1. Introduction
What is the physical mechanism of memory, and where does the brain store our memories? These are still open questions. In conventional neuroscience, the synaptic plasticity among neurons corresponds to memories stored in a brain by strengthening the connections involved in more frequent signal transmission resulting from specific activities. External stimulations change synaptic signaling and enhance signal transfer among neurons, which is called long-term potentiation. However, if synaptic plasticity is required for storage of memory, how do we explain memory in single-cell organisms [
1]? To explain memory storage, even for single-cell organisms, we need to include molecular mechanisms involving the cytoskeleton structures inside cells whose organization of internal degrees of freedom could serve as memory storage mechanisms, as previously proposed by others [
2]. Furthermore, it is important to briefly discuss the issue of energy efficiency characterizing the human brain. We require, on average,
of power for the brain to function under normal circumstances, which is extremely small compared with man-made computational technology, with a power demand as high as
(we refer to [
3]) for AlphaGo in DeepMind, where a human won one of five games, in spite of the orders-of-magnitude difference in the respective power needs. How do we realize this extremely high energy efficiency for information processing taking place in the brain within the framework of classical information processing? This is a major challenge. We propose a different approach to resolve this issue. Our strategy is to adopt quantum information processing and quantum information storage in the cytoskeleton.
Cellular cytoskeletons construct a network of proteins that are indispensable for the performance of key cellular processes involving growth, molecular transport, internal structural reorganization, cell division and motility, as shown by Lee et al. [
4]. We find microfilaments (also referred to as actin filaments), intermediate filaments and microtubules as the main components of cytoskeletons. Tryptophans in microtubules can play the role of information processors. Craddock et al. suggested that high-capacity memory storage can be achieved in microtubules by CaMKII phosphorylation of specific residues in its building blocks, namely tubular dimers [
2]. Additionally, biophoton mechanisms may play a role in fast intra- and inter-cellular communication, since both super-radiant (fast decay) and sub-radiant (slow decay) states have been experimentally determined to emerge for tryptophan excitations, as shown in [
5,
6,
7]. Moreover, Babcock et al. investigated tryptophan mega-networks in microtubules, where super-radiant processes can emerge, even under thermal equilibrium conditions [
8]. Super-radiant photon emission can lead to quantum information transfer, as two separate quantum dots conduct photonic information processing, as shown in an experimental study [
9]. Super-radiance realizes long-range atom–atom interactions under conditions where atoms are confined to one spatial dimension [
10]. Furthermore, since microtubules are surrounded by mitochondria, the production of reactive oxygen species (ROS) by mitochondria (as a byproduct of ATP-based energy production) can generate ultra-weak photon emission within cells [
11]. Since microtubules have been shown to absorb these photons, store them for a period of time and release them into the environment by de-excitation, microtubule networks may act to transfer and, perhaps, process ROS-generated photons that may contain quantum information (qubits). Zapata et al. suggested ultra-weak photon emission due to oxidative stress [
12]. Shirmovsky studied quantum entanglement in microtubule tryptophan systems [
13,
14]. Moreover, circadian clock activity might be explained by photo-reduction mediated by tryptophan [
15].
Additionally, water degrees of freedom might play a role, due to their dipole moments, by extending local events occurring in microtubules to diffused, non-local events in distal regions of the brain. In fact, quantum brain dynamics, which is a Quantum Field Theory (QFT) of water electric dipoles and photon degrees of freedom, represents a mathematically developed hypothesis of memory formation in such a system. It originated with the work reported in [
16,
17,
18]. The QFT approach to biological systems was also further elaborated on in the 1980s in a series of seminal papers [
19,
20,
21,
22,
23]. Water and photon degrees of freedom were explicitly introduced in QBD by Jibu and Yasue [
24]. These degrees of freedom might amplify the effects of polarization of dipoles in biological systems, namely tryptophan residues abundantly present in microtubules, and may extend local events taking place in tubulin’s tryptophans to non-local information diffused in the whole brain. Then, information contained in the local events might be converted to non-local holographic information. The holographic approach was first proposed by Pribram [
25,
26], who was a collaborator of Jibu and Yasue. We described holography within the framework in QBD in our earlier work [
27,
28,
29], where rotational degrees of freedom of water molecules were adopted. Super-radiant photon emission might be viewed as coherent light for interference patterns (for holography) of reference waves and object waves irradiated on microtubules [
30,
31,
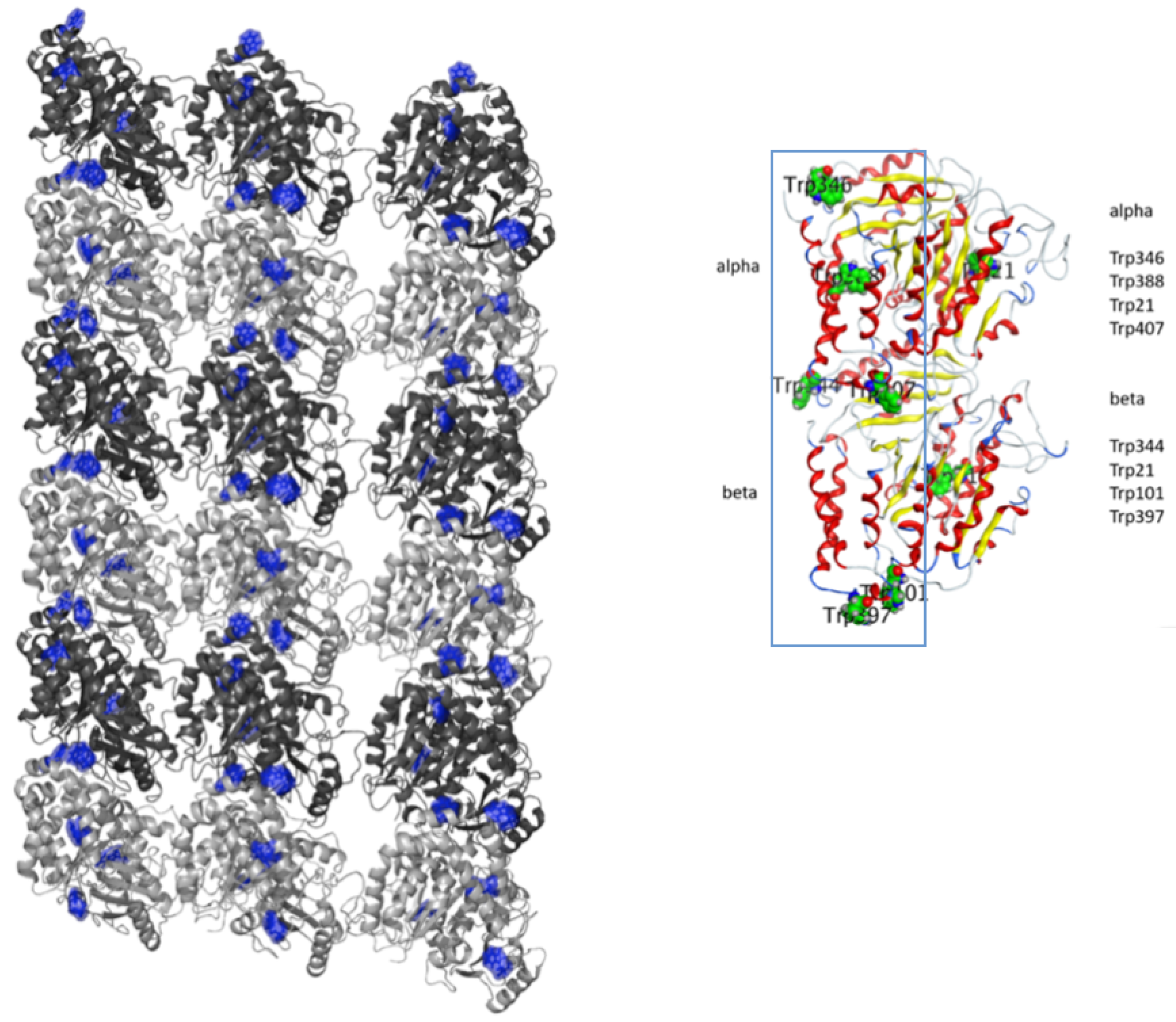
32]. We consider tryptophans in microtubules as candidates for local memory in the brain. As shown in
Figure 1, we find that six (of eight) tryptophans in a tubulin dimer form a one-dimensional waveguide; then, we can adopt waveguide quantum electrodynamics, where qubits with two energy levels are coupled with photon modes in one spatial dimension. Tryptophans might also be entangled with surrounding water molecules. Here, we can adopt properties of water molecules for absorption and emission in a large variety of wavelengths, especially with the same frequency of absorption for water molecules (electronically resonant mode) as that of tryptophans. Finally, a recent paper indicated an important role of membrane dipoles in neutrons as participants in holographic image formation in the brain [
33].
In this paper, we consider tryptophan residues in a microtubule lattice as data qubits entangled with surrounding water molecules that can be utilized for memory storage in a brain. Typically, a microtubule consists of 13 protofilaments involving tubulin dimers as their building blocks. We consider a protofilament as a potential waveguide for propagating photons among qubits aligned in one dimension. We aim to investigate the robustness of tryptophan data qubits entangled with water molecules as Josephson quantum filters adopting sub-radiance. We first introduce a Hamiltonian within the framework of waveguide quantum electrodynamics involving qubits coupled with several photon modes. Next, we adopt Heisenberg equations for operators and derive time-evolution equations for probability coefficients in both zero- and finite-temperature cases. Using the time-evolution equations, we show the robustness of data qubits coupled with Josephson quantum filters that correspond to water molecules in our numerical simulations. Sub-radiance is found to be a key concept for the resultant robustness.
This paper is organized as follows. In
Section 2, we show time-evolution equations using Heisenberg equations derived using the Hamiltonian of waveguide QED. In
Section 3, we show numerical results illustrating the robustness of data qubits. In
Section 4, we discuss our results. In
Section 5, we provide concluding remarks and future perspectives. In this paper, we use the natural units where the speed of light, the Planck constant (
ℏ) and the Boltzmann constant are all set to unity.
2. Time-Evolution Equations at Zero and Finite Temperature
In this section, we provide time-evolution equations in waveguide QED [
34] at zero and finite temperature. The degrees of freedom are qubits involving the ground state (
) and an excited state (
), as well as photons coupled with qubits arranged in
dimensions.
We begin with the Hamiltonian,
where we introduce
as well as creation and annihilation operators for photons with momentum (
k) by
and
. The integration in
is set from
to
∞ or from 0 to
∞. We introduce coupling (
) between qubits and photons as follows:
where
represents the decay rate of the
jth qubit in position
in one spatial dimension and
represents the boundary condition for the coupling. We find the commutation relations for these operators as follows:
The commutations between two creation operators and between two annihilation operators are zero.
In the Heisenberg representation, we find Heisenberg equations and the corresponding Hermitian conjugates as follows:
We then investigate relaxation processes at zero temperature. The initial state involving excitation present only in the first qubit including
is given by
with the ground state being
with
,
. Due to the Schrödinger equation, we can write
We then find
with
. Similarly, we find
Using Heisenberg Equations (
6) and (7), we then derive the following relationship:
In the above equations, we obtain the conservation law for excitation numbers as follows:
where
represents the occupation probability of state
and
is related to the distribution of photons with momentum (
k) at time
t.
The solution of
in Equation (13) is
Setting the lower bound of
k from zero to
and using the solution of
, we then expand Equation (
12) as
If the limits of integration over k are from 0 to ∞, we require the factor to be multiplied by the second term on the right-hand side of the above equation. The first term on the right-hand side represents the frequency term, while the second term involves the decay factors (). The damping oscillation for the excitation of a qubit is found to emerge in time evolution.
We now extend our approach to quantum dynamics at finite temperature. The initial density matrix (
) is represented by
with
and the ground state of qubits (
). We consider a heat bath of photons with a temperature of
. Since unitary evolution emerges for the density matrix, we write
Here,
represents the projection onto the state involving one excitation, which is the approximation adopted in our approach. The excitation numbers for the density matrix are written as
with the Bose–Einstein distribution (
) for temperature (
).
We can also introduce the other elements of the density matrix as
Using Heisenberg Equations (
6) and (7), we arrive at
The variables we should trace are
,
,
and
. Here, we can represent the conservation law as
The value of the bracket in the above equation using Equation (23) deviates from the excitation numbers represented by in Equation (19) at a finite temperature. This is because of the projection in Equation (18). Although the conservation law is modified at a finite temperature, the conserved quantity is still present in our approach.
We also solve the following case:
where we introduce the damping term to the initial thermal equilibrium state with the damping parameter (
) to describe the internal loss representing the absorption of photons by the environment.
4. Discussion
In this paper, we introduced the Hamiltonian of waveguide quantum electrodynamics (wQED) and Heisenberg equations for qubits and photons and applied this formalism to quantum information processes in microtubules. We introduced time-evolution equations for the expectation values of the product of creation and annihilation operators of qubits and photons with a density matrix at finite temperature within the framework of wQED and subsequently solved the system’s time evolution using numerical simulations. We determined the robustness of data qubits representing tryptophans as data qubits (DQs) entangled with qubits for Josephson quantum filters (JQFs) representing water molecules in numerical simulations by increasing the decay rate of qubits for JQF. We also demonstrated that sub-radiant states represent a key concept for robustness, as discussed below in this section.
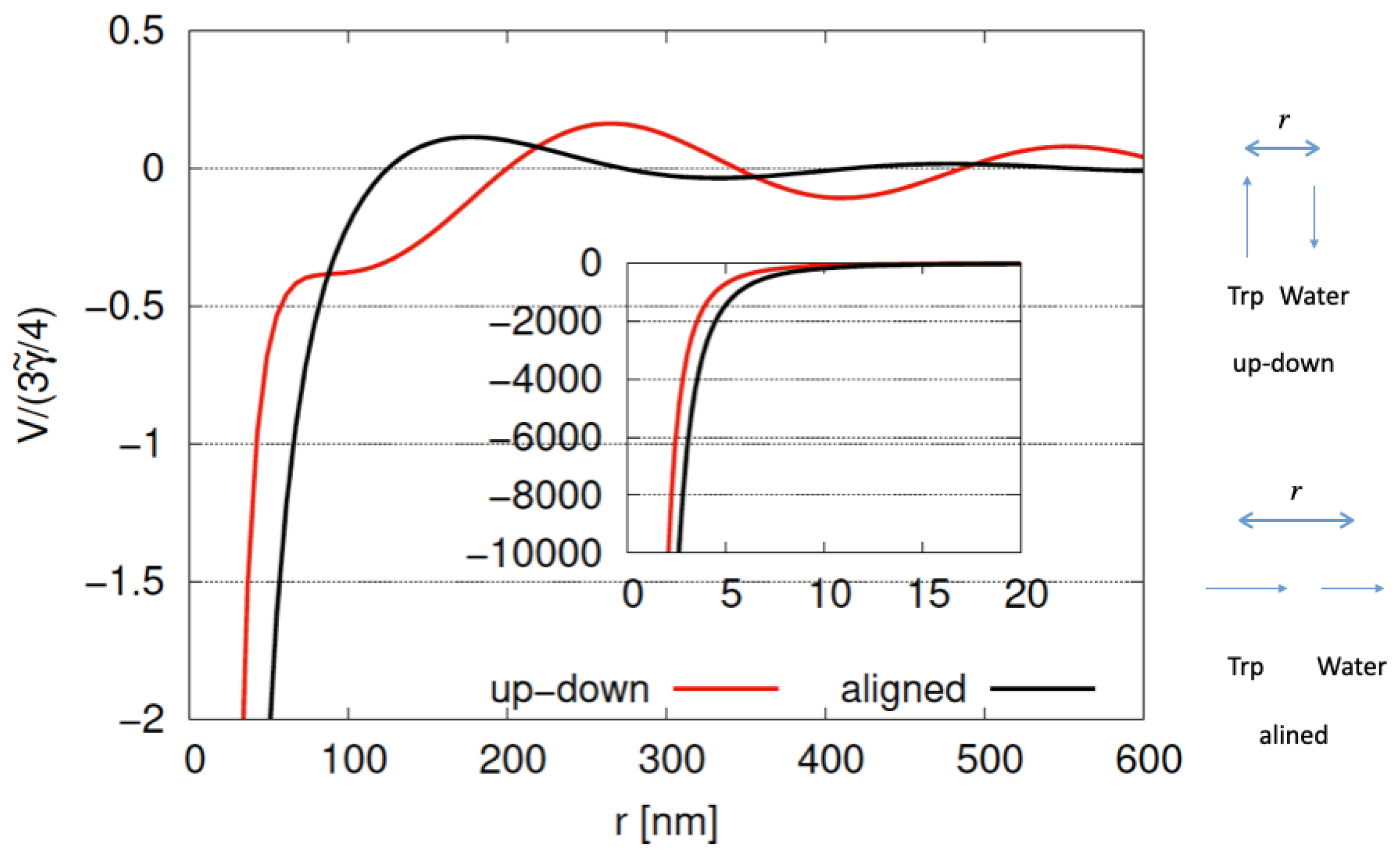
The potential energy (
V) between water and tryptophan dipoles [
7] is shown in
Figure 18 for
where
r represents the distance between water and tryptophan (Trp) molecules;
represents the energy difference between the ground state and the first excited state;
,
and
represent normalized dipole moment vectors for Trp
and water molecule
, respectively;
represents the normalized relative coordinate between Trp and water given by
; and
is
(with relative permittivity
) representing
. We show cases of an up–down configuration and an aligned configuration for Trp and water dipoles in
Figure 18. As shown in
Figure 18, we find a large decrease in potential energy for both up–down and aligned configurations, representing large attractive forces between dipoles. For
, the potential energy for the aligned configuration is lower than that for the up–down configuration. As
r increases, potential energy gradually approaches zero with a wavelength of
but still showing long-range interaction (
). Long-range interaction might play the role of collective behaviors of Trp and water systems for large scales of microtubules. Trapped water molecules for Trp have been studied in several experimental and theoretical studies. Tryptophans with trapped protonated water molecules (TrpH
+(H
2O)
3 and TrpH
+(H
2O)
5) were investigated using cold-ion spectroscopy [
35]. The dielectric property of water molecules trapped in tryptophan residue depending on pH was shown in fluorescence emission spectra [
36]. Trapped water molecules around tryptophans are key factors of protein fluorescence, since the intrinsic fluorescence of tryptophans is sensitive to the environment surrounding solvents or amino acids [
37,
38]. In cases in which Trp molecules attract surrounding water molecules and their distances are at atomic scales or extremely small compared with a wavelength of
, quantum effects, such as entanglement, superposition and so on, might emerge. Microtubules involving tryptophans have been shown to be effective light harvesters [
5,
6]. Aromatic residues, such as tryptophans and tyrosines, might be adopted for information processing roles. Therefore, we need quantum models to describe the fluorescence of tryptophans with surrounding water molecules. Such models might be applied to describe the quantum information processing of tryptophans.
We consider super-radiant and sub-radiant states for two qubits (
) with a decay rate of
. We investigate the following decay matrix (
) appearing on the right-hand side in Equation (16) as follows:
We then find two eigenstates as follows:
The eigenstate expressed as
represents a super-radiant state with a large decay rate
. On the other hand, the eigenstate expressed as
represents a sub-radiant state with a decay rate of 0. In the case of
, this sub-radiant state corresponds to
, representing a spin-0 state of two spin particles (
). The sub-radiant state is decoupled from spin-1 states, namely
,
and
, corresponding to intermediate states in a decaying processes for super-radiant states. Let us now increase the ratio
from 1. In the case of
, the sub-radiant state (
) is approximately
. As a result, the state of
for qubit 1 coupled with qubit 2 with an extremely large decay rate of
is stable over the course of time evolution. We propose the adoption of sub-radiance to achieve robustness of data qubits of tryptophans entangled with JQFs of water molecules. Our approach to maintain quantum information is distinguished from making use of quantum Zeno effects involving continuous measurement procedures [
39]. We adopt the time evolution of expectation values of operators based on Heisenberg equations. Then no measurement procedures are necessary. We prepare two qubits with decay rates of
and
. Quantum entanglement between two qubits then suggests a super-radiant state with a decay rate of
and sub-radiant state with a vanishing decay rate in a Heisenberg equation with a total decay rate of
remaining. We can then use sub-radiance with a vanishing decay rate for data qubits.
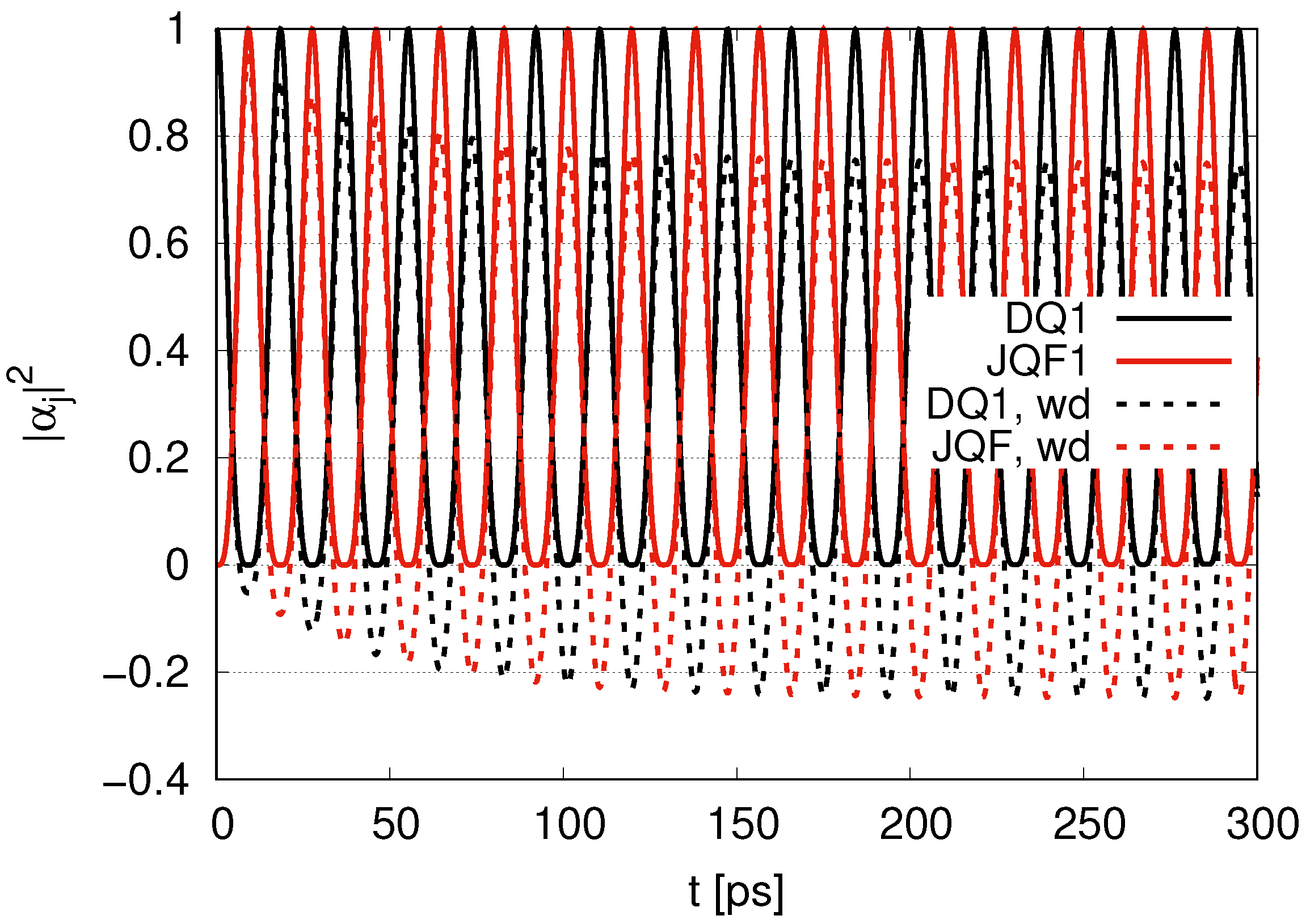
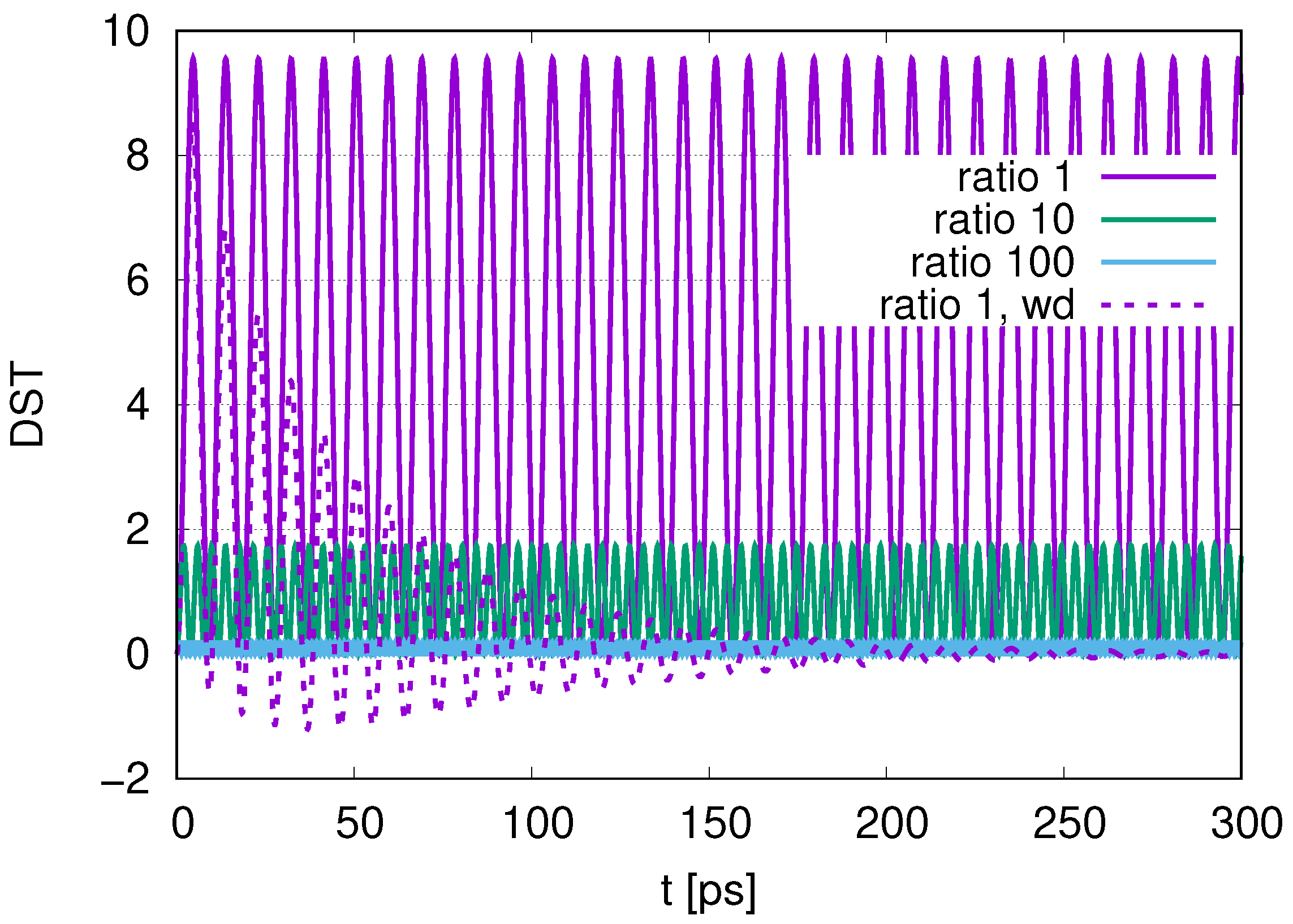
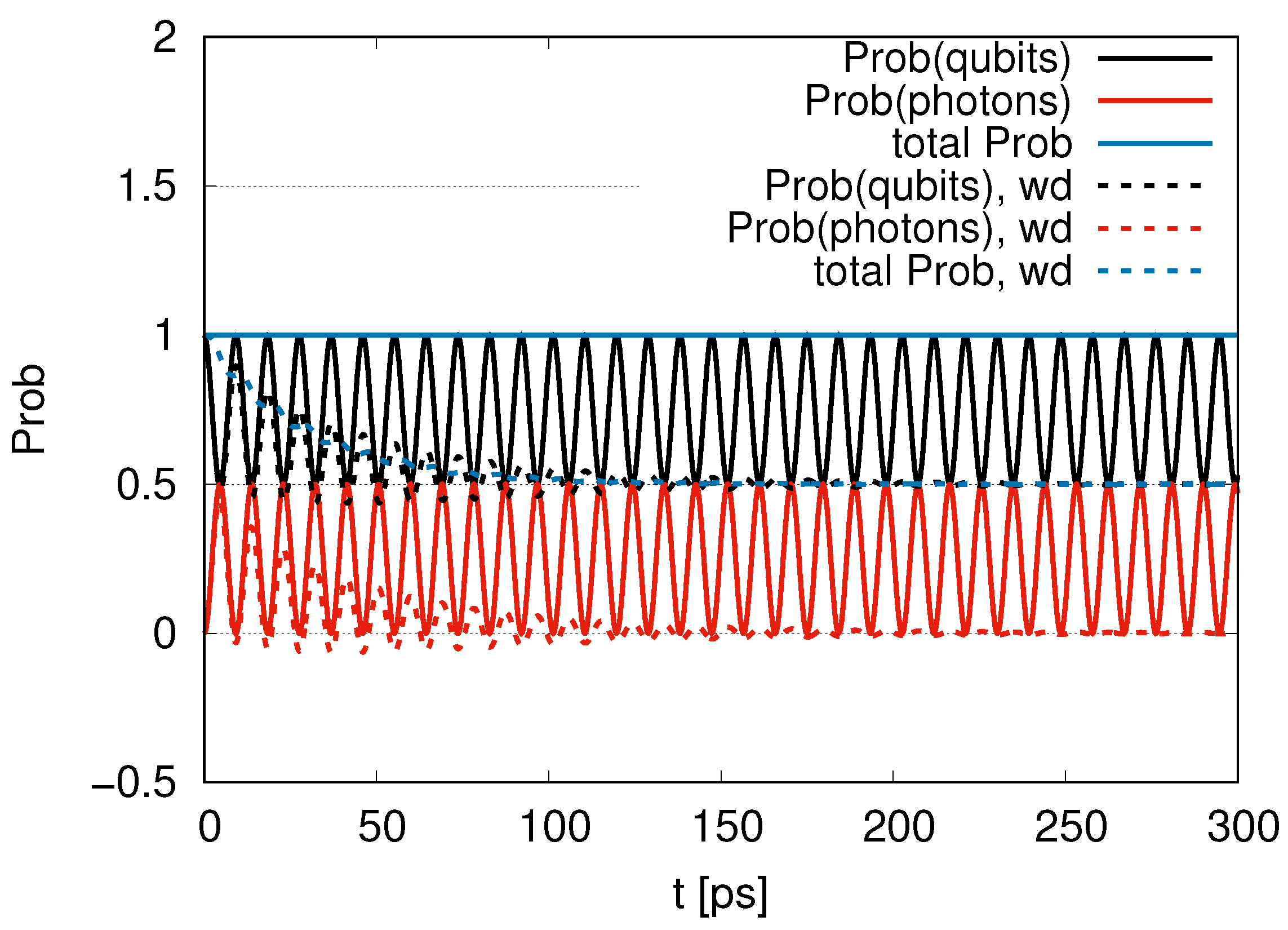
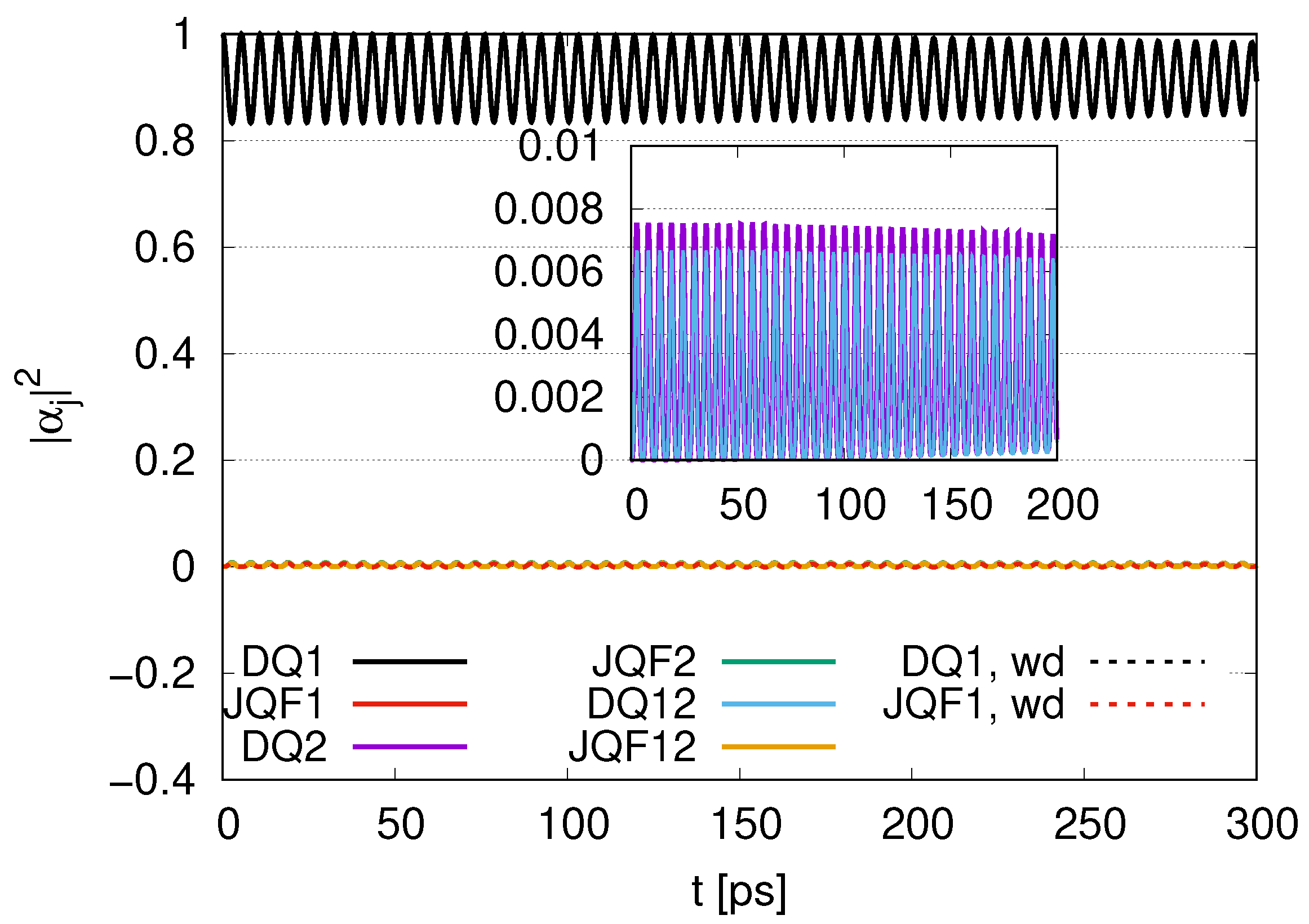
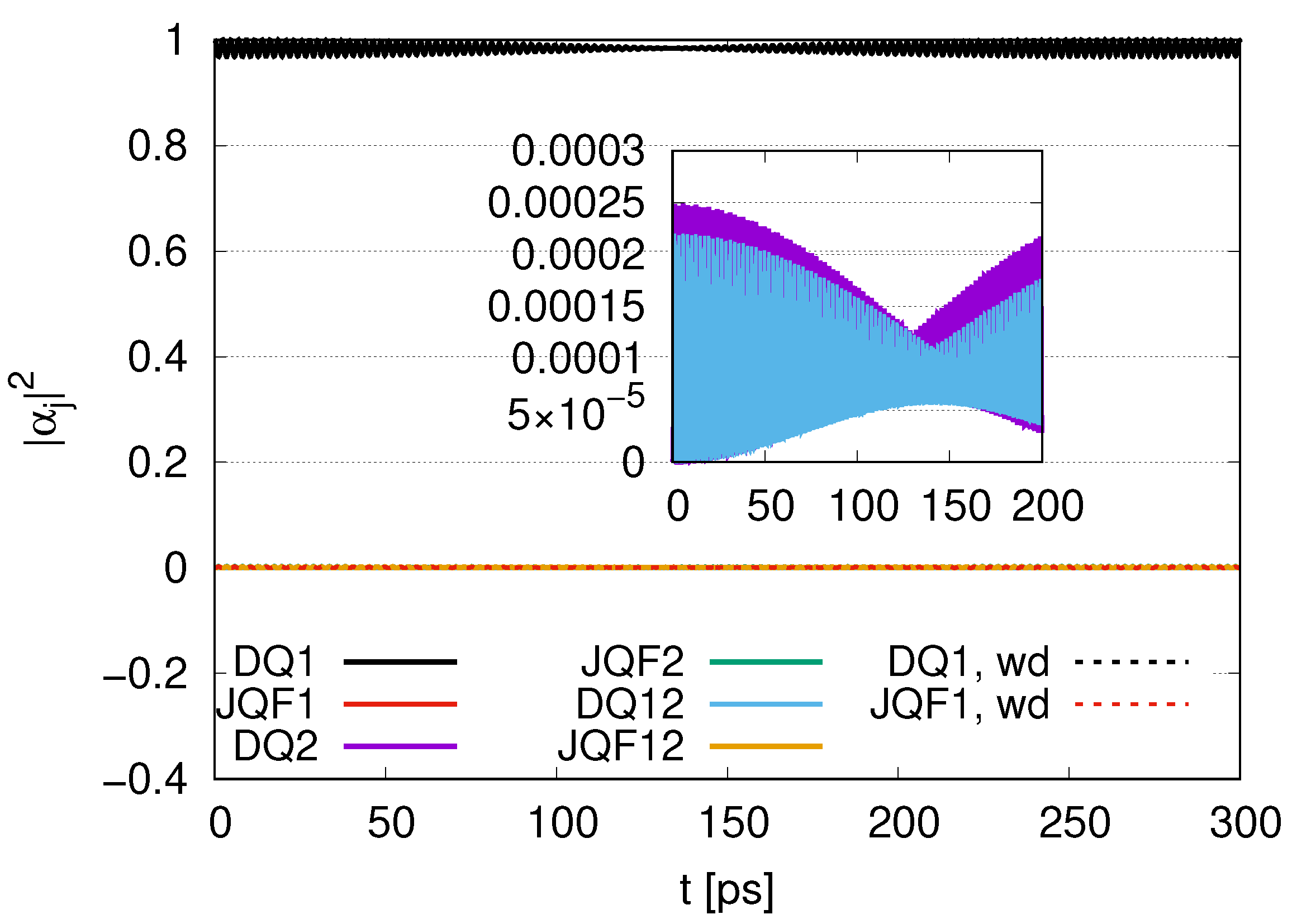
Sub-radiance emerging with an increasing ratio is shown in
Section 3.1, where we introduced two qubits (one DQ and one JQF) in the same position (
). For the ratio of the decay rates (
) in
Figure 2, we find an interplay of probability between DQ and JQF. There is no distinction between DQ and JQF. In the case of a ratio 10, as shown in
Figure 3, the average values of
for DQ and JQF are 0.83 and 0.17, respectively. Here, determine the ratio to be
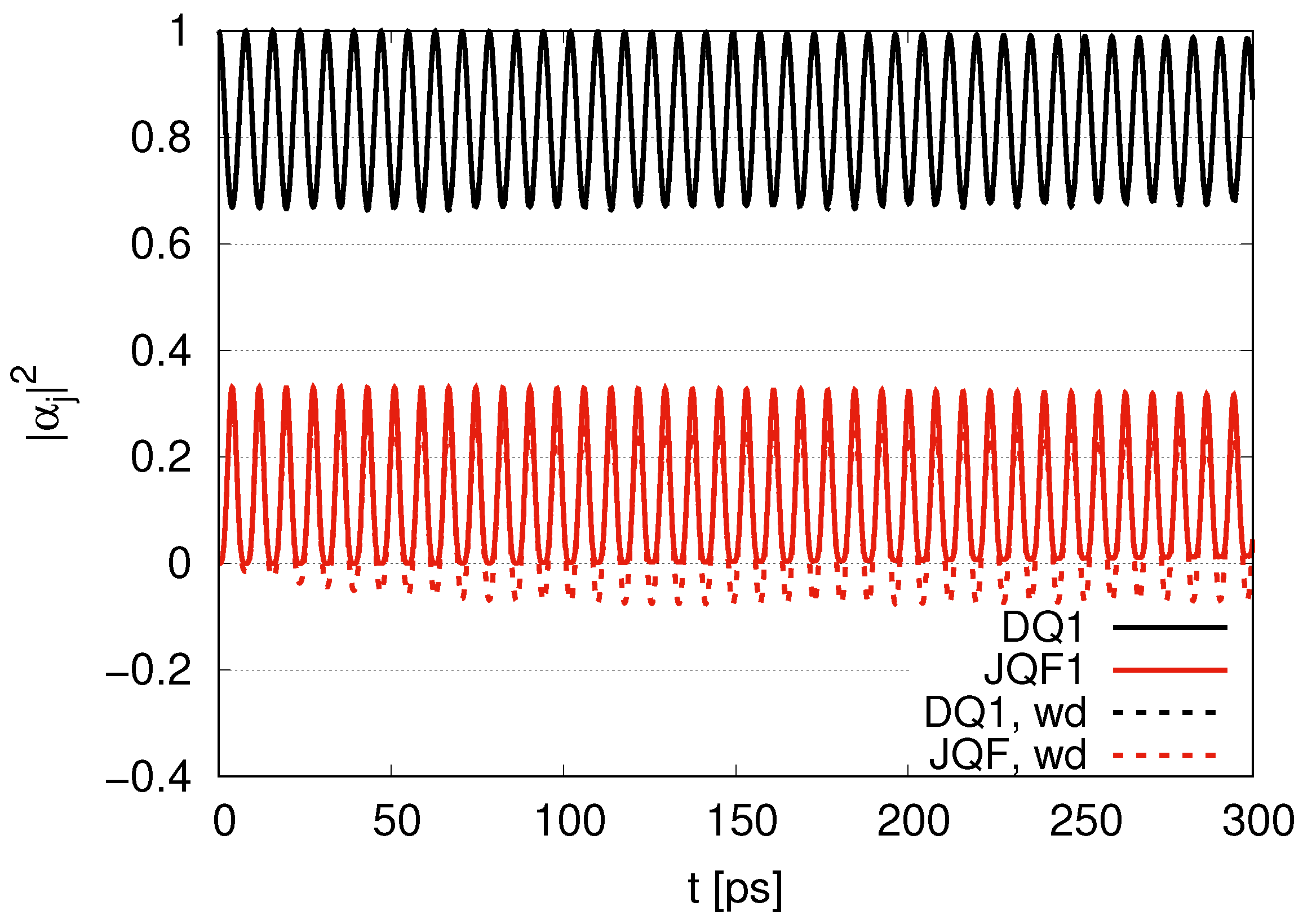
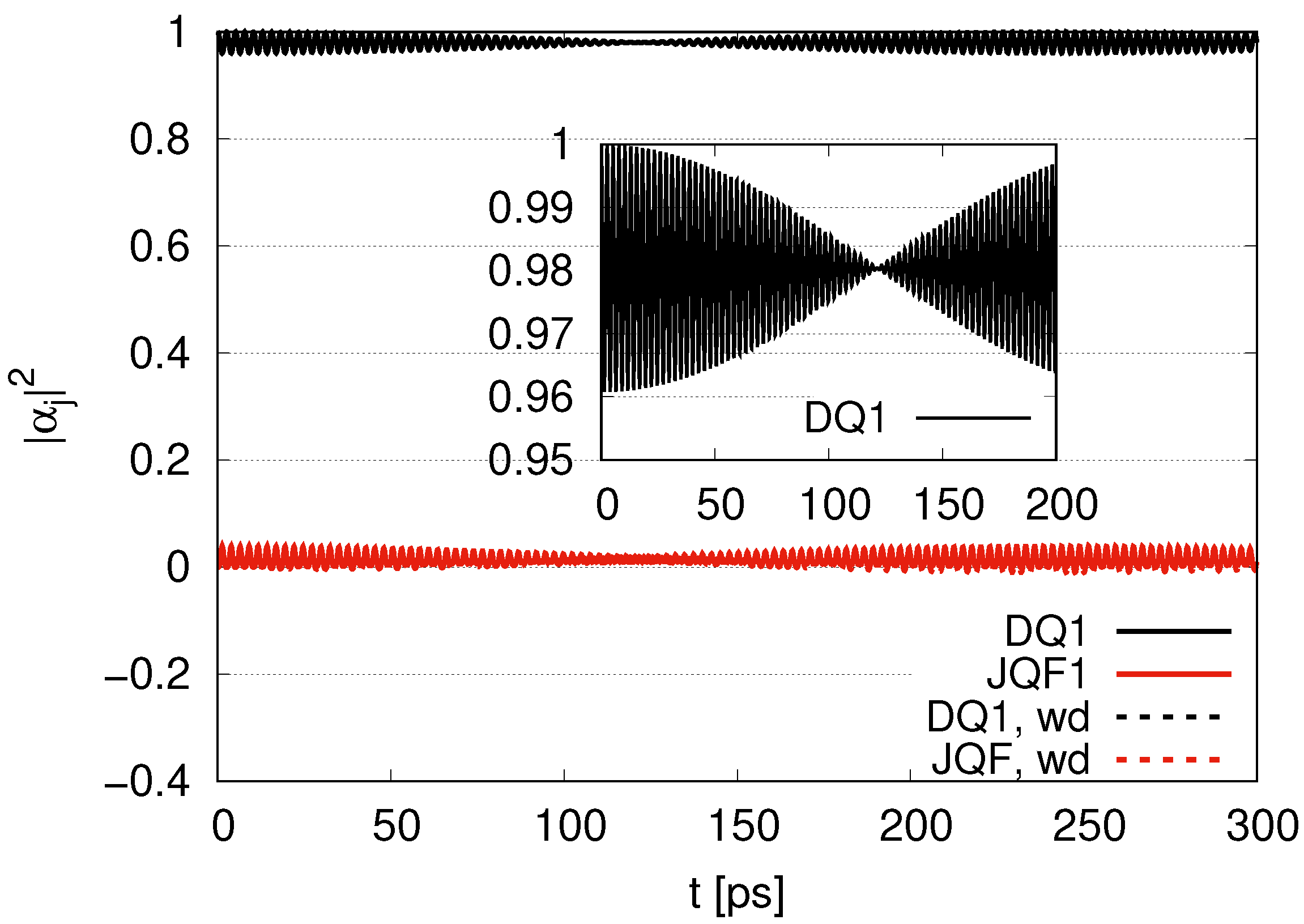
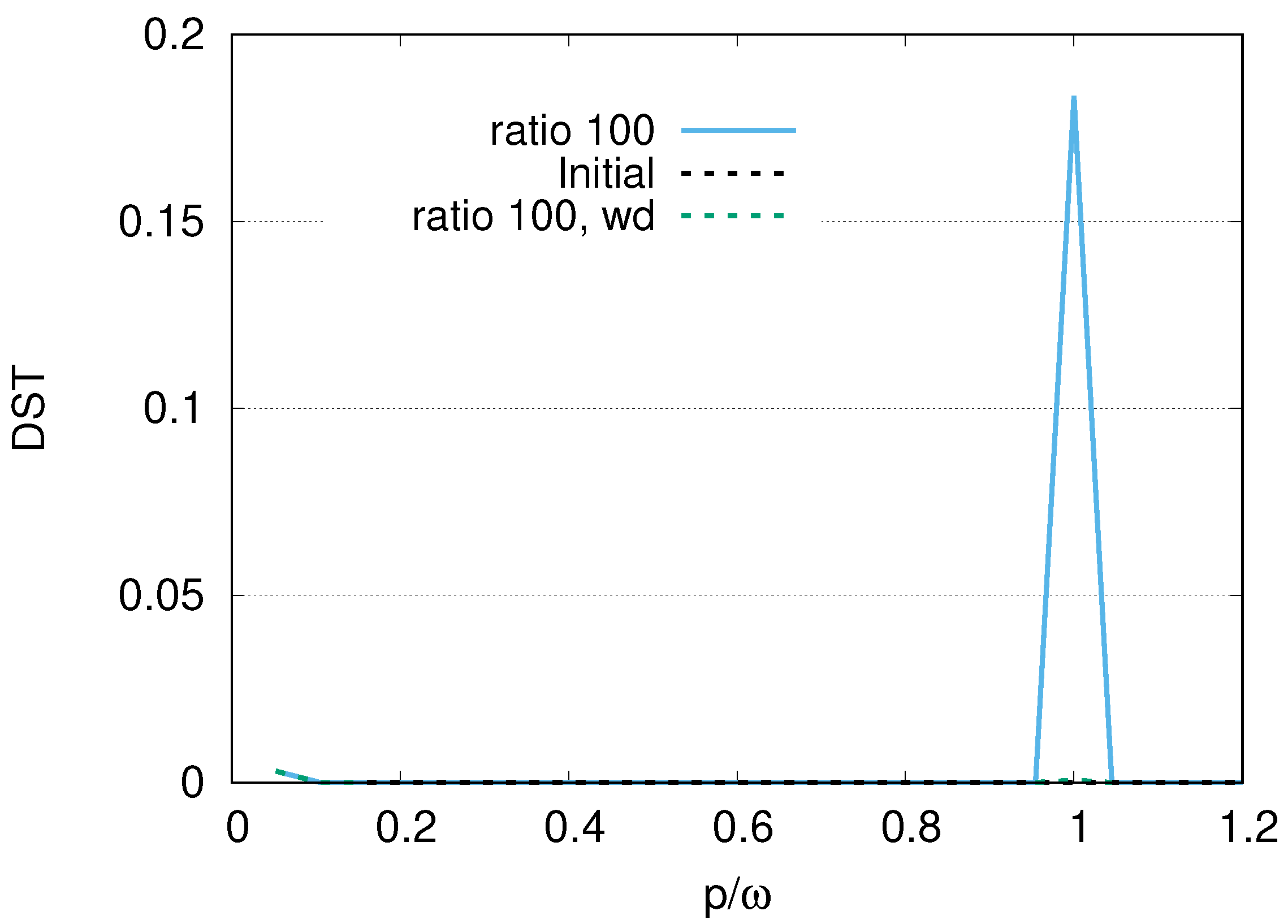
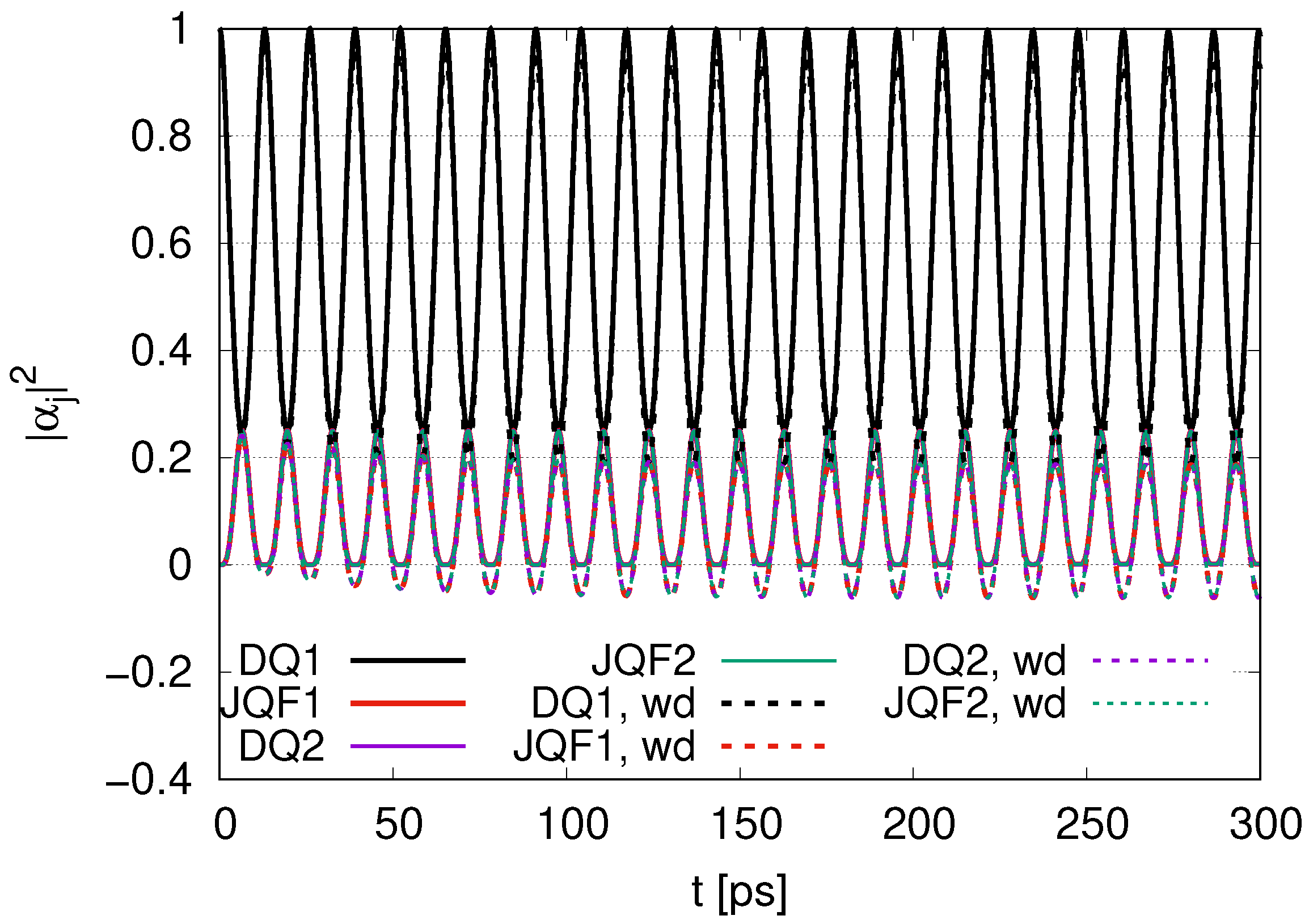
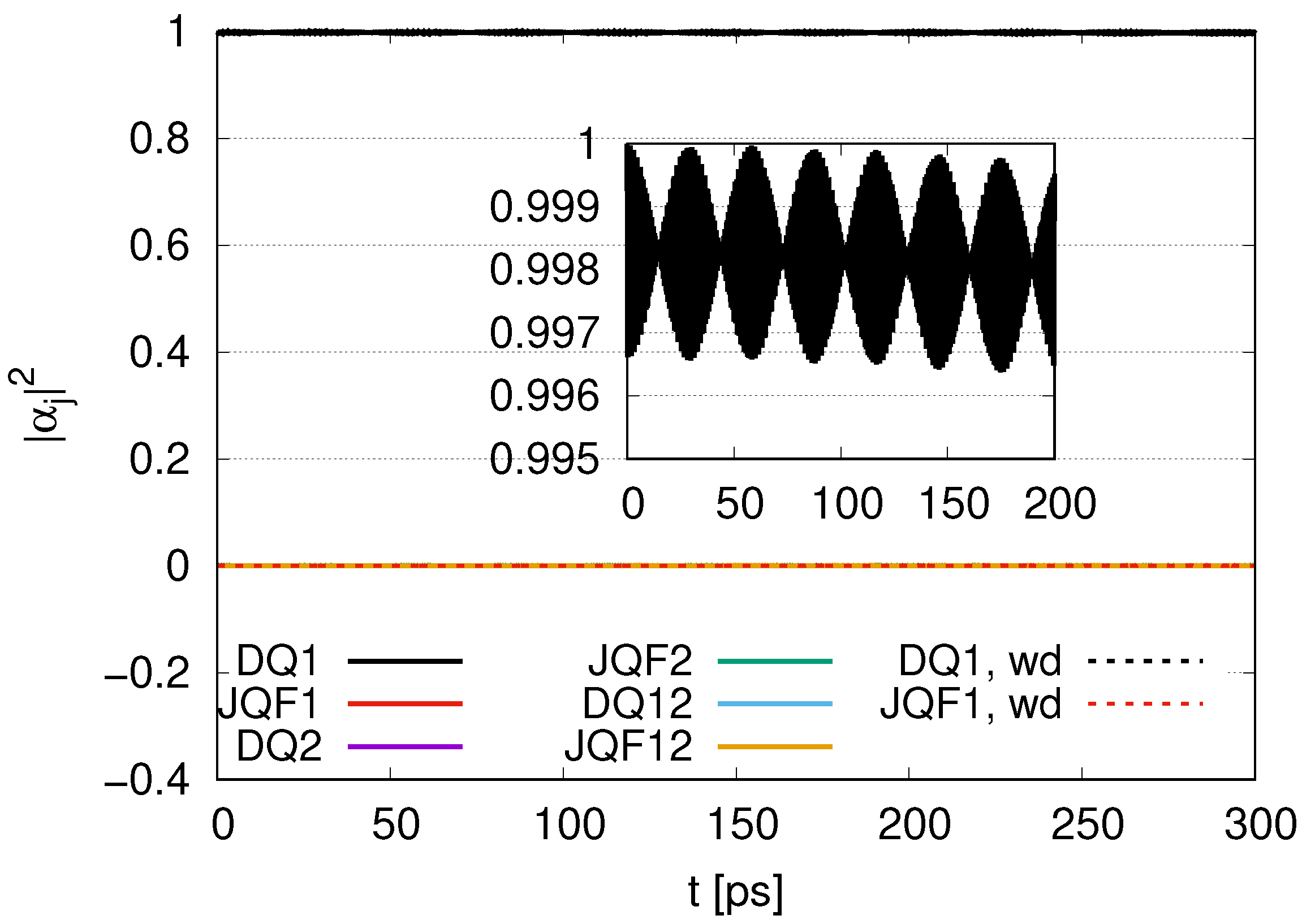
. Since the ratio of the decay rates is 10, the ratio 4.9 is roughly one half of 10. This is because part of the excitation of DQ is transferred to photon modes. In the case of a ratio 100, as shown in
Figure 4, the average values of
for DQ and JQF are 0.98 and 0.02, respectively. We find a ratio
and that the order is half of 100 due to contributions of the excitation of photons. As we increase the ratio of decay rates for DQ and JQF, the
for DQ remains around the initial value of 1. For a ratio 100, the corresponding time scales are
and
, which might correspond to the time scales of internal losses and are 20 times larger than thermalization time scales (
) for water–photon systems where we set the mean free path of water molecules to
and the mean velocity of water molecules to
, finding
with four collision processes.
The other type of sub-radiance for a ratio 1 involving 4 and 24 qubits is shown in
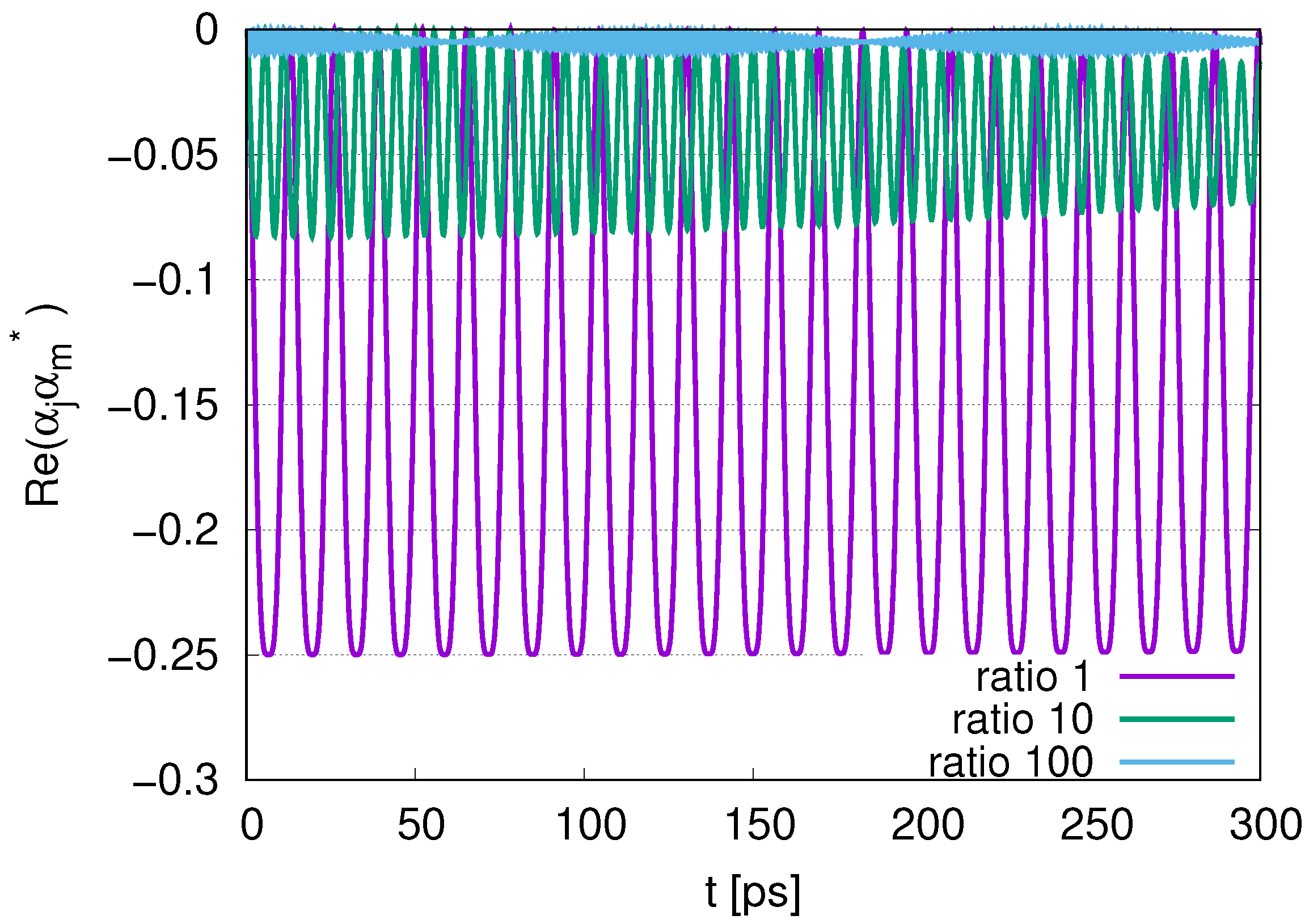
Figure 8 and
Figure 14. Even if the ratio is 1, we find that
for DQ1 oscillates between 0.25 and 1, as shown in
Figure 8. We infer that the other type of the sub-radiant state (
) appears since Re
with
DQ1 and
DQ2 has negative values between
and 0, as shown in
Figure 11. The ratio of coefficients for DQ1 and DQ2 is
, while the ratio of the average of
for DQ1 and
for DQ2 is
, which is smaller than 9 because excitations of qubits are transferred to photon excitations. Similarly, we also infer that the sub-radiant state (
) appears. The ratio of the coefficients for DQ1 and other qubits is
, while the ratio of the averages of
for DQ1 and
for DQ2 is
. The ratio of
for DQ1 and
for DQ12 is
, approaching the order of 529. The initial state of DQ1 is protected by other surrounding qubits under this type of sub-radiance. However, comparing it with the sub-radiance in making use of the different decay rates for DQ and JQF, the deviation of
for DQ1 from its initial value 1 is dependent on the number of surrounding qubits, which means that we should use more surrounding qubits for the stability of data. This represents the waste of resources in terms of qubits. Making use of surrounding water molecules with larger decay rates than JQFs might be more advantageous in protecting data contained in a microtubule.
The temperature dependence is extremely low in the course of the time evolution of qubits. This is because for tryptophan is extremely large compared with a temperature of . Although we investigated several temperatures (, and ) for a total of two qubits, when the ratio of decay rates of DQ and JQF is 10, there are minimal changes in for DQ1. As the temperature approaches the order of , the amplitudes of oscillations of gradually increase.
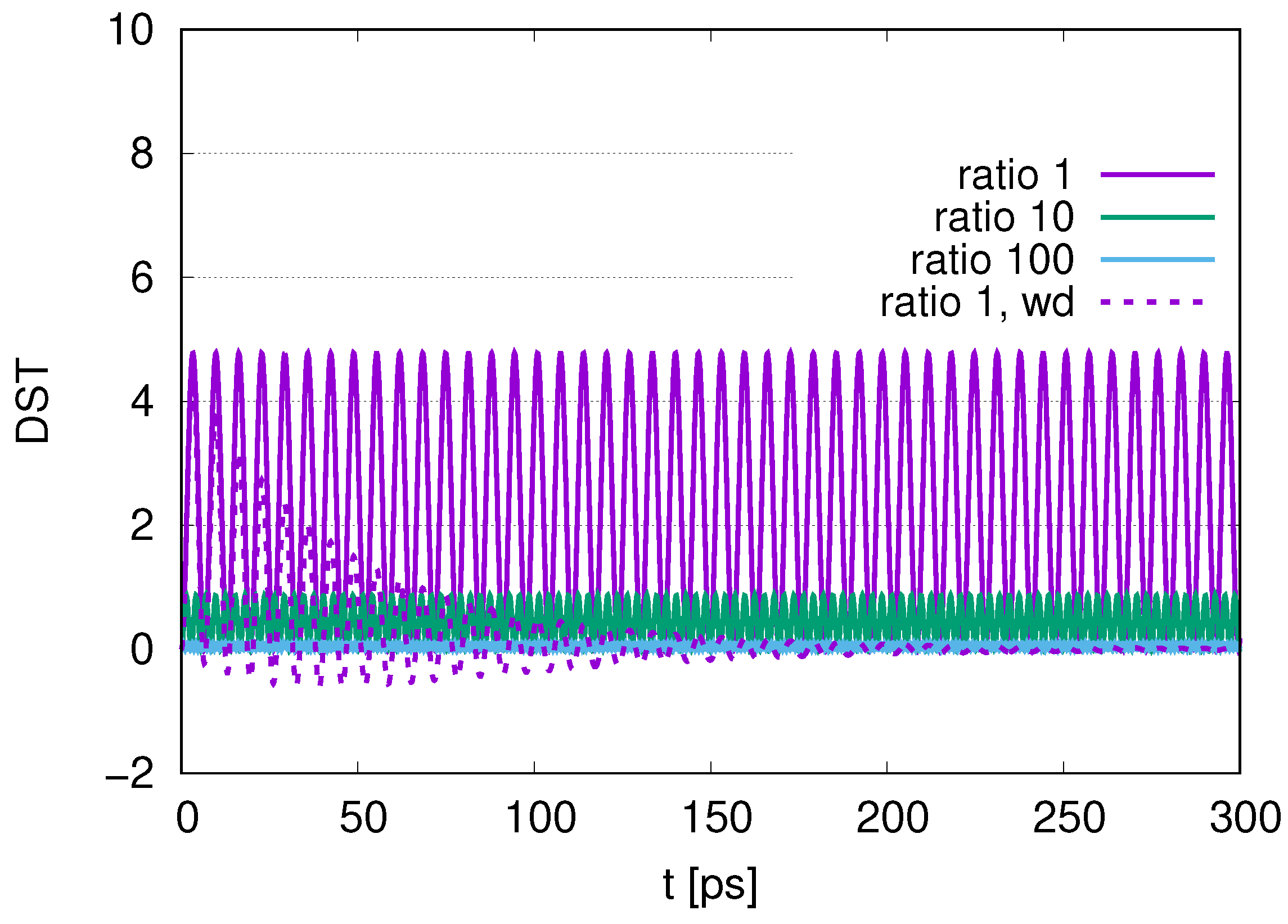
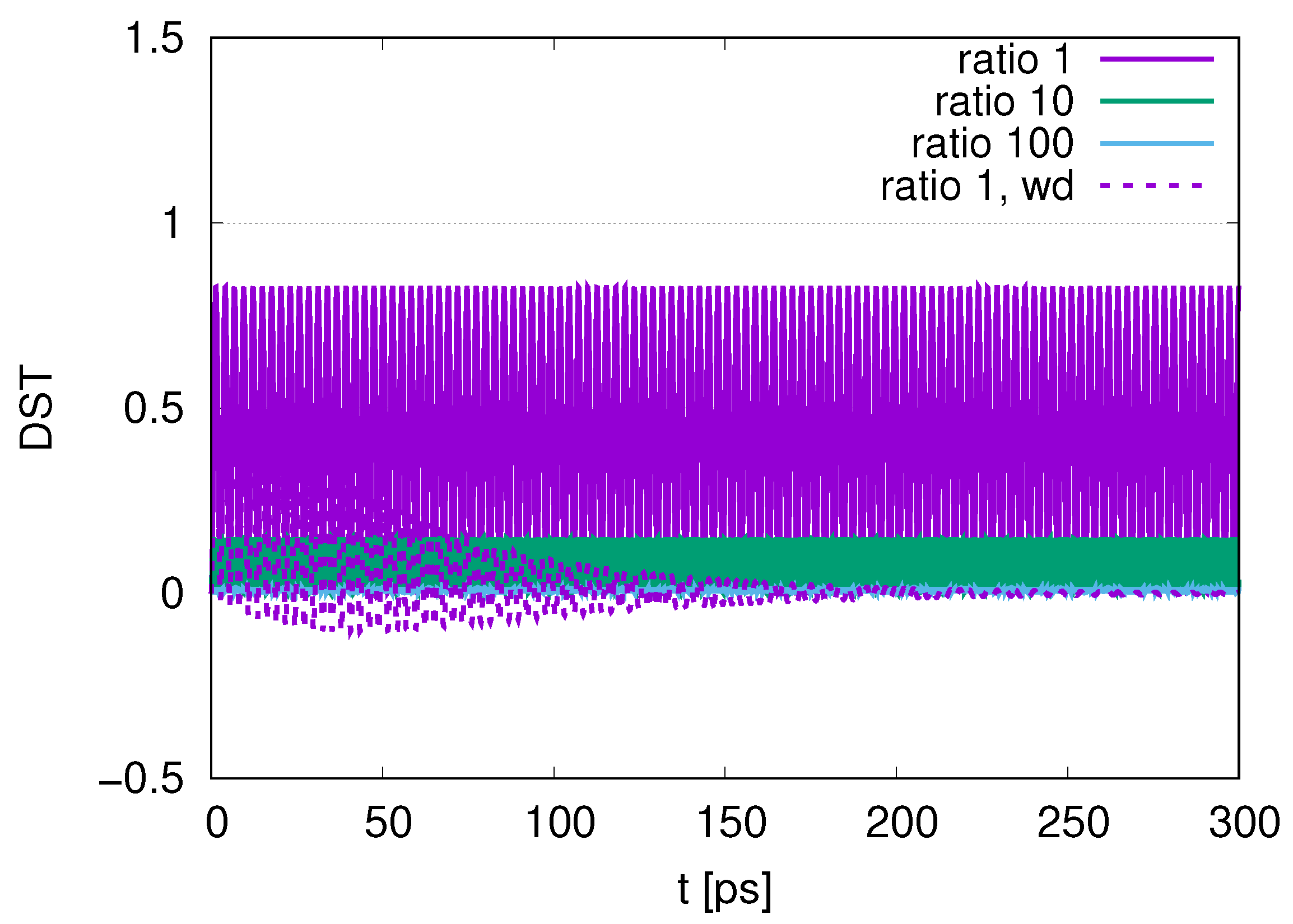
In this paper, we adopted wQED with several photon modes. As shown in
Figure 5, the contribution of photon modes is almost only the resonant mode (
) at a temperature of
. As we increase the number of DQs in
Figure 6,
Figure 12 and
Figure 17, the maximum values in the distribution of the peak of photon modes (DST) decrease in the course of time evolution. We also find that as we increase the ratio of the decay rates, the maximum values of DST decrease, with smaller leakage to the excitations of photons, which reflects the merit of adopting sub-radiance with robust states of qubits.
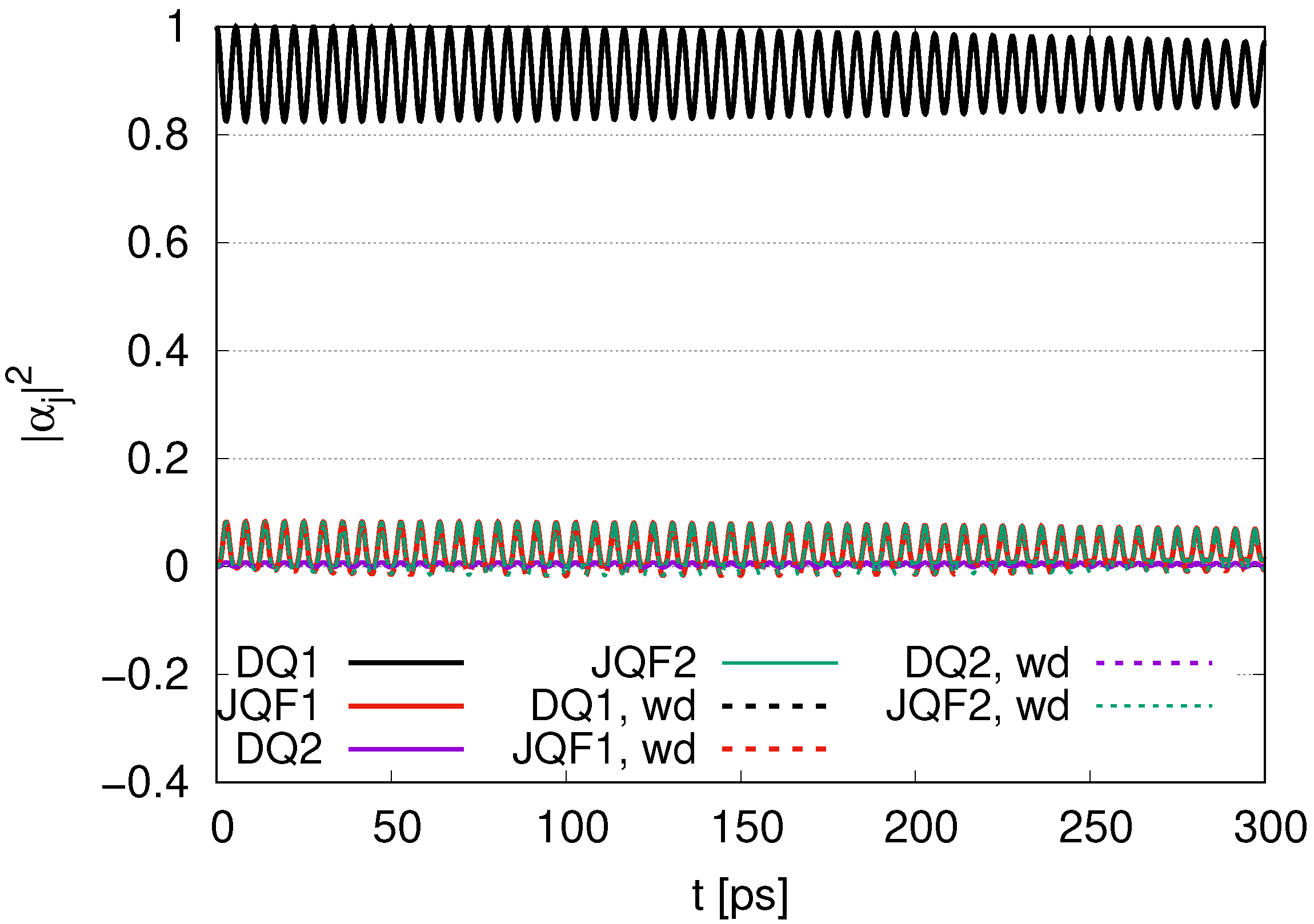
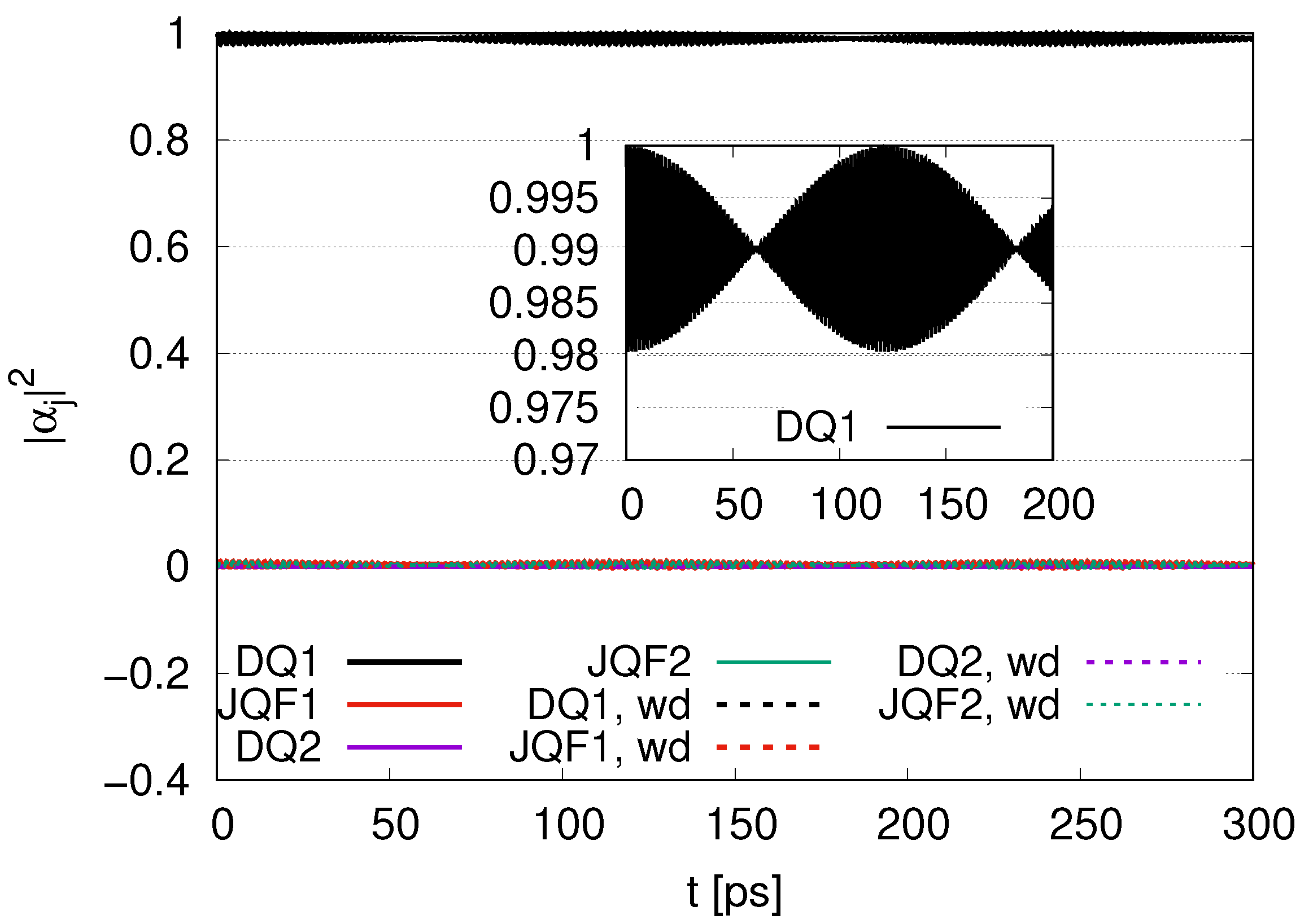
We can compare
with the ratio 100 for 1 DQ in
Figure 4, for 2 DQs in
Figure 10 and for 12 DQs in
Figure 16. As the number of DQs increases, the
for DQ1 tends to approach the initial value of 1 in the course of time evolution, with an average of 0.98 in
Figure 4, 0.99 in
Figure 10 and 0.998 in
Figure 16. In addition, the average of
for JQF1 in
Figure 4 is 0.02, that of JQF2 in
Figure 10 is 0.005 and that of JQF12 in
Figure 16 is 0.00013. The ratios of the averages of
for DQ1 and JQF are
We can conclude that sub-radiant states are realized as
for the case of 1 DQ,
for the case of 2 DQs and
for the case of 12 DQs. We now calculate the ratio of
for DQ1 and JQF. We find
for the cases of 1 DQ, 2 DQs and 12 DQs, respectively. The numerical results shown in Equation (39) correspond to those for the sub-radiant states in Equation (40).
We have described the damping processes of photon modes in Equation (29) by changing
to
. We find that the damping affects the average of
in the cases of a ratio 1 in
Figure 2 and
Figure 8. We also find convergence to the initial values for photon modes in
Figure 6 and
Figure 12. As we increase the ratio of the decay rate (
) to 10, then to 100 in
Figure 3 and
Figure 4, respectively, the effects of damping tend to disappear over time evolution. This is because
for DQ1 is nearly equal to 1, and the absolute values of the distribution of photon modes cease to increase, as shown in
Figure 6 and
Figure 12. Adopting the sub-radiant states, little changes occur due to the damping of photon modes that appear in time evolution.
We should also discuss the extension of our model to three-dimensional cases. In this paper, we considered a one-dimensional waveguide QED for the model of tryptophans in tubulin dimers. To extend this to a three-dimensional case, we extend
in Equation (
3) to
with three-dimensional momenta
for the photon modes and position
for the
jth qubit. Even if we extend our model to a thee-dimensional case, since we can estimate
for 12 DQs in two tubulin dimers, little change is expected to appear in the course of time evolution.
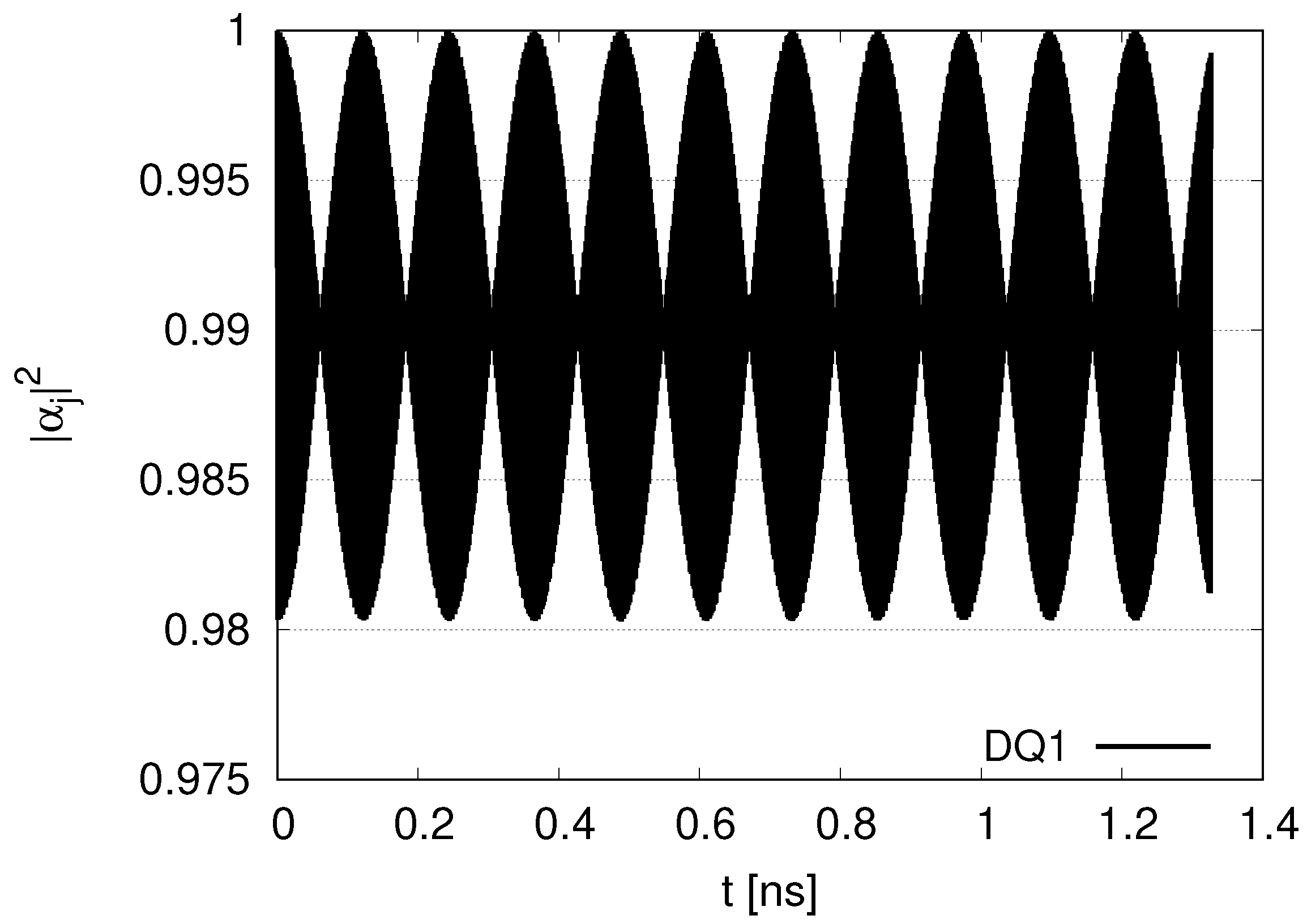
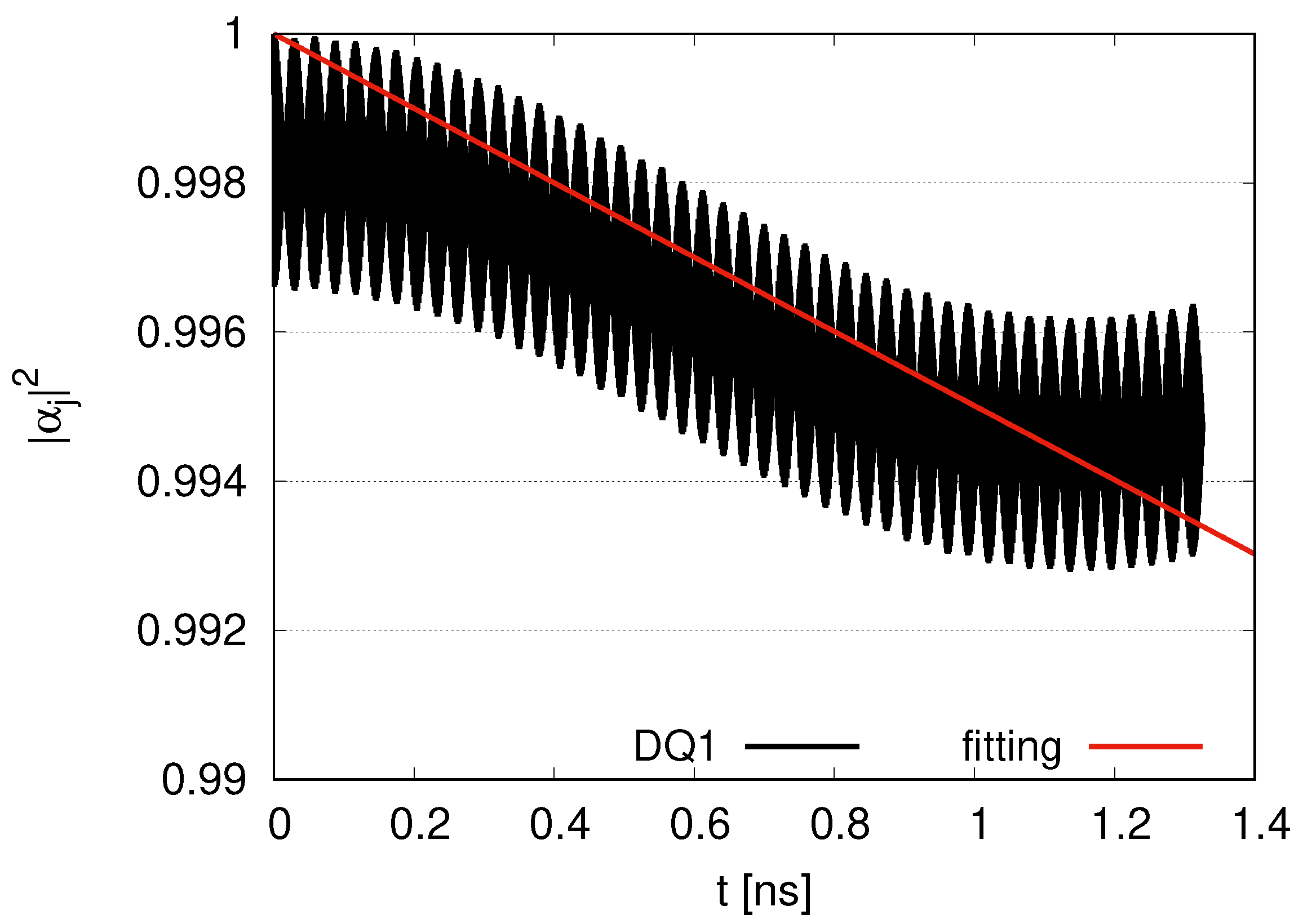
Longer simulations are shown in
Appendix A. In the case of two DQs and two JQFs in
Figure A1, little decay for
appears for DQ1 at
. On the other hand, in the case of 12 DQs and 12 JQFs in
Figure A2, we find a slight decay of fitting
with
, although the decrease seems to be moderate at
. We can consider the size of the system to discuss the decay. In the case of 2 DQs and 2 JQFs, the size of the system is
, while the size is
in the case of 12 DQs and 12 JQFs. The diffusion length of excitation energy transport in a microtubule was estimated an the experimental study as the order of
[
5]. As the size of the system in our simulations increases gradually from
, the effects of diffusion might gradually emerge. Although we encounter diffusion effects in maintaining data qubits for larger simulations, quantum entanglement might disappear for distant qubits in microtubules. Since the systems for distant qubits behave independently, the dynamics of the DQs and JQFs might be described by near qubits, as in the case of two DQs and two JQFs. Our experimental study suggests that the fluorescence lifetime of tryptophan is a few ns. Our simulation reported in this paper describes nanosecond scales comparable with the fluorescence lifetime. If we assume a larger decay rate (
) for JQFs representing water molecules (with a ratio larger than 100), the robustness of data qubits is reinforced. A balance of diffusion and robustness of sub-radiant states is significant.
Maintaining quantum coherence at high temperatures is a demanding task in biological systems. Our strategy is to regard biological systems, especially brains, as open systems. Life is a flow of energy and matter. To maintain biological systems, we prepared three systems, namely an energy supply, physical system and heat bath. We provided continuous energy flow from the energy supply, through the physical system to the heat bath for dissipation of energy. The Fröhlich condensate represents an example suggested in the flow of these three systems, with Bose–Einstein condensate proposed in biological systems [
40,
41,
42]. Self-organization and error corrections due to energy supply represent key concepts for life. We investigated the brain as an open system achieving biological order. In particular, microtubules have an energy source supplied by surrounding mitochondria and heat baths in cells [
43,
44]. In the case of error corrections by the energy supply to overcome decoherence, quantum coherence might be maintained.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}