

Immunomodulatory and Antioxidant Properties of Ipomoea batatas Flour and Extracts Obtained by Green Extraction
 , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Phytochemichal Analysis and Glycemic Index
2.2.1. Determination of Carbohydrates
Total Starch
Free Sugars
Resistant Starch
Determination of Amylose
Quantification of Insoluble Fiber
Measurement of the Glycemic Index (GI)
2.3. Identification and Quantification of Bioactive Phytochemicals
2.3.1. Total Phenolic Content Test
2.3.2. Identification and Quantification of Carotenoids
2.3.3. Determination of Mineral Materials
2.4. Evaluation of the Bioactive Potential
2.4.1. Antioxidant Activity
Determination of NO (Nitric Oxide) Scavenging
DPPH (2,2-Diphenyl-1-picrylhydrazyl) Assay
ORAC (Oxygen Radical Absorbance Capacity) Assay
2.4.2. Immunomodulatory Anti-Inflammatory Bioactivity
Macrophage Culture
Cell Viability by MTS/PMS Assay
Productions of NO, IL-6, TNF-α, MCP-1, and PGE-2
Determination of Nitrites (NO)
IL-6 (Interleukin 6) Assay
TNF-alpha (Tumor Necrosis Factor Alpha) Assay
MCP-1 (Monocyte Chemoattractant Protein-1) Assay
PGE-2 (Prostaglandin) Assay
2.5. Statistical Analysis
3. Results
3.1. Phytochemical Analysis and Glycemic Index
3.2. Identification and Quantification of Bioactive Phytochemicals
3.3. Antioxidant Bioactivity
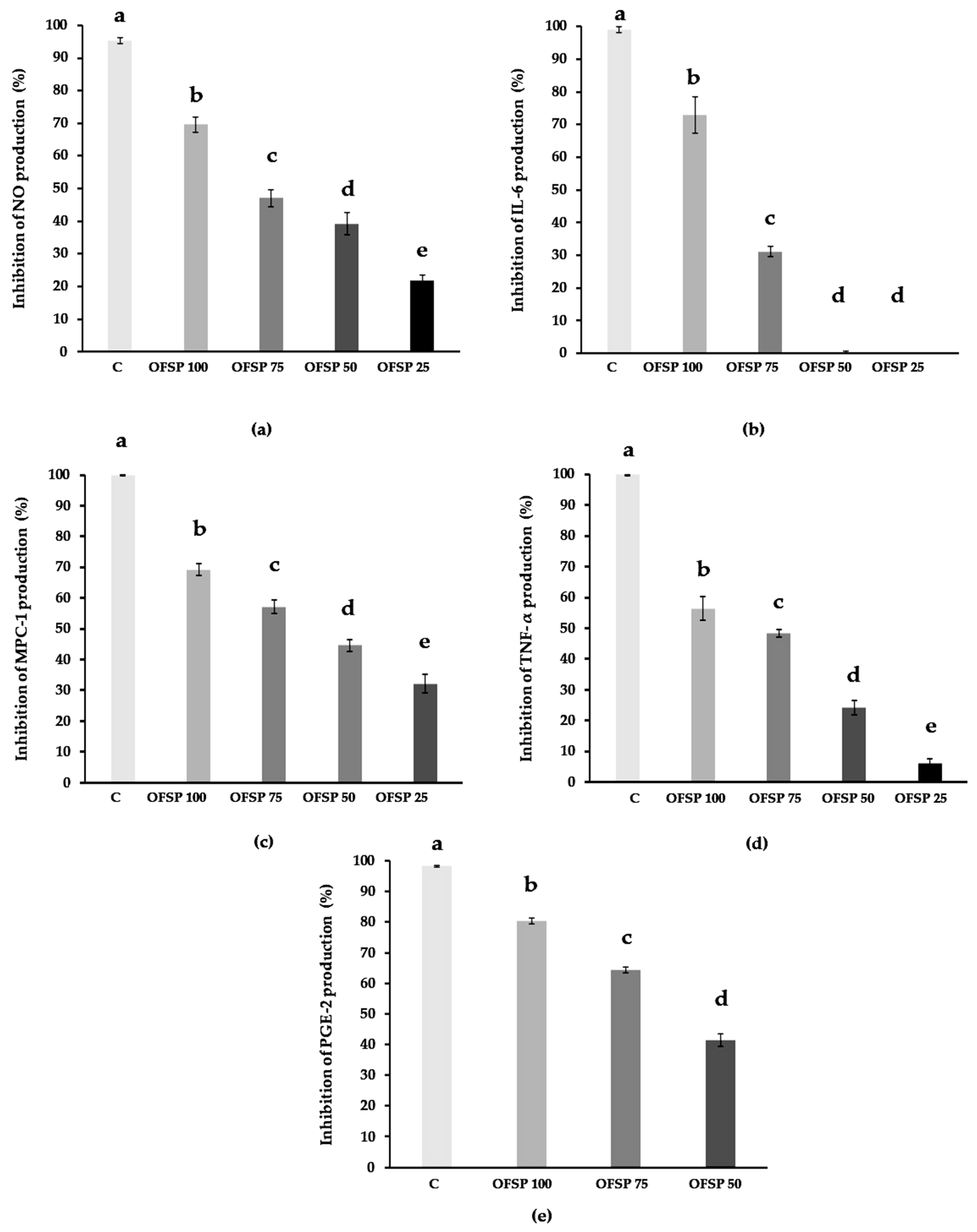
3.4. Immunomodulatory Anti-Inflammatory Bioactivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, R.; Zheng, W.; Jiang, M.; Zhang, H. A Review of Starch Biosynthesis in Cereal Crops and Its Potential Breeding Applications in Rice (Oryza sativa L.). Peer J. 2021, 9, e12678. [Google Scholar] [CrossRef]
- Van Ittersum, M.K.; van Bussel, L.G.J.; Wolf, J.; Grassini, P.; van Wart, J.; Guilpart, N.; Claessens, L.; de Groot, H.; Wiebe, K.; Mason-D’Croz, D.; et al. Can Sub-Saharan Africa Feed Itself? Proc. Natl. Acad. Sci. USA 2016, 113, 14964–14969. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, A.; Josheph Kumar, T. Roots and Tuber Crops as Functional Foods: A Review on Phytochemical Constituents and Their Potential Health Benefits. Int. J. Food Sci. 2016, 2016, e3631647. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QV (accessed on 23 March 2022).
- Escobar-Puentes, A.A.; Palomo, I.; Rodríguez, L.; Fuentes, E.; Villegas-Ochoa, M.A.; González-Aguilar, G.A.; Olivas-Aguirre, F.J.; Wall-Medrano, A. Sweet Potato (Ipomoea batatas L.) Phenotypes: From Agroindustry to Health Effects. Foods 2022, 11, 1058. [Google Scholar] [CrossRef] [PubMed]
- Le Prix Mondial de L’Alimentation—Améliorer la Qualité, la Quantité et la Disponibilité des Aliments Dans le Monde. Available online: https://www.worldfoodprize.org/ (accessed on 16 November 2022).
- Jenkins, M.; Byker Shanks, C.; Houghtaling, B. Orange-Fleshed Sweet Potato: Successes and Remaining Challenges of the Introduction of a Nutritionally Superior Staple Crop in Mozambique. Food Nutr. Bull. 2015, 36, 327–353. [Google Scholar] [CrossRef] [PubMed]
- Laurie, S.M.; Faber, M.; Claasen, N. Incorporating Orange-Fleshed Sweet Potato into the Food System as a Strategy for Improved Nutrition: The Context of South Africa. Food Res. Int. 2018, 104, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Low, J.W.; Mwanga, R.O.M.; Andrade, M.; Carey, E.; Ball, A.-M. Tackling Vitamin A Deficiency with Biofortified Sweetpotato in Sub-Saharan Africa. Glob. Food Sec. 2017, 14, 23–30. [Google Scholar] [CrossRef]
- Low, J.W.; Thiele, G. Understanding Innovation: The Development and Scaling of Orange-Fleshed Sweetpotato in Major African Food Systems. Agric. Syst. 2020, 179, 102770. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Ilangantileke, S. Dietary intervention with orange fleshed sweetpotato (Ipomoea batatas (L.) Lam.) To alleviate vitamin a deficiency in south and west asia. Acta Hortic. 2002, 583, 205–210. [Google Scholar] [CrossRef]
- Darnton-Hill, I. Public Health Aspects in the Prevention and Control of Vitamin Deficiencies. Curr. Dev. Nutr. 2019, 3, nzz075. [Google Scholar] [CrossRef]
- Xu, Y.; Shan, Y.; Lin, X.; Miao, Q.; Lou, L.; Wang, Y.; Ye, J. Global patterns in vision loss burden due to vitamin A deficiency from 1990 to 2017. Public Health Nutr. 2021, 24, 5786–5794. [Google Scholar] [CrossRef] [PubMed]
- Aibana, O.; Franke, M.F.; Huang, C.-C.; Galea, J.T.; Calderon, R.; Zhang, Z.; Becerra, M.C.; Smith, E.R.; Ronnenberg, A.G.; Contreras, C.; et al. Impact of Vitamin A and Carotenoids on the Risk of Tuberculosis Progression. Clin. Infect. Dis. 2017, 65, 900–909. [Google Scholar] [CrossRef]
- Woloszynowska-Fraser, M.U.; Kouchmeshky, A.; McCaffery, P. Vitamin A and Retinoic Acid in Cognition and Cognitive Disease. Annu. Rev. Nutr. 2020, 40, 247–272. [Google Scholar] [CrossRef]
- Global Prevalence of Vitamin A Deficiency in Populations at Risk 1995–2005: WHO Global Database on Vitamin A Deficiency. Available online: https://www.who.int/publications-detail-redirect/9789241598019 (accessed on 16 November 2022).
- Aguayo, V.M.; Kahn, S.; Ismael, C.; Meershoek, S. Vitamin A Deficiency and Child Mortality in Mozambique. Public Health Nutr. 2005, 8, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Weinstein, S.J.; Yu, K.; Männistö, S.; Albanes, D. Serum Beta Carotene and Overall and Cause-Specific Mortality. Circ. Res. 2018, 123, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, R.; Hui, J.; Li, L.; Zheng, X. β-Carotene Attenuates LPS-Induced Rat Intestinal Inflammation via Modulating Autophagy and Regulating the JAK2/STAT3 and JNK/P38 MAPK Signaling Pathways. J. Food Biochem. 2021, 45, e13544. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y.; Na, X.; Zhao, A. β-Carotene Supplementation and Risk of Cardiovascular Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2022, 14, 1284. [Google Scholar] [CrossRef]
- Anand, R.; Mohan, L.; Bharadvaja, N. Disease Prevention and Treatment Using β-Carotene: The Ultimate Provitamin A. Rev. Bras Farm. 2022, 32, 491–501. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef]
- Shimozono, H.; Kobori, M.; Shinmoto, H.; Tsushida, T. Suppression of the Melanogenesis of Mouse Melanoma B 16 Cells by Sweet Potato Extract. Nippon. Shokuhin Kogyo Gakkai Shi 1996, 43, 313–317. [Google Scholar]
- Ishiguro, K.; Yahara, S.; Yoshimoto, M. Changes in Polyphenolic Content and Radical-Scavenging Activity of Sweetpotato (Ipomoea batatas L.) during Storage at Optimal and Low Temperatures. J. Agric. Food Chem. 2007, 55, 10773–10778. [Google Scholar] [CrossRef] [PubMed]
- Mu, T.-H.; Singh, J. Chapter 1—Sweet Potato: Chemistry, Processing, and Nutrition—An Introduction. In Sweet Potato; Mu, T.-H., Singh, J., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 1–4. [Google Scholar] [CrossRef]
- Kim, M.Y.; Lee, B.W.; Lee, H.-U.; Lee, Y.Y.; Kim, M.H.; Lee, J.Y.; Lee, B.K.; Woo, K.S.; Kim, H.-J. Phenolic Compounds and Antioxidant Activity in Sweet Potato after Heat Treatment. J. Sci. Food Agric. 2019, 99, 6833–6840. [Google Scholar] [CrossRef]
- Majid, M.; Nasir, B.; Zahra, S.S.; Khan, M.R.; Mirza, B.; Haq, I. Ipomoea batatas L. Lam. Ameliorates Acute and Chronic Inflammations by Suppressing Inflammatory Mediators, a Comprehensive Exploration Using in Vitro and in Vivo Models. BMC Complement. Altern. Med. 2018, 18, 216. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Biasutto, L.; Mattarei, A.; Zoratti, M. Resveratrol and Health: The Starting Point. ChemBioChem 2012, 13, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Si, L.-Y. Resveratrol Role in Cardiovascular and Metabolic Health and Potential Mechanisms of Action. Nutr. Res. 2012, 32, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.A.; Olson, E.N. Cardiac Plasticity. N. Engl. J. Med. 2008, 358, 1370–1380. [Google Scholar] [CrossRef] [PubMed]
- Owen, R.W.; Giacosa, A.; Hull, W.E.; Haubner, R.; Spiegelhalder, B.; Bartsch, H. The Antioxidant/Anticancer Potential of Phenolic Compounds Isolated from Olive Oil. Eur. J. Cancer 2000, 36, 1235–1247. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106. [Google Scholar] [CrossRef]
- Mao, T.K.; van de Water, J.; Keen, C.L.; Schmitz, H.H.; Gershwin, M.E. Modulation of TNF-α Secretion in Peripheral Blood Mononuclear Cells by Cocoa Flavanols and Procyanidins. Dev. Immunol. 2002, 9, 135–141. [Google Scholar] [CrossRef]
- Leiherer, A.; Mündlein, A.; Drexel, H. Phytochemicals and Their Impact on Adipose Tissue Inflammation and Diabetes. Vasc. Pharmacol. 2013, 58, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Rivera, L.; Morón, R.; Sánchez, M.; Zarzuelo, A.; Galisteo, M. Quercetin Ameliorates Metabolic Syndrome and Improves the Inflammatory Status in Obese Zucker Rats. Obesity 2008, 16, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Polyphenols and Glycemic Control. Nutrients 2016, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Alfaddagh, A.; Martin, S.S.; Leucker, T.M.; Michos, E.D.; Blaha, M.J.; Lowenstein, C.J.; Jones, S.R.; Toth, P.P. Inflammation and Cardiovascular Disease: From Mechanisms to Therapeutics. Am. J. Prev. Cardiol. 2020, 4, 100130. [Google Scholar] [CrossRef]
- Otręba, M.; Kośmider, L.; Stojko, J.; Rzepecka-Stojko, A. Cardioprotective Activity of Selected Polyphenols Based on Epithelial and Aortic Cell Lines. A Review. Molecules 2020, 25, 5343. [Google Scholar] [CrossRef] [PubMed]
- ISO Standards. ISO 6647-1:2020: Determination of Amylose Content. Available online: https://www.iso.org/standard (accessed on 2 February 2023).
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Arnould, S.; Vitou, M.; Boudard, F.; Guzman, C.; Poucheret, P.; Fons, F.; Rapior, S. Antiproliferative and Antioxidant Activities of Wild Boletales Mushrooms from France. IJM 2018, 20. [Google Scholar] [CrossRef]
- Boukhers, I.; Boudard, F.; Morel, S.; Servent, A.; Portet, K.; Guzman, C.; Vitou, M.; Kongolo, J.; Michel, A.; Poucheret, P. Nutrition, Healthcare Benefits and Phytochemical Properties of Cassava (Manihot esculenta) Leaves Sourced from Three Countries (Reunion, Guinea, and Costa Rica). Foods 2022, 11, 2027. [Google Scholar] [CrossRef]
- Laurie, S.M.; Calitz, F.J.; Adebola, P.O.; Lezar, A. Characterization and Evaluation of South African Sweet Potato (Ipomoea batatas (L.) LAM) Land Races. S. Afr. J. Bot. 2013, 85, 10–16. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in Human Nutrition and Health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Abert-Vian, M.; Fabiano-Tixier, A.S.; Strube, J.; Uhlenbrock, L.; Gunjevic, V.; Cravotto, G. Green extraction of natural products. Origins, current status, and future challenges. TrAC Trends Anal. Chem. 2019, 118, 248–263. [Google Scholar] [CrossRef]
- Lai, Y.-C.; Huang, C.-L.; Chan, C.-F.; Lien, C.-Y.; Liao, W.C. Studies of Sugar Composition and Starch Morphology of Baked Sweet Potatoes (Ipomoea batatas (L.) Lam). J. Food Sci. Technol. 2013, 50, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, G.; Grosser, T. Prostanoids and Inflammatory Pain. Prostaglandins Other Lipid Mediat. 2013, 104, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Aprianita, A.; Vasiljevic, T.; Bannikova, A.; Kasapis, S. Physicochemical Properties of Flours and Starches Derived from Traditional Indonesian Tubers and Roots. J. Food Sci. Technol. 2014, 51, 3669–3679. [Google Scholar] [CrossRef] [PubMed]
- Abegunde, O.K.; Mu, T.-H.; Chen, J.-W.; Deng, F.-M. Physicochemical Characterization of Sweet Potato Starches Popularly Used in Chinese Starch Industry. Food Hydrocoll. 2013, 33, 169–177. [Google Scholar] [CrossRef]
- Kim, H.-J.; Woo, K.S.; Lee, H.-U.; Nam, S.S.; Lee, B.W.; Kim, M.Y.; Lee, Y.-Y.; Lee, J.Y.; Kim, M.H.; Lee, B. Physicochemical Characteristics of Starch in Sweet Potato Cultivars Grown in Korea. Prev. Nutr. Food Sci. 2020, 25, 212–218. [Google Scholar] [CrossRef]
- Benesi, I.R.; Labuschagne, M.T.; Dixon, A.G.; Mahungu, N.M. Stability of Native Starch Quality Parameters, Starch Extraction and Root Dry Matter of Cassava Genotypes in Different Environments. J. Sci. Food Agric. 2004, 84, 1381–1388. [Google Scholar] [CrossRef]
- Osunsami, A.T.; Akingbala, J.O.; Oguntimein, G.B. Effect of Storage on Starch Content and Modification of Cassava Starch. Starch Stärke 1989, 41, 54–57. [Google Scholar] [CrossRef]
- Benesi, I.R.M.; Labuschagne, M.T.; Herselman, L.; Mahungu, N.M.; Saka, J.K. The Effect of Genotype, Location and Season on Cassava Starch Extraction. Euphytica 2008, 160, 59–74. [Google Scholar] [CrossRef]
- Morris, K.L.; Zemel, M.B. Glycemic Index, Cardiovascular Disease, and Obesity. Nutr. Rev. 1999, 57 Pt 1, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Bhupathiraju, S.N.; Tobias, D.K.; Malik, V.S.; Pan, A.; Hruby, A.; Manson, J.E.; Willett, W.C.; Hu, F.B. Glycemic Index, Glycemic Load, and Risk of Type 2 Diabetes: Results from 3 Large US Cohorts and an Updated Meta-Analysis. Am. J. Clin. Nutr. 2014, 100, 218–232. [Google Scholar] [CrossRef] [PubMed]
- Oba, S.; Nanri, A.; Kurotani, K.; Goto, A.; Kato, M.; Mizoue, T.; Noda, M.; Inoue, M.; Tsugane, S.; Japan Public Health Center-Based Prospective Study Group. Dietary Glycemic Index, Glycemic Load and Incidence of Type 2 Diabetes in Japanese Men and Women: The Japan Public Health Center-Based Prospective Study. Nutr. J. 2013, 12, 165. [Google Scholar] [CrossRef] [PubMed]
- Brand, J.C.; Colagiuri, S.; Crossman, S.; Allen, A.; Roberts, D.C.; Truswell, A.S. Low-Glycemic Index Foods Improve Long-Term Glycemic Control in NIDDM. Diabetes Care 1991, 14, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Owen, B.; Wolever, T.M. Effect of Fat on Glycaemic Responses in Normal Subjects: A Dose-Response Study. Nutr. Res. 2003, 23, 1341–1347. [Google Scholar] [CrossRef]
- Allen, J.C.; Corbitt, A.D.; Maloney, K.P.; Butt, M.S.; Truong, V.-D. Glycemic index of sweet potato as affected by cooking methods. Open Nutr. J. 2012, 6, 1–11. [Google Scholar] [CrossRef]
- Zenel, A.M.; Stewart, M.L. High Amylose White Rice Reduces Post-Prandial Glycemic Response but Not Appetite in Humans. Nutrients 2015, 7, 5362–5374. [Google Scholar] [CrossRef]
- Belobrajdic, D.P.; Regina, A.; Klingner, B.; Zajac, I.; Chapron, S.; Berbezy, P.; Bird, A.R. High-Amylose Wheat Lowers the Postprandial Glycemic Response to Bread in Healthy Adults: A Randomized Controlled Crossover Trial. J. Nutr. 2019, 149, 1335–1345. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Wolever, T.M.; Taylor, R.H.; Barker, H.; Fielden, H.; Baldwin, J.M.; Bowling, A.C.; Newman, H.C.; Jenkins, A.L.; Goff, D.V. Glycemic Index of Foods: A Physiological Basis for Carbohydrate Exchange. Am. J. Clin. Nutr. 1981, 34, 362–366. [Google Scholar] [CrossRef]
- Cui, R.; Zhu, F. Physicochemical and Functional Properties of Sweetpotato Flour. J. Sci. Food Agric. 2019, 99, 4624–4634. [Google Scholar] [CrossRef]
- Fujii, H.; Iwase, M.; Ohkuma, T.; Ogata-Kaizu, S.; Ide, H.; Kikuchi, Y.; Idewaki, Y.; Joudai, T.; Hirakawa, Y.; Uchida, K.; et al. Impact of Dietary Fiber Intake on Glycemic Control, Cardiovascular Risk Factors and Chronic Kidney Disease in Japanese Patients with Type 2 Diabetes Mellitus: The Fukuoka Diabetes Registry. Nutr. J. 2013, 12, 159. [Google Scholar] [CrossRef]
- Mwanga, R.O.M.; Odongo, B.; Niringiye, C.; Alajo, A.; Abidin, P.E.; Kapinga, R.; Tumwegamire, S.; Lemaga, B.; Nsumba, J.; Carey, E.E. Release of Two Orange-Fleshed Sweetpotato Cultivars, ‘SPK004’ (‘Kakamega’) and ‘Ejumula’, in Uganda. HortScience 2007, 42, 1728–1730. [Google Scholar] [CrossRef]
- Mohanraj, R.; Sivasankar, S. Sweet Potato (Ipomoea batatas (L.) Lam)—A Valuable Medicinal Food: A Review. J. Med. Food 2014, 17, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Ben Amara, N.; Tourniaire, F.; Maraninchi, M.; Attia, N.; Amiot-Carlin, M.J.; Raccah, D.; Valéro, R.; Landrier, J.F.; Darmon, P. Independent Positive Association of Plasma β-Carotene Concentrations with Adiponectin among Non-Diabetic Obese Subjects. Eur. J. Nutr. 2015, 54, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Krochmal-Marczak, B.; Zagórska-Dziok, M.; Michalak, M.; Kiełtyka-Dadasiewicz, A. Comparative Assessment of Phenolic Content, Cellular Antioxidant, Antityrosinase and Protective Activities on Skin Cells of Extracts from Three Sweet Potato (Ipomoea batatas (L.) Lam.) Cultivars. J. King Saud Univ. Sci. 2021, 33, 101532. [Google Scholar] [CrossRef]
- Olatunde, G.O.; Henshaw, F.O.; Idowu, M.A.; TomLins, K. Quality Attributes of Sweet Potato Flour as Influenced by Variety, Pretreatment and Drying Method. Food Sci. Nutr. 2015, 4, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Maloney, K.; Truong, V.D.; Allen, J. Susceptibility of Sweet Potato (Ipomoea batatas) Peel Proteins to Digestive Enzymes. Food Sci. Nutr. 2014, 2, 351–360. [Google Scholar] [CrossRef]
- Ahmad, A.; Dempsey, S.K.; Daneva, Z.; Azam, M.; Li, N.; Li, P.-L.; Ritter, J.K. Role of Nitric Oxide in the Cardiovascular and Renal Systems. Int. J. Mol. Sci. 2018, 19, 2605. [Google Scholar] [CrossRef]
- Yokozawa, T.; Chen, C.P.; Tanaka, T. Direct Scavenging of Nitric Oxide by Traditional Crude Drugs. Phytomedicine 2000, 6, 453–463. [Google Scholar] [CrossRef]
- Rautenbach, F.; Faber, M.; Laurie, S.; Laurie, R. Antioxidant Capacity and Antioxidant Content in Roots of 4 Sweetpotato Varieties. J. Food Sci. 2010, 75, C400–C405. [Google Scholar] [CrossRef]
- Franková, H.; Musilová, J.; Árvay, J.; Šnirc, M.; Jančo, I.; Lidiková, J.; Vollmannová, A. Changes in Antioxidant Properties and Phenolics in Sweet Potatoes (Ipomoea batatas L.) Due to Heat Treatments. Molecules 2022, 27, 1884. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.H.; Yousef, G.G.; Gustafson, S.J.; Truong, V.-D.; Yencho, G.C.; Lila, M.A. Phytochemical Changes in Phenolics, Anthocyanins, Ascorbic Acid, and Carotenoids Associated with Sweetpotato Storage and Impacts on Bioactive Properties. Food Chem. 2014, 145, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Teow, C.C.; Truong, V.-D.; McFeeters, R.F.; Thompson, R.L.; Pecota, K.V.; Yencho, G.C. Antioxidant Activities, Phenolic and β-Carotene Contents of Sweet Potato Genotypes with Varying Flesh Colours. Food Chem. 2007, 103, 829–838. [Google Scholar] [CrossRef]
- Agunloye, O.M.; Oboh, G.; Ademiluyi, A.O.; Ademosun, A.O.; Akindahunsi, A.A.; Oyagbemi, A.A.; Omobowale, T.O.; Ajibade, T.O.; Adedapo, A.A. Cardio-Protective and Antioxidant Properties of Caffeic Acid and Chlorogenic Acid: Mechanistic Role of Angiotensin Converting Enzyme, Cholinesterase and Arginase Activities in Cyclosporine Induced Hypertensive Rats. Biomed. Pharmacother. 2019, 109, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Silva Rosario, A.C.R.; da Silva, A.H.M.; Silva, A.G.B.; Vieira, A.P.O.; Monteiro, M.C. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [CrossRef] [PubMed]
- Padda, M.; Picha, D. Quantification of Phenolic Acids and Antioxidant Activity in Sweetpotato Genotypes. Sci. Hortic. Sci. Hort Amst. 2008, 119, 17–20. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of Nitric Oxide in Inflammatory Diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Brezani, V.; Smejkal, K.; Hosek, J.; Tomasova, V. Anti-Inflammatory Natural Prenylated Phenolic Compounds—Potential Lead Substances. Curr. Med. Chem. 2018, 25, 1094–1159. [Google Scholar] [CrossRef]
- Bouhlali, E.; Dine, T.; Hmidani, A.; Bourkhis, B.; Khouya, T.; Ramchoun, M.; Filali-Zegzouti, Y.; Alem, C. Phenolic Profile and Anti-Inflammatory Activity of Four Moroccan Date (Phoenix dactylifera L.) Seed Varieties. Heliyon 2020, 6, e03436. [Google Scholar] [CrossRef]
- Yoon, J.H.; Baek, S.J. Molecular Targets of Dietary Polyphenols with Anti-Inflammatory Properties. Yonsei Med. J. 2005, 46, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Karunaweera, N.; Raju, R.; Gyengesi, E.; Münch, G. Plant Polyphenols as Inhibitors of NF-ΚB Induced Cytokine Production—A Potential Anti-Inflammatory Treatment for Alzheimer’s Disease? Front. Mol. Neurosci. 2015, 8, 24. [Google Scholar] [CrossRef] [PubMed]
- Potapovich, A.I.; Lulli, D.; Fidanza, P.; Kostyuk, V.A.; De Luca, C.; Pastore, S.; Korkina, L.G. Plant Polyphenols Differentially Modulate Inflammatory Responses of Human Keratinocytes by Interfering with Activation of Transcription Factors NFκB and AhR and EGFR–ERK Pathway. Toxicol. Appl. Pharmacol. 2011, 255, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Aruna, R.; Geetha, A.; Suguna, P. Rutin Modulates ASC Expression in NLRP3 Inflammasome: A Study in Alcohol and Cerulein-Induced Rat Model of Pancreatitis. Mol. Cell Biochem. 2014, 396, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, G.; Gurley, E.C.; Zhou, H. Flavonoid Apigenin Inhibits Lipopolysaccharide-Induced Inflammatory Response through Multiple Mechanisms in Macrophages. PLoS ONE 2014, 9, e107072. [Google Scholar] [CrossRef] [PubMed]
- Okai, Y.; Higashi-Okai, K. Possible Immunomodulating Activities of Carotenoids in in Vitro Cell Culture Experiments. Int. J. Immunopharmacol. 1996, 18, 753–758. [Google Scholar] [CrossRef]
- Kawata, A.; Murakami, Y.; Suzuki, S.; Fujisawa, S. Anti-Inflammatory Activity of β-Carotene, Lycopene and Tri-n-Butylborane, a Scavenger of Reactive Oxygen Species. In Vivo 2018, 32, 255–264. [Google Scholar] [CrossRef]
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic Effects of Inflammation on Vitamin A and Carotenoids in Humans and Animal Models. Adv. Nutr. 2017, 8, 197–212. [Google Scholar] [CrossRef]
- Bae, J.-Y.; Park, W.-S.; Kim, H.-J.; Kim, H.-S.; Kang, K.-K.; Kwak, S.-S.; Ahn, M.-J. Protective Effect of Carotenoid Extract from Orange-Fleshed Sweet Potato on Gastric Ulcer in Mice by Inhibition of NO, IL-6 and PGE2 Production. Pharmaceuticals 2021, 14, 1320. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Chemical Composition (%) of OFSP Flour | ||
|---|---|---|
| Yield (%) | 35.2 | |
| Moisture (% flour) | 8.4 | |
| Carbohydrates (% DM flour) | 42.7 ± 1.3 | |
| Total starch | 35.8 ± 1.8 | |
| Simple sugars | 6.9 ± 0.7 | |
| Amylose (% DM starch) | 5.1 ± 0.07 | |
| Resistant starch (% DM flour) | 2.2 ± 0.4 | |
| Total insoluble Fibers (% DM flour) | 6.2 ± 0.1 | |
| Hemicellulose | 1.75 ± 0.01 | |
| Cellulose | 4.24 ± 0.02 | |
| Lignin | 0.21 ± 0.01 | |
| Glycemic index Maximum glycemia | 71 122.16 ± 2.48 mg/dL | |
| Potentially Bioactive Micronutrient Composition of Orange Sweet Potato Flour | |||
|---|---|---|---|
| Sample | TPC in mg GAE/g EDW | Minerals Matters (% DM/g‧100 g−1) | beta-Carotene (% DM/mg‧100 g−1) |
| OFSP | 38.89 ± 0.52 | 5.93 ± 0.31 | 21.81 ± 2.20 |
| Antioxydant Activity | ||
|---|---|---|
| Assays | ||
| NO scavenging | Inhibition (%) at 100 µg/mL | 21.67 ± 2.43 a |
| Inhibition (%) at 75 µg/mL | 14.75 ± 2.55 b | |
| Inhibition (%) at 50 µg/mL | 4.97 ± 1.80 c | |
| DPPH | µmol TE/g EDW | 56.95 ± 1.73 |
| Inhibition (%) at 1 mg/mL | 39.80 ± 1.18 | |
| ORAC | µmol TE/g EDW | 988.66 ± 27.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boukhers, I.; Morel, S.; Kongolo, J.; Domingo, R.; Servent, A.; Ollier, L.; Kodja, H.; Petit, T.; Poucheret, P. Immunomodulatory and Antioxidant Properties of Ipomoea batatas Flour and Extracts Obtained by Green Extraction. Curr. Issues Mol. Biol. 2023, 45, 6967-6985. https://doi.org/10.3390/cimb45090440
Boukhers I, Morel S, Kongolo J, Domingo R, Servent A, Ollier L, Kodja H, Petit T, Poucheret P. Immunomodulatory and Antioxidant Properties of Ipomoea batatas Flour and Extracts Obtained by Green Extraction. Current Issues in Molecular Biology. 2023; 45(9):6967-6985. https://doi.org/10.3390/cimb45090440
Chicago/Turabian StyleBoukhers, Imane, Sylvie Morel, Joelle Kongolo, Romain Domingo, Adrien Servent, Lea Ollier, Hippolyte Kodja, Thomas Petit, and Patrick Poucheret. 2023. "Immunomodulatory and Antioxidant Properties of Ipomoea batatas Flour and Extracts Obtained by Green Extraction" Current Issues in Molecular Biology 45, no. 9: 6967-6985. https://doi.org/10.3390/cimb45090440
APA StyleBoukhers, I., Morel, S., Kongolo, J., Domingo, R., Servent, A., Ollier, L., Kodja, H., Petit, T., & Poucheret, P. (2023). Immunomodulatory and Antioxidant Properties of Ipomoea batatas Flour and Extracts Obtained by Green Extraction. Current Issues in Molecular Biology, 45(9), 6967-6985. https://doi.org/10.3390/cimb45090440