Defining Parallels between the Salivary Glands and Pancreas to Better Understand Pancreatic Carcinogenesis

{kind=link}
{kind=link}
Abstract
1. Introduction
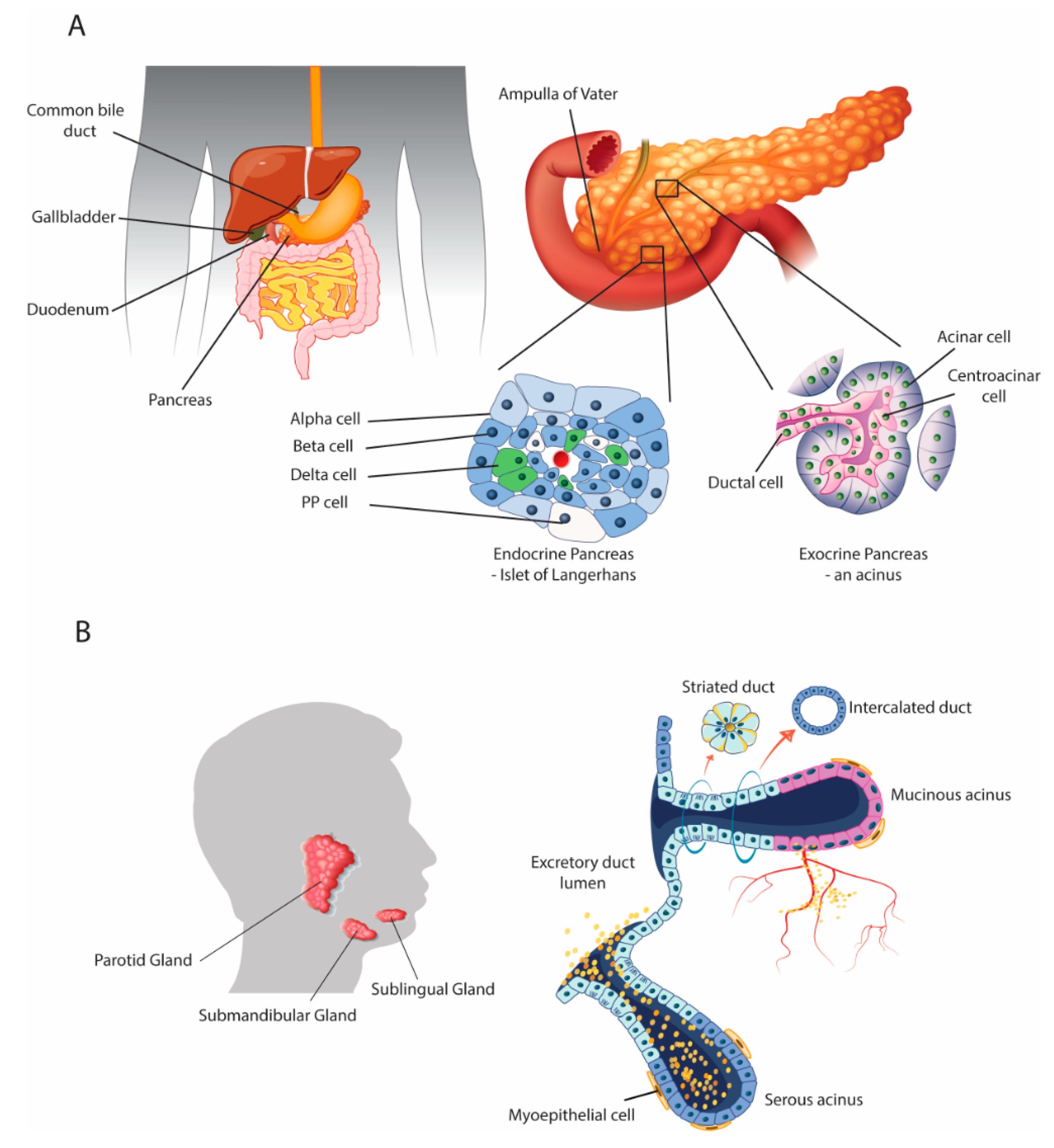
2. Parallels with the Salivary Glands
2.1. Anatomical and Physiological Parallels
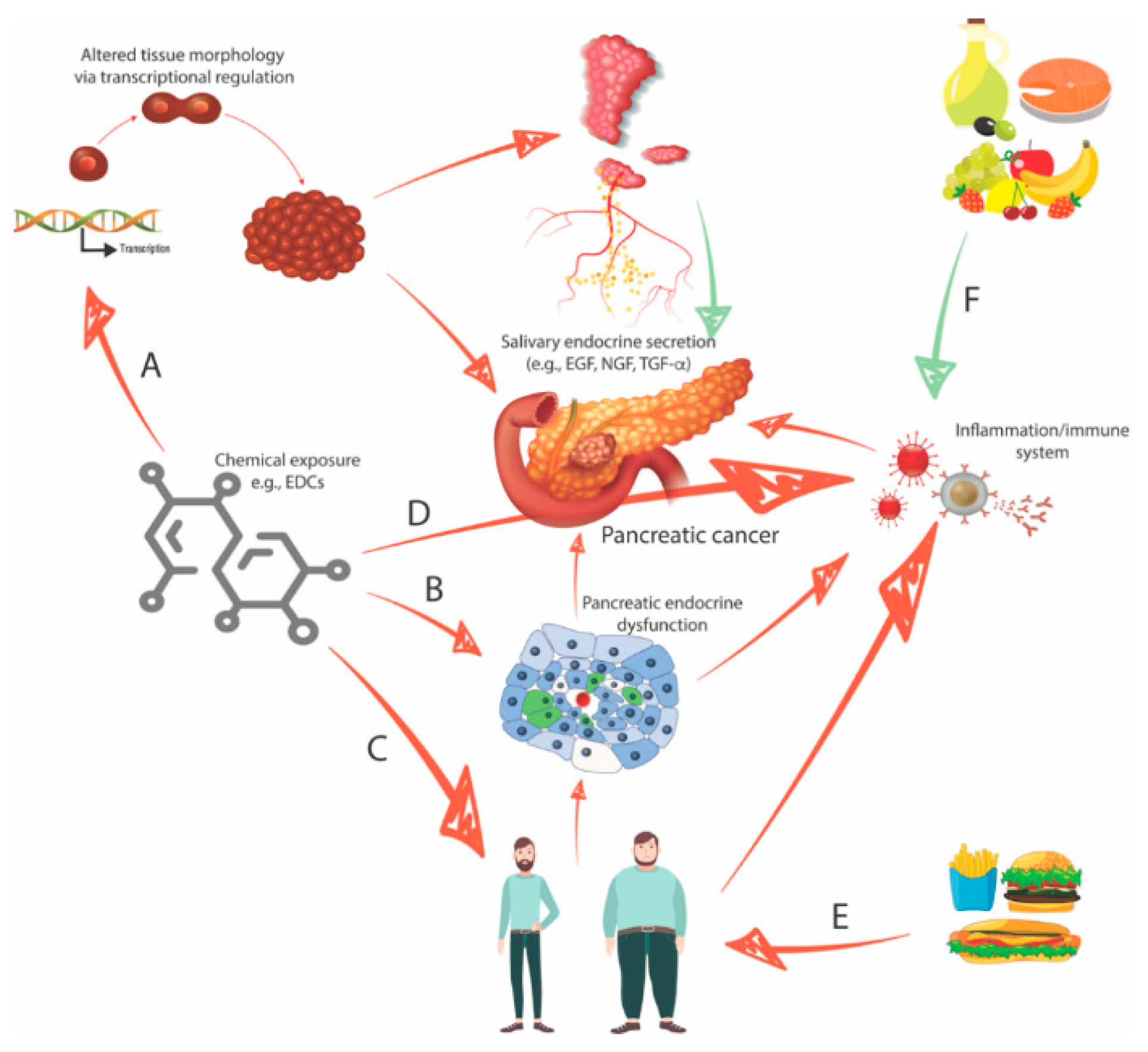
2.2. Interaction between the Organs: The Role of the Growth Factor EGF
2.3. Endocrine-Disrupting Compounds Perturb Organ Morphology and Function
2.4. Endocrine-Disrupting Compounds Contribute to the Development of Type II Diabetes and Obesity
2.5. Four Interconnected Risk Factors for Pancreatic Cancer: Chemical Exposure, Diabetes, Obesity, and Eating Behavior
3. Urgent Need for Early Diagnosis: Identification of Salivary Biomarkers and Development of Screening Tools
4. Conclusions
Funding
Conflicts of Interest
Abbreviations
| PDAC | Pancreatic ductal adenocarcinoma |
| USA | United States of America |
| FOLFIRINOX | Folinic acid, 5-fluorouracil, irinotecan, and oxaliplatin |
| EGF | Epidermal growth factor |
| NGF | Nerve growth factor |
| TGF-α | Transforming growth factor alpha |
| TGF-β | Transforming growth factor bêta |
| HGF | Hepatocyte growth factor |
| IGF-I | Insulin-like growth factor 1 |
| IGF-II | Insulin-like growth factor 2 |
| bFGF | Basic fibroblast growth factor |
| EDC | Endocrine-disrupting chemical |
| AR | Androgen receptor |
| PR | Progesterone receptor |
| NP | Nonylphenol |
| BPA | Bisphenol A |
| ERα | Estrogen receptor alpha |
| CDK4 | Cyclin-dependent kinase 4 |
| IL-6 | Interleukin 6 |
| IL-1 | Interleukin 1 |
| BMI | Body mass index |
| VEGF | Vascular endothelial growth factor |
| IDDM | Insulin-dependent diabetes mellitus |
| NIDDM | Non-insulin-dependent diabetes mellitus |
References
- Harbuzariu, A.; Oprea-Ilies, G.; Gonzalez-Perez, R.R. Pancreatic cancer, leptin, and chemoresistance: Current challenges. In Advances in Pancreatic Cancer; Rodrigo, L., Ed.; Intech Open: London, UK, 2018. [Google Scholar] [CrossRef]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Demir, I.E.; Friess, H. Pancreatic cancer—Lessons from the past decade. Indian J. Med. Paediatr. Oncol. 2015, 36, 73–76. [Google Scholar] [PubMed]
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Mace, T.A.; Shakya, R.; Pitarresi, J.R.; Swanson, B.; McQuinn, C.W.; Loftus, S.; Nordquist, E.; Cruz-Monserrate, Z.; Yu, L.; Young, G.; et al. IL-6 and PD-L1 antibody blockade combination therapy reduces tumour progression in murine models of pancreatic cancer. Gut 2018, 67, 320–332. [Google Scholar] [CrossRef]
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouché, O.; Guimbaud, R.; Bécouarn, Y.; Adenis, A.; Raoul, J.L.; Gourgou-Bourgade, S.; de la Fouchardière, C.; et al. Groupe Tumeurs Digestives of Unicancer; PRODIGE Intergroup. FOLFINIROX versus gemcitabine for metastatic pancreatic cancer. N. Engl. J. Med. 2011, 364, 1817–1825. [Google Scholar] [CrossRef]
- von Hoff, D.D.; Ervin, T.; Arena, F.P.; Chiorean, E.G.; Infante, J.; Moore, M.; Seay, T.; Tjulandin, S.A.; Ma, W.W.; Saleh, M.N.; et al. Increased survival in pancreatic cancer with nab-paclitaxel plus gemcitabine. N. Engl. J. Med. 2013, 369, 1691–1703. [Google Scholar] [CrossRef]
- Ducreux, M.; Cuhna, A.S.; Caramella, C.; Hollebecque, A.; Burtin, P.; Goéré, D.; Seufferlein, T.; Haustermans, K.; van Laethem, J.L.; Conroy, T.; et al. ESMO guidelines committee. cancer of the pancreas: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2015, 26, v56–v68. [Google Scholar] [CrossRef]
- Wei, M.Y.; Shi, S.; Liang, C.; Meng, Q.C.; Hua, J.; Zhang, Y.Y.; Liu, J.; Zhang, B.; Xu, J.; Yu, X.J. The microbiota and microbiome in pancreatic cancer: More influential than expected. Mol. Cancer 2019, 18, 97. [Google Scholar] [CrossRef]
- Yadav, D.; Lowenfels, A.B. The epidemiology of pancreatitis and pancreatic cancer. Gastroenterology 2013, 144, 1252–1261. [Google Scholar] [CrossRef]
- Iodice, S.; Gandini, S.; Maisonneuve, P.; Lowenfels, A.B. Tobacco and the risk of pancreatic cancer: A review and meta-analysis. Langenbecks Arch. Surg. 2008, 393, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Chari, S.T.; Leibson, C.L.; Rabe, K.G.; Timmons, L.J.; Ransom, J.; de Andrade, M.; Petersen, G.M. Pancreatic cancer-associated diabetes mellitus: Prevalence and temporal association with diagnosis of cancer. Gastroenterology 2008, 134, 95–101. [Google Scholar] [CrossRef] [PubMed]
- de Gonzalez, A.B.; Sweetland, S.; Spencer, E. A meta-analysis of obesity and the risk of pancreatic cancer. Br. J. Cancer 2003, 89, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Soto, A.M.; Sonnenschein, C. Environmental causes of cancer: Endocrine disruptors as carcinogens. Nat. Rev. Endocrinol. 2010, 6, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N. Endocrine disrupting chemicals and breast cancer: The saga of bisphenol, A. In Estrogen Receptor and Breast Cancer. Cancer Drug Discovery and Development; Zhang, X., Ed.; Humana Press: Cham, Switzerland, 2019; pp. 343–377. [Google Scholar]
- Bachelet, D.; Verner, M.A.; Neri, M.; Duverger, É.C.; Charlier, C.; Arveux, P.; Haddad, S.; Guénel, P. Breast cancer and exposure to organochlorines in the CECILE Study: Associations with plasma levels measured at the time of diagnosis and estimated during adolescence. Int. J. Environ. Res. Public Health 2019, 16, 271. [Google Scholar] [CrossRef]
- Kouidhi, W.; Bergès, R.; Tiffon, C.; Desmetz, C.; El May, M.; Auger, J.; Canivenc-Lavier, M. Perinatal xenohormone exposure impacts sweet preference and submandibular development in male rats. Oral. Dis. 2013, 19, 812–823. [Google Scholar] [CrossRef]
- Mathison, R. Submandibular Salivary Gland Endocrine Secretions and Systemic Pathophysiological Responses. Open Inflamm. J. 2009, 2, 9–21. [Google Scholar]
- Mathison, R. The submandibular glands: A role in homeostasis and allostasis. Biomed. Rev. 1995, 4, 61–69. [Google Scholar] [CrossRef]
- Lee, M.G.; Ohana, E.; Park, H.W.; Yang, D.; Muallem, S. Molecular mechanism of pancreatic and salivary gland fluid and HCO3 secretion. Physiol. Rev. 2012, 92, 39–74. [Google Scholar] [CrossRef]
- Kamisawa, T.; Tu, Y.; Egawa, N.; Sakaki, N.; Inokuma, S.; Kamata, N. Salivary gland involvement in chronic pancreatitis of various etiologies. Am. J. Gastroenterol. 2003, 98, 323–326. [Google Scholar] [CrossRef]
- Sagatelian, M.A.; Fravel, J.; Gallo, S.H.; Makk, L.J.; Looney, S.W.; Wright, R.A. Do parotid duct abnormalities occur in patients with chronic alcoholic pancreatitis? Am. J. Gastroenterol. 1998, 93, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Gittes, G.K. Developmental biology of the pancreas: A comprehensive review. Dev. Biol. 2009, 326, 4–35. [Google Scholar] [CrossRef] [PubMed]
- Rakonczay, Z., Jr.; Vág, J.; Földes, A.; Nagy, K.; Nagy, Á.; Hegyi, P.; Varga, G. Chronic inflammation in the pancreas and salivary glands--lessons from similarities and differences in pathophysiology and treatment modalities. Curr. Pharm. Des. 2014, 20, 1104–1120. [Google Scholar] [PubMed]
- Jennings, R.E.; Berry, A.A.; Strutt, J.P.; Gerrard, D.T.; Hanley, N.A. Human pancreas development. Development 2015, 142, 3126–3137. [Google Scholar] [CrossRef]
- Som, P.M.; Miletich, I. The embryology of the salivary glands: An Update. Neurographics 2015, 5, 167–177. [Google Scholar] [CrossRef]
- Matsuo, R. Role of saliva in the maintenance of taste sensitivity. Crit. Rev. Oral. Biol. Med. 2000, 11, 216–229. [Google Scholar] [CrossRef]
- Neyraud, E. Role of saliva in oral food perception. Monogr. Oral. Sci. 2014, 24, 61–70. [Google Scholar]
- Pedersen, A.M.; Bardow, A.; Jensen, S.B.; Nauntofte, B. Saliva and gastrointestinal functions of taste, mastication, swallowing and digestion. Oral. Dis. 2002, 8, 117–129. [Google Scholar] [CrossRef]
- Marti, U.; Burwen, S.J.; Jones, A.L. Biological effects of epidermal growth factor, with emphasis on the gastrointestinal tract and liver: An update. Hepatology 1989, 9, 126–138. [Google Scholar] [CrossRef]
- Zeng, F.; Harris, R.C. Epidermal growth factor, from gene organization to bedside. Semin. Cell Dev. Biol 2014, 28, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Kouidhi, W.; Desmetz, C.; Nahdi, A.; Bergès, R.; Cravedi, J.P.; Auger, J.; May, M.E.; Canivenc-Lavier, M.C. In utero and lactational exposure to low-dose genistein-vinclozolin mixture affects the development and growth factor mRNA expression of the submandibular salivary gland in immature female rats. Toxicol. Pathol. 2012, 40, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, J.; Wang, P.; Luo, Y.; Yang, X.F.; Yang, X.S.; Li, W.M.; Xu, J. The adverse effects of perinatal exposure to nonylphenol on carbohydrate metabolism in male offspring rats. Int. J. Environ. Health Res. 2017, 27, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, F.L.; Routledge, E.J.; Heidlberger, A.; Rentsch, D.; Guenther, K.; Giger, W.; Sumpter, J.P.; Kohler, H.P. Isomer-specific degradation and endocrine disrupting activity of nonylphenols. Environ. Sci. Technol. 2008, 42, 6399–6408. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Guieysse, B.; Jefferson, B.; Cartmell, E.; Lester, J.N. Nonylphenol in the environment: A critical review on occurrence, fate, toxicity and treatment in wastewaters. Environ. Int. 2008, 34, 1033–1049. [Google Scholar] [CrossRef] [PubMed]
- Vivacqua, A.; Recchia, A.G.; Fasanella, G.; Gabriele, S.; Carpino, A.; Rago, V.; di Gioia, M.L.; Leggio, A.; Bonofiglio, D.; Liguori, A.; et al. The food contaminants bisphenol A and 4-nonylphenol act as agonists for estrogen receptor alpha in MCF7 breast cancer cells. Endocrine 2003, 22, 275–284. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; García-Arévalo, M.; Quesada, I.; Nadal, Á. Bisphenol-A treatment during pregnancy in mice: A new window of susceptibility for the development of diabetes in mothers later in life. Endocrinology 2015, 156, 1659–1670. [Google Scholar] [CrossRef]
- Grün, F.; Blumberg, B. Endocrine disrupters as obesogens. Mol. Cell. Endocrinol. 2009, 304, 19–29. [Google Scholar] [CrossRef]
- Masuo, Y.; Morita, M.; Oka, S.; Ishido, M. Motor hyperactivity caused by a deficit in dopaminergic neurons and the effects of endocrine disruptors: A study inspired by the physiological roles of PACAP in the brain. Regul. Pept. 2004, 123, 225–234. [Google Scholar] [CrossRef]
- Sakata, I.; Tanaka, T.; Yamazaki, M.; Tanizaki, T.; Zheng, Z.; Sakai, T. Gastric estrogen directly induces ghrelin expression and production in the rat stomach. J. Endocrinol. 2006, 190, 749–757. [Google Scholar] [CrossRef]
- Kim, M.S.; Yoon, C.Y.; Jang, P.G.; Park, Y.J.; Shin, C.S.; Park, H.S.; Ryu, J.W.; Pak, Y.K.; Park, J.Y.; Lee, K.U.; et al. The mitogenic and antiapoptotic actions of ghrelin in 3T3-L1 adipocytes. Mol. Endocrinol. 2004, 18, 2291–2301. [Google Scholar] [CrossRef]
- Jubendradass, R.; D’Cruz, S.C.; Mathur, P.P. Long-term exposure to nonylphenol affects insulin signaling in the liver of adult male rats. Hum. Exp. Toxicol. 2012, 31, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Tiffon, C. Non-hereditary risk factors associated with pancreatic cancer: A focus on obesity and diabetes. Integr. Cancer Sci. Ther. 2020, 7, 1–2. [Google Scholar]
- Cascetta, P.; Cavaliere, A.; Piro, G.; Torroni, L.; Santoro, R.; Tortora, G.; Melisi, D.; Carbone, C. Pancreatic cancer and obesity: Molecular mechanisms of cell transformation and chemoresistance. Int. J. Mol. Sci. 2018, 19, 3331. [Google Scholar] [CrossRef]
- Incio, J.; Ligibel, J.A.; McManus, D.T.; Suboj, P.; Jung, K.; Kawaguchi, K.; Pinter, M.; Babykutty, S.; Chin, S.M.; Vardam, T.D.; et al. Obesity promotes resistance to anti-VEGF therapy in breast cancer by up-regulating IL-6 and potentially FGF-2. Sci. Transl. Med. 2018, 10, eaag0945. [Google Scholar] [CrossRef] [PubMed]
- Furmli, S.; Elmasry, R.; Ramos, M.; Fung, J. Therapeutic use of intermittent fasting for people with type 2 diabetes as an alternative to insulin. BMJ Case Rep. 2018, 2018, bcr-2017-221854. [Google Scholar] [CrossRef]
- Giovannucci, E.; Harlan, D.M.; Archer, M.C.; Bergenstal, R.M.; Gapstur, S.M.; Habel, L.A.; Pollak, M.; Regensteiner, J.G.; Yee, D. Diabetes and cancer: A consensus report. Diabetes Care 2010, 33, 1674–1685. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 diabetes mellitus: From a metabolic disorder to an inflammatory condition. World J. Diabetes 2015, 6, 598–612. [Google Scholar] [CrossRef]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The role of inflammation in diabetes: Current concepts and future perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The endocrine society’s second scientific statement on endocrine-disrupting chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Nowak, K.; Jabłońska, E.; Ratajczak-Wrona, W. Immunomodulatory effects of synthetic endocrine disrupting chemicals on the development and functions of human immune cells. Environ. Int. 2019, 125, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Nakayama, A.; Wenger, M.; Burki, R.; Eppler, E.; Krasnov, A.; Segner, H. Endocrine disrupting compounds: Can they target the immune system of fish? Mar. Pollut. Bull. 2011, 63, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Yang, S.N.; Kuo, P.L.; Hung, C.H. Immunomodulatory effects of environmental endocrine disrupting chemicals. Kaohsiung J. Med. Sci. 2012, 28, S37–S42. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G. Lifelong impact of perinatal endocrine disruptor exposures (faulty hormonal imprinting). IJPAES 2019, 9, 94–102. [Google Scholar]
- Kirkegård, J.; Cronin-Fenton, D.; Heide-Jørgensen, U.; Mortensen, F.V. Acute pancreatitis and pancreatic cancer risk: A nationwide matched-cohort study in Denmark. Gastroenterology 2018, 154, 1729–1736. [Google Scholar] [CrossRef]
- Kirkegård, J.; Mortensen, F.V.; Cronin-Fenton, D. Chronic pancreatitis and pancreatic cancer risk: A systematic review and meta-analysis. Am. J. Gastroenterol. 2017, 112, 1366–1372. [Google Scholar] [CrossRef]
- Singh, V.K.; Yadav, D.; Garg, P.K. Diagnosis and management of chronic pancreatitis: A Review. JAMA 2019, 322, 2422–2434. [Google Scholar] [CrossRef]
- Magruder, J.T.; Elahi, D.; Andersen, D.K. Diabetes and pancreatic cancer: Chicken or egg? Pancreas 2011, 40, 339–351. [Google Scholar] [CrossRef]
- Lee, Y.H.; Wong, D.T. Saliva: An emerging biofluid for early detection of diseases. Am. J. Dent. 2009, 22, 241–248. [Google Scholar]
- Zhang, L.; Xiao, H.; Karlan, S.; Zhou, H.; Gross, J.; Elashoff, D.; Akin, D.; Yan, X.; Chia, D.; Karlan, B.; et al. Discovery and preclinical validation of salivary transcriptomic and proteomic biomarkers for the non-invasive detection of breast cancer. PLoS ONE 2010, 5, e15573. [Google Scholar] [CrossRef]
- Katsiougiannis, S.; Wong, D.T. The Proteomics of Saliva in Sjögren’s Syndrome. Rheum. Dis. Clin. N. Am. 2016, 42, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, M.; Szulimowska, J.; Skutnik, A.; Taranta-Janusz, K.; Wasilewska, A.; Wiśniewska, N.; Zalewska, A. Salivary biomarkers of oxidative stress in children with chronic kidney disease. J. Clin. Med. 2018, 7, 209. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, M.; Skutnik-Radziszewska, A.; Zieniewska, I.; Matczuk, J.; Domel, E.; Waszkiel, D.; Żendzian-Piotrowska, M.; Szarmach, I.; Zalewska, A. Antioxidant defense, oxidative modification, and salivary gland function in an early phase of cerulein pancreatitis. Oxid. Med. Cell. Longev. 2019, 8403578-14. [Google Scholar] [CrossRef] [PubMed]
- Sawczuk, B.; Maciejczyk, M.; Sawczuk-Siemieniuk, M.; Posmyk, R.; Zalewska, A.; Car, H. Salivary gland function, antioxidant defence and oxidative damage in the saliva of patients with breast cancer: Does the BRCA1 mutation disturb the salivary redox profile? Cancers (Basel) 2019, 11, 1501. [Google Scholar] [CrossRef] [PubMed]
- Kaczor-Urbanowicz, K.E.; Trivedi, H.M.; Lima, P.O.; Camargo, P.M.; Giannobile, W.V.; Grogan, T.R.; Gleber-Netto, F.O.; Whiteman, Y.; Li, F.; Lee, H.J.; et al. Salivary exRNA biomarkers to detect gingivitis and monitor disease regression. J. Clin. Periodontol. 2018, 45, 806–817. [Google Scholar] [CrossRef]
- Kaczor-Urbanowicz, K.E.; Carreras-Presas, C.M.; Aro, K.; Tu, M.; Garcia-Godoy, F.; Wong, D.T. Saliva diagnostics—Current views and directions. Exp. Biol. Med. (Maywood) 2017, 242, 459–472. [Google Scholar] [CrossRef]
- Cheng, J.; Nonaka, T.; Wong, D.T.W. Salivary exosomes as nanocarriers for cancer biomarker delivery. Materials (Basel) 2019, 12, 654. [Google Scholar] [CrossRef]
- Wong, D.T.W. Salivaomics. J. Am. Dent. Assoc. 2012, 143, 19S–24S. [Google Scholar] [CrossRef]
- Nonaka, T.; Wong, D.T.W. Saliva-exosomics in cancer: Molecular characterization of cancer-derived exosomes in saliva. Enzymes 2017, 42, 125–151. [Google Scholar]
- Zhang, L.; Farrell, J.J.; Zhou, H.; Elashoff, D.; Akin, D.; Park, N.H.; Chia, D.; Wong, D.T. Salivary transcriptomic biomarkers for detection of resectable pancreatic cancer. Gastroenterology 2010, 138, 949–957. [Google Scholar] [CrossRef]
- Li, F.; Kaczor-Urbanowicz, K.E.; Sun, J.; Majem, B.; Lo, H.C.; Kim, Y.; Koyano, K.; Rao, S.L.; Kang, S.Y.; Kim, S.M.; et al. Characterization of human salivary extracellular RNA by next-generation sequencing. Clin. Chem. 2018, 64, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Meleti, M.; Cassi, D.; Vescovi, P.; Setti, G.; Pertinhez, T.A.; Pezzi, M.E. Salivary microRNA for diagnosis of systemic diseases and malignant tumors: A systematic review. Med. Oral Patol. Oral Cirugía Bucal 2020, 25, e299. [Google Scholar]
- Setti, G.; Pezzi, M.E.; Viani, M.V.; Pertinhez, T.A.; Cassi, D.; Magnoni, C.; Bellini, P.; Musolino, A.; Vescovi, P.; Meleti, M. Salivary MicroRNA for diagnosis of cancer and systemic diseases: A systematic review. Int. J. Mol. Sci. 2020, 21, 907. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Yin, X.; Gong, B.; Nie, W.; Wu, B.; Zhang, X.; Huang, J.; Zhang, P.; Zhou, Z.; Li, Z. Salivary microRNAs show potential as a noninvasive biomarker for detecting resectable pancreatic cancer. Cancer Prev. Res. (Phila) 2015, 8, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Farrell, J.J.; Zhang, L.; Zhou, H.; Chia, D.; Elashoff, D.; Akin, D.; Paster, B.J.; Joshipura, K.; Wong, D.T.W. Variations of oral microbiota are associated with pancreatic diseases including pancreatic cancer. Gut 2012, 61, 582–588. [Google Scholar] [CrossRef]
- Lau, C.; Kim, Y.; Chia, D.; Spielmann, N.; Eibl, G.; Elashoff, D.; Wei, F.; Lin, Y.L.; Moro, A.; Grogan, T.; et al. Role of pancreatic cancer-derived exosomes in salivary biomarker development. J. Biol. Chem. 2013, 288, 26888–26897. [Google Scholar] [CrossRef]


© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiffon, C. Defining Parallels between the Salivary Glands and Pancreas to Better Understand Pancreatic Carcinogenesis. Biomedicines 2020, 8, 178. https://doi.org/10.3390/biomedicines8060178
Tiffon C. Defining Parallels between the Salivary Glands and Pancreas to Better Understand Pancreatic Carcinogenesis. Biomedicines. 2020; 8(6):178. https://doi.org/10.3390/biomedicines8060178
Chicago/Turabian StyleTiffon, Céline. 2020. "Defining Parallels between the Salivary Glands and Pancreas to Better Understand Pancreatic Carcinogenesis" Biomedicines 8, no. 6: 178. https://doi.org/10.3390/biomedicines8060178
APA StyleTiffon, C. (2020). Defining Parallels between the Salivary Glands and Pancreas to Better Understand Pancreatic Carcinogenesis. Biomedicines, 8(6), 178. https://doi.org/10.3390/biomedicines8060178