage XC1
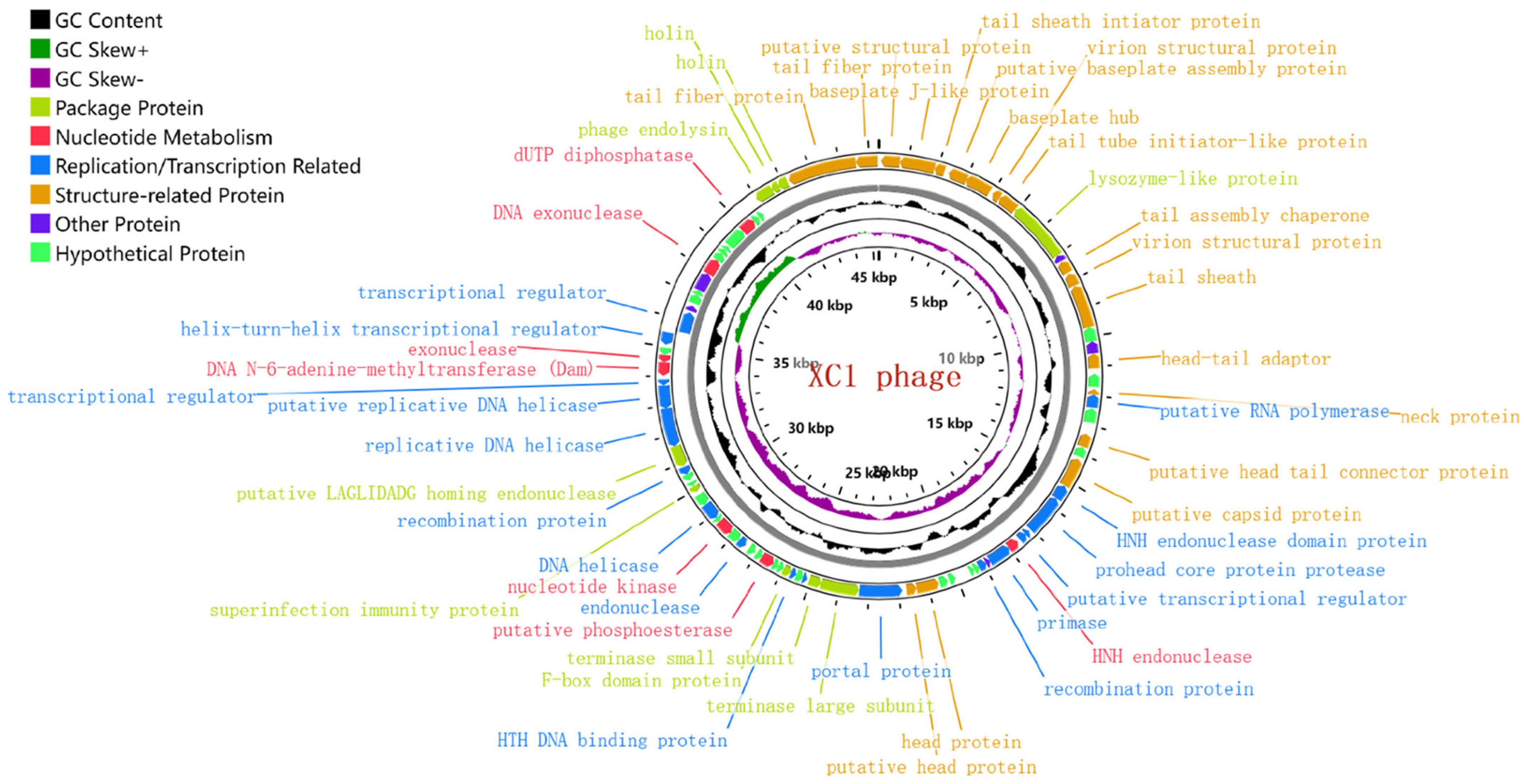
3.5. Genome-Wide Analysis and Functional Annotation of Phage XC1
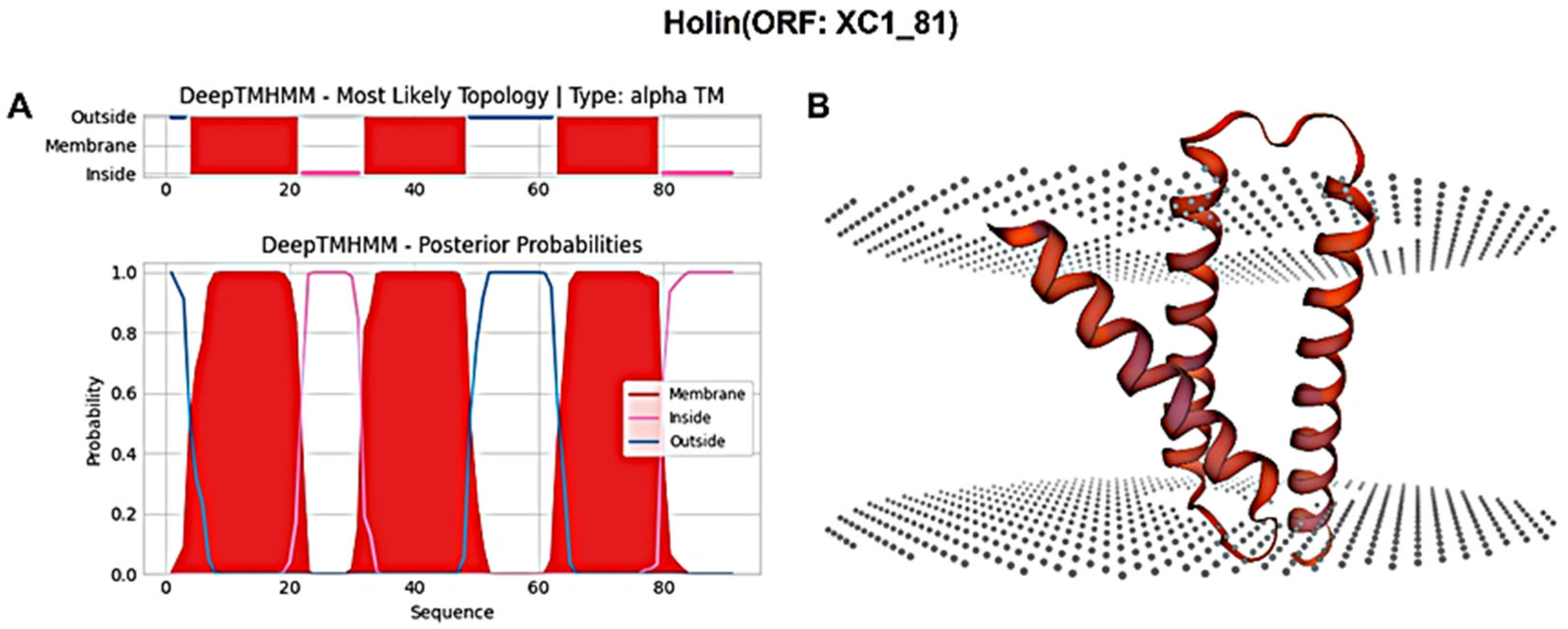
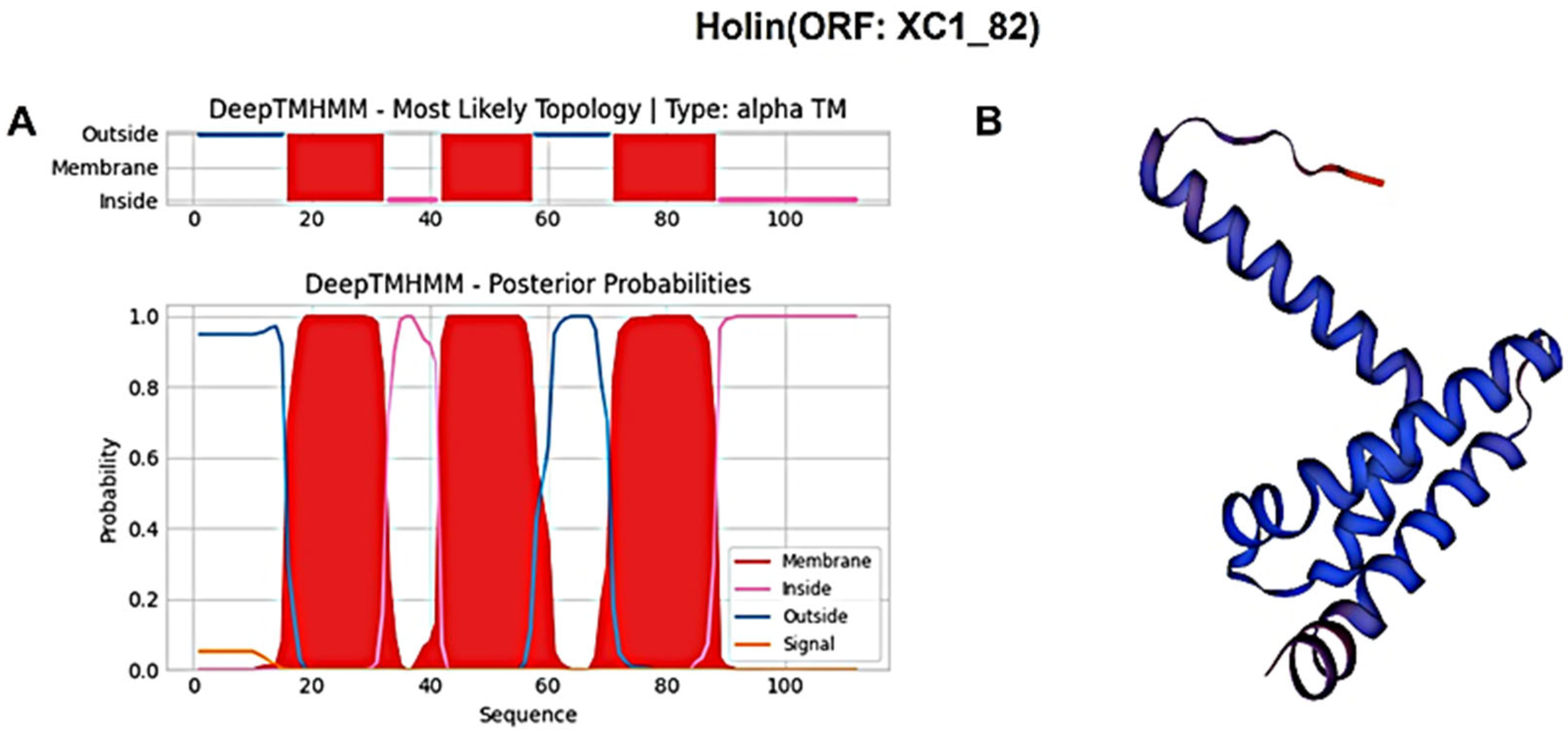
3.6. Analysis of the Lysis System of Phage XC1
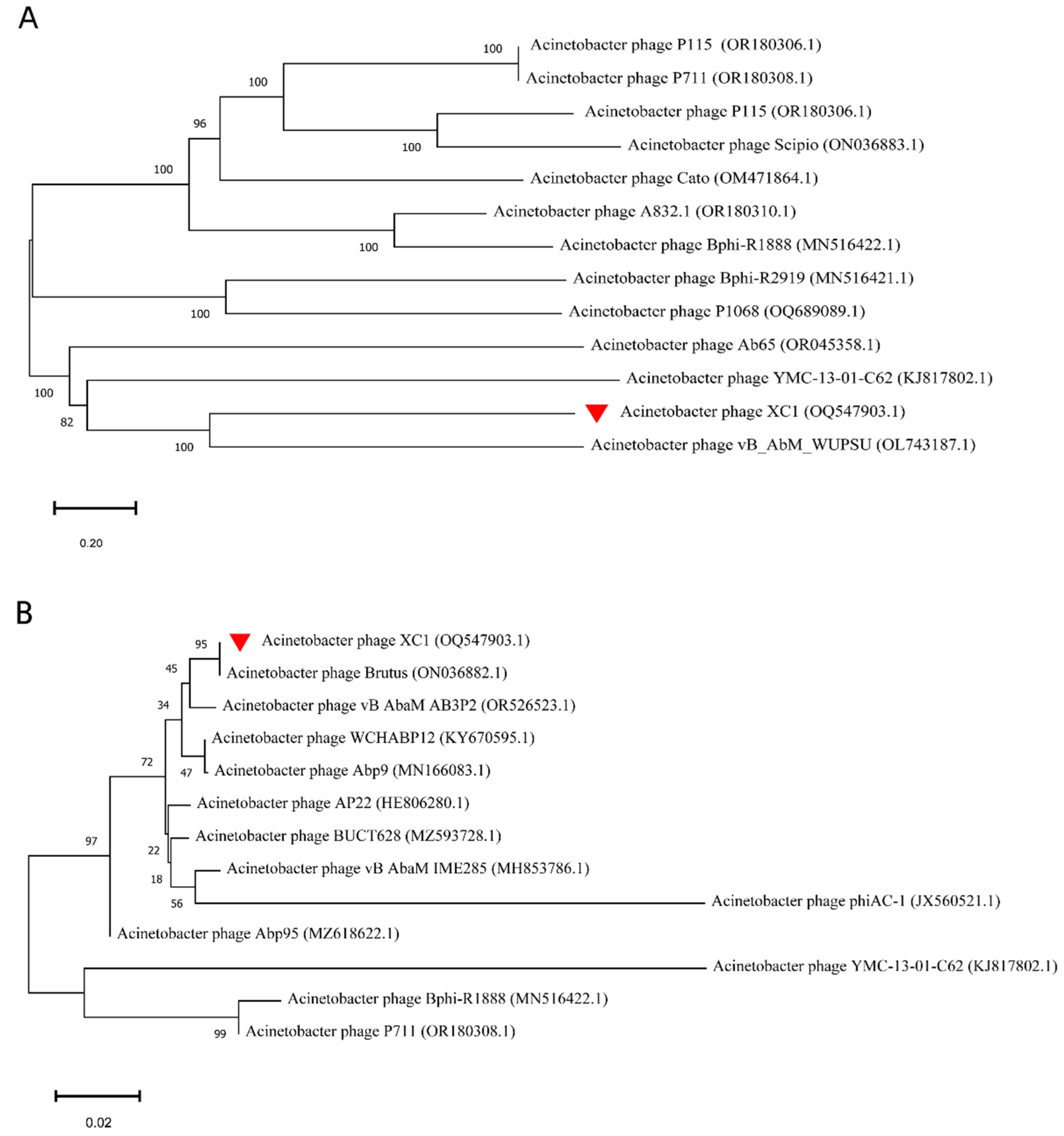
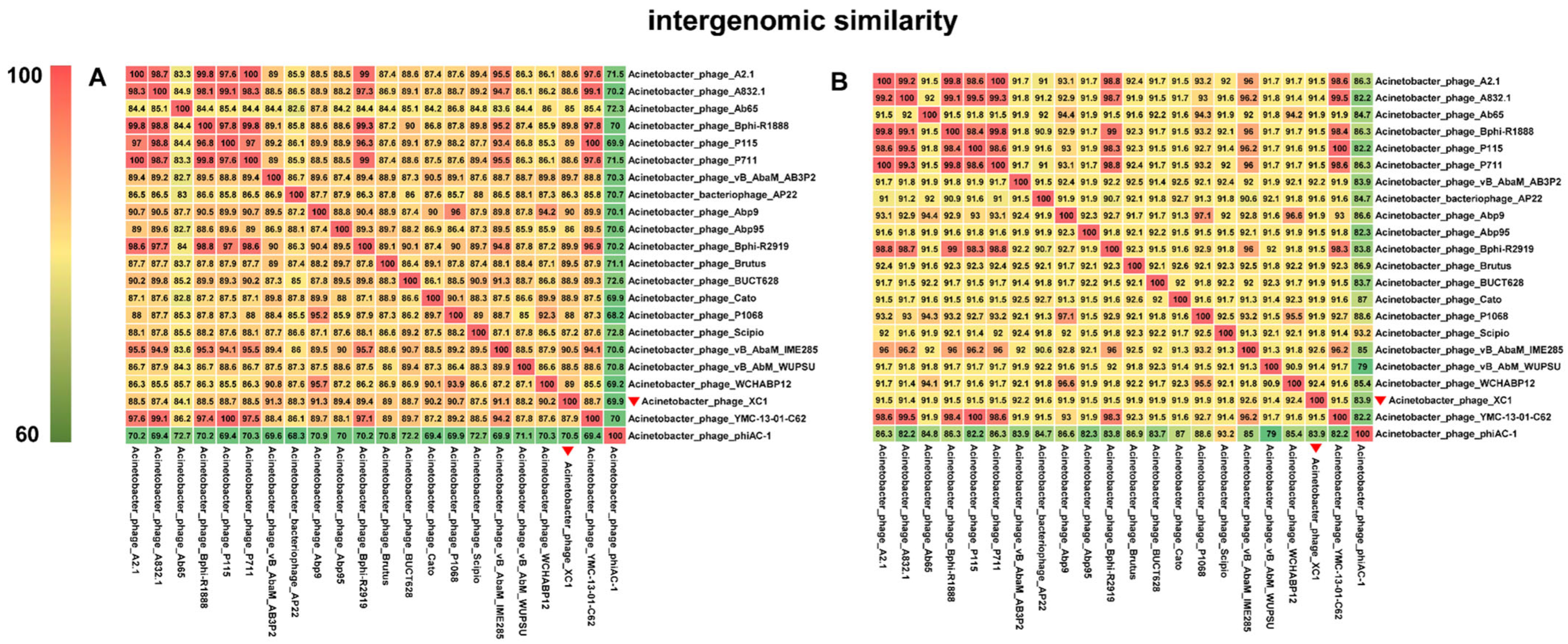
3.7. Phylogenetic Evolutionary Analysis of Phage XC1
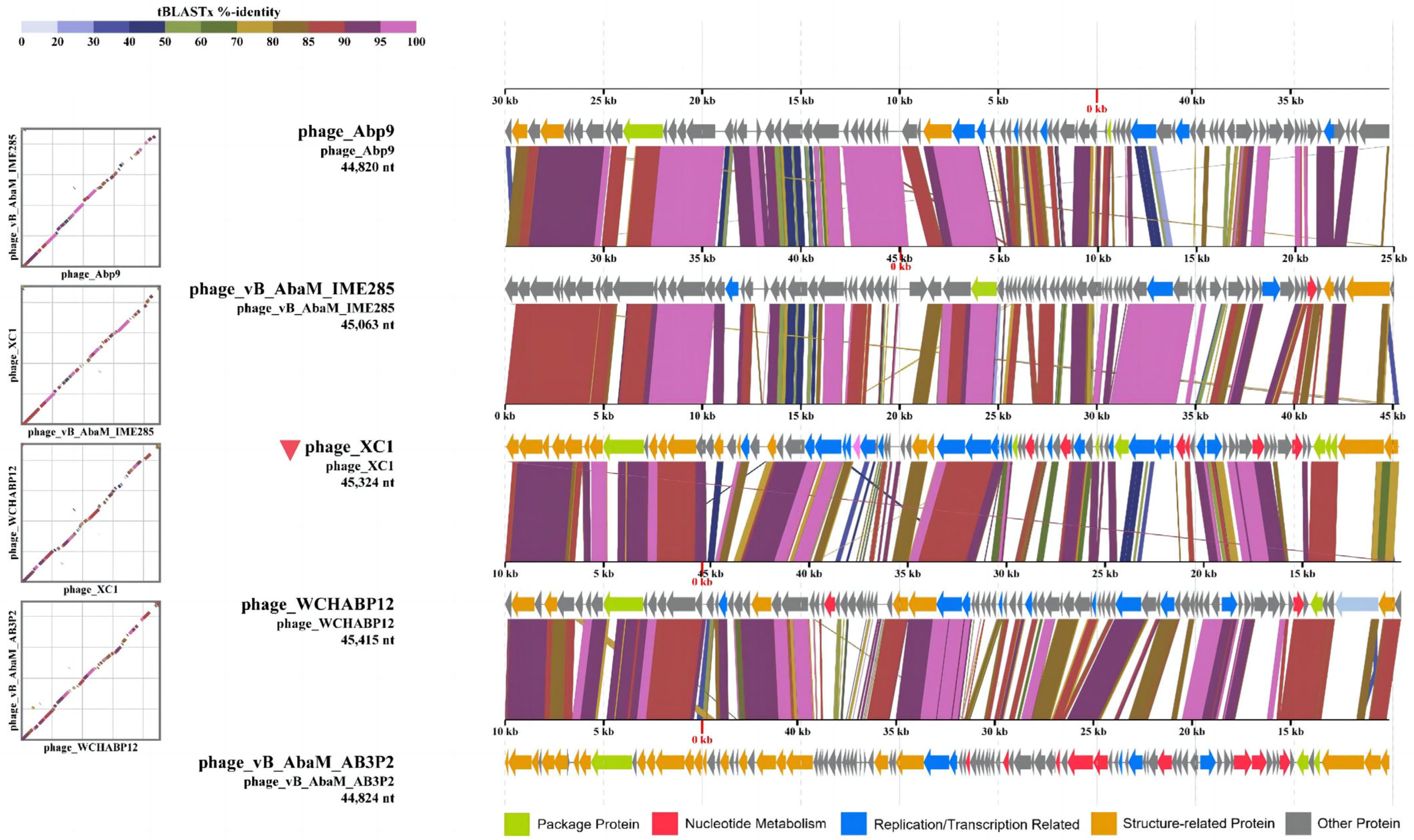
3.8. Comparative Genomics Analysis of Phage XC1
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, P.; Zhang, Y.; Yan, F.; Zhou, X. Characteristics of a Bacteriophage, vB_Kox_ZX8, Isolated from Clinical Klebsiella Oxytoca and Its Therapeutic Effect on Mice Bacteremia. Front. Microbiol. 2021, 12, 763136. [Google Scholar] [CrossRef] [PubMed]
- Sikora, A.; Zahra, F. Nosocomial Infections. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Wisplinghoff, H.; Paulus, T.; Lugenheim, M.; Stefanik, D.; Higgins, P.G.; Edmond, M.B.; Wenzel, R.P.; Seifert, H. Nosocomial Bloodstream Infections Due to Acinetobacter baumannii, Acinetobacter Pittii and Acinetobacter nosocomialis in the United States. J. Infect. 2012, 64, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Genotypic and Phenotypic Characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii Complex—Search Results. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=Genotypic+and+phenotypic+characterization+of+the+Acinetobacter+calcoaceticus-Acinetobacter+baumannii+complex+&filter=pubt.booksdocs (accessed on 4 March 2025).
- Jing, L.; Xu, Z.; Zhang, Y.; Li, D.; Song, Y.; Hu, H.; Fang, Y.; Zhu, W. Metagenomic Insights into Pathogenic Characterization of ST410 Acinetobacter nosocomialis Prevalent in China. Pathogens 2022, 11, 838. [Google Scholar] [CrossRef]
- Yamamoto, M.; Arata, S.; Fukuchi, K.; Honda, H.; Kobayashi, H.; Inagaki, M. Classification of Gases around Pseudomonas Aeruginosa and Acinetobacter baumannii by Infrared Spectroscopy. J. Microbiol. Methods 2022, 196, 106474. [Google Scholar] [CrossRef] [PubMed]
- Kalová, A.; Gelbíčová, T.; Overballe-Petersen, S.; Litrup, E.; Karpíšková, R. Characterisation of Colistin -Resistant Enterobacterales and Acinetobacter Strains Carrying Mcr Genes from Asian Aquaculture Products. Antibiotics 2021, 10, 838. [Google Scholar] [CrossRef]
- Amaral, S.C.; Pruski, B.B.; de Freitas, S.B.; Dos Santos, L.M.; Hartwig, D.D. Biofilm Formation in Drug-Resistant Acinetobacter baumannii and Acinetobacter nosocomialis Isolates Obtained from a University Hospital in Pelotas, RS, Brazil. Lett. Appl. Microbiol. 2023, 76, ovad094. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Y.; Chen, J.; Hong, X.; Xu, X.; Wu, Z.; Ahmed, T.; Loh, B.; Leptihn, S.; Hassan, S.; et al. Identification and Characterization of a New Type of Holin-Endolysin Lysis Cassette in Acidovorax Oryzae Phage AP1. Viruses 2022, 14, 167. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Zeng, F.; Wu, Z.; Jin, Z.; Li, W.; Zhu, M.; Wang, Q.; Tong, Y.; Chen, L.; Bai, Q. Isolation and Genomic Analysis of Temperate Phage 5W Targeting Multidrug-Resistant Acinetobacter baumannii. Arch. Microbiol. 2021, 204, 58. [Google Scholar] [CrossRef] [PubMed]
- Nithichanon, A.; Kewcharoenwong, C.; Da-Oh, H.; Surajinda, S.; Khongmee, A.; Koosakunwat, S.; Wren, B.W.; Stabler, R.A.; Brown, J.S.; Lertmemongkolchai, G. Acinetobacter nosocomialis Causes as Severe Disease as Acinetobacter baumannii in Northeast Thailand: Underestimated Role of A. nosocomialis in Infection. Microbiol. Spectr. 2022, 10, e0283622. [Google Scholar] [CrossRef]
- Mancilla-Rojano, J.; Ochoa, S.A.; Reyes-Grajeda, J.P.; Flores, V.; Medina-Contreras, O.; Espinosa-Mazariego, K.; Parra-Ortega, I.; Rosa-Zamboni, D.D.L.; Castellanos-Cruz, M.D.C.; Arellano-Galindo, J.; et al. Molecular Epidemiology of Acinetobacter calcoaceticus-Acinetobacter baumannii Complex Isolated from Children at the Hospital Infantil de México Federico Gómez. Front. Microbiol. 2020, 11, 576673. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, W.; Zhang, Z.; Gu, Y.; Huang, A.; Wang, J.; Hao, H. Phage Products for Fighting Antimicrobial Resistance. Microorganisms 2022, 10, 1324. [Google Scholar] [CrossRef] [PubMed]
- Duerkop, B.A.; Palmer, K.L.; Horsburgh, M.J. Enterococcal Bacteriophages and Genome Defense. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Pekkle Lam, H.Y.; Lai, M.-J.; Wu, W.-J.; Chin, Y.-H.; Chao, H.-J.; Chen, L.-K.; Peng, S.-Y.; Chang, K.-C. Isolation and Characterization of Bacteriophages with Activities against Multi-Drug-Resistant Acinetobacter nosocomialis Causing Bloodstream Infection in Vivo. J. Microbiol. Immunol. Infect. = Wei Mian Yu Gan Ran Za Zhi 2023, 56, 1026–1035. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, A.; Lei, C.; Wang, H.; Guan, Z.; Xu, C.; Liu, B.; Zhang, D.; Li, Q.; Jiang, W.; et al. Characteristics of Plasmids Coharboring 16S rRNA Methylases, CTX-M, and Virulence Factors in Escherichia coli and Klebsiella pneumoniae Isolates from Chickens in China. Foodborne Pathog. Dis. 2015, 12, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Zhang, L.; Feng, J.; Wu, D.; Wu, L.; Pan, W.; Jiang, Y.; Chen, M.; Chen, J.; Shui, P. Hypoxia-Responsive Covalent Organic Framework Nanoplatform for Breast-Cancer-Targeted Cocktail Immunotherapy via Triple Therapeutic Switch Mechanisms. Small 2025, 21, e2407553. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Xiong, Y.; Lin, Z.; Chu, X.; Lv, B.; Lu, L.; Lin, C.; Liao, J.; Ouyang, L.; Sun, Y.; et al. Novel Cocktail Therapy Based on Multifunctional Supramolecular Hydrogel Targeting Immune-Angiogenesis-Nerve Network for Enhanced Diabetic Wound Healing. J. Nanobiotechnol. 2024, 22, 749. [Google Scholar] [CrossRef] [PubMed]
- Valério, N.; Oliveira, C.; Jesus, V.; Branco, T.; Pereira, C.; Moreirinha, C.; Almeida, A. Effects of Single and Combined Use of Bacteriophages and Antibiotics to Inactivate Escherichia coli. Virus Res. 2017, 240, 8–17. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF | Locus | Length | Function | Protein ID |
|---|---|---|---|---|
| XC1_01 | − | 627 | putative structural protein | WFD61208.1 |
| XC1_02 | − | 1185 | baseplate J-like protein | WFD61209.1 |
| XC1_03 | − | 354 | tail sheath intiator protein | WFD61210.1 |
| XC1_04 | − | 648 | putative baseplate assembly protein | WFD61211.1 |
| XC1_05 | − | 891 | baseplate hub | WFD61212.1 |
| XC1_06 | − | 279 | virion structural protein | WFD61213.1 |
| XC1_07 | − | 657 | tail tube initiator-like protein | WFD61214.1 |
| XC1_08 | − | 2091 | lysozyme-like protein | WFD61215.1 |
| XC1_09 | − | 213 | putative tail-fiber/lysozyme protein | WFD61216.1 |
| XC1_10 | − | 426 | tail assembly chaperone | WFD61217.1 |
| XC1_11 | − | 450 | virion structural protein | WFD61218.1 |
| XC1_12 | − | 1464 | tail sheath | WFD61219.1 |
| XC1_13 | − | 495 | hypothetical protein | WFD61220.1 |
| XC1_14 | − | 414 | cysteine protease | WFD61221.1 |
| XC1_15 | − | 504 | head-tail adaptor | WFD61222.1 |
| XC1_16 | − | 447 | hypothetical protein | WFD61223.1 |
| XC1_17 | − | 186 | neck protein | WFD61224.1 |
| XC1_18 | − | 438 | putative RNA polymerase | WFD61225.1 |
| XC1_19 | − | 477 | hypothetical protein | WFD61226.1 |
| XC1_20 | − | 453 | putative head tail connector protein | WFD61227.1 |
| XC1_21 | − | 354 | hypothetical protein | WFD61228.1 |
| XC1_22 | − | 1008 | putative capsid protein | WFD61229.1 |
| XC1_23 | − | 495 | HNH endonuclease domain protein | WFD61230.1 |
| XC1_24 | − | 1347 | prohead core protein protease | WFD61231.1 |
| XC1_25 | − | 168 | primase | WFD61232.1 |
| XC1_26 | − | 276 | putative transcriptional regulator | WFD61233.1 |
| XC1_27 | − | 363 | HNH endonuclease | WFD61234.1 |
| XC1_28 | − | 777 | primase | WFD61235.1 |
| XC1_29 | − | 150 | putative membrane protein | WFD61236.1 |
| XC1_30 | − | 282 | recombination protein | WFD61237.1 |
| XC1_31 | − | 183 | hypothetical protein | WFD61238.1 |
| XC1_32 | − | 171 | hypothetical protein | WFD61239.1 |
| XC1_33 | − | 231 | hypothetical protein | WFD61240.1 |
| XC1_34 | − | 279 | hypothetical protein | WFD61241.1 |
| XC1_35 | − | 771 | head protein | WFD61242.1 |
| XC1_36 | − | 345 | putative head protein | WFD61243.1 |
| XC1_37 | − | 1440 | portal protein | WFD61244.1 |
| XC1_38 | − | 1302 | terminase large subunit | WFD61245.1 |
| XC1_39 | − | 432 | terminase small subunit | WFD61246.1 |
| XC1_40 | − | 165 | primase | WFD61247.1 |
| XC1_41 | − | 243 | hypothetical protein | WFD61248.1 |
| XC1_42 | − | 201 | HTH DNA binding protein | WFD61249.1 |
| XC1_43 | − | 276 | F-box domain protein | WFD61250.1 |
| XC1_44 | − | 195 | hypothetical protein | WFD61251.1 |
| XC1_45 | − | 210 | hypothetical protein | WFD61252.1 |
| XC1_46 | − | 444 | putative phosphoesterase | WFD61253.1 |
| XC1_47 | − | 222 | hypothetical protein | WFD61254.1 |
| XC1_48 | − | 240 | hypothetical protein | WFD61255.1 |
| XC1_49 | − | 282 | endonuclease | WFD61256.1 |
| XC1_50 | − | 390 | hypothetical protein | WFD61257.1 |
| XC1_51 | − | 534 | nucleotide kinase | WFD61258.1 |
| XC1_52 | − | 165 | hypothetical protein | WFD61259.1 |
| XC1_53 | − | 576 | DNA helicase | WFD61260.1 |
| XC1_54 | − | 408 | hypothetical protein | WFD61261.1 |
| XC1_55 | − | 213 | superinfection immunity protein | WFD61262.1 |
| XC1_56 | − | 162 | hypothetical protein | WFD61263.1 |
| XC1_57 | − | 222 | hypothetical protein | WFD61264.1 |
| XC1_58 | − | 267 | recombination protein | WFD61265.1 |
| XC1_59 | − | 729 | putative LAGLIDADG homing endonuclease | WFD61266.1 |
| XC1_60 | − | 1314 | replicative DNA helicase | WFD61267.1 |
| XC1_61 | − | 774 | putative replicative DNA helicase | WFD61268.1 |
| XC1_62 | − | 198 | transcriptional regulator | WFD61269.1 |
| XC1_63 | − | 456 | DNA N-6-adenine-methyltransferase | WFD61270.1 |
| XC1_64 | − | 243 | exonuclease | WFD61271.1 |
| XC1_65 | − | 252 | hypothetical protein | WFD61272.1 |
| XC1_66 | − | 426 | helix-turn-helix transcriptional regulator | WFD61273.1 |
| XC1_67 | + | 762 | transcriptional regulator | WFD61274.1 |
| XC1_68 | + | 213 | DUF551 domain-containing protein | WFD61275.1 |
| XC1_69 | + | 336 | hypothetical protein | WFD61276.1 |
| XC1_70 | + | 183 | hypothetical protein | WFD61277.1 |
| XC1_71 | + | 681 | ERF family protein | WFD61278.1 |
| XC1_72 | + | 603 | DNA exonuclease | WFD61279.1 |
| XC1_73 | + | 294 | hypothetical protein | WFD61280.1 |
| XC1_74 | + | 195 | hypothetical protein | WFD61281.1 |
| XC1_75 | + | 162 | hypothetical protein | WFD61282.1 |
| XC1_76 | + | 723 | hypothetical protein | WFD61283.1 |
| XC1_77 | + | 540 | dUTP diphosphatase | WFD61284.1 |
| XC1_78 | + | 231 | hypothetical protein | WFD61285.1 |
| XC1_79 | + | 198 | hypothetical protein | WFD61286.1 |
| XC1_80 | − | 609 | phage endolysin | WFD61287.1 |
| XC1_81 | − | 276 | holin | WFD61288.1 |
| XC1_82 | − | 339 | holin | WFD61289.1 |
| XC1_83 | − | 2328 | tail fiber protein | WFD61290.1 |
| XC1_84 | − | 726 | tail fiber protein | WFD61291.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Zhao, S.; Jiang, H.; Shi, H.; Li, J.; Zhao, C.; Huang, H. Biological Characteristics and Genomic Analysis of Acinetobacter nosocomialis Lytic Phage XC1. Curr. Issues Mol. Biol. 2025, 47, 335. https://doi.org/10.3390/cimb47050335
Wang C, Zhao S, Jiang H, Shi H, Li J, Zhao C, Huang H. Biological Characteristics and Genomic Analysis of Acinetobacter nosocomialis Lytic Phage XC1. Current Issues in Molecular Biology. 2025; 47(5):335. https://doi.org/10.3390/cimb47050335
Chicago/Turabian StyleWang, Chuhan, Shuchuan Zhao, Hailin Jiang, Hongyan Shi, Jinghua Li, Chunyan Zhao, and Honglan Huang. 2025. "Biological Characteristics and Genomic Analysis of Acinetobacter nosocomialis Lytic Phage XC1" Current Issues in Molecular Biology 47, no. 5: 335. https://doi.org/10.3390/cimb47050335
APA StyleWang, C., Zhao, S., Jiang, H., Shi, H., Li, J., Zhao, C., & Huang, H. (2025). Biological Characteristics and Genomic Analysis of Acinetobacter nosocomialis Lytic Phage XC1. Current Issues in Molecular Biology, 47(5), 335. https://doi.org/10.3390/cimb47050335