
Circular RNAs Variously Participate in Coronary Atherogenesis

 ,
,
Abstract
1. Introduction
2. Characteristics and Functions of circRNAs
3. Biogenesis and Degradation of circRNAs
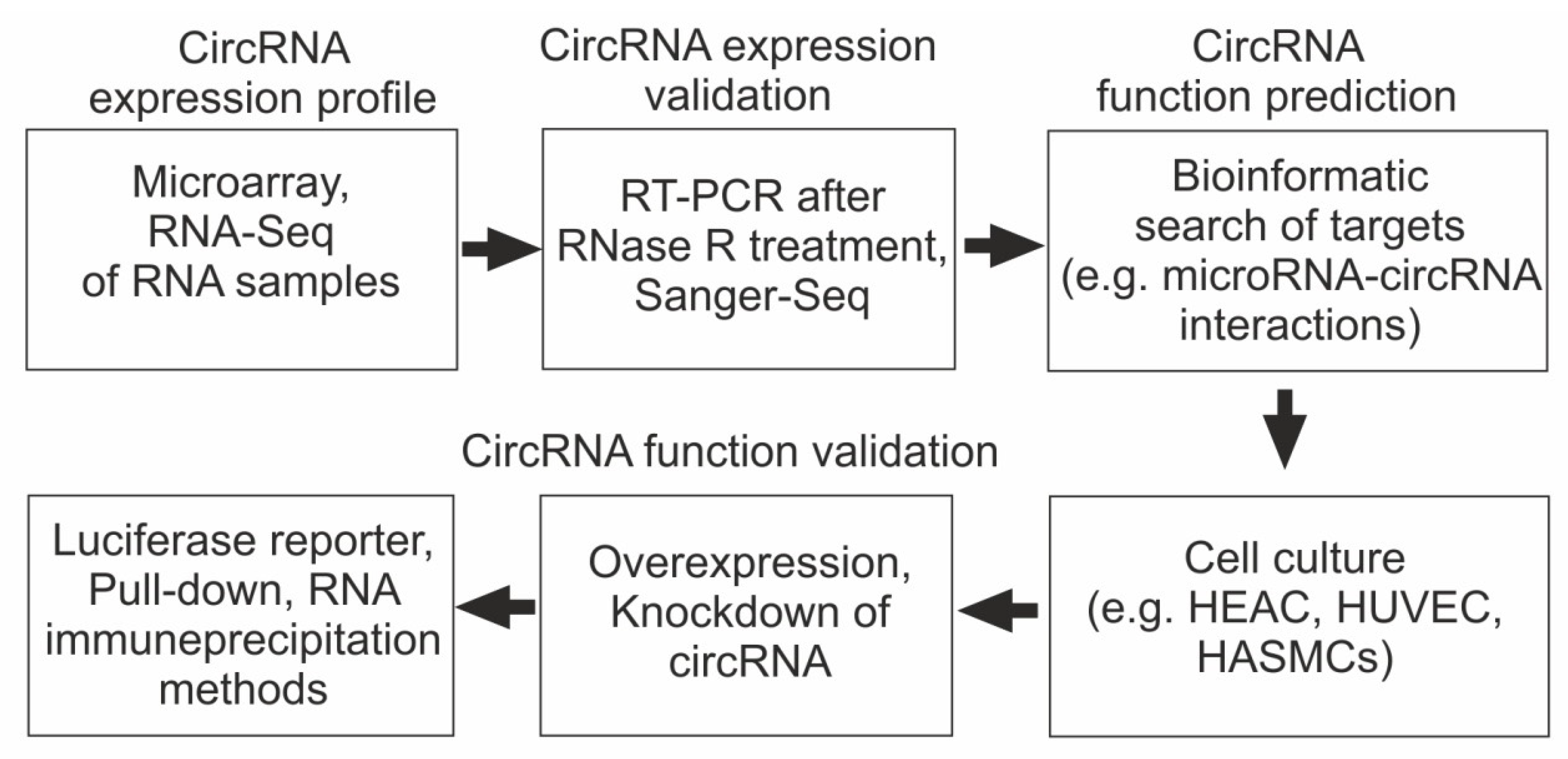
4. Main Approaches for Studying the Role of circRNA in the Pathogenesis of CAD
5. The Role of circRNAs in the Pathogenesis of CAD and Atherogenesis
6. CircRNAs Are Promising Biomarkers and Therapeutic Targets for the Treatment of CAD
7. Conclusions
8. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi, K.; Takeya, M.; Sakashita, N. Multifunctional Roles of Macrophages in the Development and Progression of Atherosclerosis in Humans and Experimental Animals. Med. Electron. Microsc. 2002, 35, 179–203. [Google Scholar] [CrossRef]
- Rudijanto, A. The Role of Vascular Smooth Muscle Cells on the Pathogenesis of Atherosclerosis. Acta Med. Indones. 2007, 39, 86–93. [Google Scholar]
- Baumer, Y.; McCurdy, S.; Alcala, M.; Mehta, N.; Lee, B.-H.; Ginsberg, M.H.; Boisvert, W.A. CD98 Regulates Vascular Smooth Muscle Cell Proliferation in Atherosclerosis. Atherosclerosis 2017, 256, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Gholipour, M.; Taheri, M. The Emerging Role of Long Non-Coding RNAs and Circular RNAs in Coronary Artery Disease. Front. Cardiovasc. Med. 2021, 8, 632393. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin Signature Reveals over a Thousand Highly Conserved Large Non-Coding RNAs in Mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many Human Large Intergenic Noncoding RNAs Associate with Chromatin-Modifying Complexes and Affect Gene Expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Lai, E.C. Endogenous Small Interfering RNAs in Animals. Nat. Rev. Mol. Cell Biol. 2008, 9, 673–678. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92. [Google Scholar] [CrossRef] [PubMed]
- Rozhkova, A.V.; Dmitrieva, V.G.; Nosova, E.V.; Dergunov, A.D.; Limborska, S.A.; Dergunova, L.V. Genomic Variants and Multilevel Regulation of ABCA1, ABCG1, and SCARB1 Expression in Atherogenesis. J. Cardiovasc. Dev. Dis. 2021, 8, 170. [Google Scholar] [CrossRef]
- Sidorkiewicz, M. Is MicroRNA-33 an Appropriate Target in the Treatment of Atherosclerosis? Nutrients 2023, 15, 902. [Google Scholar] [CrossRef]
- Tong, K.-L.; Tan, K.-E.; Lim, Y.-Y.; Tien, X.-Y.; Wong, P.-F. CircRNA-MiRNA Interactions in Atherogenesis. Mol. Cell. Biochem. 2022, 477, 2703–2733. [Google Scholar] [CrossRef]
- Cheng, C.; Wang, Y.; Xue, Q.; Huang, Y.; Wang, X.; Liao, F.; Miao, C. CircRnas in Atherosclerosis, with Special Emphasis on the Spongy Effect of CircRnas on MiRnas. Cell Cycle 2023, 22, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Li, Y.; Wu, Q.; Huang, L.; Xu, J.; Zeng, Q. Pathogenic Role of MicroRNAs in Atherosclerotic Ischemic Stroke: Implications for Diagnosis and Therapy. Genes. Dis. 2022, 9, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tussy, P.; Ruz-Maldonado, I.; Fernández-Hernando, C. MicroRNAs and Circular RNAs in Lipoprotein Metabolism. Curr. Atheroscler. Rep. 2021, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef]
- Vidigal, J.A.; Ventura, A. The Biological Functions of MiRNAs: Lessons from in Vivo Studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef]
- Sallam, T.; Jones, M.; Thomas, B.J.; Wu, X.; Gilliland, T.; Qian, K.; Eskin, A.; Casero, D.; Zhang, Z.; Sandhu, J.; et al. Transcriptional Regulation of Macrophage Cholesterol Efflux and Atherogenesis by a Long Noncoding RNA. Nat. Med. 2018, 24, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-D.; Yao, H.-H.; Wang, L.-M.; Yu, M.; Shi, S.; Yuan, Z.-X.; Liu, J. Knockdown of GAS5 Inhibits Atherosclerosis Progression via Reducing EZH2-Mediated ABCA1 Transcription in ApoE-/- Mice. Mol. Ther. Nucleic Acids 2020, 19, 84–96. [Google Scholar] [CrossRef]
- Lan, X.; Yan, J.; Ren, J.; Zhong, B.; Li, J.; Li, Y.; Liu, L.; Yi, J.; Sun, Q.; Yang, X.; et al. A Novel Long Noncoding RNA Lnc-HC Binds HnRNPA2B1 to Regulate Expressions of Cyp7a1 and Abca1 in Hepatocytic Cholesterol Metabolism. Hepatology 2016, 64, 58–72. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 159. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The Multilayered Complexity of CeRNA Crosstalk and Competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Wang, L.; Meng, X.; Li, G.; Zhou, Q.; Xiao, J. Circular RNAs in Cardiovascular Diseases. Adv. Exp. Med. Biol. 2018, 1087, 191–204. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.; Hu, L.; Liu, Y.; Zhou, Q.; Wang, M.; An, Y.; Li, P. Role of Circular RNAs in the Pathogenesis of Cardiovascular Disease. J. Cardiovasc. Transl. Res. 2020, 13, 572–583. [Google Scholar] [CrossRef]
- Sun, J.Y.; Shi, Y.; Cai, X.Y.; Liu, J. Potential Diagnostic and Therapeutic Value of Circular RNAs in Cardiovascular Diseases. Cell. Signal 2020, 71, 109604. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sharpless, N.E. Detecting and Characterizing Circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Panda, A.C. Circular RNAs Act as MiRNA Sponges. Adv. Exp. Med. Biol. 2018, 1087, 67–79. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA Circles Function as Efficient MicroRNA Sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Joaquim, V.H.A.; Pereira, N.P.; Fernandes, T.; Oliveira, E.M. Circular RNAs as a Diagnostic and Therapeutic Target in Cardiovascular Diseases. Int. J. Mol. Sci. 2023, 24, 2125. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Lv, X. Revealing New Landscape of Cardiovascular Disease through Circular RNA-MiRNA-MRNA Axis. Genomics 2020, 112, 1680–1685. [Google Scholar] [CrossRef]
- Wei, W.; Tang, M.; Wang, Q.; Li, X. Circ_HECW2 Regulates Ox-LDL-Induced Dysfunction of Cardiovascular Endothelial Cells by MiR-942-5p/TLR4 Axis. Clin. Hemorheol. Microcirc. 2022, preprint. [Google Scholar] [CrossRef] [PubMed]
- Diener, T.O. Potato Spindle Tuber “Virus”. IV. A Replicating, Low Molecular Weight RNA. Virology 1971, 45, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids Are Single-Stranded Covalently Closed Circular RNA Molecules Existing as Highly Base-Paired Rod-like Structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef]
- HSU, M.-T.; COCA-PRADOS, M. Electron Microscopic Evidence for the Circular Form of RNA in the Cytoplasm of Eukaryotic Cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs Are Abundant, Conserved, and Associated with ALU Repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Jakobsen, T.; Hager, H.; Kjems, J. The Emerging Roles of CircRNAs in Cancer and Oncology. Nat. Rev. Clin. Oncol. 2022, 19, 188–206. [Google Scholar] [CrossRef] [PubMed]
- Filippenkov, I.B.; Kalinichenko, E.O.; Limborska, S.A.; Dergunova, L.V. Circular RNAs-One of the Enigmas of the Brain. Neurogenetics 2017, 18, 1–6. [Google Scholar] [CrossRef]
- Bagheri Moghaddam, M.; Maleki, M.; Oveisee, M.; Bagheri Moghaddam, M.; Arabian, M.; Malakootian, M. Circular RNAs: New Players in Cardiomyopathy. Genes 2022, 13, 1537. [Google Scholar] [CrossRef]
- Farina, F.M.; Weber, C.; Santovito, D. The Emerging Landscape of Non-Conventional RNA Functions in Atherosclerosis. Atherosclerosis 2023, 374, 74–86. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-Intron Circular RNAs Regulate Transcription in the Nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Müller, S.; Ebermann, C.; Schnarr, T. Recent Advances in Understanding Circular RNAs. F1000Research 2020, 9, 655. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, L.; Fan, K.; Cheng, Z.-X.; Sun, Q.-C.; Wang, J.-J. Circular RNA-ITCH Suppresses Lung Cancer Proliferation via Inhibiting the Wnt/β-Catenin Pathway. Biomed. Res. Int. 2016, 2016, 1579490. [Google Scholar] [CrossRef]
- Armakola, M.; Higgins, M.J.; Figley, M.D.; Barmada, S.J.; Scarborough, E.A.; Diaz, Z.; Fang, X.; Shorter, J.; Krogan, N.J.; Finkbeiner, S.; et al. Inhibition of RNA Lariat Debranching Enzyme Suppresses TDP-43 Toxicity in ALS Disease Models. Nat. Genet. 2012, 44, 1302–1309. [Google Scholar] [CrossRef]
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA Sponges: Competitive Inhibitors of Small RNAs in Mammalian Cells. Nat. Methods 2007, 4, 721–726. [Google Scholar] [CrossRef]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA Biology from an Atlas of Mammalian MRNA-Binding Proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Tian, M.; Cao, N.; Yang, P.; Xu, Z.; Zheng, S.; Liao, Q.; Chen, C.; Zeng, C.; Jose, P.A.; et al. Circular RNA CircEsyt2 Regulates Vascular Smooth Muscle Cell Remodeling via Splicing Regulation. J. Clin. Invest. 2021, 131, e147031. [Google Scholar] [CrossRef]
- Zhou, H.; Gan, X.; He, S.; Wang, Y.; Zhang, S.; Chen, J.; Zhou, Y.; Hou, C.; Hua, L.; Zhang, Q.; et al. Identification of Circular RNA BTBD7_hsa_circ_0000563 as a Novel Biomarker for Coronary Artery Disease and the Functional Discovery of BTBD7_hsa_circ_0000563 Based on Peripheral Blood Mononuclear Cells: A Case Control Study. Clin. Proteom. 2022, 19, 37. [Google Scholar] [CrossRef]
- Prats, A.-C.; David, F.; Diallo, L.H.; Roussel, E.; Tatin, F.; Garmy-Susini, B.; Lacazette, E. Circular RNA, the Key for Translation. Int. J. Mol. Sci. 2020, 21, 8591. [Google Scholar] [CrossRef] [PubMed]
- van Rossum, D.; Verheijen, B.M.; Pasterkamp, R.J. Circular RNAs: Novel Regulators of Neuronal Development. Front. Mol. Neurosci. 2016, 9, 74. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular Intronic Long Noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Kelly, S.; Greenman, C.; Cook, P.R.; Papantonis, A. Exon Skipping Is Correlated with Exon Circularization. J. Mol. Biol. 2015, 427, 2414–2417. [Google Scholar] [CrossRef]
- Yao, T.; Chen, Q.; Fu, L.; Guo, J. Circular RNAs: Biogenesis, Properties, Roles, and Their Relationships with Liver Diseases. Hepatol. Res. 2017, 47, 497–504. [Google Scholar] [CrossRef]
- Ren, L.; Jiang, Q.; Mo, L.; Tan, L.; Dong, Q.; Meng, L.; Yang, N.; Li, G. Mechanisms of Circular RNA Degradation. Commun. Biol. 2022, 5, 1355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary Sequence-Mediated Exon Circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Wilusz, J.E. Short Intronic Repeat Sequences Facilitate Circular RNA Production. Genes. Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. CircRNA Biogenesis Competes with Pre-MRNA Splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Montañés-Agudo, P.; van der Made, I.; Aufiero, S.; Tijsen, A.J.; Pinto, Y.M.; Creemers, E.E. Quaking Regulates Circular RNA Production in Cardiomyocytes. J. Cell Sci. 2023, 136, jcs261120. [Google Scholar] [CrossRef]
- Errichelli, L.; Dini Modigliani, S.; Laneve, P.; Colantoni, A.; Legnini, I.; Capauto, D.; Rosa, A.; De Santis, R.; Scarfò, R.; Peruzzi, G.; et al. FUS Affects Circular RNA Expression in Murine Embryonic Stem Cell-Derived Motor Neurons. Nat. Commun. 2017, 8, 14741. [Google Scholar] [CrossRef]
- Dong, W.; Dai, Z.H.; Liu, F.C.; Guo, X.G.; Ge, C.M.; Ding, J.; Liu, H.; Yang, F. The RNA-Binding Protein RBM3 Promotes Cell Proliferation in Hepatocellular Carcinoma by Regulating Circular RNA SCD-CircRNA 2 Production. EBioMedicine 2019, 45, 155–167. [Google Scholar] [CrossRef]
- Hsiao, K.Y.; Sun, H.S.; Tsai, S.J. Circular RNA—New Member of Noncoding RNA with Novel Functions. Exp. Biol. Med. 2017, 242, 1136–1141. [Google Scholar] [CrossRef]
- Liang, D.; Tatomer, D.C.; Luo, Z.; Wu, H.; Yang, L.; Chen, L.L.; Cherry, S.; Wilusz, J.E. The Output of Protein-Coding Genes Shifts to Circular RNAs When the Pre-MRNA Processing Machinery Is Limiting. Mol. Cell 2017, 68, 940–954.e3. [Google Scholar] [CrossRef] [PubMed]
- Kokot, K.E.; Kneuer, J.M.; John, D.; Rebs, S.; Möbius-Winkler, M.N.; Erbe, S.; Müller, M.; Andritschke, M.; Gaul, S.; Sheikh, B.N.; et al. Reduction of A-to-I RNA Editing in the Failing Human Heart Regulates Formation of Circular RNAs. Basic Res. Cardiol. 2022, 117, 32. [Google Scholar] [CrossRef]
- Aktaş, T.; Ilik, I.A.; Maticzka, D.; Bhardwaj, V.; Pessoa Rodrigues, C.; Mittler, G.; Manke, T.; Backofen, R.; Akhtar, A. DHX9 Suppresses RNA Processing Defects Originating from the Alu Invasion of the Human Genome. Nature 2017, 544, 115–119. [Google Scholar] [CrossRef]
- Eleazer, R.; De Silva, K.; Andreeva, K.; Jenkins, Z.; Osmani, N.; Rouchka, E.C.; Fondufe-Mittendorf, Y. PARP1 Regulates Circular RNA Biogenesis Though Control of Transcriptional Dynamics. Cells 2023, 12, 1160. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.L.; Lei, Y.N.; Liu, X.Q.; Xue, W.; Zhang, Y.; Nan, F.; Gao, X.; Zhang, J.; Wei, J.; et al. Linking Circular Intronic RNA Degradation and Function in Transcription by RNase H1. Sci. China Life Sci. 2021, 64, 1795–1809. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.M. Circular RNAs and RNase L in PKR Activation and Virus Infection. Cell Biosci. 2019, 9, 43. [Google Scholar] [CrossRef]
- Liu, C.X.; Li, X.; Nan, F.; Jiang, S.; Gao, X.; Guo, S.K.; Xue, W.; Cui, Y.; Dong, K.; Ding, H.; et al. Structure and Degradation of Circular RNAs Regulate PKR Activation in Innate Immunity. Cell 2019, 177, 865–880.e21. [Google Scholar] [CrossRef]
- Park, O.H.; Ha, H.; Lee, Y.; Boo, S.H.; Kwon, D.H.; Song, H.K.; Kim, Y.K. Endoribonucleolytic Cleavage of M6A-Containing RNAs by RNase P/MRP Complex. Mol. Cell 2019, 74, 494–507.e8. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.W.; Busa, V.F.; Shao, Y.; Leung, A.K.L. Structure-Mediated RNA Decay by UPF1 and G3BP1. Mol. Cell 2020, 78, 70–84.e6. [Google Scholar] [CrossRef]
- Piwecka, M.; Glažar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Jara, C.A.C.; Fenske, P.; et al. Loss of a Mammalian Circular RNA Locus Causes MiRNA Deregulation and Affects Brain Function. Science 2017, 357, eaam8526. [Google Scholar] [CrossRef]
- Zhou, M.; Xiao, M.S.; Li, Z.; Huang, C. New Progresses of Circular RNA Biology: From Nuclear Export to Degradation. RNA Biol. 2021, 18, 1365–1373. [Google Scholar] [CrossRef]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded Identification and Characterization of Mammalian Circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef]
- Xu, J.-L.; Xu, W.-X.; Tang, J.-H. Exosomal CircRNAs: A New Communication Method in Cancer. Am. J. Transl. Res. 2021, 13, 12913. [Google Scholar]
- Lasda, E.; Parker, R. Circular RNAs Co-Precipitate with Extracellular Vesicles: A Possible Mechanism for CircRNA Clearance. PLoS ONE 2016, 11, e0148407. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Song, J.; Huang, X.; Pan, Z.; Goldbrunner, R.; Stavrinou, L.; Lin, S.; Hu, W.; Zheng, F.; Stavrinou, P. Exosomes Derived From Mesenchymal Stem Cells: Novel Effects in the Treatment of Ischemic Stroke. Front. Neurosci. 2022, 16, 899887. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.F.; Chen, J.X.; He, S.; Zhou, Y.Q.; Hua, L.; Hou, C.; Zhang, S.; Gan, X.K.; Wang, Y.J.; Zhou, H.X.; et al. Characteristics of Circular Rnas Expression of Peripheral Blood Mononuclear Cells in Humans with Coronary Artery Disease. Physiol. Genom. 2021, 53, 349–357. [Google Scholar] [CrossRef]
- Tan, W.L.W.; Lim, B.T.S.; Anene-Nzelu, C.G.O.; Ackers-Johnson, M.; Dashi, A.; See, K.; Tiang, Z.; Lee, D.P.; Chua, W.W.; Luu, T.D.A.; et al. A Landscape of Circular RNA Expression in the Human Heart. Cardiovasc. Res. 2017, 113, 298–309. [Google Scholar] [CrossRef]
- Wei, L.; Yu, Z.; Liu, L.; Zhou, Y.; Bai, X.; Wang, L.; Bai, M.; Sun, S. Integrated Analysis of the CircRNA-Based CeRNA Network in Renal Fibrosis Induced by Ischemia Reperfusion Injury. Front. Genet. 2022, 12, 793182. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in Human Diseases and Potential Use as Biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.B.; Lavenniah, A.; Foo, R.S.Y. Circles in the Heart and Cardiovascular System. Cardiovasc. Res. 2020, 116, 269–278. [Google Scholar] [CrossRef]
- Zhang, C.; Ge, S.; Gong, W.; Xu, J.; Guo, Z.; Liu, Z.; Gao, X.; Wei, X.; Ge, S. LncRNA ANRIL Acts as a Modular Scaffold of WDR5 and HDAC3 Complexes and Promotes Alteration of the Vascular Smooth Muscle Cell Phenotype. Cell Death Dis. 2020, 11, 435. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Sun, L.; Zhang, J.; Zhang, L.; Liu, X. Oxygenized Low-Density Lipoprotein-Induced ASMC Dysregulation Depends on Circ_0000345-Mediated Regulatory Mechanism. J. Atheroscler. Thromb. 2022, 29, 63327. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, P.; Chen, X.; Yang, W. Circ_CHMP5 Aggravates Oxidized Low-Density Lipoprotein-Induced Damage to Human Umbilical Vein Endothelial Cells through MiR-516b-5p/TGFβR2 Axis. Clin. Hemorheol. Microcirc. 2023, preprint. [Google Scholar] [CrossRef]
- Lin, H.; Gao, D.; Wang, S.; Wang, Z.; Guan, H.; Wang, Y.; Zhou, Y. Inhibition of Circ_0000231 Suppresses Oxidized Low Density Lipoprotein-Induced Apoptosis, Autophagy and Inflammation in Human Umbilical Vein Endothelial Cells by Regulating MiR-590-5p/PDCD4 Axis. Clin. Hemorheol. Microcirc. 2023, preprint. [Google Scholar] [CrossRef]
- Moore, K.J.; Tabas, I. Macrophages in the Pathogenesis of Atherosclerosis. Cell 2011, 145, 341–355. [Google Scholar] [CrossRef]
- Hansson, G.K.; Hermansson, A. The Immune System in Atherosclerosis. Nat. Immunol. 2011, 12, 204–212. [Google Scholar] [CrossRef]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Zernecke, A.; Weber, C. Chemokines in the Vascular Inflammatory Response of Atherosclerosis. Cardiovasc. Res. 2010, 86, 192–201. [Google Scholar] [CrossRef]
- Brunetti, N.D.; Salvemini, G.; Cuculo, A.; Ruggiero, A.; De Gennaro, L.; Gaglione, A.; Di Biase, M. Coronary Artery Ectasia Is Related to Coronary Slow Flow and Inflammatory Activation. Atherosclerosis 2014, 233, 636–640. [Google Scholar] [CrossRef]
- Shen, L.; Hu, Y.; Lou, J.; Yin, S.; Wang, W.; Wang, Y.; Xia, Y.; Wu, W. CircRNA-0044073 Is Upregulated in Atherosclerosis and Increases the Proliferation and Invasion of Cells by Targeting MiR-107. Mol. Med. Rep. 2019, 19, 3923–3932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, W.; Wu, X.; Zhou, X. Regulatory Roles of Circular RNAs in Coronary Artery Disease. Mol. Ther. Nucleic Acids 2020, 21, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Lv, B.; Jiang, S.; Lin, J. The Landscape of Circular RNAs in Cardiovascular Diseases. Int. J. Mol. Sci. 2023, 24, 4571. [Google Scholar] [CrossRef]
- Pan, R.-Y.; Liu, P.; Zhou, H.-T.; Sun, W.-X.; Song, J.; Shu, J.; Cui, G.-J.; Yang, Z.-J.; Jia, E.-Z. Circular RNAs Promote TRPM3 Expression by Inhibiting Hsa-MiR-130a-3p in Coronary Artery Disease Patients. Oncotarget 2017, 8, 60280–60290. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.-Y.; Zhao, C.-H.; Yuan, J.-X.; Zhang, Y.-J.; Jin, J.-L.; Gu, M.-F.; Mao, Z.-Y.; Sun, H.-J.; Jia, Q.-W.; Ji, M.-Y.; et al. Circular RNA Profile in Coronary Artery Disease. Am. J. Transl. Res. 2019, 11, 7115. [Google Scholar]
- Hou, C.; Gu, L.; Guo, Y.; Zhou, Y.; Hua, L.; Chen, J.; He, S.; Zhang, S.; Jia, Q.; Zhao, C.; et al. Association between Circular RNA Expression Content and Severity of Coronary Atherosclerosis in Human Coronary Artery. J. Clin. Lab. Anal. 2020, 34, e23552. [Google Scholar] [CrossRef]
- Yang, L.; Chen, W.; Li, B.; Hu, Y.; Lu, H.; Zhang, P.; Yang, H.; Zhang, M.; Pan, D. Circular RNA Circ_0026218 Suppressed Atherosclerosis Progression via MiR-338-3p/SIRT6 Axis. Biomed. Res. Int. 2023, 2023, 5647758. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Ma, W.; Song, B.; Wang, S.; Li, X.; Wang, Z. Hsa_circ_0030042 Ameliorates Oxidized Low-Density Lipoprotein-Induced Endothelial Cell Injury via the MiR-616-3p/RFX7 Axis. Int. Heart J. 2022, 63, 22–065. [Google Scholar] [CrossRef]
- Zhang, W.-B.; Qi, Y.-F.; Xiao, Z.-X.; Chen, H.; Liu, S.-H.; Li, Z.-Z.; Zeng, Z.-F.; Wu, H.-F. CircHIPK3 Regulates Vascular Smooth Muscle Cell Calcification Via the MiR-106a-5p/MFN2 Axis. J. Cardiovasc. Transl. Res. 2022, 15, 1315–1326. [Google Scholar] [CrossRef]
- WEI, M.Y.; LV, R.R.; TENG, Z. Circular RNA CircHIPK3 as a Novel CircRNA Regulator of Autophagy and Endothelial Cell Dysfunction in Atherosclerosis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12849–12858. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Jia, H.; Huang, B.; Lu, S.; Chen, Z.; Shen, J.; Zou, Y.; Wang, C.; Sun, Y. Identification of Differently Expressed MRNAs in Atherosclerosis Reveals CDK6 Is Regulated by CircHIPK3/MiR-637 Axis and Promotes Cell Growth in Human Vascular Smooth Muscle Cells. Front. Genet. 2021, 12, 596169. [Google Scholar] [CrossRef]
- Ji, N.; Wang, Y.; Gong, X.; Ni, S.; Zhang, H. CircMTO1 Inhibits Ox-LDL-Stimulated Vascular Smooth Muscle Cell Proliferation and Migration via Regulating the MiR-182-5p/RASA1 Axis. Mol. Med. 2021, 27, 73. [Google Scholar] [CrossRef]
- Wei, Z.; Ran, H.; Yang, C. CircRSF1 Contributes to Endothelial Cell Growth, Migration and Tube Formation under Ox-LDL Stress through Regulating MiR-758/CCND2 Axis. Life Sci. 2020, 259, 118241. [Google Scholar] [CrossRef] [PubMed]
- Tiliwaldi, H.; Tursun, A.; Tohti, A.; Mamatzunun, M.; Wu, Z. Circ_0000345 Protects Endothelial Cells From Oxidized Low-Density Lipoprotein-Induced Injury by MiR-129-5p/Ten-Eleven Translocation Axis. J. Cardiovasc. Pharmacol. 2021, 77, 603–613. [Google Scholar] [CrossRef]
- Yuan, R.; Xin, Q.; Ma, X.; Yu, M.; Miao, Y.; Chen, K.; Cong, W. Identification of a Novel Angiogenesis Signalling CircSCRG1/MiR-1268b/NR4A1 Pathway in Atherosclerosis and the Regulatory Effects of TMP-PF In Vitro. Molecules 2023, 28, 1271. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Lu, Z.; Lai, C. Circ_0093887 Regulated Ox-LDL Induced Human Aortic Endothelial Cells Viability, Apoptosis, and Inflammation through Modulating MiR-758-3p/BAMBI Axis in Atherosclerosis. Clin. Hemorheol. Microcirc. 2022, 81, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, G.; Fan, S.; Wang, Y.; Wei, H.; Li, M.; Li, X. Circ_0093887 Upregulates CCND2 and SUCNR1 to Inhibit the Ox-LDL-Induced Endothelial Dysfunction in Atherosclerosis by Functioning as a MiR-876-3p Sponge. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Yu, Y.; Wang, L.; Dou, Y.-Q.; Zhang, X.-H.; Cui, Y.; Wang, H.-Y.; Yong, Y.-T.; Liu, Y.-B.; Hu, H.-J.; et al. Circ-Sirt1 Controls NF-ΚB Activation via Sequence-Specific Interaction and Enhancement of SIRT1 Expression by Binding to MiR-132/212 in Vascular Smooth Muscle Cells. Nucleic Acids Res. 2019, 47, 3580–3593. [Google Scholar] [CrossRef]
- Yang, Z.; Liang, X.; Yang, L. Circular RNA Circ_0001445 Alleviates the Ox-LDL-Induced Endothelial Injury in Human Primary Aortic Endothelial Cells through Regulating ABCG1 via Acting as a Sponge of MiR-208b-5p. Gen. Thorac. Cardiovasc. Surg. 2022, 70, 779–792. [Google Scholar] [CrossRef]
- Ryu, J.; Choe, N.; Kwon, D.-H.; Shin, S.; Lim, Y.-H.; Yoon, G.; Kim, J.H.; Kim, H.S.; Lee, I.-K.; Ahn, Y.; et al. Circular RNA CircSmoc1-2 Regulates Vascular Calcification by Acting as a MiR-874-3p Sponge in Vascular Smooth Muscle Cells. Mol. Ther. Nucleic Acids 2022, 27, 645–655. [Google Scholar] [CrossRef]
- Kou, L.; Yang, N.; Dong, B.; Yang, J.; Song, Y.; Li, Y.; Qin, Q. Circular RNA Testis-Expressed 14 Overexpression Induces Apoptosis and Suppresses Migration of Ox-LDL-Stimulated Vascular Smooth Muscle Cells via Regulating the MicroRNA 6509-3p/Thanatos-Associated Domain-Containing Apoptosis-Associated Protein 1 Axis. Bioengineered 2022, 13, 13150–13161. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, H.; Wang, Y. Circ_0003423 Alleviates Oxidized Low-Density Lipoprotein-Induced Endothelial Cell Injury by Sponging MiR-142-3p and Activating Sirtuin 3/Superoxide Dismutase 2 Pathway. J. Surg. Res. 2022, 277, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Vilades, D.; Martínez-Camblor, P.; Ferrero-Gregori, A.; Bär, C.; Lu, D.; Xiao, K.; Vea, À.; Nasarre, L.; Sanchez Vega, J.; Leta, R.; et al. Plasma Circular RNA Hsa_circ_0001445 and Coronary Artery Disease: Performance as a Biomarker. FASEB J. 2020, 34, 4403–4414. [Google Scholar] [CrossRef]
- Cai, Y.; Xu, L.; Xu, C.; Wang, Y.; Fan, C. Hsa_circ_0001445 Inhibits Ox-LDL-Induced HUVECs Inflammation, Oxidative Stress and Apoptosis by Regulating MiRNA-640. Perfusion 2022, 37, 86–94. [Google Scholar] [CrossRef]
- Dinh, P.; Peng, J.; Tran, T.; Wu, D.; Tran, C.; Dinh, T.; Pan, S. Identification of Hsa_circ_0001445 of a Novel CircRNA-MiRNA-MRNA Regulatory Network as Potential Biomarker for Coronary Heart Disease. Front. Cardiovasc. Med. 2023, 10, 1104223. [Google Scholar] [CrossRef]
- Li, R.; Jiang, Q.; Zheng, Y. Circ_0002984 Induces Proliferation, Migration and Inflammation Response of VSMCs Induced by Ox-LDL through MiR-326-3p/VAMP3 Axis in Atherosclerosis. J. Cell Mol. Med. 2021, 25, 8028–8038. [Google Scholar] [CrossRef]
- Wang, Y.; Pei, W.; Lu, P. Circ_ARHGAP32 Acts as MiR-665 Sponge to Upregulate FGF2 to Promote Ox-LDL Induced Vascular Smooth Muscle Cells Proliferation and Migration. Clin. Hemorheol. Microcirc. 2022, 82, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Shi, S.; Zhao, J.; Pan, Q.; Huang, C.; Shen, Q.; Liu, Z. Propofol Inhibits Cell Apoptosis and Inflammatory Response in Ox-LDL-Induced Human Umbilical Vein Endothelial Cells through the Modulation of the Circ_0003645/MiR-149-3p/TRAF7 Axis. Clin. Hemorheol. Microcirc. 2022, 84, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, L.; Ye, B.; Chen, W.; Zheng, G.; Xie, H.; Guo, Y. Knockdown of Hsa_circ_0005699 Attenuates Inflammation and Apoptosis Induced by Ox-LDL in Human Umbilical Vein Endothelial Cells through Regulation of the MiR-450b-5p/NFKB1 Axis. Mol. Med. Rep. 2022, 26, 290. [Google Scholar] [CrossRef]
- Geng, H.-H.; Li, R.; Su, Y.-M.; Xiao, J.; Pan, M.; Cai, X.-X.; Ji, X.-P. The Circular RNA Cdr1as Promotes Myocardial Infarction by Mediating the Regulation of MiR-7a on Its Target Genes Expression. PLoS ONE 2016, 11, e0151753. [Google Scholar] [CrossRef]
- Huang, S.; Zeng, Z.; Sun, Y.; Cai, Y.; Xu, X.; Li, H.; Wu, S. Association Study of Hsa_circ_0001946, Hsa-MiR-7-5p and PARP1 in Coronary Atherosclerotic Heart Disease. Int. J. Cardiol. 2021, 328, 1–7. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Yu, Z.; Zhang, T. Circ_0026218 Ameliorates Oxidized Low-Density Lipoprotein-Induced Vascular Endothelial Cell Dysfunction by Regulating MiR-188-3p/TLR4/NF-ΚB Pathway. Cardiovasc. Drugs Ther. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Li, Y.; Wang, B. Circular RNA CircCHFR Downregulation Protects against Oxidized Low-Density Lipoprotein-Induced Endothelial Injury via Regulation of MicroRNA-15b-5p/Growth Arrest and DNA Damage Inducible Gamma. Bioengineered 2022, 13, 4481–4492. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yang, F.; Zhao, H.; Wang, M.; Zhang, Y. Circular RNA CircCHFR Facilitates the Proliferation and Migration of Vascular Smooth Muscle via MiR-370/FOXO1/Cyclin D1 Pathway. Mol. Ther. Nucleic Acids 2019, 16, 434–441. [Google Scholar] [CrossRef]
- He, D.; Li, Z.; Chen, Y.; Huang, M. Circular RNA Circ_0029589 Promotes Ox-LDL-Induced Endothelial Cell Injury through Regulating RAB22A by Serving as a Sponge of MiR-1197. Clin. Hemorheol. Microcirc. 2023, 83, 359–376. [Google Scholar] [CrossRef]
- Li, X.; Kang, X.; Di, Y.; Sun, S.; Yang, L.; Wang, B.; Ji, Z. CircCHMP5 Contributes to Ox-LDL-Induced Endothelial Cell Injury Through the Regulation of MiR-532-5p/ROCK2 Axis. Cardiovasc. Drugs Ther. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Ni, Q.; Wang, H.; Wang, Y.; Yao, Y.; Li, Y.; Wang, W.; Yang, S.; Chen, J.; Lv, L.; et al. CircRNA CircCOL1A1 Acts as a Sponge of MiR-30a-5p to Promote Vascular Smooth Cell Phenotype Switch through Regulation of Smad1 Expression. Thromb. Haemost. 2023, 123, 097–107. [Google Scholar] [CrossRef]
- Wang, K.; Bai, X.; Mei, L.; Miao, Y.; Jin, F. CircRNA_0050486 Promotes Cell Apoptosis and Inflammation by Targeting MiR-1270 in Atherosclerosis. Ann. Transl. Med. 2022, 10, 905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, W.; Li, M. Role and Mechanism of Circular RNA Circ_0050486 in Regulating Oxidized Low-Density Lipoprotein-Induced Injury in Endothelial Cells. Clin. Hemorheol. Microcirc. 2022, 82, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Gao, B.; Wang, Y. CircIRAK1 Aggravates Ox-LDL-Induced Endothelial Cell Injury in Atherosclerosis via TRIM14 Upregulation by Binding to MiR-330-5p1. Clin. Hemorheol. Microcirc. 2022, preprint. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhao, P.; Meng, X.; Jin, H.; Tuo, B. Circ_0091822 Aggravates Ox-LDL-Induced Endothelial Cell Injury through Targeting the MiR-661/RAB22A Axis. Clin. Hemorheol. Microcirc. 2022, 83, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, Z.; Peng, X.; Zheng, Z.; Le, A.; Guo, J.; Ma, L.; Shi, H.; Yao, K.; Zhang, S.; et al. Extracellular Vesicle-Derived CircITGB1 Regulates Dendritic Cell Maturation and Cardiac Inflammation via MiR-342-3p/NFAM1. Oxid. Med. Cell. Longev. 2022, 2022, 8392313. [Google Scholar] [CrossRef]
- Li, F.; Chen, Y.; He, Z.; Wang, C.; Wang, X.; Pan, G.; Peng, J.Y.; Chen, Q.; Wang, X. Hsa_circ_0001879 Promotes the Progression of Atherosclerosis by Regulating the Proliferation and Migration of Oxidation of Low Density Lipoprotein (Ox-LDL)-Induced Vascular Endothelial Cells via the MiR-6873-5p-HDAC9 Axis. Bioengineered 2021, 12, 10420–10429. [Google Scholar] [CrossRef]
- Xiong, F.; Mao, R.; Zhang, L.; Zhao, R.; Tan, K.; Liu, C.; Xu, J.; Du, G.; Zhang, T. CircNPHP4 in Monocyte-Derived Small Extracellular Vesicles Controls Heterogeneous Adhesion in Coronary Heart Atherosclerotic Disease. Cell Death Dis. 2021, 12, 948. [Google Scholar] [CrossRef]
- Jing, B.; Hui, Z. Circular RNA_0033596 Aggravates Endothelial Cell Injury Induced by Oxidized Low-Density Lipoprotein via MicroRNA-217-5p /Chloride Intracellular Channel 4 Axis. Bioengineered 2022, 13, 3410–3421. [Google Scholar] [CrossRef]
- Hou, X.; Dai, H.; Zheng, Y. Circular RNA Hsa_circ_0008896 Accelerates Atherosclerosis by Promoting the Proliferation, Migration and Invasion of Vascular Smooth Muscle Cells via Hsa-MiR-633/CDC20B (Cell Division Cycle 20B) Axis. Bioengineered 2022, 13, 5987–5998. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L. CircRNA-PTPRA Promoted the Progression of Atherosclerosis through Sponging with MiR-636 and Upregulating the Transcription Factor SP1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12437–12449. [Google Scholar] [CrossRef]
- Mei, R.; Wu, M.; Ren, F. Knockdown of Circ_0002194 Protects against Oxidized Low-Density Lipoprotein-Induced Cell Damage via the Regulation of the MiR-637/PACS2 Axis in Human Vascular Endothelial Cells. Interact. Cardiovasc. Thorac. Surg. 2022, 35, ivac210. [Google Scholar] [CrossRef]
- Huang, H.; Huang, X.; Yu, H.; Xue, Y.; Zhu, P. Circular RNA Circ-RELL1 Regulates Inflammatory Response by MiR-6873-3p/MyD88/NF-ΚB Axis in Endothelial Cells. Biochem. Biophys. Res. Commun. 2020, 525, 512–519. [Google Scholar] [CrossRef]
- Lin, D.-S.; Zhang, C.-Y.; Li, L.; Ye, G.-H.; Jiang, L.-P.; Jin, Q. Circ_ROBO2/MiR-149 Axis Promotes the Proliferation and Migration of Human Aortic Smooth Muscle Cells by Activating NF-ΚB Signaling. Cytogenet. Genome Res. 2021, 161, 414–424. [Google Scholar] [CrossRef]
- Mao, X.; Wang, L.; Chen, C.; Tao, L.; Ren, S.; Zhang, L. Circ_0124644 Enhances Ox-LDL-Induced Cell Damages in Human Umbilical Vein Endothelial Cells through Upregulating FOXO4 by Sponging MiR-370-3p. Clin. Hemorheol. Microcirc. 2022, 81, 135–147. [Google Scholar] [CrossRef]
- Ye, Q.; Ju, C.; Ye, Z.; Tong, J. Circ_ROBO2/MiR-186-5p/TRIM14 Axis Regulates Oxidized Low-Density Lipoprotein-Induced Cardiac Microvascular Endothelial Cell Injury. Regen. Ther. 2022, 20, 138–146. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Shao, D.; Hao, S.; Yuan, Y.; Liu, H.; Liu, F.; Mu, Q. CircSCAP Aggravates Oxidized Low-Density Lipoprotein-Induced Macrophage Injury by Upregulating PDE3B by MiR-221-5p in Atherosclerosis. J. Cardiovasc. Pharmacol. 2021, 78, e749–e760. [Google Scholar] [CrossRef] [PubMed]
- Du, N.; Li, M.; Yang, D. Hsa_circRNA_102541 Regulates the Development of Atherosclerosis by Targeting MiR-296-5p/PLK1 Pathway. Ir. J. Med. Sci. 2022, 191, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, S.; Guo, S.; Zhang, X.; Yang, C.; Su, G.; Wan, J. Circ_0004104 Participates in the Regulation of Ox-LDL-Induced Endothelial Cells Injury via MiR-942-5p/ROCK2 Axis. BMC Cardiovasc. Disord. 2022, 22, 517. [Google Scholar] [CrossRef]
- Guan, Z.; Lu, R.; Sun, Y.; Wang, X.; Yu, C.; Song, T. Regulation of Oxidized LDL-Induced Proliferation and Migration in Human Vascular Smooth Muscle Cells by a Novel Circ_0007478/MiR-638/ROCK2 CeRNA Network. Vasc. Med. 2023, 28, 6–17. [Google Scholar] [CrossRef]
- Ye, B.; Liang, X.; Zhao, Y.; Cai, X.; Wang, Z.; Lin, S.; Wang, W.; Shan, P.; Huang, W.; Huang, Z. Hsa_circ_0007478 Aggravates NLRP3 Inflammasome Activation and Lipid Metabolism Imbalance in Ox-LDL-Stimulated Macrophage via MiR-765/EFNA3 Axis. Chem. Biol. Interact. 2022, 368, 110195. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Liu, C.; Xu, J.; Li, S.; Dai, D.; Zhang, L.; Yonghui, P. Circ_0021155 Can Participate in the Phenotypic Transformation of Human Vascular Smooth Muscle Cells via the MiR-4459/TRPM7 Axis. Biochem. Biophys. Res. Commun. 2022, 630, 133–142. [Google Scholar] [CrossRef]
- Chen, M.; Li, F.; Jiang, Q.; Zhang, W.; Li, Z.; Tang, W. Role of MiR-181b/Notch1 Axis in Circ_TNPO1 Promotion of Proliferation and Migration of Atherosclerotic Vascular Smooth Muscle Cells. J. Healthc. Eng. 2022, 2022, 4086935. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Chen, Z.; Wang, M. Blocking Circ_UBR4 Suppressed Proliferation, Migration, and Cell Cycle Progression of Human Vascular Smooth Muscle Cells in Atherosclerosis. Open Life Sci. 2021, 16, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.; Ding, Y.; Tian, Y.; Lei, X. Circular RNA Circ_0010283 Regulates the Viability and Migration of Oxidized Low-density Lipoprotein-induced Vascular Smooth Muscle Cells via an MiR-370-3p/HMGB1 Axis in Atherosclerosis. Int. J. Mol. Med. 2020, 46, 1399–1408. [Google Scholar] [CrossRef]
- Feng, Z.; Zhu, Y.; Zhang, J.; Yang, W.; Chen, Z.; Li, B. Hsa-Circ_0010283 Regulates Oxidized Low-Density Lipoprotein-Induced Proliferation and Migration of Vascular Smooth Muscle Cells by Targeting the MiR-133a-3p/Pregnancy-Associated Plasma Protein A Axis. Circ. J. 2020, 84, 2259–2269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, Y.; Zhu, J.; Xie, Y.; Wu, R.; Zhong, J.; Qiu, Z.; Jiang, L. Circ_0086296 Induced Atherosclerotic Lesions via the IFIT1/STAT1 Feedback Loop by Sponging MiR-576-3p. Cell. Mol. Biol. Lett. 2022, 27, 80. [Google Scholar] [CrossRef]
- Zhao, Q.; Lu, Y.-H.; Wang, X.; Zhang, X.-J. Circ_USP36/MiR-182-5p/KLF5 Axis Regulates the Ox-LDL-Induced Injury in Human Umbilical Vein Smooth Muscle Cells. Am. J. Transl. Res. 2020, 12, 7855–7869. [Google Scholar]
- Wan, H.; You, T.; Luo, W. Circ_0003204 Regulates Cell Growth, Oxidative Stress, and Inflammation in Ox-LDL-Induced Vascular Endothelial Cells via Regulating MiR-942-5p/HDAC9 Axis. Front. Cardiovasc. Med. 2021, 8, 646832. [Google Scholar] [CrossRef]
- Lei, X.; Yang, Y. Oxidized Low-Density Lipoprotein Contributes to Injury of Endothelial Cells via the Circ_0090231/MiR-9-5p/TXNIP Axis. Cent. Eur. J. Immunol. 2022, 47, 41–57. [Google Scholar] [CrossRef]
- Peng, H.; Sun, J.; Li, Y.; Zhang, Y.; Zhong, Y. Circ-USP9X Inhibition Reduces Oxidized Low-Density Lipoprotein–Induced Endothelial Cell Injury via the MicroRNA 599/Chloride Intracellular Channel 4 Axis. J. Cardiovasc. Pharmacol. 2021, 78, 560–571. [Google Scholar] [CrossRef]
- Wen, Y.; Chun, Y.; Lian, Z.; Yong, Z.; Lan, Y.; Huan, L.; Xi, C.; Juan, L.; Qing, Z.; Jia, C.; et al. CircRNA-0006896-miR1264-DNMT1 Axis Plays an Important Role in Carotid Plaque Destabilization by Regulating the Behavior of Endothelial Cells in Atherosclerosis. Mol. Med. Rep. 2021, 23, 311. [Google Scholar] [CrossRef]
- Chen, W.; Liu, Y.; Li, L.; Liang, B.; Wang, S.; Xu, X.; Xing, D.; Wu, X. The Potential Role and Mechanism of CircRNAs in Foam Cell Formation. Noncoding RNA Res. 2023, 8, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Dergunov, A.D.; Baserova, V.B. Different Pathways of Cellular Cholesterol Efflux. Cell Biochem. Biophys. 2022, 80, 471–481. [Google Scholar] [CrossRef]
- Altesha, M.; Ni, T.; Khan, A.; Liu, K.; Zheng, X. Circular RNA in Cardiovascular Disease. J. Cell Physiol. 2019, 234, 5588–5600. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, T.; Xiao, J. Circular RNAs: Promising Biomarkers for Human Diseases. EBioMedicine 2018, 34, 267–274. [Google Scholar] [CrossRef]
- Li, M.; Ding, W.; Liu, G.; Wang, J. Extracellular Circular RNAs Act as Novel First Messengers Mediating Cell Cross-Talk in Ischemic Cardiac Injury and Myocardial Remodeling. J. Cardiovasc. Transl. Res. 2022, 15, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, X.; Gao, C.; Jian, D.; Hao, P.; Rao, L.; Li, M. Peripheral Blood Circular RNA Hsa_circ_0124644 Can Be Used as a Diagnostic Biomarker of Coronary Artery Disease. Sci. Rep. 2017, 7, 39918. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shen, C.; Wang, Y.; Zou, T.; Zhu, H.; Lu, X.; Li, L.; Yang, B.; Chen, J.; Chen, S.; et al. Identification of Circular RNA Hsa_circ_0001879 and Hsa_circ_0004104 as Novel Biomarkers for Coronary Artery Disease. Atherosclerosis 2019, 286, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.P.; Pan, Y.H.; Cai, M.Y.; Cen, J.M.; Chen, C.; Zheng, L.; Liu, X.; Xiong, X.D. Plasma-Derived Exosomal Circular RNA Hsa_circ_0005540 as a Novel Diagnostic Biomarker for Coronary Artery Disease. Dis. Markers 2020, 2020, 3178642. [Google Scholar] [CrossRef]
- Chen, H.H.; Stewart, A.F.R. Transcriptomic Signature of Atherosclerosis in the Peripheral Blood: Fact or Fiction? Curr. Atheroscler. Rep. 2016, 18, 77. [Google Scholar] [CrossRef]
- Bazan, H.A.; Hatfield, S.A.; Brug, A.; Brooks, A.J.; Lightell, D.J.; Woods, T.C. Carotid Plaque Rupture Is Accompanied by an Increase in the Ratio of Serum CircR-284 to MiR-221 Levels. Circ. Cardiovasc. Genet. 2017, 10, e001720. [Google Scholar] [CrossRef]
- Wang, L.; Xu, G.; Spanos, M.; Li, G.; Lei, Z.; Sluijter, J.P.G.; Xiao, J. Circular RNAs in Cardiovascular Diseases: Regulation and Therapeutic Applications. Research 2023, 6, 38. [Google Scholar] [CrossRef]
- Wesselhoeft, R.A.; Kowalski, P.S.; Anderson, D.G. Engineering Circular RNA for Potent and Stable Translation in Eukaryotic Cells. Nat. Commun. 2018, 9, 2629. [Google Scholar] [CrossRef]
- Hu, X.; Qin, H.; Yan, Y.; Wu, W.; Gong, S.; Wang, L.; Jiang, R.; Zhao, Q.; Sun, Y.; Wang, Q.; et al. Exosomal Circular RNAs: Biogenesis, Effect, and Application in Cardiovascular Diseases. Front. Cell Dev. Biol. 2022, 10, 948256. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, S.; Tehrani, F.R.; Tahmasebi, S.; Shafiee, A.; Hashemi, S.M. Exosome Engineering in Cell Therapy and Drug Delivery. Inflammopharmacology 2023, 31, 145–169. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, F.; Chen, R.; Wang, Z.; Su, X.; Yang, C. Therapeutic Potential of Exosomes Derived From CircRNA_0002113 Lacking Mesenchymal Stem Cells in Myocardial Infarction. Front. Cell Dev. Biol. 2022, 9, 779524. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Qiu, C.; Tang, X.; Wan, R.; Wu, Z.; Zou, D.; Wang, W.; Luo, Y.; Liu, T. Investigation of the Clinicopathological and Prognostic Role of CircMTO1 in Multiple Cancers. Expert. Rev. Mol. Diagn. 2023, 23, 159–170. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, Z.; Lv, B.; Xu, X. The Diagnostic and Therapeutic Role of Circular RNA HIPK3 in Human Diseases. Diagnostics 2022, 12, 2469. [Google Scholar] [CrossRef]
- Li, G.; Zhu, C.; Qiao, D.; Chen, R. Circ_0001946 Promotes the Development of Acute Myeloid Leukemia by Upregulating PDL1. Turk. J. Haematol. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Fu, W.; Wang, X.; Xiang, J.; Chen, S.; Xia, R.; Xie, F.; Chi, B.; Qin, F.; Li, Z.; Mou, L.; et al. CircPTPRA Promotes the Progression of Pancreatic Ductal Adenocarcinoma via the MiR-140-5p/LMNB1 Axis. Cancer Med. 2023, 12, 11651–11671. [Google Scholar] [CrossRef]
- Hossain, M.T.; Li, S.; Reza, M.S.; Feng, S.; Zhang, X.; Jin, Z.; Wei, Y.; Peng, Y. Identification of CircRNA Biomarker for Gastric Cancer through Integrated Analysis. Front. Mol. Biosci. 2022, 9, 857320. [Google Scholar] [CrossRef]
- Du, W.W.; Zhang, C.; Yang, W.; Yong, T.; Awan, F.M.; Yang, B.B. Identifying and Characterizing CircRNA-Protein Interaction. Theranostics 2017, 7, 4183–4191. [Google Scholar] [CrossRef]
- Gao, X.; Xia, X.; Li, F.; Zhang, M.; Zhou, H.; Wu, X.; Zhong, J.; Zhao, Z.; Zhao, K.; Liu, D.; et al. Circular RNA-Encoded Oncogenic E-Cadherin Variant Promotes Glioblastoma Tumorigenicity through Activation of EGFR-STAT3 Signalling. Nat. Cell Biol. 2021, 23, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Sinha, T.; Panigrahi, C.; Das, D.; Chandra Panda, A. Circular RNA Translation, a Path to Hidden Proteome. Wiley Interdiscip. Rev. RNA 2022, 13, e1685. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Xu, Y.; Zhang, X.; Deng, S.; Yuan, Y.; Luo, X.; Hossain, M.T.; Zhu, X.; Du, K.; Hu, F.; et al. A Novel Protein AXIN1-295aa Encoded by CircAXIN1 Activates the Wnt/β-Catenin Signaling Pathway to Promote Gastric Cancer Progression. Mol. Cancer 2021, 20, 158. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.Y.; Qadir, J.; Yang, B.B. Circular RNA Translation: Novel Protein Isoforms and Clinical Significance. Trends Mol. Med. 2022, 28, 405–420. [Google Scholar] [CrossRef]
- Li, F.; Tang, H.; Zhao, S.; Gao, X.; Yang, L.; Xu, J. Circ-E-Cad Encodes a Protein That Promotes the Proliferation and Migration of Gastric Cancer via the TGF-β/Smad/C-E-Cad/PI3K/AKT Pathway. Mol. Carcinog. 2023, 62, 360–368. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| circRNA (Host Gene) | Source * | Interactions miRNA/mRNA | Confirmation * | Effect, (Potential Application) | Reference |
|---|---|---|---|---|---|
| circ_0026218 (CERS5) | HUVEC | miR-338-3p/SIRT6 | Microarray, RT-PCR, DLR, RIP | Proliferation ↑ Inflammation ↓ Oxidative stress ↓ Apoptosis ↓ (Target for therapy) | [98] |
| circ_0030042 (FOXO1) | HUVEC | miR-616-3p/RFX7 | RT-PCR, DLR, RIP, RPD | Proliferation ↑ Apoptosis ↓ Inflammation ↓ (Target for therapy) | [99] |
| circHIPK3 (HIPK3) | Serum, VSMC, HUVEC | miR-106a-5p/MFN2 | RT-PCR, DLR | Osteogenic and cartilage differentiation ↓ Vascular calcification ↓ (Target for therapy) | [100] |
| miR-190b/ATG7 | [101,102] | ||||
| circMTO1 (MTO1) | Serum, VSMC | miR-182-5p/RASA1 | RT-PCR, DLR | Apoptosis ↑ Proliferation ↓ (Target for therapy) | [103] |
| circ_0000345 (RSF1) | Serum, ASMC | miR-647/PAPD5 | RT-PCR, DLR, RIP | Apoptosis ↑ Proliferation ↓ Inflammation ↓ (Target for therapy) | [84] |
| Serum, HAEC, HUVEC | miR-758/CCND2, miR-129-5p/TET2 | RT-PCR, DLR, RIP | Proliferation ↑ Apoptosis ↓ (Target for therapy) | [104,105] | |
| circ_06206 (SCRG1) | Serum, HUVEC | miR-1268b/NR4A1 | RNA-Seq, RT-PCR, DLR | Angiogenesis ↓ (Target for therapy) | [106] |
| circ_0093887 (Sirt1) | Serum, HAEC, VSMC | miR-758-3p/BAMBI, miR-876-3p/CCND2, miR-876/SUCNRA, miR-132/212/SIRT1 | RT-PCR, DLR, RIP, RPD | Proliferation ↑ Apoptosis ↓ Inflammation ↓ (Target for therapy) | [107,108,109] |
| circ_0001445 (SMARCA5) | HUVEC | miR-208b-5p/ABCG1 | RT-PCR, DLR, RIP, RPD | Proliferation ↑ Inflammation ↓ Foam cells transformation ↓ (Biomarker, Target for therapy) | [110] |
| circSmoc1-2 (Smoc1-2) | VSMC | miR-874-3p/ADAM19 | RNA-Seq, RT-PCR, Calcium assay, RISH | Vascular calcification ↓ (Target for therapy) | [111] |
| circ_0107197 (TEX14) | Serum, VSMC | miR-6509-3p/THAP1 | RT-PCR, DLR, RIP | Apoptosis ↑ Proliferation ↓ (Target for therapy) | [112] |
| circ_0003423 (ZNF532) | HUVEC | miR-142-3p/SIRT3, SOD2 | RT-PCR, DLR, RIP | Proliferation ↑ Oxidative stress ↓ Apoptosis ↓ (Target for therapy) | [113] |
| circRNA (Host Gene) | Source * | Interactions miRNA/mRNA | Confirmation * | Effect,(Potential Application) | Reference |
|---|---|---|---|---|---|
| circ_0002984 (ARHGAP32) | Serum, VSMC | miR-326-3p/VAMP3, miR-665/FGF2 | RT-PCR, DLR, RIP, RPD | Inflammation↑ Proliferation ↑ (Target for therapy) | [117,118] |
| circ_0003645 (C16orf62) | HUVEC | miR-149-3p/TRAF7 | RT-PCR, DLR, RIP | Apoptosis ↑ Proliferation ↓ (Target for therapy) | [119] |
| circ_0005699 (C16orf62) | HUVEC | miR-450b-5P/NFKB1 | RT-PCR, DLR | Inflammation ↑ (Target for therapy) | [120] |
| circ_0001946 (CDR1) | Serum, mouse ventricles | miR-7-5p/PARP1 | RT-PCR, DLR, target prediction in silico | Apoptosis ↑ (Target for therapy, Biomarker) | [121,122] |
| circ_0026218 (CERS5) | Serum, HUVEC | miR-188-3p/TLR4 | RT-PCR, DLR, RIP, RPD | Apoptosis ↑ Inflammation↑ Oxidative stress ↑ Proliferation ↓ (Target for therapy, Biomarker) | [123] |
| circ_0029589 (CHFR) | Serum, HUVEC | miR-15b-5p/GADD45G, miR-1197/RAB22A, miR-370/FOXO1 | Microarray, RT-PCR, DLR, RPD | Apoptosis ↑ Inflammation ↑ Oxidative stress ↑ proliferation ↓ (Target for therapy) | [124,125,126] |
| circ_0003575 (CHMP5) | HUVEC | miR-532-5p/ROCK2 | RT-PCR, DLR, RIP | Apoptosis ↑ Inflammation ↑ Proliferation ↓ (Target for therapy) | [127] |
| circCOL1A1 (COL1A1) | Serum, VSMC | miR-30a-5p/SMAD1 | RT-PCR, RISH, DLR, RPD | VSMC transformation ↑ (Target for therapy, Biomarker) | [128] |
| circ_0050486 (GPI) | THP-1 | miR-1270, miR-145/NF1A, MMP16, USP31 | RT-PCR, DLR | Inflammation ↑ Apoptosis ↑ (Target for therapy) | [129] |
| Serum, HAEC | miR-182-5p/MYD88 | RT-PCR, DLR, RIP | Proliferation ↓ (Target for therapy, Biomarker) | [130] | |
| circ_0044073 (GRN) | Serum, HUVEC, HUVSMC | miR-107/JAK1 | RT-PCR, DLR, RPD | Proliferation ↑ (Target for therapy) | [92] |
| circ_0057583 (HECW2) | Serum, CMEC | miR-942-5p/TLR4 | RT-PCR, DLR, RPD | Apoptosis ↑ Proliferation ↓ (Target for therapy) | [31] |
| circ_0091822 (IRAK1) | Serum, HUVEC | miR-330-5p/TRIM14, miR-661/RAB22A | RT-PCR, DLR, RIP, RPD | Inflammation ↑ Apoptosis ↑ Oxidative stress ↑ Proliferation ↓ (Biomarker, Target for therapy,) | [131,132] |
| circ_0018146 (ITGB1) | Serum | miR-342-3p/NFAM1 | Microarray, RT-PCR, DLR, RPD | Dendritic cell maturation ↓ (Target for therapy, Biomarker) | [133] |
| circ_0001879 (NIPSNAP3A) | Serum HUVEC | miR-6873-5p/HDAC9 | RT-PCR, DLR, RIP | Inflammation ↑ Proliferation ↓ Cholesterol transport ↓ (Target for therapy) | [134] |
| circ_0009135 (NPHP4) | Serum Monocytes, EVs | miR-1231/EGFR | Microarray, RT-PCR, DLR, RIP, RPD | Heterogeneous adhesion ↑ (Target for therapy, Biomarker) | [135] |
| circ_0033596 (PACS2) | HUVEC | miR-217-5p/CLIC4 | RT-PCR, DLR, RIP | Apoptosis ↑ Proliferation ↓ (Target for therapy) | [136] |
| circ_0008896 (PPAPDC1A) | VSMC | miR-633/CDC20B | RT-PCR, DLR, RIP | Proliferation ↑ (Target for therapy) | [137] |
| circPTPRA (PTPRA) | Serum, VSMC | miR-636/SP1 | RT-PCR, DLR | Proliferation ↑ Apoptosis ↓ (Target for therapy, Biomarker) | [138] |
| circ_0002194 (RELL1) | HUVEC | miR-637/PACS2, miR-6873-3p/MYD88 | Microarray, RT-PCR, DLR, RIP, | Apoptosis ↑ Proliferation ↓ Inflammation ↑ Oxidative stress ↑ (Target for therapy) | [139,140] |
| circ_0124644 (ROBO2) | Serum, HASMC | miR-149/TRAF6 | Microarray, RT-PCR, DLR, RPD | Proliferation ↑ Inflammation ↑ Apoptosis ↓ (Biomarker) | [141] |
| HUVEC, Serum, CMEC | miR-370-3p/FOXO4, miR-186-5p/TRIM14 | RT-PCR, DLR, RIP, RPD | Inflammation ↑ Apoptosis ↑ Proliferation ↓ (Biomarker, Target for therapy) | [142,143] | |
| circ_0001292 (SCAP) | Serum, THP-1 | miR-221-5p/PDE3B | RT-PCR, DLR, RPD | Inflammation ↑ Oxidative stress ↑ Lipid accumulation ↑ (Biomarker) | [144] |
| circ_102541 (SIPA1L1) | Serum, HUVEC | miR-296-5p/PLK1 | RT-PCR, DLR | Proliferation ↑ Apoptosis ↓ (Target for therapy, Biomarker) | [145] |
| circ_0004104 (SPARC) | Serum, HUVEC | miR-942-5p/ROCK2 | RT-PCR, DLR, RIP, RPD | Inflammation ↑ Apoptosis ↑ Proliferation ↓ (Target for therapy, Biomarker) | [146] |
| circ_0007478 (TM7SF3) | Serum, VSMC | miR-638/ROCK2 | RT-PCR, DLR, RIP | Proliferation ↑ (Target for therapy Biomarker) | [147,148] |
| THP-1 | miR-765/EFNA3 | Microarray, RT-PCR, DLR | Foam cells transformation ↑ (Target for therapy, Biomarker) | ||
| circ_0021155 (TMEM41B) | VSMC | miR-4459/TRPM7 | RNA-Seq, RISH, RT-PCR, DLR | Proliferation ↑ VSMC transformation ↑ (Target for therapy) | [149] |
| circ_0072951 (TNPO1) | Serum, VSMC | miR-181b/NOTCH1 | RT-PCR, DLR | Proliferation ↑ (Target for therapy, Biomarker) | [150] |
| circ_0010283 (UBR4) | Serum, VSMC | miR-107/ROCK, miR-370–3p/HMGB1, miR-133a-3p/PAPPA | RT-PCR, DLR, RIP, RPD | Proliferation ↑ (Target for therapy, Biomarker) | [151,152,153] |
| circ_0086296 (UHRF2) | Carotid plaque, HUVEC, aorta of atherosclerotic mice | miR-576-3p/IFIT1, STAT1 | Microarray, RT-PCR, RISH, Sanger-Seq, DLR, RIP, RPD | Inflammation ↑ Lipid accumulation ↑ (Target for therapy) | [154] |
| circ_0003204 (USP36) | Serum, VSMC | miR-182-5p/KLF5, miR-942-5p/HDAC9 | RT-PCR, DLR, RPD, RIP | Proliferation ↑ (Target for therapy, Biomarker) | [155,156] |
| circ_0090231 (USP9X) | Serum, HUVEC | miR-9-5p/TXNIP, miR-599/CLIC4 | RT-PCR, DLR, RIP | Apoptosis ↑ Oxidative stress ↑ Inflammation ↑ Proliferation ↓ (Target for therapy, Biomarker) | [157,158] |
| circ_0006896 (VIRMA) | EVs, HUVEC | miR-1264/DNMT1 | Microarray, RISH, RT-PCR, DLR | Plaque formation ↑ Proliferation ↑ (Target for therapy, Biomarker) | [159] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dergunova, L.V.; Vinogradina, M.A.; Filippenkov, I.B.; Limborska, S.A.; Dergunov, A.D. Circular RNAs Variously Participate in Coronary Atherogenesis. Curr. Issues Mol. Biol. 2023, 45, 6682-6700. https://doi.org/10.3390/cimb45080422
Dergunova LV, Vinogradina MA, Filippenkov IB, Limborska SA, Dergunov AD. Circular RNAs Variously Participate in Coronary Atherogenesis. Current Issues in Molecular Biology. 2023; 45(8):6682-6700. https://doi.org/10.3390/cimb45080422
Chicago/Turabian StyleDergunova, Liudmila V., Margarita A. Vinogradina, Ivan B. Filippenkov, Svetlana A. Limborska, and Alexander D. Dergunov. 2023. "Circular RNAs Variously Participate in Coronary Atherogenesis" Current Issues in Molecular Biology 45, no. 8: 6682-6700. https://doi.org/10.3390/cimb45080422
APA StyleDergunova, L. V., Vinogradina, M. A., Filippenkov, I. B., Limborska, S. A., & Dergunov, A. D. (2023). Circular RNAs Variously Participate in Coronary Atherogenesis. Current Issues in Molecular Biology, 45(8), 6682-6700. https://doi.org/10.3390/cimb45080422