
Cytochalasin B-Induced Membrane Vesicles from TRAIL-Overexpressing Mesenchymal Stem Cells Induce Extrinsic Pathway of Apoptosis in Breast Cancer Mouse Model
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Culture Conditions
2.2. Lentivirus Production
2.3. Genetic Modification and Selection
2.4. Quantitative Polymerase Chain Reaction (qPCR)
2.5. Western Blot Analysis
2.6. Confirmation of TRAIL Protein Membrane Localization
2.7. Immunophenotyping of MSCs
2.8. Cell Viability Assay
2.9. Cell Proliferation Assay
2.10. Analysis of the Cytokine Profile in the Conditioned Medium
2.11. Isolation of Cytochalasin B-Induced Membrane Vesicles
2.12. Assessment of TRAIL mRNA and Protein Inside and on the Surface of the CIMVs
2.13. Analysis of Typical Extracellular Vesicle Markers in CIMVs
2.14. Determination of CIMV Size
2.15. In Vitro Assessment of CIMV-TRAIL Anti-Cancer Activity
2.16. Analysis of Changes in the Expression of Apoptotic Regulator Genes in Tumor Cells after Interaction with CIMVs
2.17. Animals
2.18. Creation of Breast Cancer Mouse Tumor Model
2.19. Treatment of Mice with CIMVs
2.20. Histological Analysis of the Tumors after CIMV Administration
2.21. Analysis of Induction of Apoptosis in Tumor Cells after CIMV Administration
2.22. Statistical Analysis
3. Results
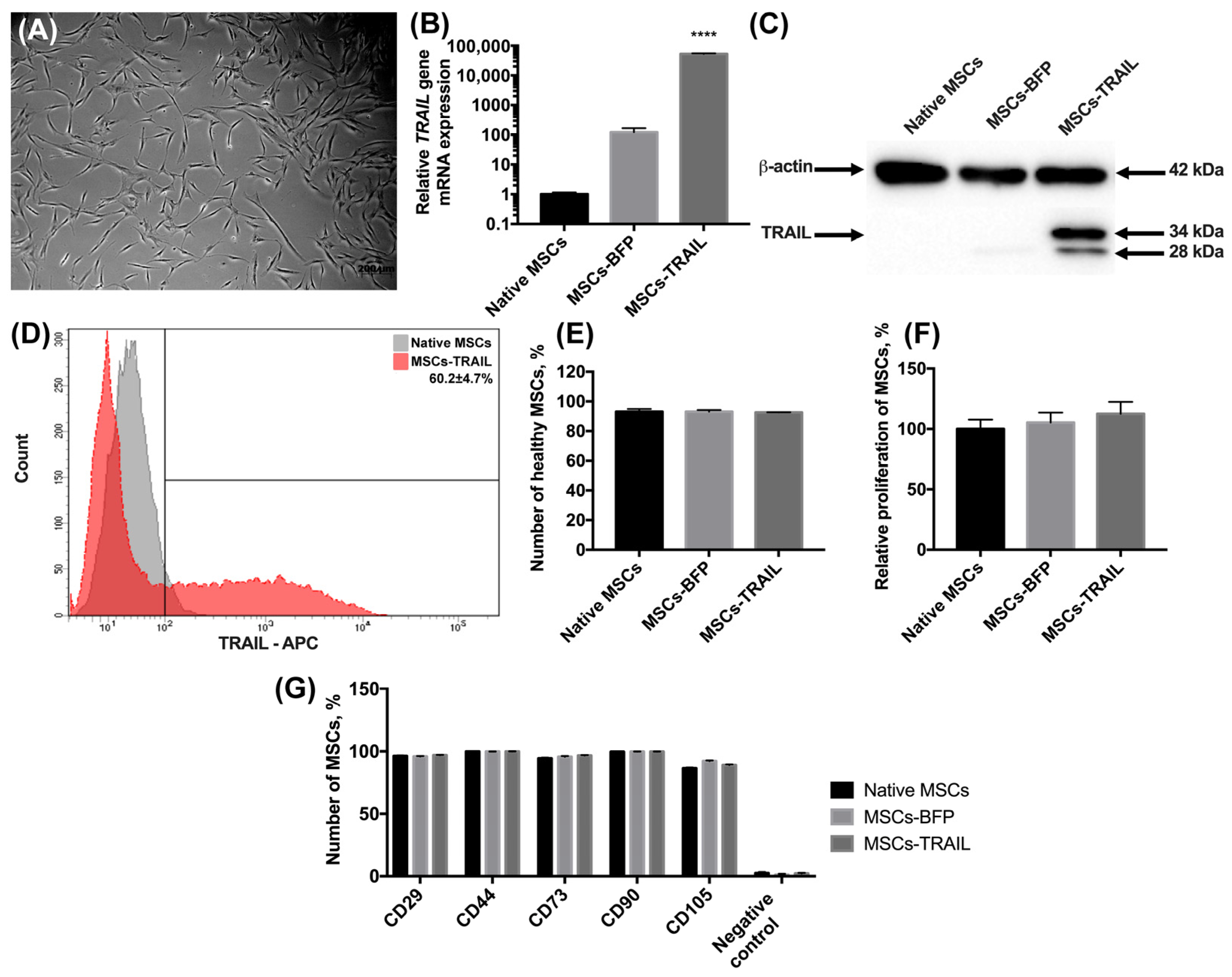
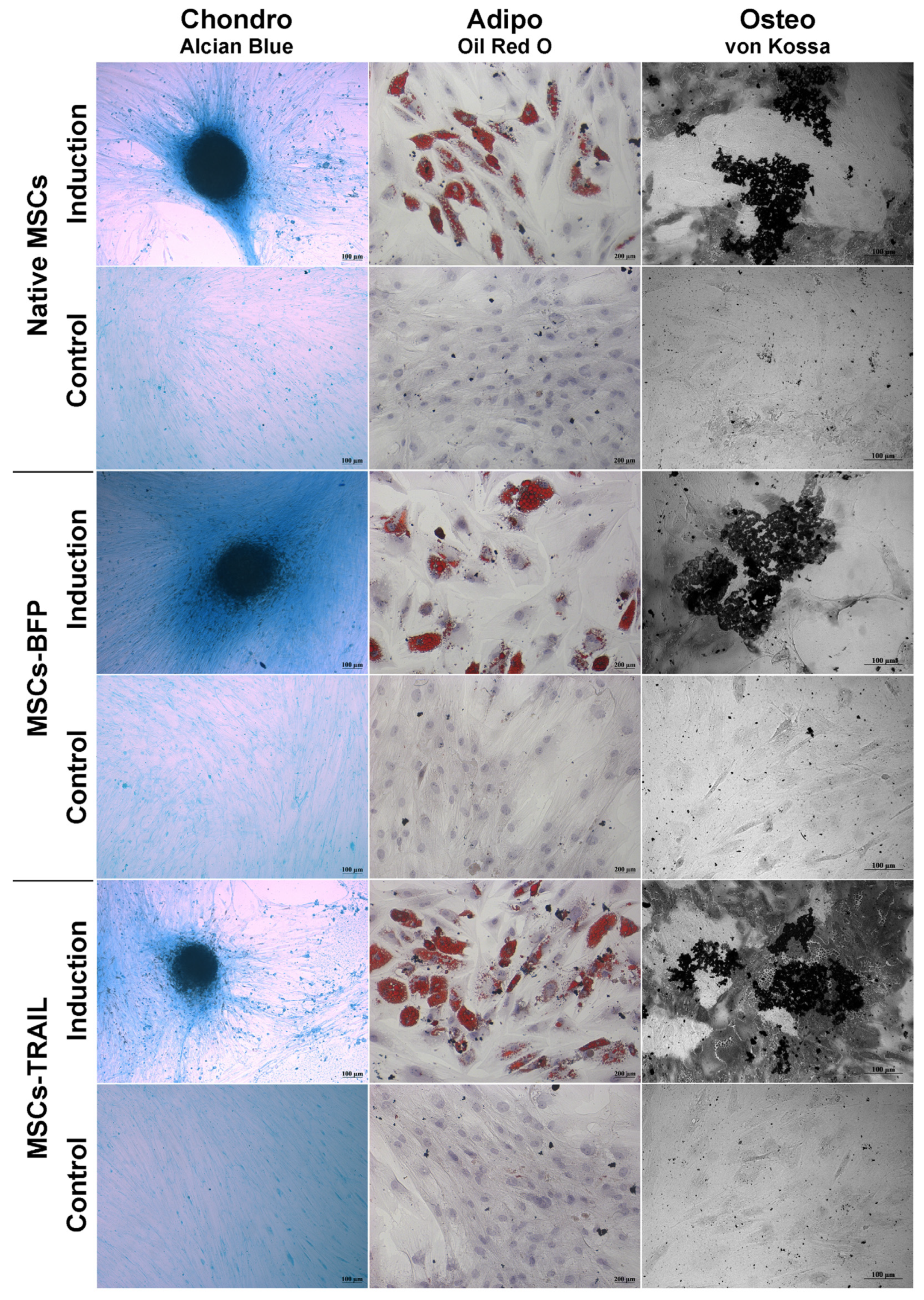
3.1. Genetically Modified MSCs-TRAIL Remain Properties of Stem Cells
3.2. TRAIL Overexpression Results in the Significant Decrease of Pro-Inflammatory Cytokine Secretion
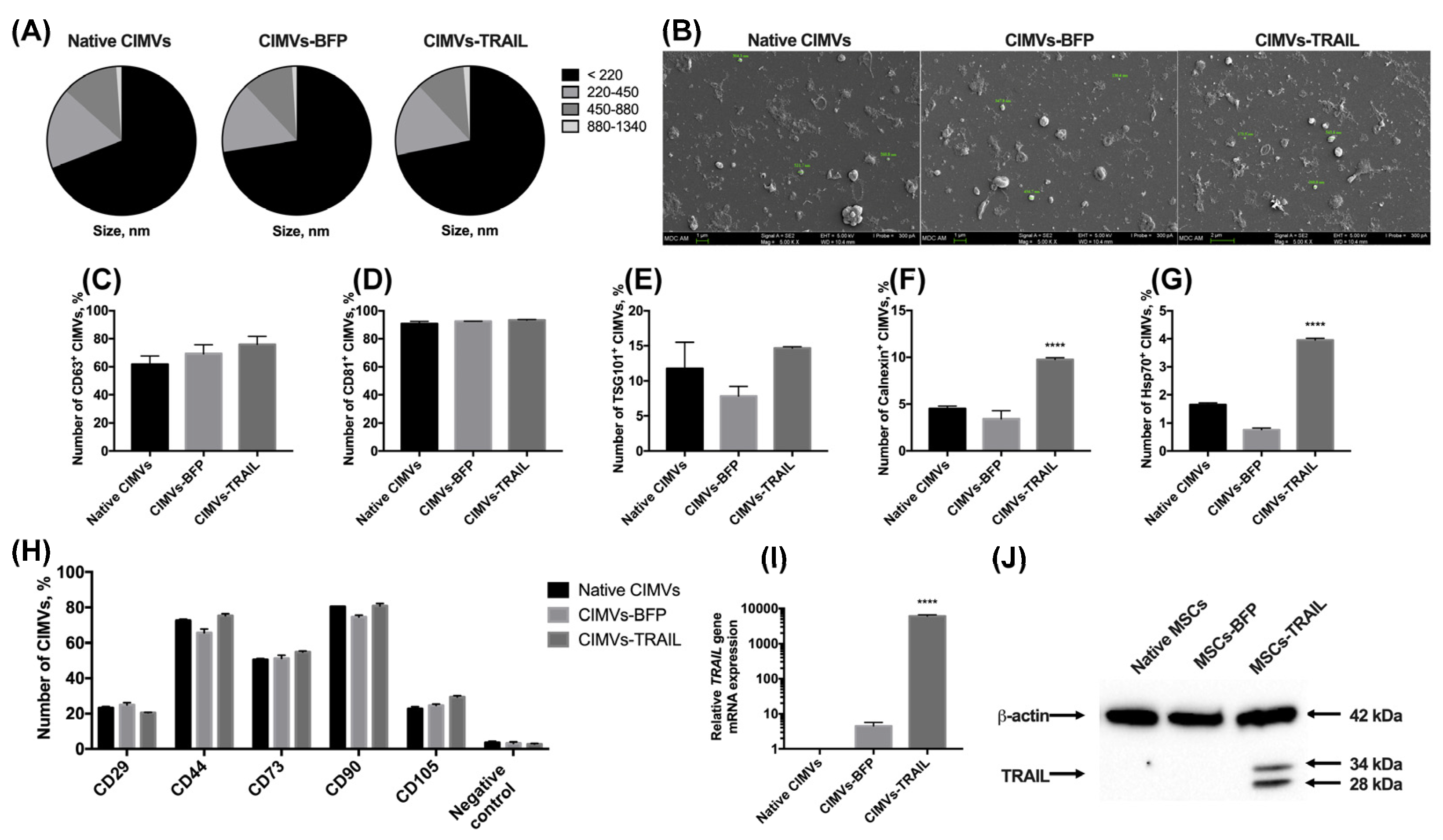
3.3. CIMVs Isolated from MSCs-TRAIL Carry TRAIL Protein
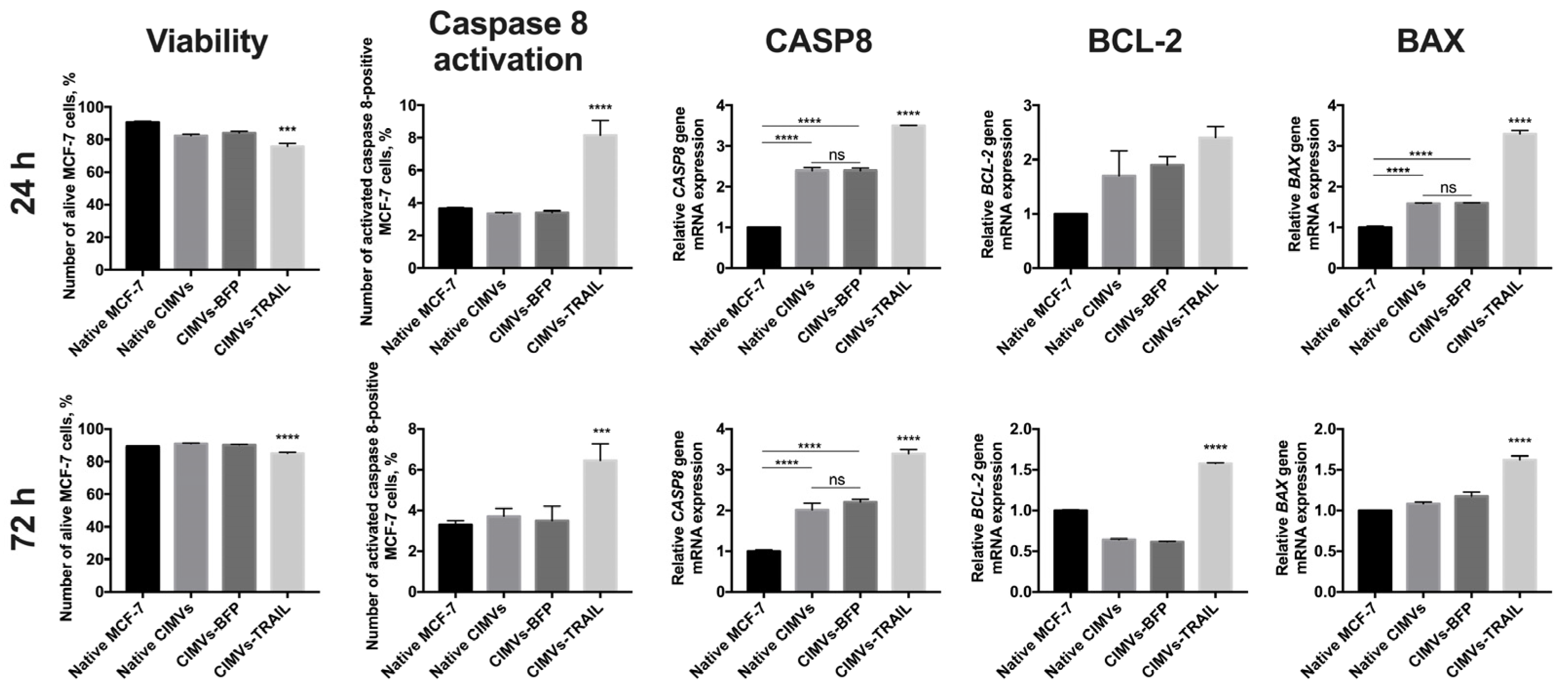
3.4. CIMVs-TRAIL Induce Apoptosis in MCF-7 Cells In Vitro
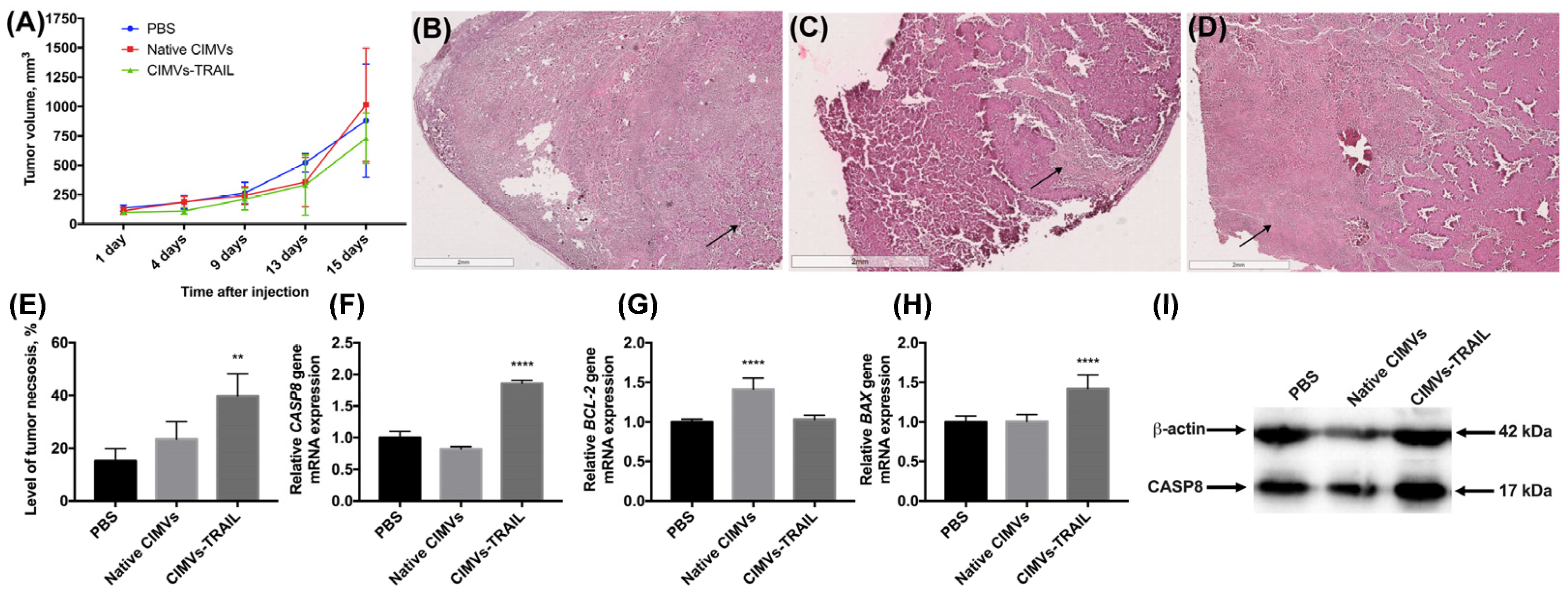
3.5. CIMVs-TRAIL Mediate Tumor Cell Death in breast Cancer Mouse Model In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashkenazi, A. Targeting the extrinsic apoptotic pathway in cancer: Lessons learned and future directions. J. Clin. Investig. 2015, 125, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Thapa, B.; Kc, R.; Uludag, H. TRAIL therapy and prospective developments for cancer treatment. J. Control Release 2020, 326, 335–349. [Google Scholar] [CrossRef]
- Snajdauf, M.; Havlova, K.; Vachtenheim, J., Jr.; Ozaniak, A.; Lischke, R.; Bartunkova, J.; Smrz, D.; Strizova, Z. The TRAIL in the Treatment of Human Cancer: An Update on Clinical Trials. Front. Mol. Biosci. 2021, 8, 628332. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Eckhardt, S.G.; Kurzrock, R.; Ebbinghaus, S.; O’Dwyer, P.J.; Gordon, M.S.; Novotny, W.; Goldwasser, M.A.; Tohnya, T.M.; Lum, B.L.; et al. Phase I dose-escalation study of recombinant human Apo2L/TRAIL, a dual proapoptotic receptor agonist, in patients with advanced cancer. J. Clin. Oncol. 2010, 28, 2839–2846. [Google Scholar] [CrossRef] [PubMed]
- Tolcher, A.W.; Mita, M.; Meropol, N.J.; von Mehren, M.; Patnaik, A.; Padavic, K.; Hill, M.; Mays, T.; McCoy, T.; Fox, N.L.; et al. Phase I pharmacokinetic and biologic correlative study of mapatumumab, a fully human monoclonal antibody with agonist activity to tumor necrosis factor-related apoptosis-inducing ligand receptor-1. J. Clin. Oncol. 2007, 25, 1390–1395. [Google Scholar] [CrossRef]
- Von Pawel, J.; Harvey, J.H.; Spigel, D.R.; Dediu, M.; Reck, M.; Cebotaru, C.L.; Humphreys, R.C.; Gribbin, M.J.; Fox, N.L.; Camidge, D.R. Phase II trial of mapatumumab, a fully human agonist monoclonal antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 1 (TRAIL-R1), in combination with paclitaxel and carboplatin in patients with advanced non-small-cell lung cancer. Clin. Lung Cancer 2014, 15, 188–196 e182. [Google Scholar] [CrossRef]
- Trarbach, T.; Moehler, M.; Heinemann, V.; Kohne, C.H.; Przyborek, M.; Schulz, C.; Sneller, V.; Gallant, G.; Kanzler, S. Phase II trial of mapatumumab, a fully human agonistic monoclonal antibody that targets and activates the tumour necrosis factor apoptosis-inducing ligand receptor-1 (TRAIL-R1), in patients with refractory colorectal cancer. Br. J. Cancer 2010, 102, 506–512. [Google Scholar] [CrossRef]
- Soria, J.C.; Mark, Z.; Zatloukal, P.; Szima, B.; Albert, I.; Juhasz, E.; Pujol, J.L.; Kozielski, J.; Baker, N.; Smethurst, D.; et al. Randomized phase II study of dulanermin in combination with paclitaxel, carboplatin, and bevacizumab in advanced non-small-cell lung cancer. J. Clin. Oncol. 2011, 29, 4442–4451. [Google Scholar] [CrossRef]
- Younes, A.; Vose, J.M.; Zelenetz, A.D.; Smith, M.R.; Burris, H.A.; Ansell, S.M.; Klein, J.; Halpern, W.; Miceli, R.; Kumm, E.; et al. A Phase 1b/2 trial of mapatumumab in patients with relapsed/refractory non-Hodgkin’s lymphoma. Br. J. Cancer 2010, 103, 1783–1787. [Google Scholar] [CrossRef]
- Kindler, H.L.; Richards, D.A.; Garbo, L.E.; Garon, E.B.; Stephenson, J.J., Jr.; Rocha-Lima, C.M.; Safran, H.; Chan, D.; Kocs, D.M.; Galimi, F.; et al. A randomized, placebo-controlled phase 2 study of ganitumab (AMG 479) or conatumumab (AMG 655) in combination with gemcitabine in patients with metastatic pancreatic cancer. Ann. Oncol. 2012, 23, 2834–2842. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Shah, K. TRAIL of Hope Meeting Resistance in Cancer. Trends Cancer 2020, 6, 989–1001. [Google Scholar] [CrossRef]
- Chulpanova, D.S.; Solovyeva, V.V.; James, V.; Arkhipova, S.S.; Gomzikova, M.O.; Garanina, E.E.; Akhmetzyanova, E.R.; Tazetdinova, L.G.; Khaiboullina, S.F.; Rizvanov, A.A. Human Mesenchymal Stem Cells Overexpressing Interleukin 2 Can Suppress Proliferation of Neuroblastoma Cells in Co-Culture and Activate Mononuclear Cells In Vitro. Bioengineering 2020, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.A.; Hwang, S.K.; Wang, K.C.; Cho, B.K.; Phi, J.H.; Lee, J.Y.; Jung, H.W.; Lee, D.H.; Kim, S.K. Therapeutic efficacy and safety of TRAIL-producing human adipose tissue-derived mesenchymal stem cells against experimental brainstem glioma. Neuro Oncol. 2011, 13, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Guiho, R.; Biteau, K.; Grisendi, G.; Taurelle, J.; Chatelais, M.; Gantier, M.; Heymann, D.; Dominici, M.; Redini, F. TRAIL delivered by mesenchymal stromal/stem cells counteracts tumor development in orthotopic Ewing sarcoma models. Int. J. Cancer 2016, 139, 2802–2811. [Google Scholar] [CrossRef] [PubMed]
- Chulpanova, D.S.; Kitaeva, K.V.; Tazetdinova, L.G.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Application of Mesenchymal Stem Cells for Therapeutic Agent Delivery in Anti-tumor Treatment. Front. Pharmacol. 2018, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Chulpanova, D.S.; Kitaeva, K.V.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Therapeutic Prospects of Extracellular Vesicles in Cancer Treatment. Front. Immunol. 2018, 9, 1534. [Google Scholar] [CrossRef] [PubMed]
- Chulpanova, D.S.; Gilazieva, Z.E.; Kletukhina, S.K.; Aimaletdinov, A.M.; Garanina, E.E.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Cytochalasin B-Induced Membrane Vesicles from Human Mesenchymal Stem Cells Overexpressing IL2 Are Able to Stimulate CD8(+) T-Killers to Kill Human Triple Negative Breast Cancer Cells. Biology 2021, 10, 141. [Google Scholar] [CrossRef]
- Shamili, F.H.; Bayegi, H.R.; Salmasi, Z.; Sadri, K.; Mahmoudi, M.; Kalantari, M.; Ramezani, M.; Abnous, K. Exosomes derived from TRAIL-engineered mesenchymal stem cells with effective anti-tumor activity in a mouse melanoma model. Int. J. Pharm. 2018, 549, 218–229. [Google Scholar] [CrossRef]
- Ke, C.; Hou, H.; Li, J.; Su, K.; Huang, C.; Lin, Y.; Lu, Z.; Du, Z.; Tan, W.; Yuan, Z. Extracellular Vesicle Delivery of TRAIL Eradicates Resistant Tumor Growth in Combination with CDK Inhibition by Dinaciclib. Cancers 2020, 12, 1157. [Google Scholar] [CrossRef]
- Yuan, Z.; Kolluri, K.K.; Gowers, K.H.; Janes, S.M. TRAIL delivery by MSC-derived extracellular vesicles is an effective anticancer therapy. J. Extracell. Vesicles 2017, 6, 1265291. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Yuan, Z.; Kolluri, K.K.; Sage, E.K.; Gowers, K.H.; Janes, S.M. Mesenchymal stromal cell delivery of full-length tumor necrosis factor-related apoptosis-inducing ligand is superior to soluble type for cancer therapy. Cytotherapy 2015, 17, 885–896. [Google Scholar] [CrossRef]
- Szegezdi, E.; O’Reilly, A.; Davy, Y.; Vawda, R.; Taylor, D.L.; Murphy, M.; Samali, A.; Mehmet, H. Stem cells are resistant to TRAIL receptor-mediated apoptosis. J. Cell Mol. Med. 2009, 13, 4409–4414. [Google Scholar] [CrossRef]
- Naval, J.; de Miguel, D.; Gallego-Lleyda, A.; Anel, A.; Martinez-Lostao, L. Importance of TRAIL Molecular Anatomy in Receptor Oligomerization and Signaling. Implications for Cancer Therapy. Cancers 2019, 11, 444. [Google Scholar] [CrossRef]
- Cunningham, H.D.; Shannon, L.A.; Calloway, P.A.; Fassold, B.C.; Dunwiddie, I.; Vielhauer, G.; Zhang, M.; Vines, C.M. Expression of the C-C chemokine receptor 7 mediates metastasis of breast cancer to the lymph nodes in mice. Transl. Oncol. 2010, 3, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Rizeq, B.; Malki, M.I. The Role of CCL21/CCR7 Chemokine Axis in Breast Cancer Progression. Cancers 2020, 12, 1036. [Google Scholar] [CrossRef]
- Koizumi, K.; Kozawa, Y.; Ohashi, Y.; Nakamura, E.S.; Aozuka, Y.; Sakurai, H.; Ichiki, K.; Doki, Y.; Misaki, T.; Saiki, I. CCL21 promotes the migration and adhesion of highly lymph node metastatic human non-small cell lung cancer Lu-99 in vitro. Oncol. Rep. 2007, 17, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Liu, P.; Li, J.; Zhang, Y. Eotaxin-1 promotes prostate cancer cell invasion via activation of the CCR3-ERK pathway and upregulation of MMP-3 expression. Oncol. Rep. 2014, 31, 2049–2054. [Google Scholar] [CrossRef]
- Cao, Z.; Fu, B.; Deng, B.; Zeng, Y.; Wan, X.; Qu, L. Overexpression of Chemokine (C-X-C) ligand 1 (CXCL1) associated with tumor progression and poor prognosis in hepatocellular carcinoma. Cancer Cell Int. 2014, 14, 86. [Google Scholar] [CrossRef]
- Zhang, F.; Jiang, J.; Xu, B.; Xu, Y.; Wu, C. Over-expression of CXCL2 is associated with poor prognosis in patients with ovarian cancer. Medicine 2021, 100, e24125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cai, Y.; Liu, L.; Wu, Y.; Xiong, X. Crucial biological functions of CCL7 in cancer. PeerJ 2018, 6, e4928. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Cao, Y.; Zhang, X.; Zhao, H. Increased CCL19 expression is associated with progression in cervical cancer. Oncotarget 2017, 8, 73817–73825. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Tang, D.; Fan, L.; Gao, W.; Lin, H. CCL25 promotes the migration and invasion of non-small cell lung cancer cells by regulating VEGF and MMPs in a CCR9-dependent manner. Exp. Ther. Med. 2020, 19, 3571–3580. [Google Scholar] [CrossRef] [PubMed]
- Sheikhpour, E.; Noorbakhsh, P.; Foroughi, E.; Farahnak, S.; Nasiri, R.; Neamatzadeh, H. A Survey on the Role of Interleukin-10 in Breast Cancer: A Narrative. Rep. Biochem. Mol. Biol. 2018, 7, 30–37. [Google Scholar]
- Liu, L.Z.; Zhang, Z.; Zheng, B.H.; Shi, Y.; Duan, M.; Ma, L.J.; Wang, Z.C.; Dong, L.Q.; Dong, P.P.; Shi, J.Y.; et al. CCL15 Recruits Suppressive Monocytes to Facilitate Immune Escape and Disease Progression in Hepatocellular Carcinoma. Hepatology 2019, 69, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhang, X.; Dong, Y.; Liu, J.; Zhang, W.; Wu, F.; Bo, H.; Shao, H.; Zhang, R.; Shen, H. Chemokine CCL17 Affects Local Immune Infiltration Characteristics and Early Prognosis Value of Lung Adenocarcinoma. Front. Cell Dev. Biol. 2022, 10, 816927. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, H.; Sun, M.; Deng, X.; Wu, X.; Ma, Y.; Li, M.; Shuoa, S.M.; You, Q.; Miao, L. CXCL5/CXCR2 axis in tumor microenvironment as potential diagnostic biomarker and therapeutic target. Cancer Commun. 2020, 40, 69–80. [Google Scholar] [CrossRef]
- Mantovani, A.; Barajon, I.; Garlanda, C. IL-1 and IL-1 regulatory pathways in cancer progression and therapy. Immunol. Rev. 2018, 281, 57–61. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, F.; Li, G.; Li, G.; Yang, X.; Liu, L.; Zhang, R.; Zhang, B.; Feng, Y. Human colorectal cancer-derived mesenchymal stem cells promote colorectal cancer progression through IL-6/JAK2/STAT3 signaling. Cell Death Dis. 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhou, Y.; Li, W.; Zhang, B.; Zhang, H.; Zhao, S.; Zheng, P.; Wu, H.; Yang, J. Tumor-derived mesenchymal-stem-cell-secreted IL-6 enhances resistance to cisplatin via the STAT3 pathway in breast cancer. Oncol. Lett. 2018, 15, 9142–9150. [Google Scholar] [CrossRef]
- Rivoltini, L.; Chiodoni, C.; Squarcina, P.; Tortoreto, M.; Villa, A.; Vergani, B.; Burdek, M.; Botti, L.; Arioli, I.; Cova, A.; et al. TNF-Related Apoptosis-Inducing Ligand (TRAIL)-Armed Exosomes Deliver Proapoptotic Signals to Tumor Site. Clin. Cancer Res. 2016, 22, 3499–3512. [Google Scholar] [CrossRef] [PubMed]
- Khatri, M.; Richardson, L.A.; Meulia, T. Mesenchymal stem cell-derived extracellular vesicles attenuate influenza virus-induced acute lung injury in a pig model. Stem Cell Res. Ther. 2018, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Campos-Silva, C.; Suarez, H.; Jara-Acevedo, R.; Linares-Espinos, E.; Martinez-Pineiro, L.; Yanez-Mo, M.; Vales-Gomez, M. High sensitivity detection of extracellular vesicles immune-captured from urine by conventional flow cytometry. Sci. Rep. 2019, 9, 2042. [Google Scholar] [CrossRef]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, B. TRAIL resistance of breast cancer cells is associated with constitutive endocytosis of death receptors 4 and 5. Mol. Cancer Res. 2008, 6, 1861–1871. [Google Scholar] [CrossRef]
- Mueller, L.P.; Luetzkendorf, J.; Widder, M.; Nerger, K.; Caysa, H.; Mueller, T. TRAIL-transduced multipotent mesenchymal stromal cells (TRAIL-MSC) overcome TRAIL resistance in selected CRC cell lines in vitro and in vivo. Cancer Gene Ther. 2011, 18, 229–239. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, H.; Wang, P.; Zhou, M.; Li, G.; Hu, Z.; Hu, Q.; Zhao, J.; Liu, X.; Wu, L.; et al. Site-Specific Integration of TRAIL in iPSC-Derived Mesenchymal Stem Cells for Targeted Cancer Therapy. Stem Cells Transl. Med. 2022, 11, 297–309. [Google Scholar] [CrossRef]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Wieder, T.; Sturm, I.; Daniel, P.T.; Orfanos, C.E.; Geilen, C.C. The Bax/Bcl-2 ratio determines the susceptibility of human melanoma cells to CD95/Fas-mediated apoptosis. J. Investig. Dermatol. 2001, 117, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Salakou, S.; Kardamakis, D.; Tsamandas, A.C.; Zolota, V.; Apostolakis, E.; Tzelepi, V.; Papathanasopoulos, P.; Bonikos, D.S.; Papapetropoulos, T.; Petsas, T.; et al. Increased Bax/Bcl-2 ratio up-regulates caspase-3 and increases apoptosis in the thymus of patients with myasthenia gravis. In Vivo 2007, 21, 123–132. [Google Scholar]
- Zhong, B.; Liu, M.; Bai, C.; Ruan, Y.; Wang, Y.; Qiu, L.; Hong, Y.; Wang, X.; Li, L.; Li, B. Caspase-8 Induces Lysosome-Associated Cell Death in Cancer Cells. Mol. Ther. 2020, 28, 1078–1091. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.; Lin, F.; Ding, R.; Wang, W.; Hong, W. Extracellular vesicles carrying miR-193a derived from mesenchymal stem cells impede cell proliferation, migration and invasion of colon cancer by downregulating FAK. Exp. Cell Res. 2020, 394, 112144. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Wang, C.; He, C.; Chen, L. Bone mesenchymal stem cells derived extracellular vesicles promote TRAIL-related apoptosis of hepatocellular carcinoma cells via the delivery of microRNA-20a-3p. Cancer Biomark. 2021, 30, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Spano, C.; Grisendi, G.; Golinelli, G.; Rossignoli, F.; Prapa, M.; Bestagno, M.; Candini, O.; Petrachi, T.; Recchia, A.; Miselli, F.; et al. Soluble TRAIL Armed Human MSC As Gene Therapy for Pancreatic Cancer. Sci. Rep. 2019, 9, 1788. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, S.; Ma, T.; Zeng, J.; Zhou, X.; Li, H.; Tang, M.; Liu, X.; Li, F.; Jiang, B.; et al. Secreted TRAIL gene-modified adipose-derived stem cells exhibited potent tumor-suppressive effect in hepatocellular carcinoma cells. Immun. Inflamm. Dis. 2021, 9, 144–156. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward Primer (5′−3′) | Reverse Primer (5′−3′) | TaqMan Probe (5′−3′) |
|---|---|---|---|
| 18S rRNA | GCCGCTAGAGGTGAAATTCTTG | CATTCTTGGCAAATGCTTTCG | [HEX] ACCGGCGCAAGACGGACCAG [BH2] |
| TRAIL | GGCACATGCCTGTAGTCCCA | GCCAGGCTGGAGTGTAGTGG | [FAM] CCACACTGCAACCTCTGCCTCCCGGG [BH1] |
| CASP8 | CTGGACAGTGAAGATCTGGC | GATGGGTTCTTGCTTCCTTTG | [FAM] CCTGAGCCTGGACTACATTCCGC [BH1] |
| BCL-2 | GTGGATGACTGAGTACCTGAAC | GCCAGGAGAAATCAAACAGAGG | [FAM] CAGGATAACGGAGGCTGGGATGC [BH1] |
| BAX | GACATGTTTTCTGACGGCAAC | AAGTCCAATGTCCAGCCC | [FAM] CTGGCAAAGTAGAAAAGGGCGACAAC [BH1] |
| Cytokine | Concentration, pg/µL | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | |||||||
| Native MScs | MSCs-BFP | MSCs-TRAIL | Native MScs | MSCs-BFP | MSCs-TRAIL | Native MScs | MSCs-BFP | MSCs-TRAIL | |
| CCL21 | 40.1 ± 2.1 | 37.7 ± 0.3 | 32.7 ± 0.3 | 44.0 ± 0.08 | 38.0 ± 1.3 | 35.2 ± 2.5 | 45.4 ± 0.97 | 43.3 ± 0.1 | 34.1 ± 0.4 |
| CXCL5 | 803.7 ± 115.5 | 691.0 ± 16.4 | 418.6 ± 11.4 | 897.2 ± 44.7 | 563.6 ± 65.2 | 418.1 ± 12.2 | 547.9 ± 16.4 | 545.5 ± 4.8 | 359.9 ± 16.3 |
| Eotaxin1 (CCL11) | 25.5 ± 2.2 | 22.7 ± 0.3 | 18.8 ± 0.3 | 24.5 ± 2.0 | 21.1 ± 0.5 | 18.7 ± 0.1 | 29.1 ± 2.5 | 24.2 1.5 | 19.1 0.7 |
| Gro-α (CXCL1) | 1310.1 ± 1.7 | 640.0 ± 6.3 | 307.4 ± 17.7 | 3624.8 ± 98.0 | 532.2 ± 3.8 | 454.1 ± 3.0 | 885.7 ± 14.0 | 906.3 ± 21.0 | 367.1 ± 14.4 |
| Gro-β (CXCL2) | 18.4 ± 0.2 | 10.86 ± 0.09 | 5.4 ± 0.5 | 14.0 ± 0.1 | 7.33 ± 0.1 | 6.7 ± 0.5 | 8.0 ± 0.4 | 8.0 ± 0.2 | 6.2 ± 0.6 |
| IL1β | 5.8 ± 0.3 | 5.0 ± 0.2 | 3.0 ± 0.4 | 5.2 ± 0.2 | 5.3 ± 0.2 | 3.5 ± 0.2 | 9.2 ± 0.1 | 8.9 ± 0.5 | 4.0 ± 0.2 |
| IL6 | 27346.1 ± 2595.5 | 24060.0 ± 783.7 | 6184.8 ± 98.8 | 12142.9 ± 1675.6 | 9242.1 ± 21.7 | 9099.9 ± 181.0 | 27925.7 ± 5796.7 | 28071.0 ± 1435.16 | 14239.3 ± 583.6 |
| IL8 | 11666.5 ± 496.4 | 9857.5 ± 3.7 | 2189.0 ± 22.0 | 11314.3 ± 250.0 | 3838.4 ± 7.6 | 1637.6 ± 212.6 | 2158.4 ± 104.6 | 2038.0 ± 10.7 | 672.7 ± 19.0 |
| IL10 | 26.7 ± 2.1 | 24.5 ± 1.5 | 17.8 ± 0.7 | 29.0 ± 1.0 | 20.2 ± 0.2 | 17.3 ± 0.04 | 18.7 ± 1.2 | 18.7 ± 0.2 | 12.0 ± 0.7 |
| MCP-3 (CCL7) | 30.4 ± 0.9 | 28.7 ± 0.4 | 16.23 ± 0.0 | 30.5 ± 0.2 | 25.0 ± 1.9 | 19.6 ± 0.5 | 30.7 ± 0.4 | 28.6 ± 0.7 | 22.5 ± 3.5 |
| MCP-4 (CCL13) | 29.8 ± 1.1 | 19.7 ± 0.6 | 12.6 ± 0.5 | 46.5 ± 1.7 | 15.7 ± 0.8 | 13.5 ± 0.6 | 19.7 ± 1.0 | 18.9 ± 0.4 | 9.4 ± 1.0 |
| MIP1β (CCL15) | 5.3 ± 0.1 | 4.4 ± 0.3 | 2.6 ± 0.5 | 5.9 ± 0.8 | 3.5 ± 0.3 | 2.8 ± 0.2 | 4.4 ± 0.04 | 4.2 ± 0.09 | 2.2 ± 0.5 |
| MIP3-β (CCL19) | 12.5 ± 1.8 | 10.3 ± 0.6 | 8.1 ± 0.6 | 12.6 ± 1.0 | 8.9 ± 0.09 | 7.95 ± 0.06 | 10.1 ± 1.1 | 9.0 ± 0.4 | 7.1 ± 0.1 |
| MPIF-1 (CCL23) | 7.9 ± 0.1 | 7.8 ± 0.8 | 5.4 ± 0.1 | 7.4 ± 0.7 | 7.1 ± 0.3 | 5.7 ± 0.8 | 7.8 ± 0.5 | 7.9 ± 0.1 | 5.8 ± 0.1 |
| CCL17 | 17.6 ± 0.1 | 16.8 ± 0.3 | 13.2 ± 0.2 | 16.1 ± 0.4 | 15.8 ± 0.5 | 14.2 ± 0.7 | 18.1 ± 0.8 | 17.6 ± 0.7 | 14.3 ± 0.2 |
| CCL25 | 93.5 ± 0.5 | 83.2 ± 1.7 | 56.9 ± 0.1 | 84.7 ± 2.4 | 75.5 ± 3.6 | 65.3 ± 5.7 | 89.4 ± 1.7 | 87.1 ± 1.5 | 67.9 ± 2.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chulpanova, D.S.; Pukhalskaia, T.V.; Gilazieva, Z.E.; Filina, Y.V.; Mansurova, M.N.; Rizvanov, A.A.; Solovyeva, V.V. Cytochalasin B-Induced Membrane Vesicles from TRAIL-Overexpressing Mesenchymal Stem Cells Induce Extrinsic Pathway of Apoptosis in Breast Cancer Mouse Model. Curr. Issues Mol. Biol. 2023, 45, 571-592. https://doi.org/10.3390/cimb45010038
Chulpanova DS, Pukhalskaia TV, Gilazieva ZE, Filina YV, Mansurova MN, Rizvanov AA, Solovyeva VV. Cytochalasin B-Induced Membrane Vesicles from TRAIL-Overexpressing Mesenchymal Stem Cells Induce Extrinsic Pathway of Apoptosis in Breast Cancer Mouse Model. Current Issues in Molecular Biology. 2023; 45(1):571-592. https://doi.org/10.3390/cimb45010038
Chicago/Turabian StyleChulpanova, Daria S., Tamara V. Pukhalskaia, Zarema E. Gilazieva, Yuliya V. Filina, Milana N. Mansurova, Albert A. Rizvanov, and Valeriya V. Solovyeva. 2023. "Cytochalasin B-Induced Membrane Vesicles from TRAIL-Overexpressing Mesenchymal Stem Cells Induce Extrinsic Pathway of Apoptosis in Breast Cancer Mouse Model" Current Issues in Molecular Biology 45, no. 1: 571-592. https://doi.org/10.3390/cimb45010038
APA StyleChulpanova, D. S., Pukhalskaia, T. V., Gilazieva, Z. E., Filina, Y. V., Mansurova, M. N., Rizvanov, A. A., & Solovyeva, V. V. (2023). Cytochalasin B-Induced Membrane Vesicles from TRAIL-Overexpressing Mesenchymal Stem Cells Induce Extrinsic Pathway of Apoptosis in Breast Cancer Mouse Model. Current Issues in Molecular Biology, 45(1), 571-592. https://doi.org/10.3390/cimb45010038