
Enterobacter sp. AA26 as a Protein Source in the Larval Diet of Drosophila suzukii

 , ,
, ,  ,
,  , and
, and 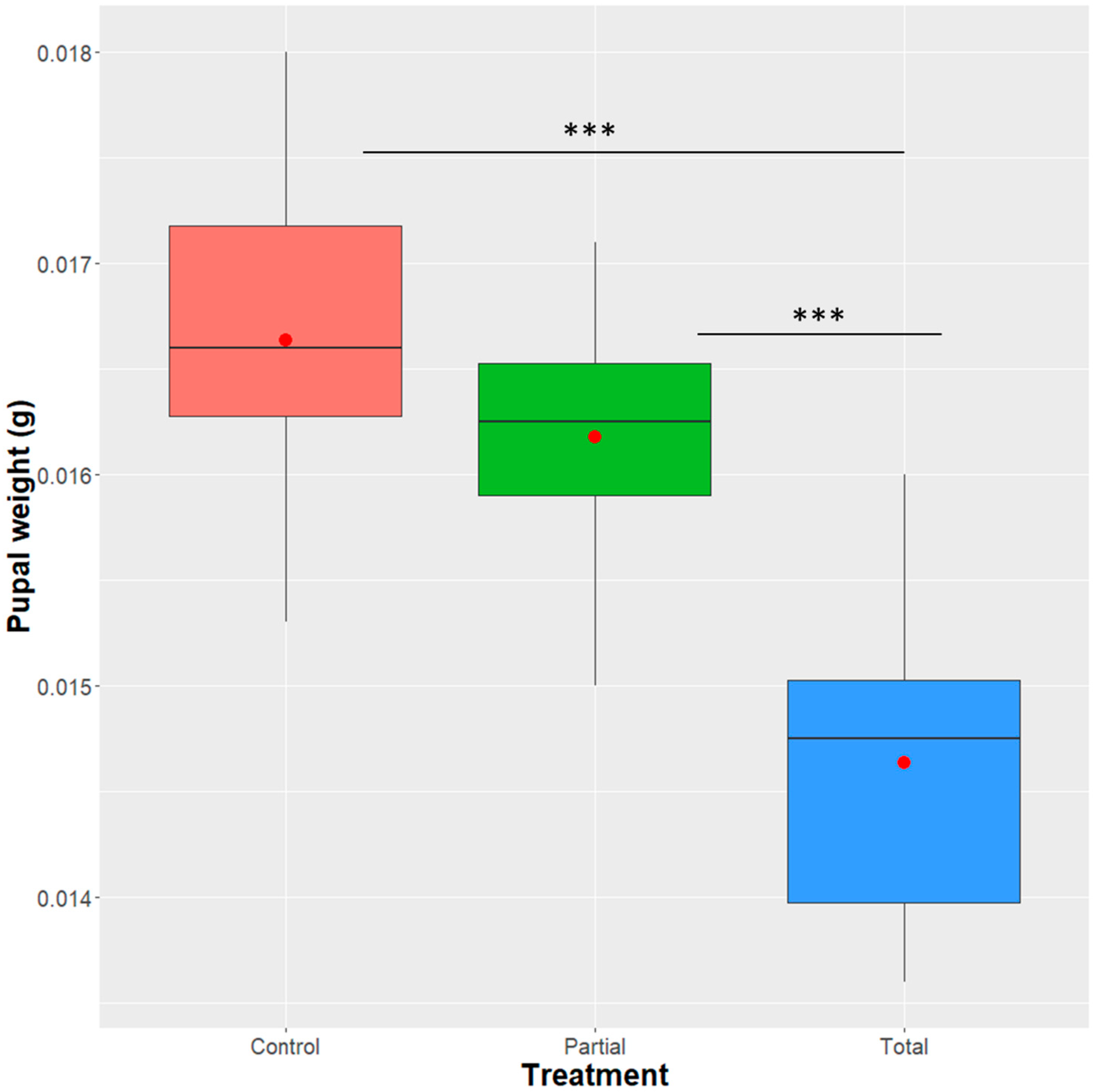{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila Suzukii Rearing Colony
2.2. Enterobacter sp. AA26 Biomass Production and Larval Diet Preparation
2.3. Drosophila Suzukii Egg Collection
2.4. Effect of Enterobacter sp. AA26 on Pupae and Adult Recovery
2.5. Effect of Enterobacter sp. AA26 on Pupal Weight
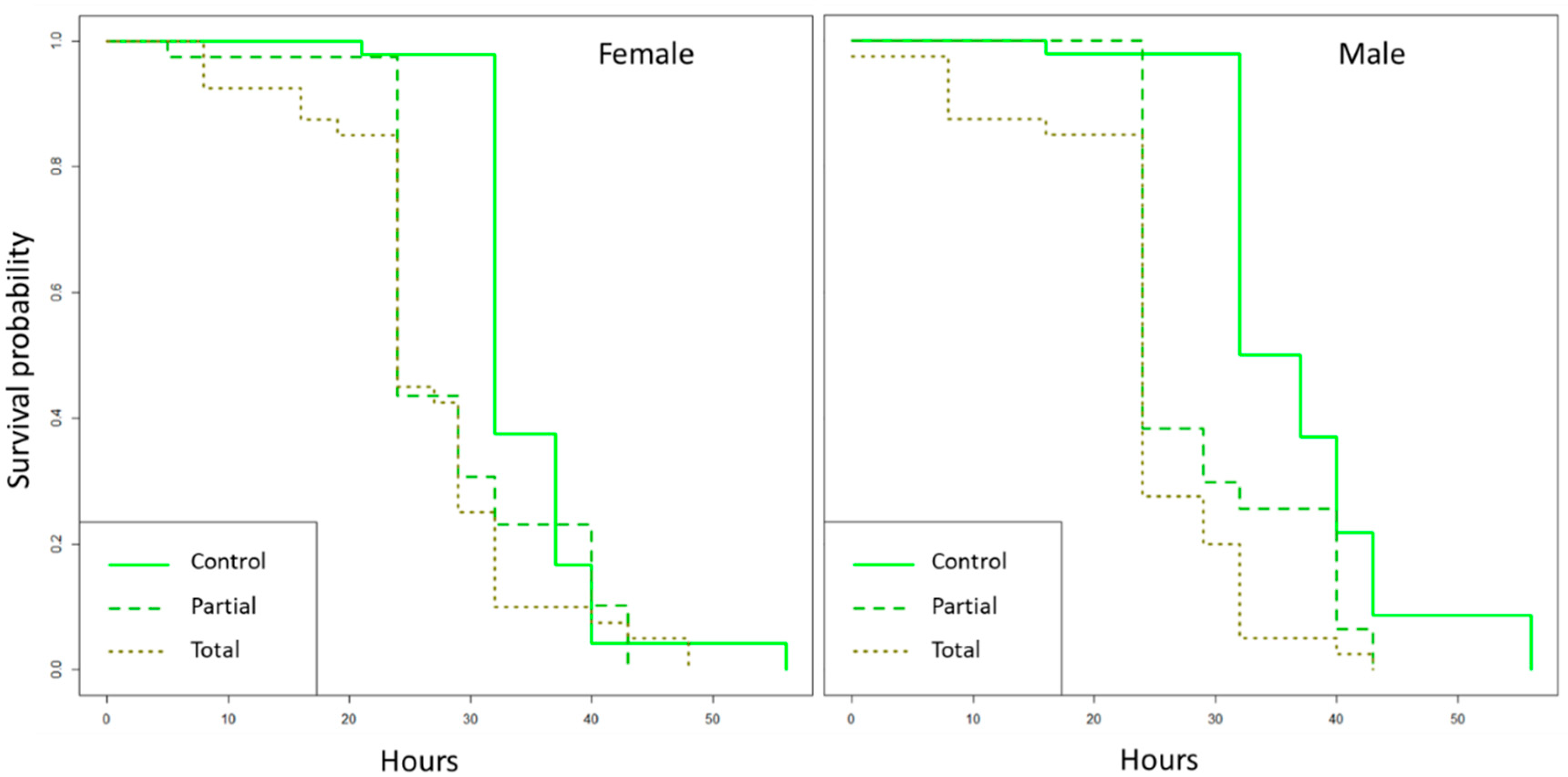
2.6. Effect of Enterobacter sp. AA26 on Survival
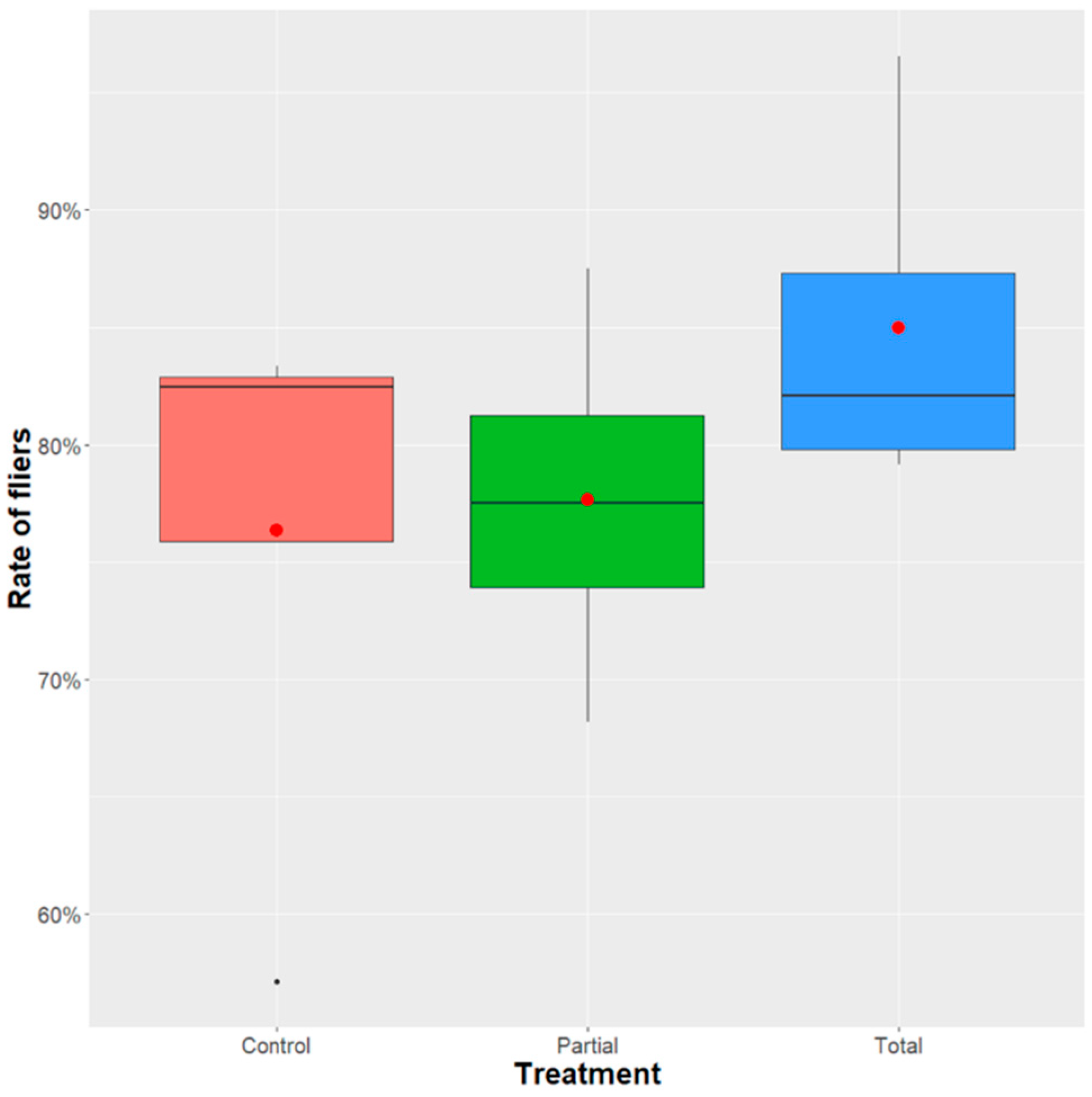
2.7. Effect of Enterobacter sp. AA26 on Flight Ability
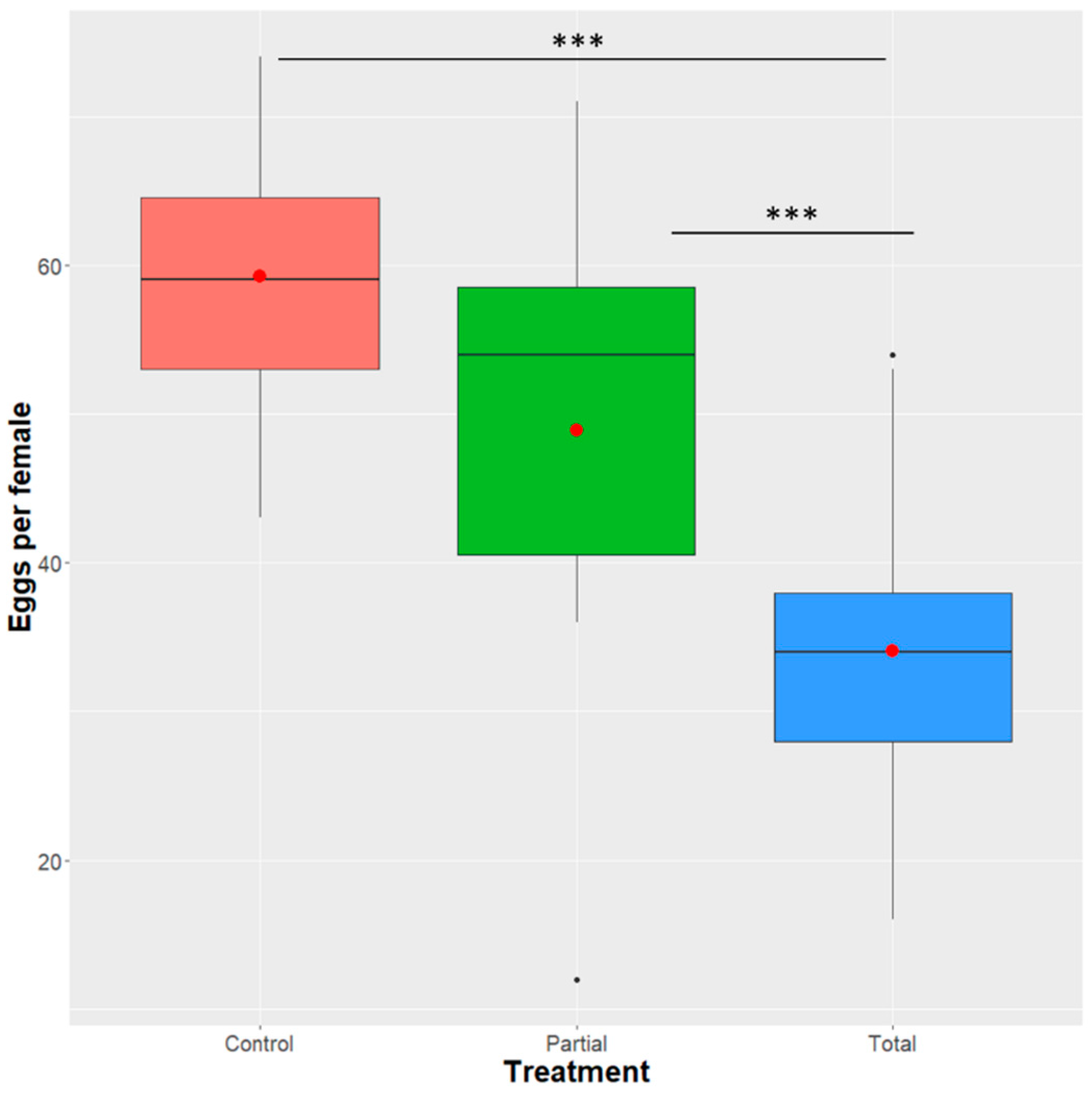
2.8. Effect of Enterobacter sp. AA26 on Female Fecundity
2.9. Statistical Analyses
3. Results
3.1. Effect of Enterobacter sp. AA26 on Pupal Weight
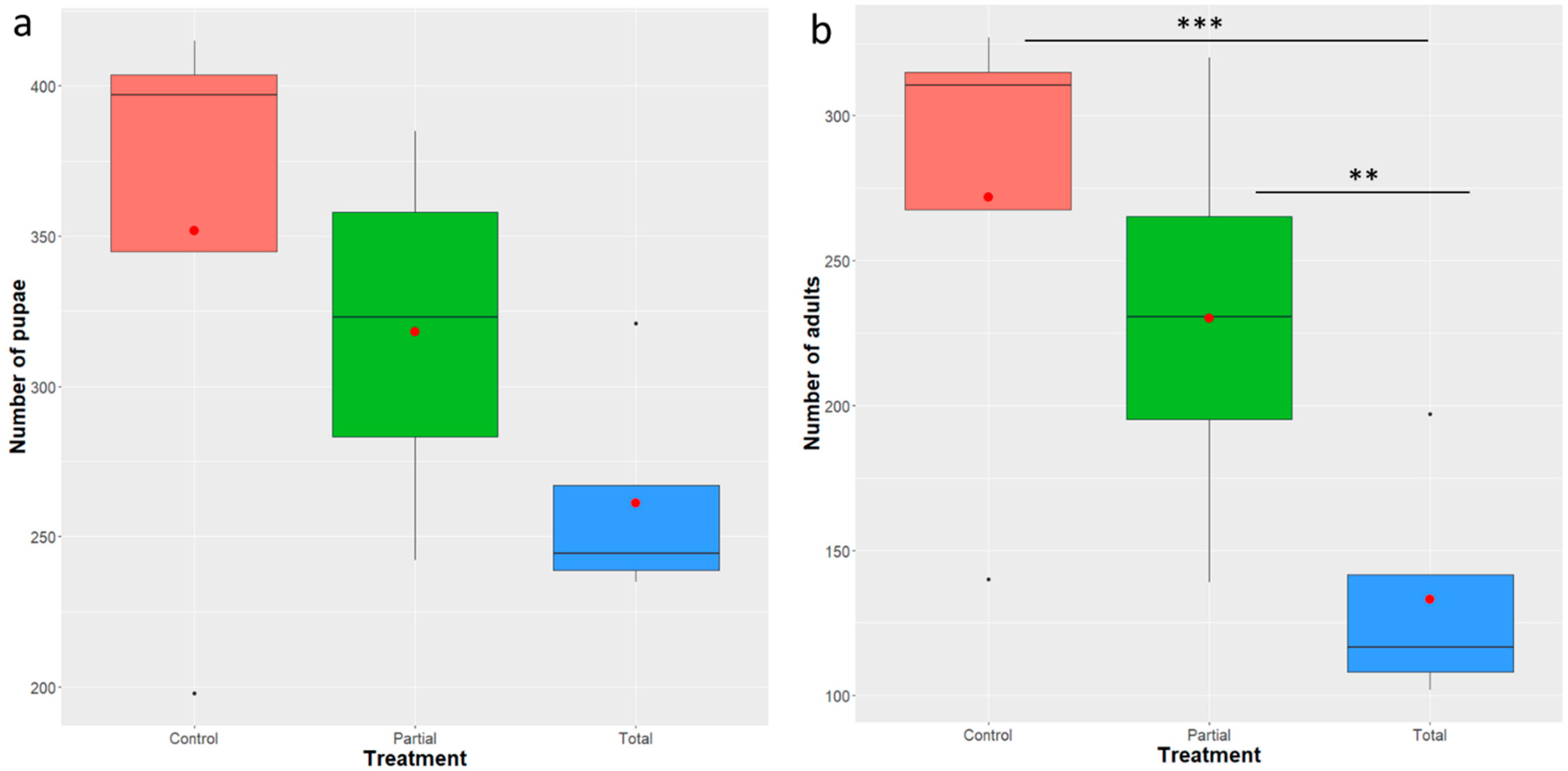
3.2. Effect of Enterobacter sp. AA26 on Developmental Parameters
3.2.1. Pupae Recovery
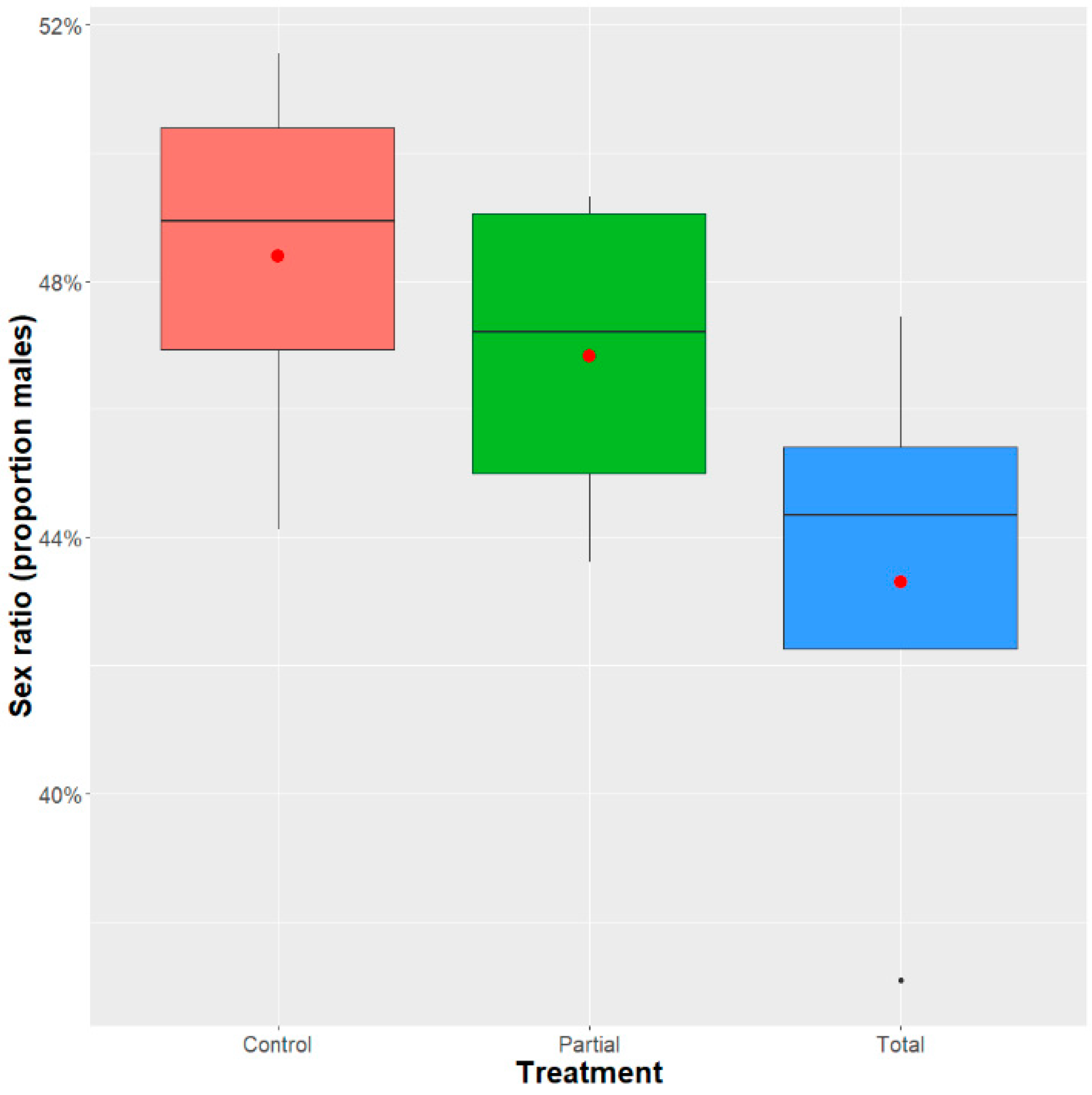
3.2.2. Adult Recovery and Sex Ratio
3.3. Effect of Enterobacter sp. AA26 on Adult Survival under Food and Water Starvation
3.4. Effect of Enterobacter sp. AA26 on Flight Ability
3.5. Effect of Enterobacter sp. AA26 on Fecundity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bourtzis, K.; Miller, T. Insect Symbiosis; CRC Press, Taylor and Francis Group, LLC: Boca Raton, FL, USA, 2003. [Google Scholar]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut Microbiota Affects Development and Olfactory Behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222, jeb.192500. [Google Scholar] [CrossRef] [PubMed]
- Lizé, A.; McKay, R.; Lewis, Z. Kin Recognition in Drosophila: The Importance of Ecology and Gut Microbiota. ISME J. 2014, 8, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Farine, J.-P.; Habbachi, W.; Cortot, J.; Roche, S.; Ferveur, J.-F. Maternally-Transmitted Microbiota Affects Odor Emission and Preference in Drosophila Larva. Sci. Rep. 2017, 7, 6062. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Multiorganismal Insects: Diversity and Function of Resident Microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef]
- Kouser, S.; Qaim, M. Impact of Bt Cotton on Pesticide Poisoning in Smallholder Agriculture: A Panel Data Analysis. Ecol. Econ. 2011, 70, 2105–2113. [Google Scholar] [CrossRef]
- Crotti, E.; Balloi, A.; Hamdi, C.; Sansonno, L.; Marzorati, M.; Gonella, E.; Favia, G.; Cherif, A.; Bandi, C.; Alma, A.; et al. Microbial Symbionts: A Resource for the Management of Insect-Related Problems: Microbial Resource Management Applied to Insects. Microb. Biotechnol. 2012, 5, 307–317. [Google Scholar] [CrossRef]
- Deutscher, A.T.; Reynolds, O.L.; Chapman, T.A. Yeast: An Overlooked Component of Bactrocera tryoni (Diptera: Tephritidae) Larval Gut Microbiota. J. Econ. Entomol. 2016, 110, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Yuval, B.; Ben-Ami, E.; Behar, A.; Ben-Yosef, M.; Jurkevitch, E. The Mediterranean Fruit Fly and Its Bacteria—Potential for Improving Sterile Insect Technique Operations: Bacteria of Ceratitis capitata and the SIT. J. Appl. Entomol. 2013, 137, 39–42. [Google Scholar] [CrossRef]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-303557-2. [Google Scholar]
- Enkerlin, W.R. Impact of Fruit Fly Control Programmes Using the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-303557-2. [Google Scholar]
- Knipling, E.F. Possibilities of Insect Control or Eradication Through the Use of Sexually Sterile Males1. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Parker, A.G. Mass-Rearing for the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-303557-2. [Google Scholar]
- Chaudhury, M.F.B. Handbook of Insect Rearing, Vol. I and II; Singh, P., Moore, R.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Cohen, A.C. Ecology of Insect Rearing Systems: A Mini-Review of Insect Rearing Papers from 1906-2017. Adv. Entomol. 2018, 6, 86–115. [Google Scholar] [CrossRef]
- Shimoji, Y.; Miyatake, T. Adaptation to Artificial Rearing During Successive Generations in the West Indian Sweetpotato Weevil, Euscepes postfasciatus (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 2002, 95, 735–739. [Google Scholar] [CrossRef]
- Niyazi, N.; Lauzon, C.R.; Shelly, T.E. Effect of Probiotic Adult Diets on Fitness Components of Sterile Male Mediterranean Fruit Flies (Diptera: Tephritidae) Under Laboratory and Field Cage Conditions. J. Econ. Entomol. 2004, 97, 1570–1580. [Google Scholar] [CrossRef]
- Ami, E.B.; Yuval, B.; Jurkevitch, E. Manipulation of the Microbiota of Mass-Reared Mediterranean Fruit Flies Ceratitis capitata (Diptera: Tephritidae) Improves Sterile Male Sexual Performance. ISME J. 2010, 4, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Rull, J.; Lasa, R.; Rodriguez, C.; Ortega, R.; Velazquez, O.E.; Aluja, M. Artificial Selection, Pre-Release Diet, and Gut Symbiont Inoculation Effects on Sterile Male Longevity for Area-Wide Fruit-Fly Management. Entomol. Exp. Appl. 2015, 157, 325–333. [Google Scholar] [CrossRef]
- Meats, A.; Streamer, K.; Gilchrist, A.S. Bacteria as Food Had No Effect on Fecundity during Domestication of the Fruit Fly, Bactrocera tryoni. J. Appl. Entomol. 2009, 133, 633–639. [Google Scholar] [CrossRef]
- Gavriel, S.; Jurkevitch, E.; Gazit, Y.; Yuval, B. Bacterially Enriched Diet Improves Sexual Performance of Sterile Male Mediterranean Fruit Flies: Bacteria and Medfly Sexual Performance. J. Appl. Entomol. 2011, 135, 564–573. [Google Scholar] [CrossRef]
- Hamden, H.; M’saad Guerfali, M.; Fadhl, S.; Saidi, M.; Chevrier, C. Fitness Improvement of Mass-Reared Sterile Males of Ceratitis capitata (Vienna 8 Strain) (Diptera: Tephritidae) After Gut Enrichment With Probiotics. J. Econ. Entomol. 2013, 106, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Augustinos, A.A.; Kyritsis, G.A.; Papadopoulos, N.T.; Abd-Alla, A.M.M.; Cáceres, C.; Bourtzis, K. Exploitation of the Medfly Gut Microbiota for the Enhancement of Sterile Insect Technique: Use of Enterobacter sp. in Larval Diet-Based Probiotic Applications. PLoS ONE 2015, 10, e0136459. [Google Scholar] [CrossRef]
- Kyritsis, G.A.; Augustinos, A.A.; Cáceres, C.; Bourtzis, K. Medfly Gut Microbiota and Enhancement of the Sterile Insect Technique: Similarities and Differences of Klebsiella oxytoca and Enterobacter sp. AA26 Probiotics during the Larval and Adult Stages of the VIENNA 8D53+ Genetic Sexing Strain. Front. Microbiol. 2017, 8, 2064. [Google Scholar] [CrossRef]
- Cai, Z.; Yao, Z.; Li, Y.; Xi, Z.; Bourtzis, K.; Zhao, Z.; Bai, S.; Zhang, H. Intestinal Probiotics Restore the Ecological Fitness Decline of Bactrocera dorsalis by Irradiation. Evol. Appl. 2018, 11, 1946–1963. [Google Scholar] [CrossRef]
- Khaeso, K.; Andongma, A.A.; Akami, M.; Souliyanonh, B.; Zhu, J.; Krutmuang, P.; Niu, C.-Y. Assessing the Effects of Gut Bacteria Manipulation on the Development of the Oriental Fruit Fly, Bactrocera dorsalis (Diptera; Tephritidae). Symbiosis 2018, 74, 97–105. [Google Scholar] [CrossRef]
- Sacchetti, P.; Ghiardi, B.; Granchietti, A.; Stefanini, F.M.; Belcari, A. Development of Probiotic Diets for the Olive Fly: Evaluation of Their Effects on Fly Longevity and Fecundity. Ann. Appl. Biol. 2014, 164, 138–150. [Google Scholar] [CrossRef]
- Kyritsis, G.A.; Augustinos, A.A.; Ntougias, S.; Papadopoulos, N.T.; Bourtzis, K.; Cáceres, C. Enterobacter sp. AA26 Gut Symbiont as a Protein Source for Mediterranean Fruit Fly Mass-Rearing and Sterile Insect Technique Applications. BMC Microbiol. 2019, 19, 288. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Pasternak, Z.; Jurkevitch, E.; Yuval, B. Symbiotic Bacteria Enable Olive Fly Larvae to Overcome Host Defences. R. Soc. Open Sci. 2015, 2, 150170. [Google Scholar] [CrossRef] [PubMed]
- Capuzzo, C.; Firrao, G.; Mazzon, L.; Squartini, A.; Girolami, V. Candidatus Erwinia dacicola, a Coevolved Symbiotic Bacterium of the Olive Fly Bactrocera Oleae (Gmelin). Int. J. Syst. Evol. Microbiol. 2005, 55, 1641–1647. [Google Scholar] [CrossRef]
- Hauser, M. A Historic Account of the Invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the Continental United States, with Remarks on Their Identification. Pest. Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, L.A.; Mendes, M.F.; Krüger, A.P.; Blauth, M.L.; Gottschalk, M.S.; Garcia, F.R.M. Global Potential Distribution of Drosophila suzukii (Diptera: Drosophilidae). PLoS ONE. 2017, 12, e0174318. [Google Scholar] [CrossRef] [PubMed]
- Calabria, G.; Máca, J.; Bächli, G.; Serra, L.; Pascual, M. First Records of the Potential Pest Species Drosophila suzukii (Diptera: Drosophilidae) in Europe: First Record of Drosophila suzukii in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Deprá, M.; Poppe, J.L.; Schmitz, H.J.; De Toni, D.C.; Valente, V.L.S. The First Records of the Invasive Pest Drosophila suzukii in the South American Continent. J. Pest. Sci. 2014, 87, 379–383. [Google Scholar] [CrossRef]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying Host Potentials: Indexing Postharvest Fresh Fruits for Spotted Wing Drosophila, Drosophila suzukii. PLoS ONE 2013, 8, e61227. [Google Scholar] [CrossRef]
- Mitsui, H.; Takahashi, K.H.; Kimura, M.T. Spatial Distributions and Clutch Sizes of Drosophila Species Ovipositing on Cherry Fruits of Different Stages. Popul. Ecol. 2006, 48, 233–237. [Google Scholar] [CrossRef]
- De Ros, G.; Grassi, A.; Pantezzi, T. Recent Trends in the Economic Impact of Drosophila suzukii. In Drosophila suzukii Management; Garcia, F.R.M., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 11–27. ISBN 978-3-030-62691-4. [Google Scholar]
- Lee, J.C.; Wang, X.; Daane, K.M.; Hoelmer, K.A.; Isaacs, R.; Sial, A.A.; Walton, V.M. Biological Control of Spotted-Wing Drosophila (Diptera: Drosophilidae)—Current and Pending Tactics. J. Integr. Pest. Manag. 2019, 10, 13. [Google Scholar] [CrossRef]
- Leach, H.; Van Timmeren, S.; Isaacs, R. Exclusion Netting Delays and Reduces Drosophila suzukii (Diptera: Drosophilidae) Infestation in Raspberries. J. Econ. Entomol. 2016, 109, 2151–2158. [Google Scholar] [CrossRef] [PubMed]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Cáceres, C.; Stauffer, C.; Pereira, R.; et al. Sterile Insect Technique and Wolbachia Symbiosis as Potential Tools for the Control of the Invasive Species Drosophila suzukii. J. Pest. Sci. 2018, 91, 489–503. [Google Scholar] [CrossRef]
- Nikolouli, K.; Sassù, F.; Mouton, L.; Stauffer, C.; Bourtzis, K. Combining Sterile and Incompatible Insect Techniques for the Population Suppression of Drosophila suzukii. J. Pest. Sci. 2020, 93, 647–661. [Google Scholar] [CrossRef]
- Sassù, F.; Nikolouli, K.; Stauffer, C.; Bourtzis, K.; Caceres, C. Sterile insect technique and incompatible insect technique for the integrated Drosophila suzukii management. In Drosophila suzukii Management; Garcia, F.R.M., Ed.; Springer: Cham, Switzerland. [CrossRef]
- Sassù, F.; Nikolouli, K.; Pereira, R.; Vreysen, M.J.B.; Stauffer, C.; Cáceres, C. Irradiation Dose Response under Hypoxia for the Application of the Sterile Insect Technique in Drosophila suzukii. PLoS ONE 2019, 14, e0226582. [Google Scholar] [CrossRef]
- Sassù, F.; Nikolouli, K.; Caravantes, S.; Taret, G.; Pereira, R.; Vreysen, M.J.B.; Stauffer, C.; Cáceres, C. Mass-Rearing of Drosophila suzukii for Sterile Insect Technique Application: Evaluation of Two Oviposition Systems. Insects 2019, 10, 448. [Google Scholar] [CrossRef]
- Hamby, K.A.; Becher, P.G. Current Knowledge of Interactions between Drosophila suzukii and Microbes, and Their Potential Utility for Pest Management. J. Pest. Sci. 2016, 89, 621–630. [Google Scholar] [CrossRef]
- Fountain, M.T.; Bennett, J.; Cobo-Medina, M.; Conde Ruiz, R.; Deakin, G.; Delgado, A.; Harrison, R.; Harrison, N. Alimentary Microbes of Winter-Form Drosophila suzukii. Insect. Mol. Biol. 2018, 27, 383–392. [Google Scholar] [CrossRef]
- Vacchini, V.; Gonella, E.; Crotti, E.; Prosdocimi, E.M.; Mazzetto, F.; Chouaia, B.; Callegari, M.; Mapelli, F.; Mandrioli, M.; Alma, A.; et al. Bacterial Diversity Shift Determined by Different Diets in the Gut of the Spotted Wing Fly Drosophila suzukii Is Primarily Reflected on Acetic Acid Bacteria: Acetic Acid Bacteria of Drosophila suzukii. Environ. Microbiol. Rep. 2017, 9, 91–103. [Google Scholar] [CrossRef]
- Jiménez-Padilla, Y.; Esan, E.O.; Floate, K.D.; Sinclair, B.J. Persistence of Diet Effects on the Microbiota of Drosophila suzukii (Diptera: Drosophilidae). Can. Entomol. 2020, 152, 516–531. [Google Scholar] [CrossRef]
- Bing, X.; Gerlach, J.; Loeb, G.; Buchon, N. Nutrient-Dependent Impact of Microbes on Drosophila suzukii Development. mBio 2018, 9, e02199-17. [Google Scholar] [CrossRef]
- Chandler, J.A.; James, P.M.; Jospin, G.; Lang, J.M. The Bacterial Communities of Drosophila suzukii Collected from Undamaged Cherries. PeerJ 2014, 2, e474. [Google Scholar] [CrossRef]
- Chandler, J.A.; Morgan Lang, J.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial Communities of Diverse Drosophila Species: Ecological Context of a Host–Microbe Model System. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef]
- Solomon, G.M.; Dodangoda, H.; McCarthy-Walker, T.; Ntim-Gyakari, R.; Newell, P.D. The Microbiota of Drosophila suzukii Influences the Larval Development of Drosophila melanogaster. PeerJ 2019, 7, e8097. [Google Scholar] [CrossRef]
- Martinez-Sañudo, I.; Simonato, M.; Squartini, A.; Mori, N.; Marri, L.; Mazzon, L. Metagenomic Analysis Reveals Changes of the Drosophila suzukii Microbiota in the Newly Colonized Regions: Microbiota Associated to Drosophila suzukii. Insect Sci. 2018, 25, 833–846. [Google Scholar] [CrossRef]
- Brummel, T.; Ching, A.; Seroude, L.; Simon, A.F.; Benzer, S. Drosophila Lifespan Enhancement by Exogenous Bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 12974–12979. [Google Scholar] [CrossRef]
- Cox, C.R.; Gilmore, M.S. Native Microbial Colonization of Drosophila melanogaster and Its Use as a Model of Enterococcus faecalis Pathogenesis. Infect. Immun. 2007, 75, 1565–1576. [Google Scholar] [CrossRef]
- Hiebert, N.; Carrau, T.; Bartling, M.; Vilcinskas, A.; Lee, K.-Z. Identification of Entomopathogenic Bacteria Associated with the Invasive Pest Drosophila suzukii in Infested Areas of Germany. J. Invertebr. Pathol. 2020, 173, 107389. [Google Scholar] [CrossRef]
- Shu, R.; Hahn, D.A.; Jurkevitch, E.; Liburd, O.E.; Yuval, B.; Wong, A.C.-N. Sex-Dependent Effects of the Microbiome on Foraging and Locomotion in Drosophila suzukii. Front. Microbiol. 2021, 12, 656406. [Google Scholar] [CrossRef]
- Keebaugh, E.S.; Yamada, R.; Obadia, B.; Ludington, W.B.; Ja, W.W. Microbial Quantity Impacts Drosophila Nutrition, Development, and Lifespan. iScience 2018, 4, 247–259. [Google Scholar] [CrossRef]
- Azis, K.; Zerva, I.; Melidis, P.; Caceres, C.; Bourtzis, K.; Ntougias, S. Biochemical and Nutritional Characterization of the Medfly Gut Symbiont Enterobacter sp. AA26 for Its Use as Probiotics in Sterile Insect Technique Applications. BMC Biotechnol. 2019, 19, 90. [Google Scholar] [CrossRef]
- FAO/IAEA/USDA. Product Quality Control for Sterile Mass-Reared and Released Tephritid Fruit Flies. 2019. Available online: https://www.iaea.org/sites/default/files/qcv7.pdf (accessed on 17 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.R-project.org/ (accessed on 17 September 2021).
- Demétrio, C.G.B.; Hinde, J.; Moral, R.A. Models for Overdispersed Data in Entomology. In Ecological Modelling Applied to Entomology; Ferreira, C.P., Godoy, W.A.C., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 219–259. ISBN 978-3-319-06876-3. [Google Scholar]
- Ver Hoef, J.M.; Boveng, P.L. Quasi-Poisson vs. Negative Binomial Regression: How Should We Model Overdispersed Count Data? Ecology 2007, 88, 2766–2772. [Google Scholar] [CrossRef]
- Nelder, J.A.; Wedderburn, R.W.M. Generalized Linear Models. J. R. Stat. Soc. Ser. A. Stat. Soc. 1972, 135, 370. [Google Scholar] [CrossRef]
- Dunn, P.K.; Smyth, G.K. Generalized Linear Models With Examples in R., 1st ed.; Springer: New York, NY, USA, 2018; ISBN 978-1-4419-0118-7. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000; ISBN 978-0-387-98784-2. [Google Scholar]
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-Normal Plots and Overdispersed Models in R: The Hnp Package. J. Stat. Soft. 2017, 81(10), 1–23. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2021. R Package Version. Available online: http://florianhartig.github.io/DHARMa/ (accessed on 17 September 2021).
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Soft. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Lesperance, D.N.; Broderick, N.A. Microbiomes as Modulators of Drosophila melanogaster Homeostasis and Disease. Curr. Opin. Insect Sci. 2020, 39, 84–90. [Google Scholar] [CrossRef]
- Corby-Harris, V.; Pontaroli, A.C.; Shimkets, L.J.; Bennetzen, J.L.; Habel, K.E.; Promislow, D.E.L. Geographical Distribution and Diversity of Bacteria Associated with Natural Populations of Drosophila melanogaster. Appl. Environ. Microbiol. 2007, 73, 3470–3479. [Google Scholar] [CrossRef]
- Broderick, N.A.; Lemaitre, B. Gut-Associated Microbes of Drosophila melanogaster. Gut Microbes 2012, 3, 307–321. [Google Scholar] [CrossRef]
- Augustinos, A.A. Insect Symbiosis in Support of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-303557-2. [Google Scholar]
- Geiger, A.; Fardeau, M.-L.; Grebaut, P.; Vatunga, G.; Josénando, T.; Herder, S.; Cuny, G.; Truc, P.; Ollivier, B. First Isolation of Enterobacter, Enterococcus, and Acinetobacter spp. as Inhabitants of the Tsetse Fly (Glossina palpalis palpalis) Midgut. Infect. Genet. Evol. 2009, 9, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Alvarez, C.; Kukutla, P.; Yu, W.; Xu, J. Draft Genome Sequences of Enterobacter sp. Isolate Ag1 from the Midgut of the Malaria Mosquito Anopheles Gambiae. J. Bacteriol. 2012, 194, 5481. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gujjar, N.R.; Govindan, S.; Verghese, A.; Subramaniam, S.; More, R. Diversity of the Cultivable Gut Bacterial Communities Associated with the Fruit Flies Bactrocera dorsalis and Bactrocera cucurbitae (Diptera: Tephritidae). Phytoparasitica 2017, 45, 453–460. [Google Scholar] [CrossRef]
- Hardin, J.A.; Kraus, D.A.; Burrack, H.J. Diet Quality Mitigates Intraspecific Larval Competition in Drosophila suzukii. Entomol. Exp. Appl. 2015, 156, 59–65. [Google Scholar] [CrossRef]
- Rendon, D.; Walton, V.; Tait, G.; Buser, J.; Lemos Souza, I.; Wallingford, A.; Loeb, G.; Lee, J. Interactions among Morphotype, Nutrition, and Temperature Impact Fitness of an Invasive Fly. Ecol. Evol. 2019, 9, 2615–2628. [Google Scholar] [CrossRef]
- Lee, J.C.; Burrack, H.J.; Barrantes, L.D.; Beers, E.H.; Dreves, A.J.; Hamby, K.A.; Haviland, D.R.; Isaacs, R.; Richardson, T.A.; Shearer, P.W.; et al. Evaluation of Monitoring Traps for Drosophila suzukii (Diptera: Drosophilidae) in North America. J. Econ. Entomol. 2012, 105, 1350–1357. [Google Scholar] [CrossRef]
- Lewis, M.T.; Hamby, K.A. Differential Impacts of Yeasts on Feeding Behavior and Development in Larval Drosophila suzukii (Diptera:Drosophilidae). Sci. Rep. 2019, 9, 13370. [Google Scholar] [CrossRef]
- Bellutti, N.; Gallmetzer, A.; Innerebner, G.; Schmidt, S.; Zelger, R.; Koschier, E.H. Dietary Yeast Affects Preference and Performance in Drosophila suzukii. J. Pest. Sci. 2018, 91, 651–660. [Google Scholar] [CrossRef]
- Silva-Soares, N.F.; Nogueira-Alves, A.; Beldade, P.; Mirth, C.K. Adaptation to New Nutritional Environments: Larval Performance, Foraging Decisions, and Adult Oviposition Choices in Drosophila suzukii. BMC Ecol. 2017, 17, 21. [Google Scholar] [CrossRef]
- Young, Y.; Buckiewicz, N.; Long, T.A.F. Nutritional Geometry and Fitness Consequences in Drosophila suzukii, the Spotted-Wing Drosophila. Ecol. Evol. 2018, 8, 2842–2851. [Google Scholar] [CrossRef]
- Priyadarsini, S.; Sahoo, M.; Sahu, S.; Jayabalan, R.; Mishra, M. An Infection of Enterobacter ludwigii Affects Development and Causes Age-Dependent Neurodegeneration in Drosophila melanogaster. Invert. Neurosci. 2019, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Zografou, E.N.; Tsiropoulos, G.J.; Margaritis, L.H. Survival, Fecundity and Fertility of Bactrocera oleae, as Affected by Amino Acid Analogues. Entomol. Exp. Appl. 1998, 87, 125–132. [Google Scholar] [CrossRef]
- Chang, C.L.; Albrecht, C.; El-Shall, S.S.A.; Kurashima, R. Adult Reproductive Capacity of Ceratitis capitata (Diptera: Tephritidae) on a Chemically Defined Diet. Ann. Entomol. Soc. Am. 2001, 94, 702–706. [Google Scholar] [CrossRef]
- Jacob, F.F.; Michel, M.; Zarnkow, M.; Hutzler, M.; Methner, F.-J. The Complexity of Yeast Extracts and Its Consequences on the Utility in Brewing: A Review. Brew. Sci. 2019, 50–62. [Google Scholar] [CrossRef]
- Yamada, E.A.; Sgarbieri, V.C. Yeast (Saccharomyces cerevisiae) Protein Concentrate: Preparation, Chemical Composition, and Nutritional and Functional Properties. J. Agric. Food Chem. 2005, 53, 3931–3936. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolouli, K.; Sassù, F.; Ntougias, S.; Stauffer, C.; Cáceres, C.; Bourtzis, K. Enterobacter sp. AA26 as a Protein Source in the Larval Diet of Drosophila suzukii. Insects 2021, 12, 923. https://doi.org/10.3390/insects12100923
Nikolouli K, Sassù F, Ntougias S, Stauffer C, Cáceres C, Bourtzis K. Enterobacter sp. AA26 as a Protein Source in the Larval Diet of Drosophila suzukii. Insects. 2021; 12(10):923. https://doi.org/10.3390/insects12100923
Chicago/Turabian StyleNikolouli, Katerina, Fabiana Sassù, Spyridon Ntougias, Christian Stauffer, Carlos Cáceres, and Kostas Bourtzis. 2021. "Enterobacter sp. AA26 as a Protein Source in the Larval Diet of Drosophila suzukii" Insects 12, no. 10: 923. https://doi.org/10.3390/insects12100923
APA StyleNikolouli, K., Sassù, F., Ntougias, S., Stauffer, C., Cáceres, C., & Bourtzis, K. (2021). Enterobacter sp. AA26 as a Protein Source in the Larval Diet of Drosophila suzukii. Insects, 12(10), 923. https://doi.org/10.3390/insects12100923