
Microbiomic and Metabolomic Analyses Unveil the Protective Effect of Saffron in a Mouse Colitis Model


 , , and
, , and 
Abstract
1. Introduction
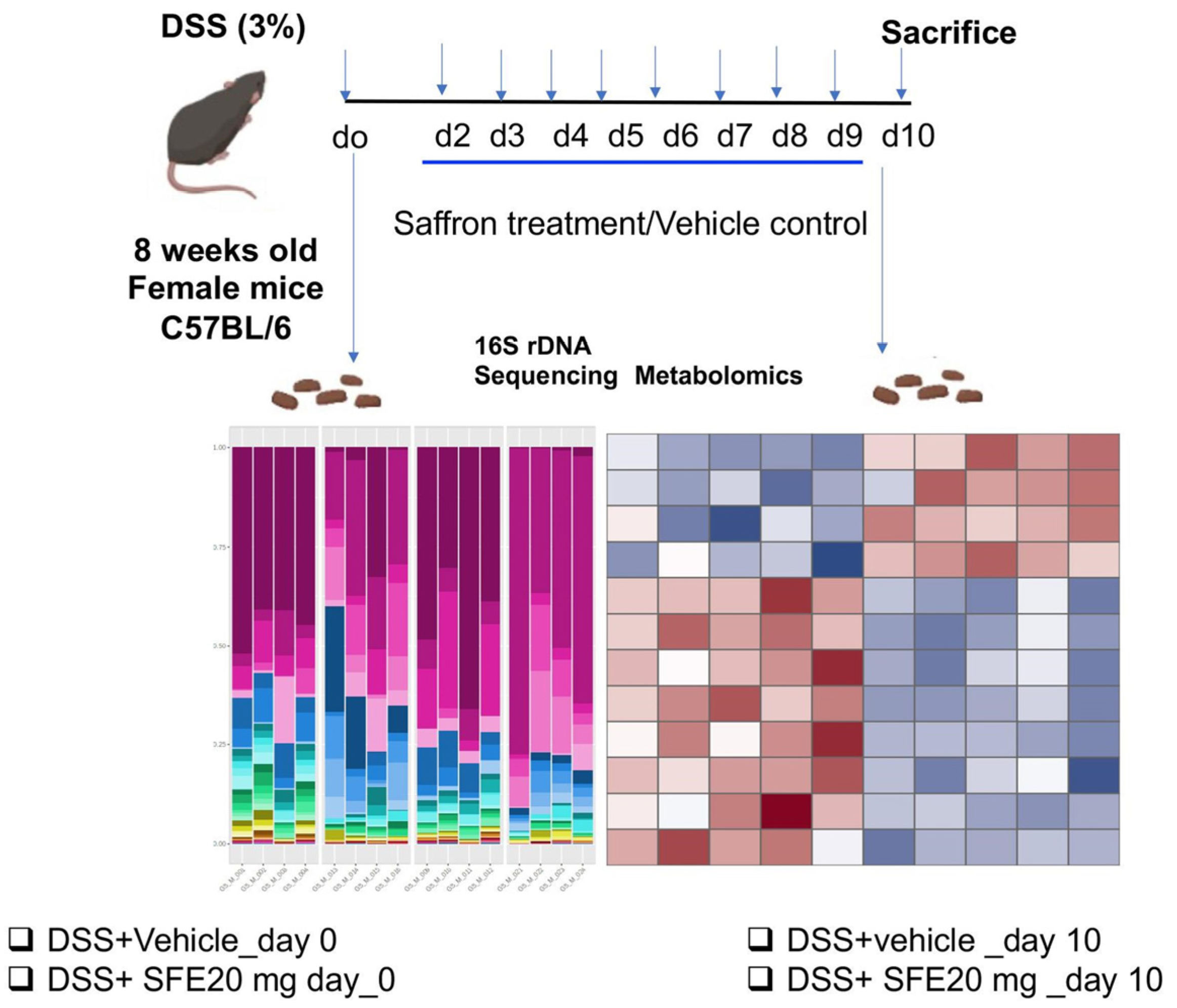
2. Materials and Methods
2.1. Animals
2.2. Saffron Aqueous Extract
2.3. Dextran Sodium Sulfate (DSS) Colitis Model
2.4. Microbiome Analysis
2.5. Untargeted Metabolomics
2.6. Statistical Analysis
3. Results
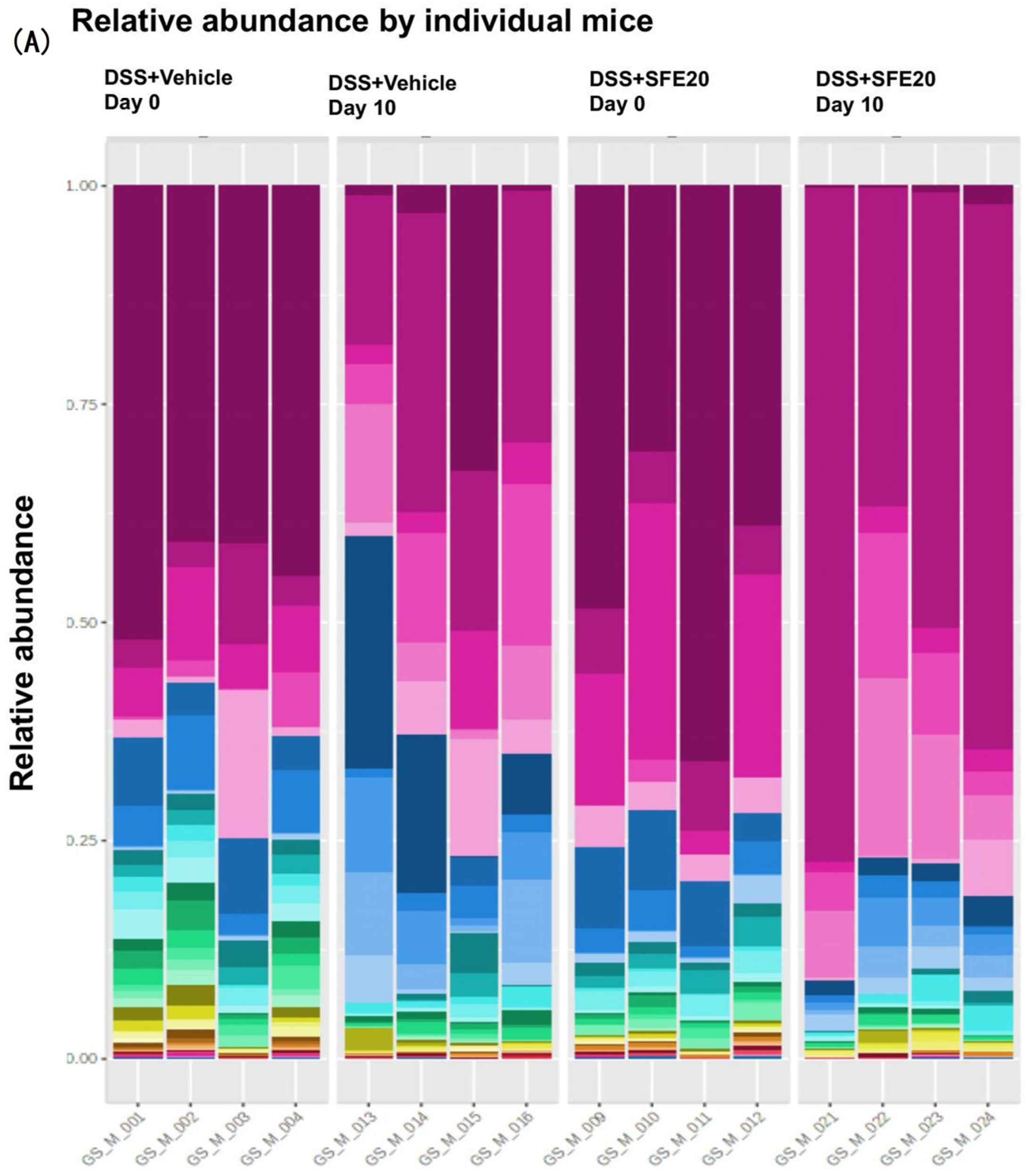
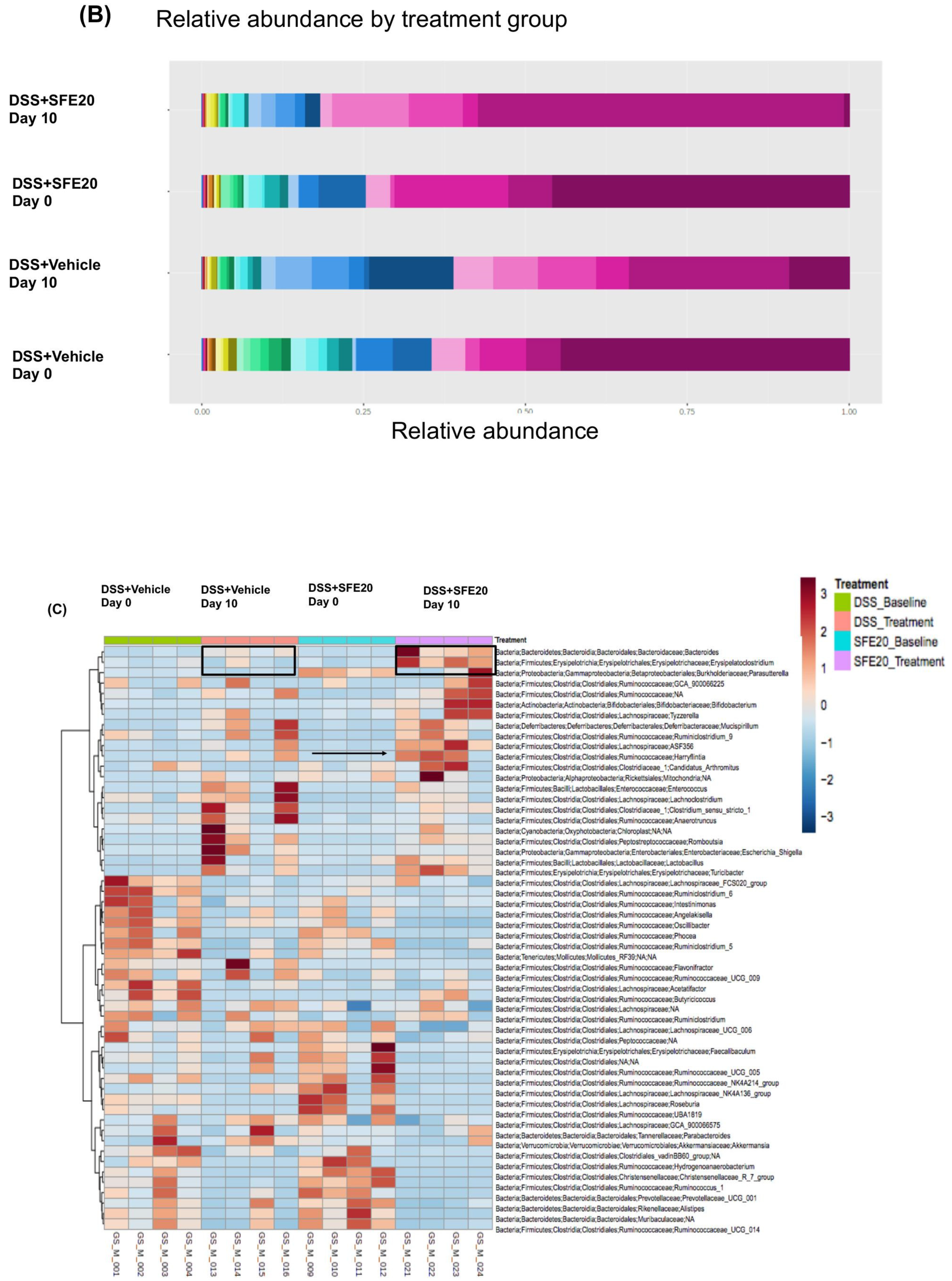
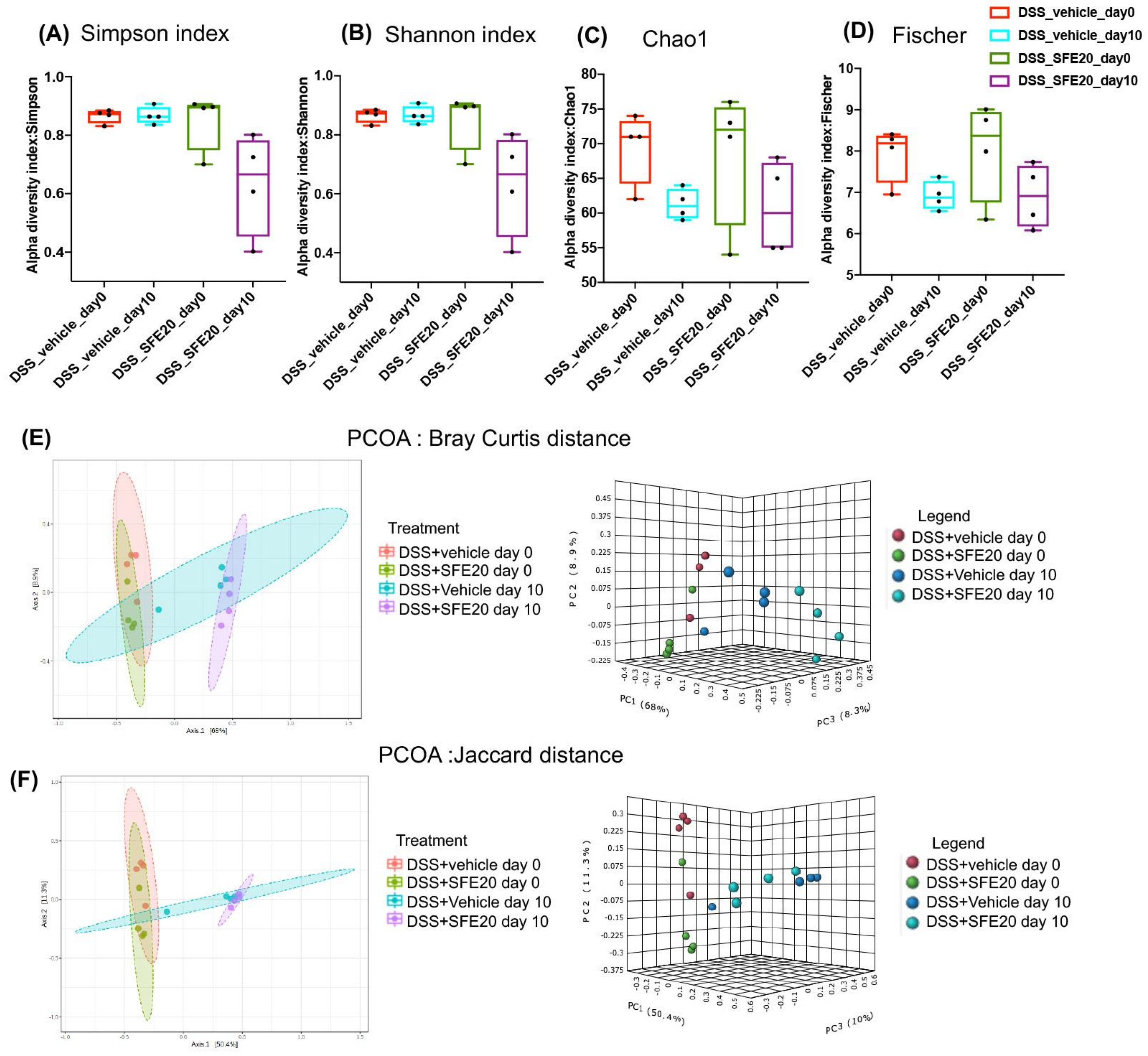
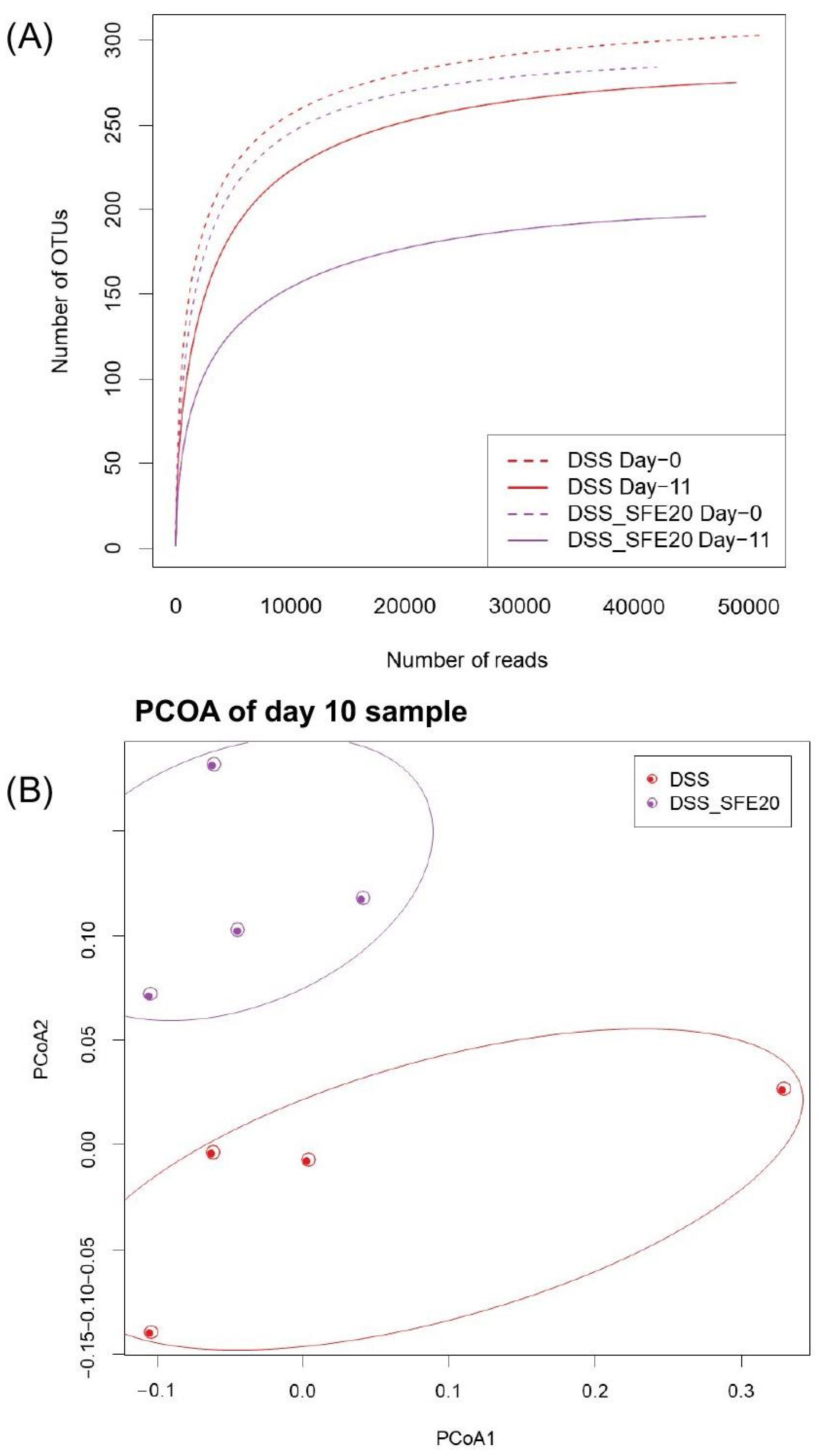
3.1. Saffron Treatment Alters DSS-Induced Microbial Profile
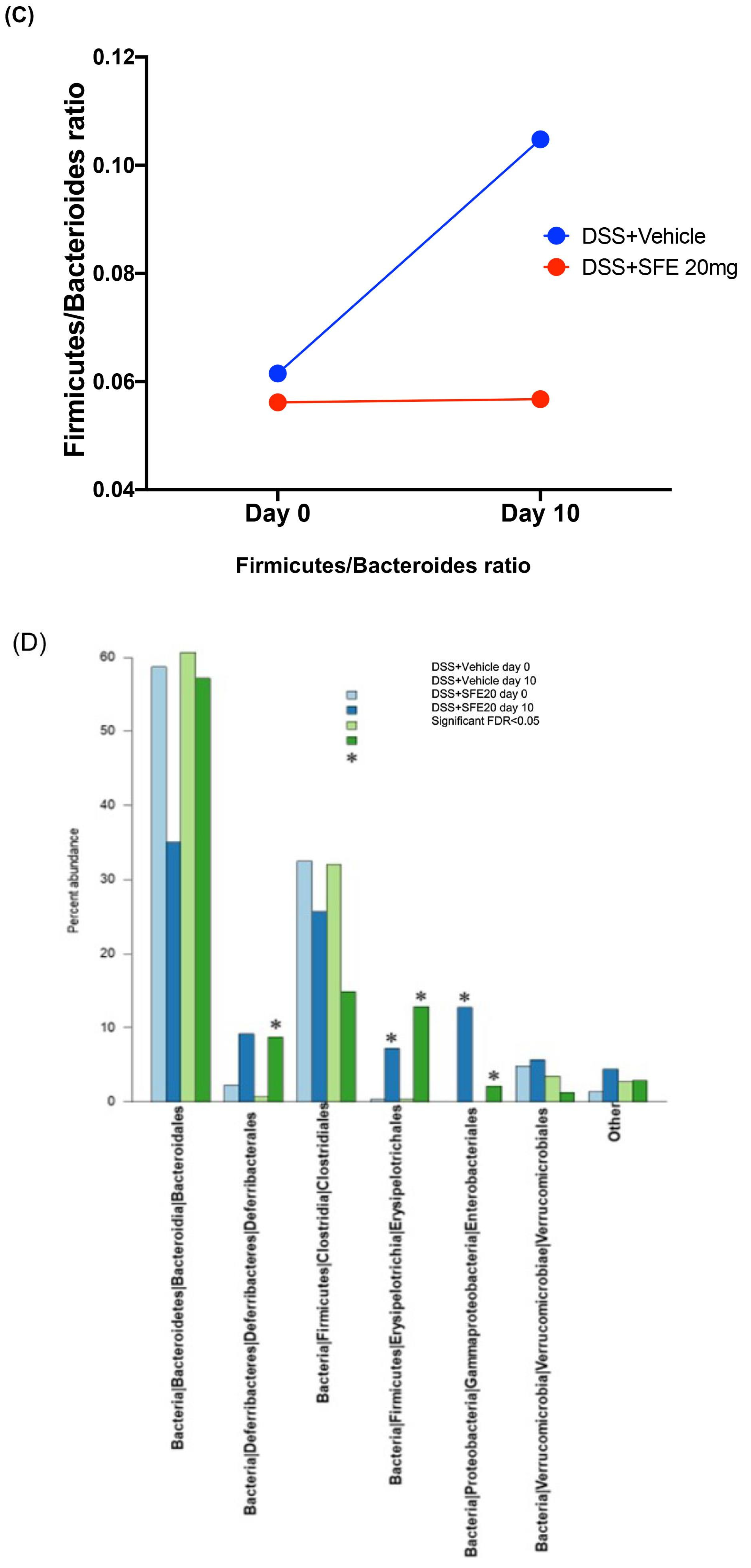
3.2. Saffron Alters the Abundance of Microbial Taxa Associated with Inflammation
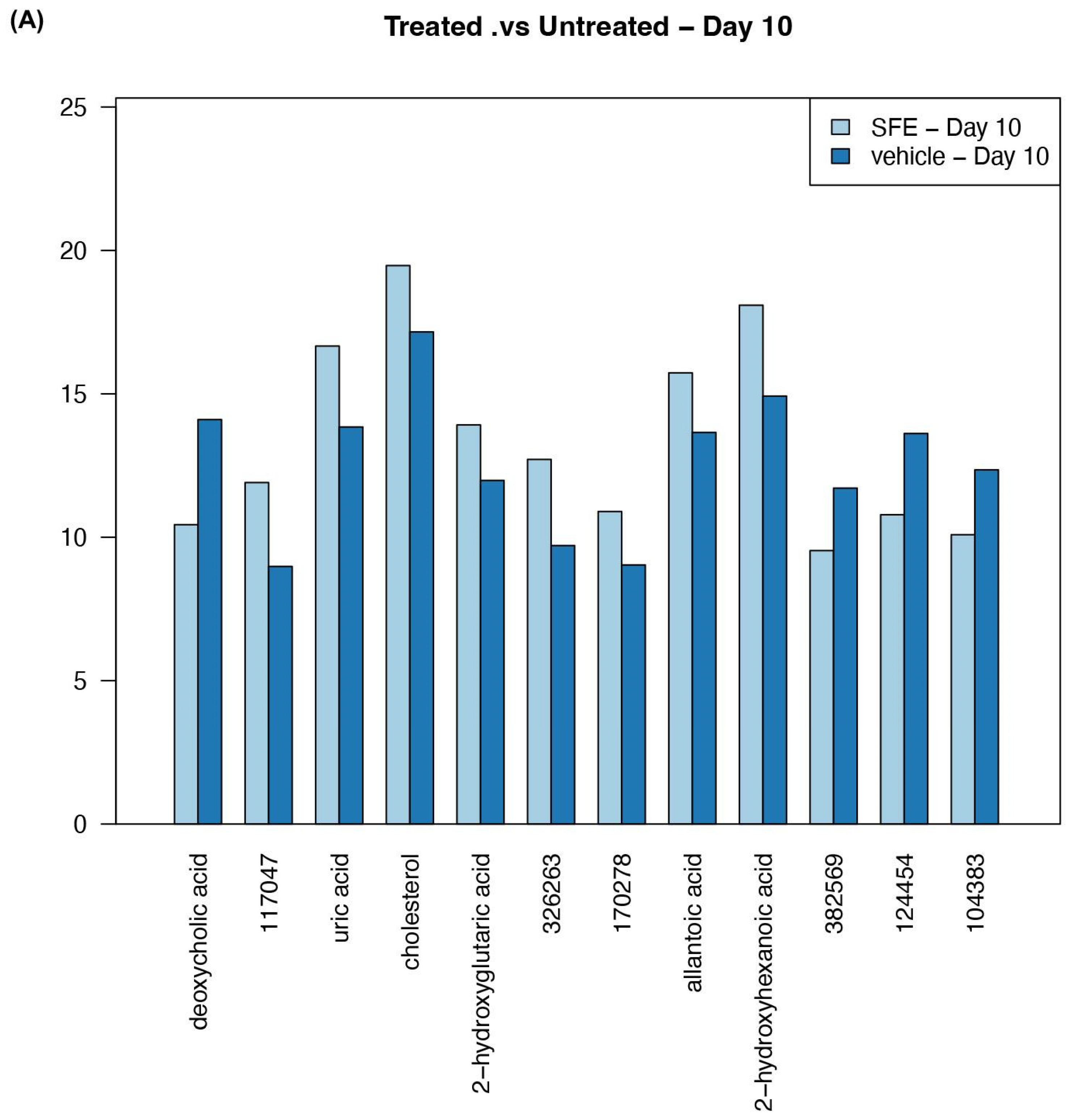
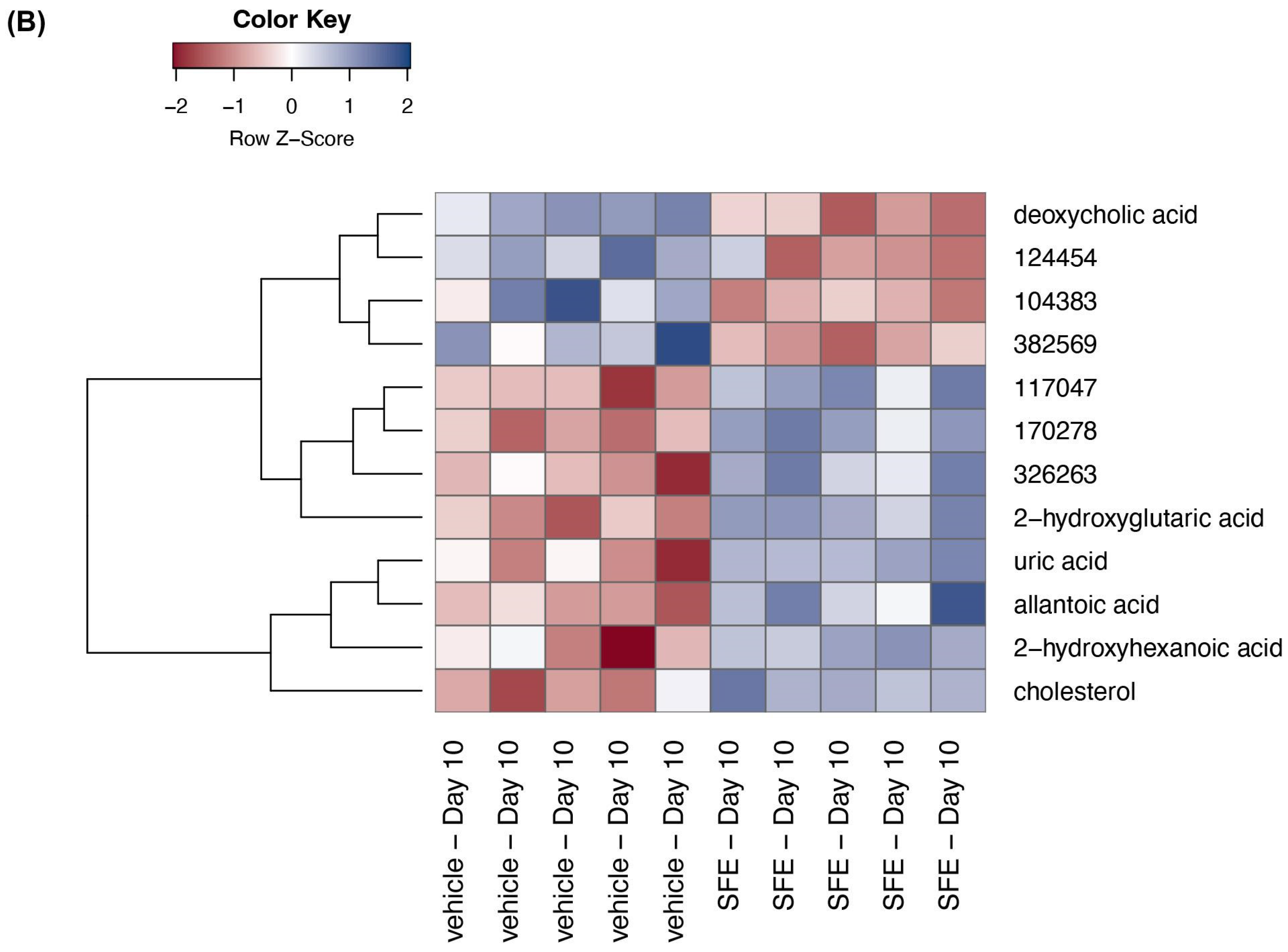
3.3. Saffron Treatment Alters Primary Metabolites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jairath, V.; Feagan, B.G. Global burden of inflammatory bowel disease. Lancet Gastroenterol. Hepatol. 2020, 5, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Mandip, K.C.; Shaukat, A.; Rector, T. All-cause hospitalizations for inflammatory bowel diseases: Can the reason for admission provide information on inpatient resource use? A study from a large veteran affairs hospital. Mil. Med. Res. 2016, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Shin, J.S.; Noh, Y.S.; Cho, Y.W.; Hong, S.J.; Park, J.H.; Lee, J.Y.; Lee, J.Y.; Lee, K.T. Anti-inflammatory effects of methanol extract of Patrinia scabiosaefolia in mice with ulcerative colitis. J. Ethnopharmacol. 2011, 136, 428–435. [Google Scholar] [CrossRef]
- Sergent, T.; Piront, N.; Meurice, J.; Toussaint, O.; Schneider, Y.J. Anti-inflammatory effects of dietary phenolic compounds in an in vitro model of inflamed human intestinal epithelium. Chem. Biol. Interact. 2010, 188, 659–667. [Google Scholar] [CrossRef]
- McLean, L.P.; Cross, R.K. Adverse events in IBD: To stop or continue immune suppressant and biologic treatment. Expert. Rev. Gastroenterol. Hepatol. 2014, 8, 223–240. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Comar, K.M.; Kirby, D.F. Herbal Remedies in Gastroenterology. J. Clin. Gastroenterol. 2005, 39, 457–468. [Google Scholar] [CrossRef]
- Joo, Y.E. Natural product-derived drugs for the treatment of inflammatory bowel diseases. Intest. Res. 2014, 12, 103–109. [Google Scholar] [CrossRef]
- Langmead, L.; Makins, R.J.; Rampton, D.S. Anti-inflammatory effects of aloe vera gel in human colorectal mucosa in vitro. Aliment. Pharm. 2004, 19, 521–527. [Google Scholar] [CrossRef]
- Ben-Arye, E.; Goldin, E.; Wengrower, D.; Stamper, A.; Kohn, R.; Berry, E. Wheat grass juice in the treatment of active distal ulcerative colitis: A randomized double-blind placebo-controlled trial. Scand. J. Gastroenterol. 2002, 37, 444–449. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Gupta, S.C.; Sung, B. Curcumin: An orally bioavailable blocker of TNF and other pro-inflammatory biomarkers. Br. J. Pharmacol. 2013, 169, 1672–1692. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Sugimoto, K. Curcumin has bright prospects for the treatment of inflammatory bowel disease. Curr. Pharm. Des. 2009, 15, 2087–2094. [Google Scholar] [CrossRef]
- Liu, D.-Y.; Gao, L.; Zhang, J.; Huo, X.-W.; Ni, H.; Cao, L. Anti-inflammatory and Anti-oxidant Effects of Licorice Flavonoids on Ulcerative Colitis in Mouse Model. Chin. Herb. Med. 2017, 9, 358–368. [Google Scholar] [CrossRef]
- Langmead, L.; Rampton, D.S. Review article: Complementary and alternative therapies for inflammatory bowel disease. Aliment. Pharm. 2006, 23, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Franzosa, E.A.; Sirota-Madi, A.; Avila-Pacheco, J.; Fornelos, N.; Haiser, H.J.; Reinker, S.; Vatanen, T.; Hall, A.B.; Mallick, H.; McIver, L.J.; et al. Gut microbiome structure and metabolic activity in inflammatory bowel disease. Nat. Microbiol. 2019, 4, 293–305. [Google Scholar] [CrossRef]
- Sheehan, D.; Moran, C.; Shanahan, F. The microbiota in inflammatory bowel disease. J. Gastroenterol. 2015, 50, 495–507. [Google Scholar] [CrossRef]
- Alam, M.T.; Amos, G.C.A.; Murphy, A.R.J.; Murch, S.; Wellington, E.M.H.; Arasaradnam, R.P. Microbial imbalance in inflammatory bowel disease patients at different taxonomic levels. Gut Pathog. 2020, 12, 1. [Google Scholar] [CrossRef]
- Sinha, S.R.; Haileselassie, Y.; Nguyen, L.P.; Tropini, C.; Wang, M.; Becker, L.S.; Sim, D.; Jarr, K.; Spear, E.T.; Singh, G.; et al. Dysbiosis-Induced Secondary Bile Acid Deficiency Promotes Intestinal Inflammation. Cell. Host Microbe 2020, 27, 659–670. [Google Scholar] [CrossRef]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Abdullaev, F.I.; Espinosa-Aguirre, J.J. Biomedical properties of saffron and its potential use in cancer therapy and chemoprevention trials. Cancer Detect. Prev. 2004, 28, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Ashktorab, H.; Soleimani, A.; Singh, G.; Amr, A.; Tabtabaei, S.; Latella, G.; Stein, U.; Akhondzadeh, S.; Solanki, N.; Gondre-Lewis, M.C.; et al. Saffron: The Golden Spice with Therapeutic Properties on Digestive Diseases. Nutrients 2019, 11, 943. [Google Scholar] [CrossRef]
- Festuccia, C.; Colapietro, A.; Mancini, A.; D’Alessandro, A. Crocetin and crocin from saffron in cancer chemotherapy and chemoprevention. Anticancer. Agents Med. Chem. 2018, 19, 38–47. [Google Scholar] [CrossRef]
- Siddiqui, M.J.; Saleh, M.S.M.; Basharuddin, S.; Zamri, S.H.B.; Mohd Najib, M.H.B.; Che Ibrahim, M.Z.B.; Binti Mohd Noor, N.A.; Binti Mazha, H.N.; Mohd Hassan, N.; Khatib, A. Saffron (Crocus sativus L.): As an Antidepressant. J. Pharm. Bioallied Sci. 2018, 10, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Haileselassie, Y.; Ji, A.R.; Maecker, H.T.; Sinha, S.R.; Brim, H.; Habtezion, A.; Ashktorab, H. Protective Effect of Saffron in Mouse Colitis Models Through Immune Modulation. Dig. Dis. Sci. 2022, 67, 2922–2935. [Google Scholar] [CrossRef]
- Jadhav, P.; Jiang, Y.; Jarr, K.; Layton, C.; Ashouri, J.F.; Sinha, S.R. Efficacy of Dietary Supplements in Inflammatory Bowel Disease and Related Autoimmune Diseases. Nutrients 2020, 12, 2156. [Google Scholar] [CrossRef]
- Banskota, S.; Brim, H.; Kwon, Y.H.; Singh, G.; Sinha, S.R.; Wang, H.; Khan, W.I.; Ashktorab, H. Saffron Pre-Treatment Promotes Reduction in Tissue Inflammatory Profiles and Alters Microbiome Composition in Experimental Colitis Mice. Molecules 2021, 26, 3351. [Google Scholar] [CrossRef]
- Tabtabaei, S.; D’Archivio, A.A.; Maggi, M.A.; Brutus, M.; Bajracharya, D.H.; Konakbayeva, D.; Soleimani, A.; Brim, H.; Ashktorab, H. Geographical classification of Iranian and Italian saffron sources based on HPLC analysis and UV-Vis spectra of aqueous extracts. Eur. Food Res. Technol. 2019, 245, 2435–2446. [Google Scholar] [CrossRef]
- Brim, H.; Yooseph, S.; Lee, E.; Sherif, Z.A.; Abbas, M.; Laiyemo, A.O.; Varma, S.; Torralba, M.; Dowd, S.E.; Nelson, K.E.; et al. A Microbiomic Analysis in African Americans with Colonic Lesions Reveals Streptococcus sp.VT162 as a Marker of Neoplastic Transformation. Genes 2017, 8, 314. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kim, M.; Lee, C.H. Treatment of Dextran Sulfate Sodium-Induced Colitis with Mucosa-Associated Lymphoid Tissue Lymphoma Translocation 1 Inhibitor MI-2 Is Associated with Restoration of Gut Immune Function and the Microbiota. Infect. Immun. 2018, 86, e00091-18. [Google Scholar] [CrossRef]
- Kozhakhmetov, S.; Babenko, D.; Kozhakhmetova, S.; Tuyakova, A.; Nurgaziyev, M.; Nurgozhina, A.; Muhanbetganov, N.; Chulenbayeva, L.; Sergazy, S.; Gulyayev, A.; et al. Gut modulation of dysbiosis induced by dextran sulfate sodium. Food Sci. 2021, 42, 101167. [Google Scholar] [CrossRef]
- Jun, G.; Aguilar, D.; Evans, C.; Burant, C.F.; Hanis, C.L. Metabolomic profiles associated with subtypes of prediabetes among Mexican Americans in Starr County, Texas, USA. Diabetologia 2020, 63, 287–295. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- Cerda-Bernad, D.; Valero-Cases, E.; Pastor, J.J.; Frutos, M.J. Saffron bioactives crocin, crocetin and safranal: Effect on oxidative stress and mechanisms of action. Crit. Rev. Food Sci. Nutr. 2022, 62, 3232–3249. [Google Scholar] [CrossRef] [PubMed]
- Finley, J.W.; Gao, S. A Perspective on Crocus sativus L. (Saffron) Constituent Crocin: A Potent Water-Soluble Antioxidant and Potential Therapy for Alzheimer’s Disease. J. Agric. Food Chem. 2017, 65, 1005–1020. [Google Scholar] [CrossRef]
- Chandra, P.; Sharma, R.K.; Arora, D.S. Antioxidant compounds from microbial sources: A review. Food Res. Int. 2020, 129, 108849. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef]
- Ahmadinejad, F.; Geir Moller, S.; Hashemzadeh-Chaleshtori, M.; Bidkhori, G.; Jami, M.S. Molecular Mechanisms behind Free Radical Scavengers Function against Oxidative Stress. Antioxidants 2017, 6, 51. [Google Scholar] [CrossRef]
- Zeinali, M.; Zirak, M.R.; Rezaee, S.A.; Karimi, G.; Hosseinzadeh, H. Immunoregulatory and anti-inflammatory properties of Crocus sativus (Saffron) and its main active constituents: A review. Iran. J. Basic. Med. Sci. 2019, 22, 334–344. [Google Scholar] [CrossRef]
- Asbaghi, O.; Sadeghian, M.; Sadeghi, O.; Rigi, S.; Tan, S.C.; Shokri, A.; Mousavi, S.M. Effects of saffron (Crocus sativus L.) supplementation on inflammatory biomarkers: A systematic review and meta-analysis. Phytother. Res. 2021, 35, 20–32. [Google Scholar] [CrossRef]
- Nair, S.C.; Kurumboor, S.K.; Hasegawa, J.H. Saffron chemoprevention in biology and medicine: A review. Cancer Biother. 1995, 10, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Maggi, M.A.; Bisti, S.; Picco, C. Saffron: Chemical Composition and Neuroprotective Activity. Molecules 2020, 25, 5618. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, T.; Shimeno, H.; Mishima, K.; Iwasaki, K.; Fujiwara, M.; Tanaka, H.; Shoyama, Y.; Toda, A.; Eyanagi, R.; Soeda, S. Protective effects of carotenoids from saffron on neuronal injury in vitro and in vivo. Biochim. Biophys. Acta 2007, 1770, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Skladnev, N.V.; Johnstone, D.M. Neuroprotective properties of dietary saffron: More than just a chemical scavenger? Neural Regen. Res. 2017, 12, 210–211. [Google Scholar] [CrossRef]
- Bisti, S.; Maccarone, R.; Falsini, B. Saffron and retina: Neuroprotection and pharmacokinetics. Vis. Neurosci. 2014, 31, 355–361. [Google Scholar] [CrossRef]
- Monchaux De Oliveira, C.; Pourtau, L.; Vancassel, S.; Pouchieu, C.; Capuron, L.; Gaudout, D.; Castanon, N. Saffron Extract-Induced Improvement of Depressive-Like Behavior in Mice Is Associated with Modulation of Monoaminergic Neurotransmission. Nutrients 2021, 13, 904. [Google Scholar] [CrossRef]
- Khazdair, M.R.; Boskabady, M.H.; Hosseini, M.; Rezaee, R.; Tsatsakis, A.M. The effects of Crocus sativus (saffron) and its constituents on nervous system: A review. Avicenna J. Phytomed 2015, 5, 376–391. [Google Scholar]
- Purushothuman, S.; Nandasena, C.; Peoples, C.L.; El Massri, N.; Johnstone, D.M.; Mitrofanis, J.; Stone, J. Saffron pre-treatment offers neuroprotection to Nigral and retinal dopaminergic cells of MPTP-Treated mice. J. Park. Dis. 2013, 3, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.; Brim, H.; Ashktorab, H. Saffron, Its Active Components, and Their Association with DNA and Histone Modification: A Narrative Review of Current Knowledge. Nutrients 2022, 14, 3317. [Google Scholar] [CrossRef] [PubMed]
- Lopresti, A.L.; Drummond, P.D. Saffron (Crocus sativus) for depression: A systematic review of clinical studies and examination of underlying antidepressant mechanisms of action. Hum. Psychopharmacol. 2014, 29, 517–527. [Google Scholar] [CrossRef]
- Cerda-Bernad, D.; Costa, L.; Serra, A.T.; Bronze, M.R.; Valero-Cases, E.; Perez-Llamas, F.; Candela, M.E.; Arnao, M.B.; Barberan, F.T.; Villalba, R.G.; et al. Saffron against Neuro-Cognitive Disorders: An Overview of Its Main Bioactive Compounds, Their Metabolic Fate and Potential Mechanisms of Neurological Protection. Nutrients 2022, 14, 5368. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Schwab, C.; Berry, D.; Rauch, I.; Rennisch, I.; Ramesmayer, J.; Hainzl, E.; Heider, S.; Decker, T.; Kenner, L.; Müller, M.; et al. Longitudinal study of murine microbiota activity and interactions with the host during acute inflammation and recovery. Isme J. 2014, 8, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Gevers, D.; Kugathasan, S.; Knights, D.; Kostic, A.D.; Knight, R.; Xavier, R.J. A Microbiome Foundation for the Study of Crohn’s Disease. Cell. Host Microbe 2017, 21, 301–304. [Google Scholar] [CrossRef]
- Huang, S.; Mao, J.; Zhou, L.; Xiong, X.; Deng, Y. The imbalance of gut microbiota and its correlation with plasma inflammatory cytokines in pemphigus vulgaris patients. Scand. J. Immunol. 2019, 90, e12799. [Google Scholar] [CrossRef]
- Silwedel, C.; Haarmann, A.; Fehrholz, M.; Claus, H.; Speer, C.P.; Glaser, K. More than just inflammation: Ureaplasma species induce apoptosis in human brain microvascular endothelial cells. J. Neuroinflammation 2019, 16, 38. [Google Scholar] [CrossRef]
- Chen, W.; Fan, H.; Liang, R.; Zhang, R.; Zhang, J.; Zhu, J. Taraxacum officinale extract ameliorates dextran sodium sulphate-induced colitis by regulating fatty acid degradation and microbial dysbiosis. J. Cell. Mol. Med. 2019, 23, 8161–8172. [Google Scholar] [CrossRef]
- Jia, W.; Xie, G.; Jia, W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef]
- Pavlidis, P.; Powell, N.; Vincent, R.P.; Ehrlich, D.; Bjarnason, I.; Hayee, B. Systematic review: Bile acids and intestinal inflammation-luminal aggressors or regulators of mucosal defence? Aliment. Pharm. 2015, 42, 802–817. [Google Scholar] [CrossRef]
- Zhao, S.; Gong, Z.; Zhou, J.; Tian, C.; Gao, Y.; Xu, C.; Chen, Y.; Cai, W.; Wu, J. Deoxycholic Acid Triggers NLRP3 Inflammasome Activation and Aggravates DSS-Induced Colitis in Mice. Front. Immunol. 2016, 7, 536. [Google Scholar] [CrossRef] [PubMed]
- Stenman, L.K.; Holma, R.; Forsgård, R.; Gylling, H.; Korpela, R. Higher fecal bile acid hydrophobicity is associated with exacerbation of dextran sodium sulfate colitis in mice. J. Nutr. 2013, 143, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- El Ridi, R.; Tallima, H. Physiological functions and pathogenic potential of uric acid: A review. J. Adv. Res. 2017, 8, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Beeharry, N.; Lowe, J.E.; Hernandez, A.R.; Chambers, J.A.; Fucassi, F.; Cragg, P.J.; Green, M.H.; Green, I.C. Linoleic acid and antioxidants protect against DNA damage and apoptosis induced by palmitic acid. Mutat. Res. 2003, 530, 27–33. [Google Scholar] [CrossRef]
- Wang, J.P.; Lee, J.H.; Yoo, J.S.; Cho, J.H.; Kim, H.J.; Kim, I.H. Effects of phenyllactic acid on growth performance, intestinal microbiota, relative organ weight, blood characteristics, and meat quality of broiler chicks. Poult. Sci. 2010, 89, 1549–1555. [Google Scholar] [CrossRef]
- Aldini, R.; Micucci, M.; Cevenini, M.; Fato, R.; Bergamini, C.; Nanni, C.; Cont, M.; Camborata, C.; Spinozzi, S.; Montagnani, M.; et al. Antiinflammatory effect of phytosterols in experimental murine colitis model: Prevention, induction, remission study. PLoS ONE 2014, 9, e108112. [Google Scholar] [CrossRef]
- Feng, S.; Dai, Z.; Liu, A.; Wang, H.; Chen, J.; Luo, Z.; Yang, C.S. β-Sitosterol and stigmasterol ameliorate dextran sulfate sodium-induced colitis in mice fed a high fat Western-style diet. Food Funct. 2017, 8, 4179–4186. [Google Scholar] [CrossRef]
- Ma, X.; Sun, Q.; Sun, X.; Chen, D.; Wei, C.; Yu, X.; Liu, C.; Li, Y.; Li, J. Activation of GABA(A) Receptors in Colon Epithelium Exacerbates Acute Colitis. Front. Immunol. 2018, 9, 987. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BinBase Name | Treated vs. Untreated Day 10 Log2-Fold-Change | Treated vs. Untreated Day 10 p-Value | Treated vs. Untreated Day 10 FDR |
|---|---|---|---|
| deoxycholic acid | −3.664 | <0.0001 | 0.015 |
| 2-hydroxyglutaric acid | 1.934 | <0.0001 | 0.039 |
| cholesterol | 2.314 | <0.0001 | 0.039 |
| uric acid | 2.822 | <0.0001 | 0.039 |
| 2-hydroxyhexanoic acid | 3.168 | 0.001 | 0.044 |
| allantoic acid | 2.077 | 0.001 | 0.044 |
| linoleic acid | 2.060 | 0.001 | 0.050 |
| 3-phenyllactic acid | 2.016 | 0.002 | 0.061 |
| beta-sitosterol | 2.039 | 0.003 | 0.074 |
| 2-aminobutyric acid | 1.963 | 0.003 | 0.075 |
| melibiose | −2.559 | 0.003 | 0.076 |
| urocanic acid | 2.248 | 0.004 | 0.076 |
| aconitic acid | 2.193 | 0.004 | 0.081 |
| N-acetylglycine | −2.132 | 0.006 | 0.105 |
| N-acetylornithine | 1.191 | 0.007 | 0.108 |
| fructose | −3.187 | 0.009 | 0.116 |
| homovanillic acid | 2.411 | 0.008 | 0.116 |
| malonic acid | 1.719 | 0.007 | 0.116 |
| mannose | −1.596 | 0.009 | 0.116 |
| mevalonic acid | 1.537 | 0.008 | 0.116 |
| N-acetylglutamate | 1.123 | 0.009 | 0.121 |
| citramalic acid | 2.546 | 0.010 | 0.126 |
| phytol | 1.454 | 0.010 | 0.126 |
| 3-aminoisobutyric acid | 1.689 | 0.015 | 0.145 |
| arachidonic acid | 1.531 | 0.013 | 0.145 |
| beta-glutamic acid | 1.792 | 0.013 | 0.145 |
| gluconic acid | 2.386 | 0.014 | 0.145 |
| lactitol | −1.326 | 0.015 | 0.145 |
| N-acetyl-D-mannosamine | −1.818 | 0.015 | 0.145 |
| pseudo uridine | 1.656 | 0.015 | 0.145 |
| thymine | 1.995 | 0.015 | 0.145 |
| sucrose | −1.998 | 0.016 | 0.146 |
| 2-deoxypentitol | 1.011 | 0.018 | 0.146 |
| tyrosine | −1.158 | 0.018 | 0.146 |
| 4-hydroxyphenylacetic acid | −2.761 | 0.019 | 0.147 |
| beta-alanine | 1.313 | 0.020 | 0.151 |
| glucose | −1.772 | 0.020 | 0.151 |
| orotic acid | 1.444 | 0.022 | 0.155 |
| 1-kestose | −2.516 | 0.024 | 0.165 |
| quinolinic acid | 1.073 | 0.025 | 0.170 |
| galactitol | −1.096 | 0.028 | 0.172 |
| N-acetyl-D-galactosamine | −2.163 | 0.027 | 0.172 |
| squalene | 1.114 | 0.029 | 0.179 |
| cytosine | 1.161 | 0.032 | 0.189 |
| 3-hydroxy-3-methylglutaric acid | 0.861 | 0.033 | 0.190 |
| glutaric acid | −1.407 | 0.034 | 0.191 |
| 3,6-anhydro-D-galactose | −0.833 | 0.038 | 0.204 |
| xylulose | 0.898 | 0.037 | 0.204 |
| 4-hydroxybutyric acid | 1.196 | 0.043 | 0.222 |
| dihydrocholesterol | 1.292 | 0.045 | 0.227 |
| epsilon-caprolactam | −1.141 | 0.045 | 0.227 |
| cis-gondoic acid | 1.116 | 0.049 | 0.230 |
| pipecolinic acid | 1.087 | 0.049 | 0.230 |
| raffinose | −3.357 | 0.048 | 0.230 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, G.; Brim, H.; Haileselassie, Y.; Varma, S.; Habtezion, A.; Rashid, M.; Sinha, S.R.; Ashktorab, H. Microbiomic and Metabolomic Analyses Unveil the Protective Effect of Saffron in a Mouse Colitis Model. Curr. Issues Mol. Biol. 2023, 45, 5558-5574. https://doi.org/10.3390/cimb45070351
Singh G, Brim H, Haileselassie Y, Varma S, Habtezion A, Rashid M, Sinha SR, Ashktorab H. Microbiomic and Metabolomic Analyses Unveil the Protective Effect of Saffron in a Mouse Colitis Model. Current Issues in Molecular Biology. 2023; 45(7):5558-5574. https://doi.org/10.3390/cimb45070351
Chicago/Turabian StyleSingh, Gulshan, Hassan Brim, Yeneneh Haileselassie, Sudhir Varma, Aida Habtezion, Mudasir Rashid, Sidhartha R. Sinha, and Hassan Ashktorab. 2023. "Microbiomic and Metabolomic Analyses Unveil the Protective Effect of Saffron in a Mouse Colitis Model" Current Issues in Molecular Biology 45, no. 7: 5558-5574. https://doi.org/10.3390/cimb45070351
APA StyleSingh, G., Brim, H., Haileselassie, Y., Varma, S., Habtezion, A., Rashid, M., Sinha, S. R., & Ashktorab, H. (2023). Microbiomic and Metabolomic Analyses Unveil the Protective Effect of Saffron in a Mouse Colitis Model. Current Issues in Molecular Biology, 45(7), 5558-5574. https://doi.org/10.3390/cimb45070351