
Antioxidant, Anti-Apoptotic, and Anti-Inflammatory Effects of Farrerol in a Mouse Model of Obstructive Uropathy


Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Assessment of Renal Function and Cytokine Levels
2.3. Histological Analysis, Immunohistochemical (IHC) Staining, and Immunofluorescence (IF) Staining
2.4. Western Blotting
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Evaluation of Oxidative Stress and Antioxidant Enzyme Activities
2.7. TdT-Mediated dUTP Nick End Labeling (TUNEL) Assay
2.8. Statistical Analysis
3. Results
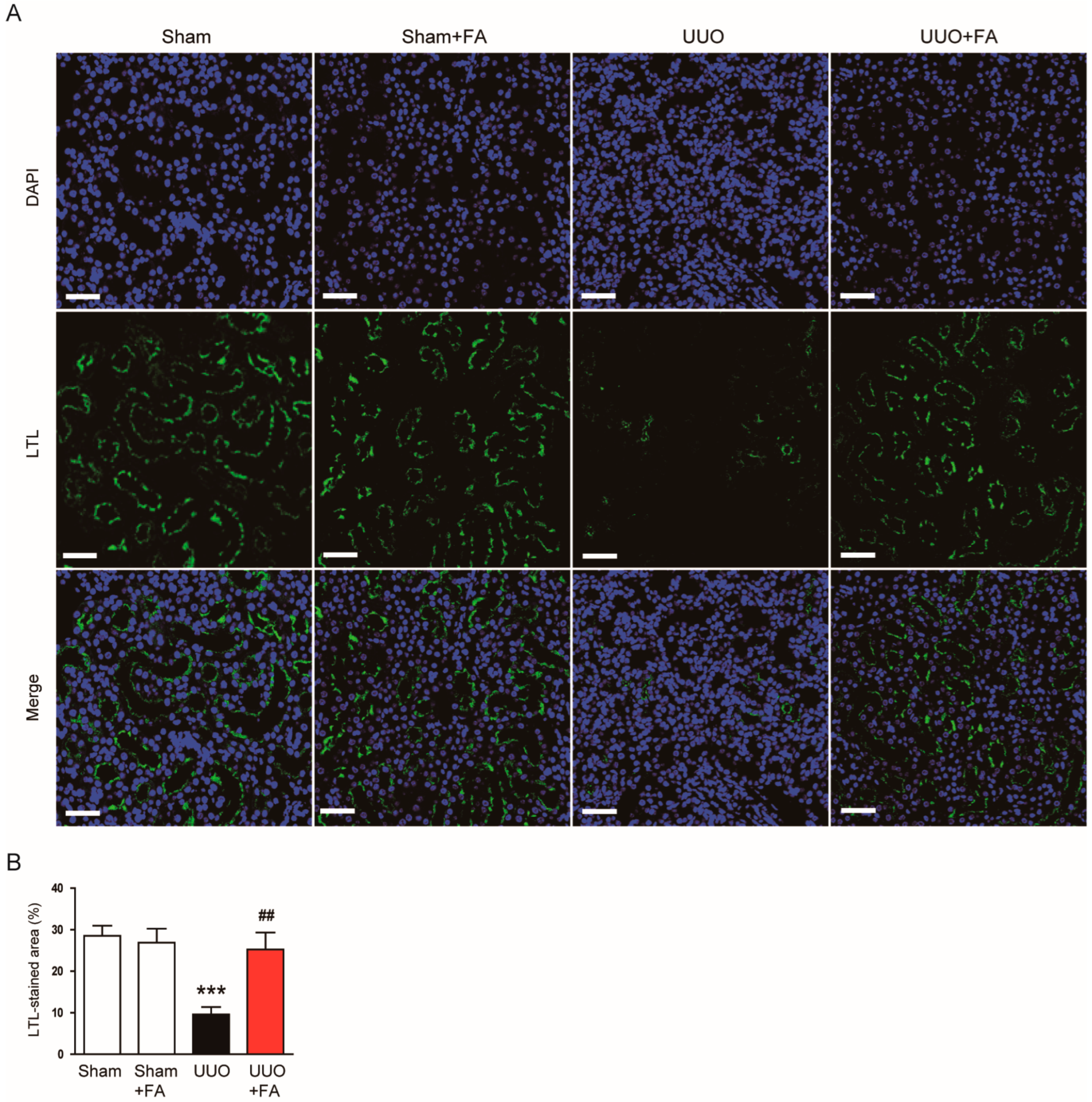
3.1. FA Improved Renal Function and Attenuated Tubular Injury in UUO Mice
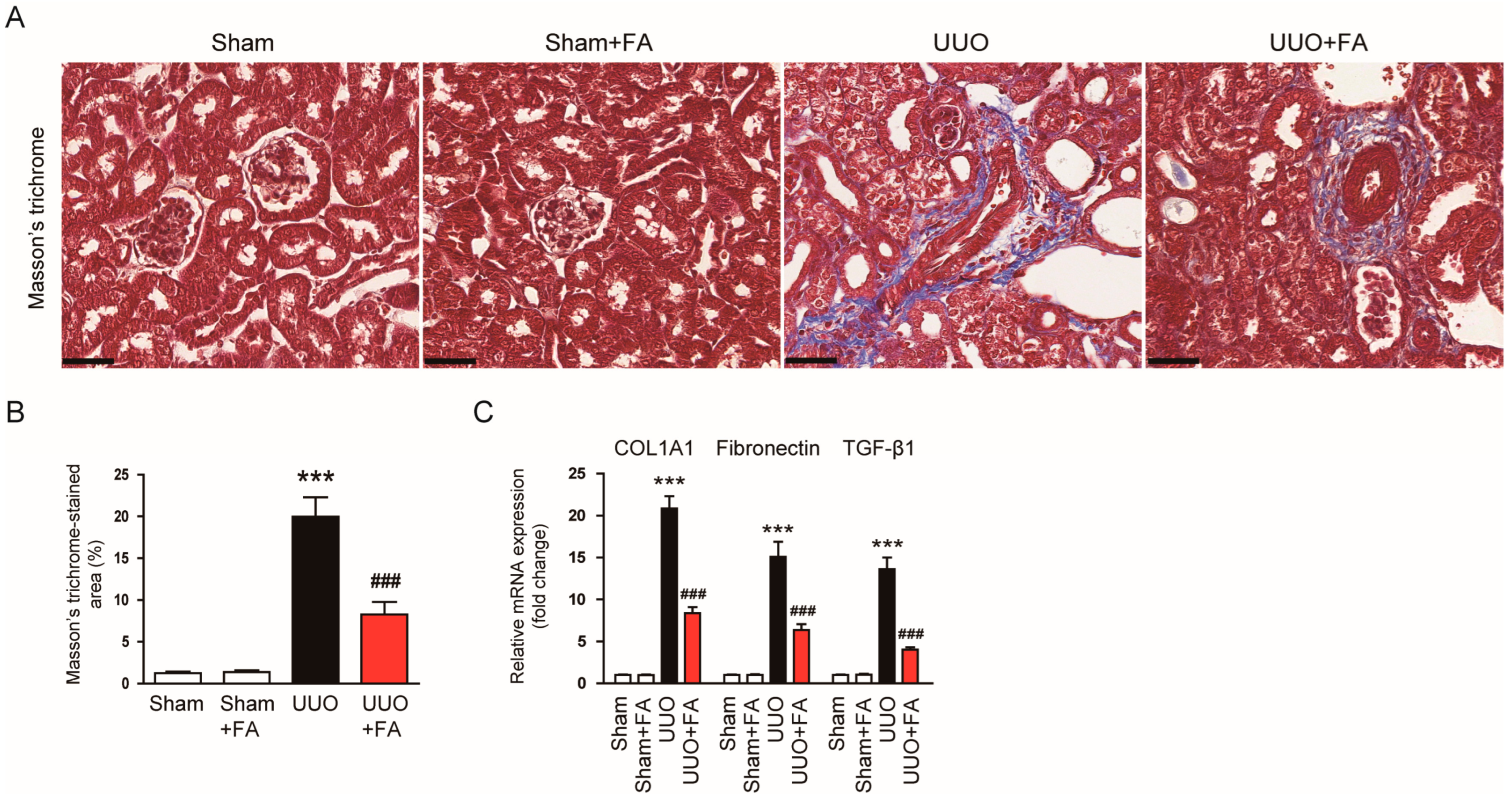
3.2. FA Attenuated Renal Fibrosis in UUO Mice
3.3. FA Suppressed Oxidative Damage in UUO Mice
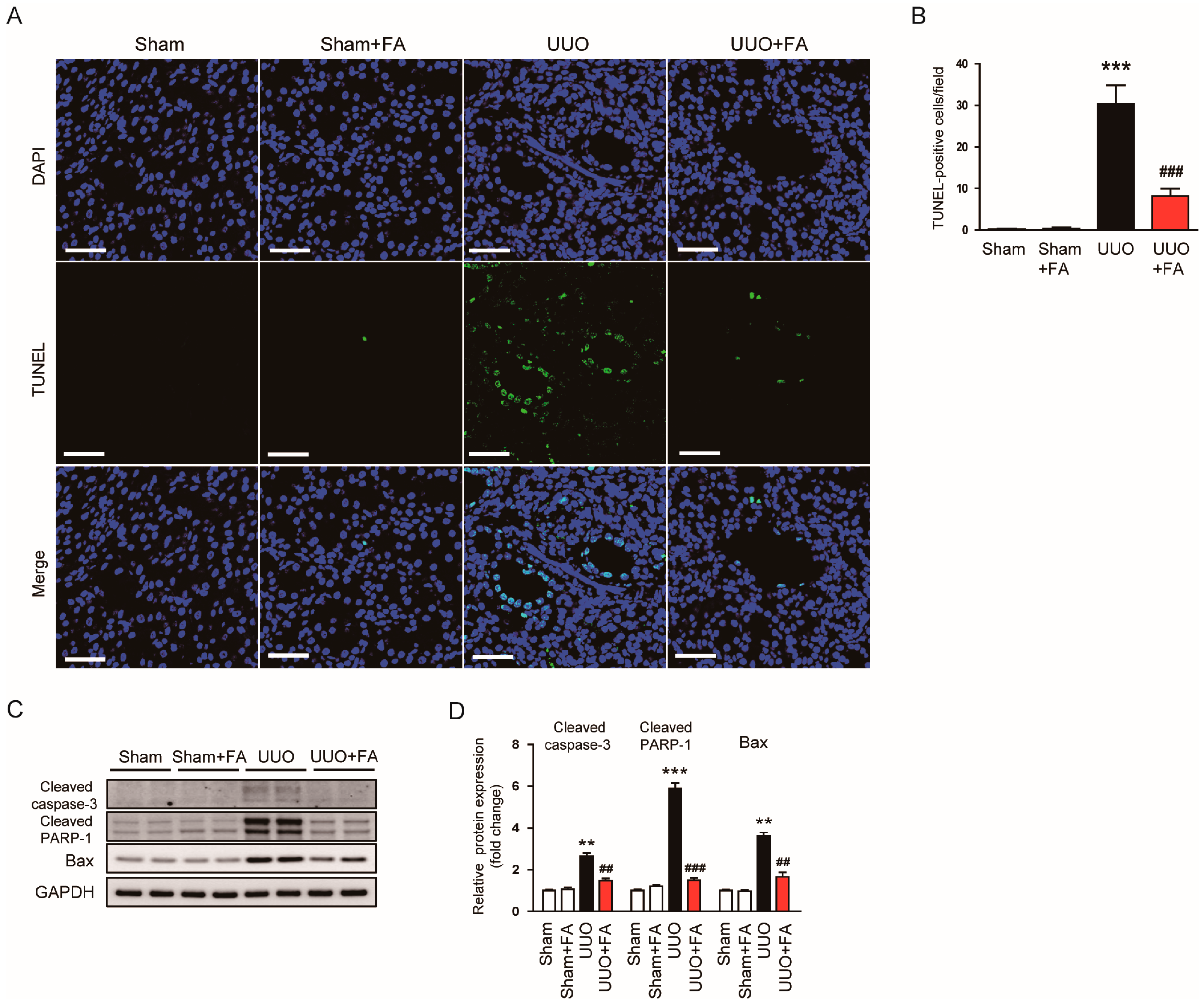
3.4. FA Inhibited Apoptotic Cell Death in UUO Mice
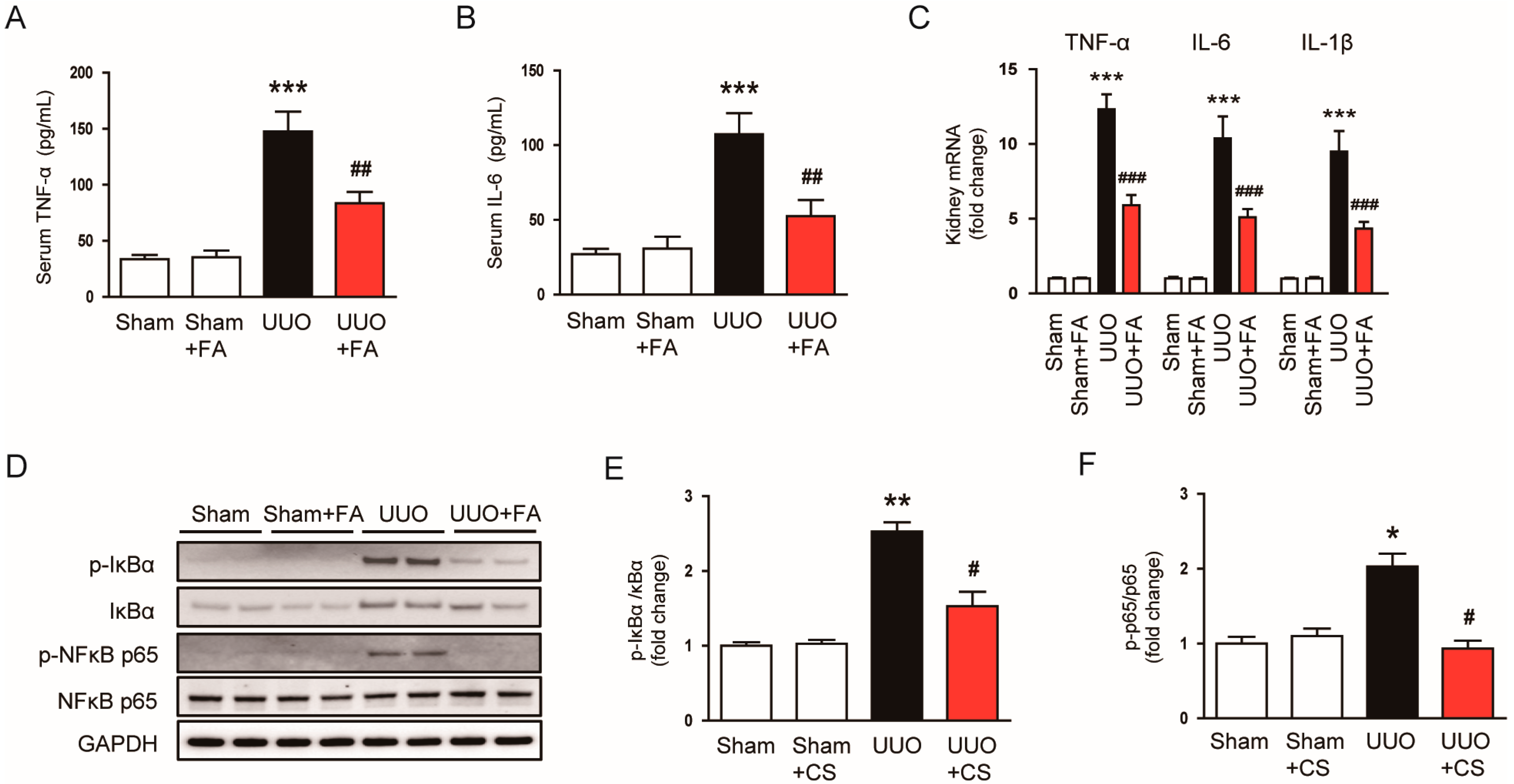
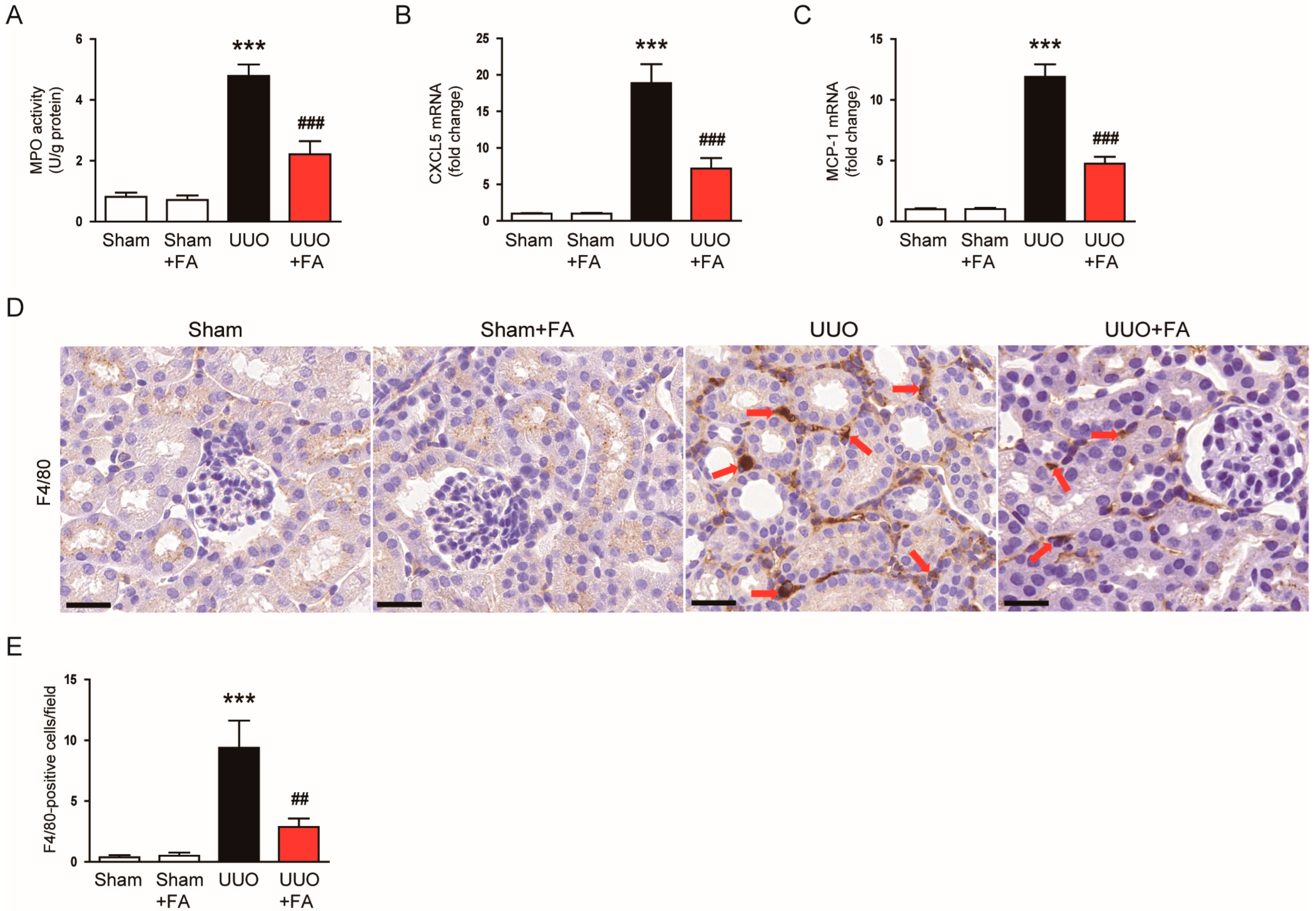
3.5. FA Suppressed Inflammatory Responses in UUO Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tseng, T.Y.; Stoller, M.L. Obstructive uropathy. Clin. Geriatr. Med. 2009, 25, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Benfield, M.R.; McDonald, R.A.; Bartosh, S.; Ho, P.L.; Harmon, W. Changing trends in pediatric transplantation: 2001 Annual Report of the North American Pediatric Renal Transplant Cooperative Study. Pediatr. Transplant. 2003, 7, 321–335. [Google Scholar] [CrossRef]
- Ito, K.; Chen, J.; El Chaar, M.; Stern, J.M.; Seshan, S.V.; Khodadadian, J.J.; Richardson, I.; Hyman, M.J.; Vaughan, E.D., Jr.; Poppas, D.P.; et al. Renal damage progresses despite improvement of renal function after relief of unilateral ureteral obstruction in adult rats. Am. J. Physiol. Renal Physiol. 2004, 287, F1283–F1293. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.A.; Shokeir, A.A.; Mosbah, A.; Abol-Enein, H.; Barakat, N.; Abou-Bieh, E.; Wafa, E.W. Recoverability of renal function after relief of chronic partial unilateral ureteral obstruction: Study of the effect of angiotensin receptor blocker (losartan). Urology 2010, 75, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Tomino, Y. Pathogenesis and treatment of chronic kidney disease: A review of our recent basic and clinical data. Kidney Blood Press. Res. 2014, 39, 450–489. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Tan, R.J.; Liu, Y. Myofibroblast in Kidney Fibrosis: Origin, Activation, and Regulation. Adv. Exp. Med. Biol. 2019, 1165, 253–283. [Google Scholar]
- Kim, J.-Y.; Park, J.-H.; Jeon, E.J.; Leem, J.; Park, K.-K. Melatonin Prevents Transforming Growth Factor-β1-Stimulated Transdifferentiation of Renal Interstitial Fibroblasts to Myofibroblasts by Suppressing Reactive Oxygen Species-Dependent Mechanisms. Antioxidants 2020, 9, 39. [Google Scholar] [CrossRef]
- Braga, P.C.; Alves, M.G.; Rodrigues, A.S.; Oliveira, P.F. Mitochondrial Pathophysiology on Chronic Kidney Disease. Int. J. Mol. Sci. 2022, 23, 1776. [Google Scholar] [CrossRef]
- Galvan, D.L.; Green, N.H.; Danesh, F.R. The hallmarks of mitochondrial dysfunction in chronic kidney disease. Kidney Int. 2017, 92, 1051–1057. [Google Scholar] [CrossRef]
- Qin, X.; Xu, X.; Hou, X.; Liang, R.; Chen, L.; Hao, Y.; Gao, A.; Du, X.; Zhao, L.; Shi, Y.; et al. The pharmacological properties and corresponding mechanisms of farrerol: A comprehensive review. Pharm. Biol. 2022, 60, 9–16. [Google Scholar] [CrossRef]
- Wu, Y.; Qian, J.; Li, K.; Li, W.; Yin, W.; Jiang, H. Farrerol alleviates collagenase-induced tendinopathy by inhibiting ferroptosis in rats. J. Cell. Mol. Med. 2022, 26, 3483–3494. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, S.; Zou, X. Farrerol Alleviates Myocardial Ischemia/Reperfusion Injury by Targeting Macrophages and NLRP3. Front. Pharmacol. 2022, 13, 879232. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Zhang, H.; Shang, C. Farrerol ameliorate adjuvant-induced ankle injury via alteration of PPAR-γ signal pathway. J. Food Biochem. 2021, 45, e13585. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Li, Y.; Chen, G.; Fu, S.; He, D.; Huang, B.; Wei, L.; Lin, Y.; Guo, Y.; Hu, G. Farrerol Ameliorates TNBS-Induced Colonic Inflammation by Inhibiting ERK1/2, JNK1/2, and NF-κB Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 2037. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wei, W.; Xiao, Q.; Yang, H.; Ci, X. Farrerol Ameliorates APAP-induced Hepatotoxicity via Activation of Nrf2 and Autophagy. Int. J. Biol. Sci. 2019, 15, 788–799. [Google Scholar] [CrossRef]
- Li, Y.; Gong, Q.; Guo, W.; Kan, X.; Xu, D.; Ma, H.; Fu, S.; Liu, J. Farrerol Relieve Lipopolysaccharide (LPS)-Induced Mastitis by Inhibiting AKT/NF-κB p65, ERK1/2 and P38 Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 1770. [Google Scholar] [CrossRef] [PubMed]
- Ci, X.; Chu, X.; Wei, M.; Yang, X.; Cai, Q.; Deng, X. Different effects of farrerol on an OVA-induced allergic asthma and LPS-induced acute lung injury. PloS ONE 2012, 7, e34634. [Google Scholar] [CrossRef]
- Ma, N.; Wei, W.; Fan, X.; Ci, X. Farrerol Attenuates Cisplatin-Induced Nephrotoxicity by Inhibiting the Reactive Oxygen Species-Mediated Oxidation, Inflammation, and Apoptotic Signaling Pathways. Front. Physiol. 2019, 10, 1419. [Google Scholar] [CrossRef]
- Ma, N.; Wei, Z.; Hu, J.; Gu, W.; Ci, X. Farrerol Ameliorated Cisplatin-Induced Chronic Kidney Disease Through Mitophagy Induction via Nrf2/PINK1 Pathway. Front. Pharmacol. 2021, 12, 768700. [Google Scholar] [CrossRef]
- Chevalier, R.L.; Forbes, M.S.; Thornhill, B.A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int. 2009, 75, 1145–1152. [Google Scholar] [CrossRef]
- Martínez-Klimova, E.; Aparicio-Trejo, O.E.; Tapia, E.; Pedraza-Chaverri, J. Unilateral Ureteral Obstruction as a Model to Investigate Fibrosis-Attenuating Treatments. Biomolecules 2019, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Han, L.; Zhang, X.; Yang, W.; Gao, Y.; Shen, Y.; Li, B.; Wang, S.; Qin, M.; Lowe, S.; et al. Tongluo Yishen Decoction Ameliorates Renal Fibrosis via Regulating Mitochondrial Dysfunction Induced by Oxidative Stress in Unilateral Ureteral Obstruction Rats. Front. Pharmacol. 2021, 12, 762756. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-M.; Lin, C.-Y.; Chung, Y.-P.; Liu, C.-H.; Huang, K.-T.; Guan, S.-S.; Wu, C.-T.; Liu, S.-H. Protective Effects of Nootkatone on Renal Inflammation, Apoptosis, and Fibrosis in a Unilateral Ureteral Obstructive Mouse Model. Nutrients 2021, 13, 3921. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y.; Jo, J.; Leem, J.; Park, K.-K. Kahweol Ameliorates Cisplatin-Induced Acute Kidney Injury through Pleiotropic Effects in Mice. Biomedicines 2020, 8, 572. [Google Scholar] [CrossRef] [PubMed]
- Gwon, M.-G.; Gu, H.; Leem, J.; Park, K.-K. Protective Effects of 6-Shogaol, an Active Compound of Ginger, in a Murine Model of Cisplatin-Induced Acute Kidney Injury. Molecules 2021, 26, 5931. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Leem, J.; Hong, H.-L. Melittin Ameliorates Endotoxin-Induced Acute Kidney Injury by Inhibiting Inflammation, Oxidative Stress, and Cell Death in Mice. Oxid. Med. Cell. Longev. 2021, 2021, 8843051. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Leem, J.; Park, K.-K. Antioxidative, Antiapoptotic, and Anti-Inflammatory Effects of Apamin in a Murine Model of Lipopolysaccharide-Induced Acute Kidney Injury. Molecules 2020, 25, 5717. [Google Scholar] [CrossRef]
- Baisantry, A.; Bhayana, S.; Rong, S.; Ermeling, E.; Wrede, C.; Hegermann, J.; Pennekamp, P.; Sörensen-Zender, I.; Haller, H.; Melk, A.; et al. Autophagy Induces Prosenescent Changes in Proximal Tubular S3 Segments. J. Am. Soc. Nephrol. 2016, 27, 1609–1616. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Lee, S.-J.; Maeng, Y.-I.; Leem, J.; Park, K.-K. Protective Effects of Bee Venom against Endotoxemia-Related Acute Kidney Injury in Mice. Biology 2020, 9, 154. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Choi, Y.; Leem, J.; Song, J.E. Heme Oxygenase-1 Induction by Cobalt Protoporphyrin Ameliorates Cholestatic Liver Disease in a Xenobiotic-Induced Murine Model. Int. J. Mol. Sci. 2021, 22, 8253. [Google Scholar] [CrossRef]
- Dendooven, A.; Ishola Jr, D.A.; Nguyen, T.Q.; Van der Giezen, D.M.; Kok, R.J.; Goldschmeding, R.; Joles, J.A. Oxidative stress in obstructive nephropathy. Int. J. Exp. Pathol. 2011, 92, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Aparicio-Trejo, O.E.; Ortega-Lozano, A.J.; Pedraza-Chaverri, J. Redox signaling pathways in unilateral ureteral obstruction (UUO)-induced renal fibrosis. Free Radic. Biol. Med. 2021, 172, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y.; Jo, J.; Kim, K.; An, H.-J.; Gwon, M.-G.; Gu, H.; Kim, H.-J.; Yang, A.Y.; Kim, S.-W.; Jeon, E.J.; et al. Pharmacological Activation of Sirt1 Ameliorates Cisplatin-Induced Acute Kidney Injury by Suppressing Apoptosis, Oxidative Stress, and Inflammation in Mice. Antioxidants 2019, 8, 322. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Leem, J. Hispidulin Ameliorates Endotoxin-Induced Acute Kidney Injury in Mice. Molecules 2022, 27, 2019. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Jo, J.; Leem, J.; Park, K.-K. Inhibition of p300 by Garcinol Protects against Cisplatin-Induced Acute Kidney Injury through Suppression of Oxidative Stress, Inflammation, and Tubular Cell Death in Mice. Antioxidants 2020, 9, 1271. [Google Scholar] [CrossRef]
- Lv, W.; Booz, G.W.; Fan, F.; Wang, Y.; Roman, R.J. Oxidative Stress and Renal Fibrosis: Recent Insights for the Development of Novel Therapeutic Strategies. Front. Physiol. 2018, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y.; Leem, J.; Hong, H.-L. Protective Effects of SPA0355, a Thiourea Analogue, Against Lipopolysaccharide-Induced Acute Kidney Injury in Mice. Antioxidants 2020, 9, 585. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Leem, J.; Jeon, E.J. Protective Effects of Melatonin Against Aristolochic Acid-Induced Nephropathy in Mice. Biomolecules 2020, 10, 11. [Google Scholar] [CrossRef]
- Jiménez-Uribe, A.P.; Gómez-Sierra, T.; Aparicio-Trejo, O.E.; Orozco-Ibarra, M.; Pedraza-Chaverri, J. Backstage players of fibrosis: NOX4, mTOR, HDAC, and S1P; companions of TGF-β. Cell. Signal. 2021, 87, 110123. [Google Scholar] [CrossRef]
- Truong, L.D.; Choi, Y.J.; Tsao, C.C.; Ayala, G.; Sheikh-Hamad, D.; Nassar, G.; Suki, W.N. Renal cell apoptosis in chronic obstructive uropathy: The roles of caspases. Kidney Int. 2001, 60, 924–934. [Google Scholar] [CrossRef]
- Wang, Y.; Harris, D.C.H. Macrophages in renal disease. J. Am. Soc. Nephrol. 2011, 22, 21–27. [Google Scholar] [CrossRef]
- Nishida, M.; Hamaoka, K. Macrophage phenotype and renal fibrosis in obstructive nephropathy. Nephron Exp. Nephrol. 2008, 110, e31–e36. [Google Scholar] [CrossRef] [PubMed]
- Frangie, C.; Daher, J. Role of myeloperoxidase in inflammation and atherosclerosis (Review). Biomed. Rep. 2022, 16, 53. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Hong, H.-L.; Kim, G.M.; Leem, J.; Kwon, H.H. Protective Effects of Carnosic Acid on Lipopolysaccharide-Induced Acute Kidney Injury in Mice. Molecules 2021, 26, 7589. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Gwon, M.-G.; Kim, J.H.; Leem, J.; Lee, S.-J. Oridonin Attenuates Cisplatin-Induced Acute Kidney Injury via Inhibiting Oxidative Stress, Apoptosis, and Inflammation in Mice. Biomed Res. Int. 2022, 2022, 3002962. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.K.; Pratap, R. The biological potential of flavones. Nat. Prod. Rep. 2010, 27, 1571–1593. [Google Scholar] [CrossRef]
- Ma, N.; Wang, X.; Qi, C.; Yu, Q.; Zhu, C.; Ren, H. Farrerol Enhances Nrf2-Mediated Defense Mechanisms against Hydrogen Peroxide-Induced Oxidative Damage in Human Retinal Pigment Epithelial Cells by Activating Akt and MAPK. Oxid. Med. Cell. Longev. 2021, 2021, 8847844. [Google Scholar] [CrossRef]
- Yan, C.; Zhang, X.; Miao, J.; Yuan, H.; Liu, E.; Liang, T.; Li, Q. Farrerol Directly Targets GSK-3 β to Activate Nrf2-ARE Pathway and Protect EA.hy926 Cells against Oxidative Stress-Induced Injuries. Oxid. Med. Cell. Longev. 2020, 2020, 5967434. [Google Scholar] [CrossRef]
- Cui, B.; Zhang, S.; Wang, Y.; Guo, Y. Farrerol attenuates β-amyloid-induced oxidative stress and inflammation through Nrf2/Keap1 pathway in a microglia cell line. Biomed. Pharmacother. 2019, 109, 112–119. [Google Scholar] [CrossRef]
- Ci, X.; Lv, H.; Wang, L.; Wang, X.; Peng, L.; Qin, F.X.; Cheng, G. The antioxidative potential of farrerol occurs via the activation of Nrf2 mediated HO-1 signaling in RAW 264.7 cells. Chem. Biol. Interact. 2015, 239, 192–199. [Google Scholar] [CrossRef]
- Chen, Z.; Gao, H.; Wang, L.; Ma, X.; Tian, L.; Zhao, W.; Li, K.; Zhang, Y.; Ma, F.; Lu, J.; et al. Farrerol alleviates high glucose-induced renal mesangial cell injury through the ROS/Nox4/ERK1/2 pathway. Chem. Biol. Interact. 2020, 316, 108921. [Google Scholar] [CrossRef]
- Li, J.-K.; Ge, R.; Tang, L.; Li, Q.-S. Protective effects of farrerol against hydrogen-peroxide-induced apoptosis in human endothelium-derived EA.hy926 cells. Can. J. Physiol. Pharmacol. 2013, 91, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Hue, M.; Rayego-Mateos, S.; Vázquez-Carballo, C.; Palomino-Antolín, A.; García-Caballero, C.; Opazo-Rios, L.; Morgado-Pascual, J.L.; Herencia, C.; Mas, S.; Ortiz, A.; et al. Protective Role of Nrf2 in Renal Disease. Antioxidants 2021, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Miyajima, A.; Chen, J.; Lawrence, C.; Ledbetter, S.; Soslow, R.A.; Stern, J.; Jha, S.; Pigato, J.; Lemer, M.L.; Poppas, D.P.; et al. Antibody to transforming growth factor-beta ameliorates tubular apoptosis in unilateral ureteral obstruction. Kidney Int. 2000, 58, 2301–2313. [Google Scholar] [CrossRef]
- Chen, Y.; Mu, L.; Xing, L.; Li, S.; Fu, S. Rhein alleviates renal interstitial fibrosis by inhibiting tubular cell apoptosis in rats. Biol. Res. 2019, 52, 50. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, Q.; Xia, R.; Cai, C. Farrerol exhibits inhibitory effects on lung adenocarcinoma cells by activating the mitochondrial apoptotic pathway. J. Biochem. Mol. Toxicol. 2022, 36, e23157. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.; Kim, J.S.; Choi, s.t.; Lee, S.G.; Ojulari, O.V.; Kang, Y.J.; Kwon, T.K.; Nam, J.-O. Farrerol Induces Cancer Cell Death via ERK Activation in SKOV3 Cells and Attenuates TNF-α-Mediated Lipolysis. Int. J. Mol. Sci. 2021, 22, 9400. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-M. Inflammatory Mediators and Renal Fibrosis. Adv. Exp. Med. Biol. 2019, 1165, 381–406. [Google Scholar]
- Cui, B.; Guo, X.; You, Y.; Fu, R. Farrerol attenuates MPP+ -induced inflammatory response by TLR4 signaling in a microglia cell line. Phytother. Res. 2019, 33, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, B.; Yu, J.-L. Farrerol inhibits IL-6 and IL-8 production in LPS-stimulated human gingival fibroblasts by suppressing PI3K/AKT/NF-κB signaling pathway. Arch. Oral Biol. 2016, 62, 28–32. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, J.; Zhuang, Y.; Han, G. Anti-inflammatory effects of farrerol on IL-1β-stimulated human osteoarthritis chondrocytes. Eur. J. Pharmacol. 2015, 764, 443–447. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) | Accession No. |
|---|---|---|
| COL1A1 | F: TCCTCCAGGGATCCAACGA R: GGCAGGCGGGAGGTCTT | NM_007742 |
| Fibronectin | F: CGAGGTGACAGAGACCACAA R: CTGGAGTCAAGCCAGACACA | NM_010233 |
| TGF-β1 | F: GCCCTGGATACCAACTATTGCTT R: AGTTGGCATGGTAGCCCTTG | NM_011577 |
| NOX4 | F: CCCAAGTTCCAAGCTCATTTCC R: TGGTGACAGGTTTGTTGCTCCT | NM_015760 |
| TNF-α | F: CACAGAAAGCATGATCCGCGACGT R: CGGCAGAGAGGAGGTTGACTTTCT | NM_013693 |
| IL-6 | F: TAGTCCTTCCTACCCCAATTTCC R: TTGGTCCTTAGCCACTCCTTC | NM_031168 |
| IL-1β | F: CGCAGCAGCACATCAACAAGAGC R: TGTCCTCATCCTGGAAGGTCCACG | NM_008361 |
| CXCL5 | F: TCATGAGAAGGCAATGCT R: ACATTATGCCATACTACGAAGA | NM_009141 |
| MCP-1 | F: TAAAAACCTGGATCGGAACCAA R: GCATTAGCTTCAGATTTACGGGT | NM_011333 |
| GAPDH | F: ACTCCACTCACGGCAAATTC R: TCTCCATGGTGGTGAAGACA | NM_001289726 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-Y.; Leem, J.; Park, K.-K. Antioxidant, Anti-Apoptotic, and Anti-Inflammatory Effects of Farrerol in a Mouse Model of Obstructive Uropathy. Curr. Issues Mol. Biol. 2023, 45, 337-352. https://doi.org/10.3390/cimb45010024
Kim J-Y, Leem J, Park K-K. Antioxidant, Anti-Apoptotic, and Anti-Inflammatory Effects of Farrerol in a Mouse Model of Obstructive Uropathy. Current Issues in Molecular Biology. 2023; 45(1):337-352. https://doi.org/10.3390/cimb45010024
Chicago/Turabian StyleKim, Jung-Yeon, Jaechan Leem, and Kwan-Kyu Park. 2023. "Antioxidant, Anti-Apoptotic, and Anti-Inflammatory Effects of Farrerol in a Mouse Model of Obstructive Uropathy" Current Issues in Molecular Biology 45, no. 1: 337-352. https://doi.org/10.3390/cimb45010024
APA StyleKim, J.-Y., Leem, J., & Park, K.-K. (2023). Antioxidant, Anti-Apoptotic, and Anti-Inflammatory Effects of Farrerol in a Mouse Model of Obstructive Uropathy. Current Issues in Molecular Biology, 45(1), 337-352. https://doi.org/10.3390/cimb45010024