
Steviol Represses Glucose Metabolism and Translation Initiation in Pancreatic Cancer Cells




 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Antibodies
2.3. Proliferation Assay
2.4. Western Blotting
2.5. Clonogenic Assay
2.6. Real-Time xCELLigence Assays
2.7. Migration Assay
2.8. Invasion Assay
2.9. Cell Cycle Studies
2.10. Lactate Assay/ Glucose Assay
2.11. Statistical Analysis
3. Results
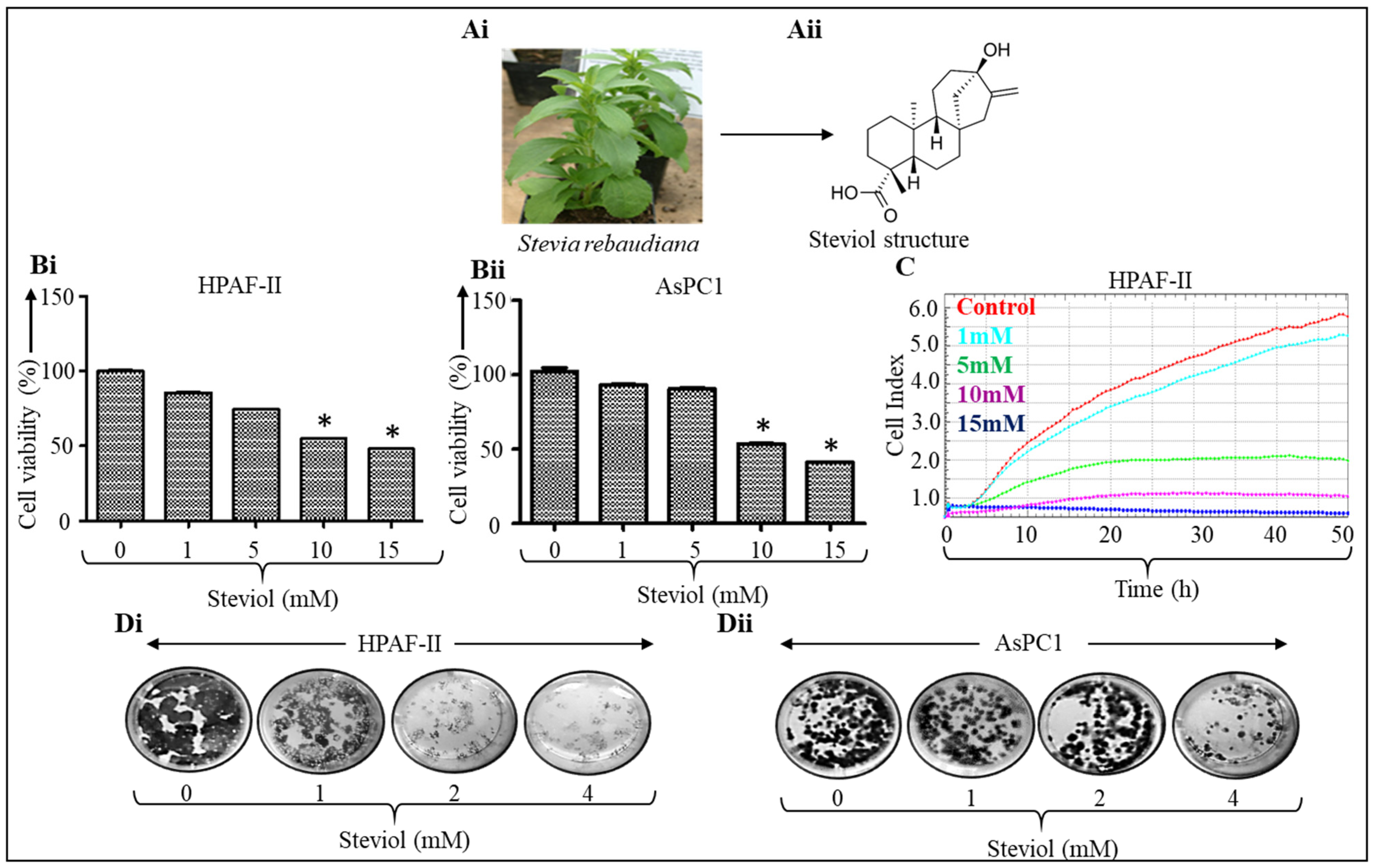
3.1. Growth Inhibitory Effects of Steviol in Pancreatic Cancer Cells
3.2. Steviol Suppresses Migration and Invasion Capabilities of Pancreatic Cancer Cells
3.3. Steviol Regulates Cell Cycle Machinery
3.4. Steviol Induces Apoptosis in Pancreatic Cancer Cells
3.5. Steviol Suppresses Translation Initiation in Pancreatic Cancer Cells
3.6. Steviol Decreases Glucose Uptake and Lactate Secretion in Pancreatic Cancer Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Tao, J.; Yang, G.; Zhou, W.; Qiu, J.; Chen, G.; Luo, W.; Zhao, F.; You, L.; Zheng, L.; Zhang, T.; et al. Targeting hypoxic tumor microenvironment in pancreatic cancer. J. Hematol. Oncol. 2021, 14, 14. [Google Scholar] [CrossRef]
- Mohd-Radzman, N.H.; Ismail, W.I.; Jaapar, S.S.; Adam, Z.; Adam, A. Stevioside from Stevia rebaudiana Bertoni Increases Insulin Sensitivity in 3T3-L1 Adipocytes. Evid.-Based Complement. Altern. Med. Ecam 2013, 2013, 938081. [Google Scholar] [CrossRef] [Green Version]
- Boonkaewwan, C.; Burodom, A. Anti-inflammatory and immunomodulatory activities of stevioside and steviol on colonic epithelial cells. J. Sci. Food Agric. 2013, 93, 3820–3825. [Google Scholar] [CrossRef]
- Chatsudthipong, V.; Muanprasat, C. Stevioside and related compounds: Therapeutic benefits beyond sweetness. Pharmacol. Ther. 2009, 121, 41–54. [Google Scholar] [CrossRef]
- Wu, Y.; Rodenburg, D.L.; Ibrahim, M.A.; McChesney, J.D.; Avery, M.A. Stevioside methanol tetra-solvate. Acta Crystallogr. Sect. E Struct. Rep. Online 2013, 69, o410–o411. [Google Scholar] [CrossRef]
- López, V.; Pérez, S.; Vinuesa, A.; Zorzetto, C.; Abian, O. Stevia rebaudiana ethanolic extract exerts better antioxidant properties and antiproliferative effects in tumour cells than its diterpene glycoside stevioside. Food Funct. 2016, 7, 2107–2113. [Google Scholar] [CrossRef]
- Geuns, J.M.; Augustijns, P.; Mols, R.; Buyse, J.G.; Driessen, B. Metabolism of stevioside in pigs and intestinal absorption characteristics of stevioside, rebaudioside A and steviol. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2003, 41, 1599–1607. [Google Scholar] [CrossRef]
- Ray, J.; Kumar, S.; Laor, D.; Shereen, N.; Nwamaghinna, F.; Thomson, A.; Perez Perez, J.; Soni, L.; McFarlane, S.I. Effects of Stevia Rebaudiana on Glucose Homeostasis, Blood Pressure and Inflammation: A Critical Review of Past and Current Research Evidence. Int. J. Clin. Res. Trials 2020, 5, 142. [Google Scholar] [CrossRef]
- Takasaki, M.; Konoshima, T.; Kozuka, M.; Tokuda, H.; Takayasu, J.; Nishino, H.; Miyakoshi, M.; Mizutani, K.; Lee, K.H. Cancer preventive agents. Part 8: Chemopreventive effects of stevioside and related compounds. Bioorganic Med. Chem. 2009, 17, 600–605. [Google Scholar] [CrossRef]
- Gregersen, S.; Jeppesen, P.B.; Holst, J.J.; Hermansen, K. Antihyperglycemic effects of stevioside in type 2 diabetic subjects. Metab. Clin. Exp. 2004, 53, 73–76. [Google Scholar] [CrossRef]
- Jeppesen, P.B.; Gregersen, S.; Rolfsen, S.E.; Jepsen, M.; Colombo, M.; Agger, A.; Xiao, J.; Kruhoffer, M.; Orntoft, T.; Hermansen, K. Antihyperglycemic and blood pressure-reducing effects of stevioside in the diabetic Goto-Kakizaki rat. Metab. Clin. Exp. 2003, 52, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Boonkaewwan, C.; Ao, M.; Toskulkao, C.; Rao, M.C. Specific immunomodulatory and secretory activities of stevioside and steviol in intestinal cells. J. Agric. Food Chem. 2008, 56, 3777–3784. [Google Scholar] [CrossRef]
- Gupta, E.; Kaushik, S.; Purwar, S.; Sharma, R.; Balapure, A.K.; Sundaram, S. Anticancer Potential of Steviol in MCF-7 Human Breast Cancer Cells. Pharmacog. Mag. 2017, 13, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Mattison, J.A.; Roth, G.S.; Beasley, T.M.; Tilmont, E.M.; Handy, A.M.; Herbert, R.L.; Longo, D.L.; Allison, D.B.; Young, J.E.; Bryant, M.; et al. Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nature 2012, 489, 318–321. [Google Scholar] [CrossRef] [Green Version]
- Slattery, M.L.; Benson, J.; Berry, T.D.; Duncan, D.; Edwards, S.L.; Caan, B.J.; Potter, J.D. Dietary sugar and colon cancer. Cancer Epidemiol. Biomark. Prev. A Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 1997, 6, 677–685. [Google Scholar]
- Fiechter, A.; Gmunder, F.K. Metabolic control of glucose degradation in yeast and tumor cells. Adv. Biochem. Eng./Biotechnol. 1989, 39, 1–28. [Google Scholar]
- Yan, L.; Raj, P.; Yao, W.; Ying, H. Glucose Metabolism in Pancreatic Cancer. Cancers 2019, 11, 1460. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Khan, S.; Sekhri, R.; Mandil, H.; Behrman, S.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. Protein kinase D1 regulates metabolic switch in pancreatic cancer via modulation of mTORC1. Br. J. Cancer 2020, 122, 121–131. [Google Scholar] [CrossRef]
- Janapala, Y.; Preiss, T.; Shirokikh, N.E. Control of Translation at the Initiation Phase During Glucose Starvation in Yeast. Int. J. Mol. Sci. 2019, 20, 4043. [Google Scholar] [CrossRef] [Green Version]
- Karaki, S.; Andrieu, C.; Ziouziou, H.; Rocchi, P. Chapter One—The Eukaryotic Translation Initiation Factor 4E (eIF4E) as a Therapeutic Target for Cancer. In Adv Protein Chem Struct Biol; Donev, R., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 101, pp. 1–26. [Google Scholar]
- Jiang, S.-L.; Mo, J.-L.; Peng, J.; Lei, L.; Yin, J.-Y.; Zhou, H.-H.; Liu, Z.-Q.; Hong, W.-X. Targeting translation regulators improves cancer therapy. Genomics 2021, 113, 1247–1256. [Google Scholar] [CrossRef]
- Lambert, A.; Schwarz, L.; Borbath, I.; Henry, A.; Van Laethem, J.L.; Malka, D.; Ducreux, M.; Conroy, T. An update on treatment options for pancreatic adenocarcinoma. Adv. Med. Oncol. 2019, 11, 1758835919875568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Liang, Y.; Xing, C.; Wang, H.; Hu, P.; Li, J.; Huang, H.; Wang, W.; Jiang, C. Cancer-associated adipocytes exhibit distinct phenotypes and facilitate tumor progression in pancreatic cancer. Oncol. Rep. 2019, 42, 2537–2549. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.L.; Rehman, A.A.; Gondi, C.S. Therapeutic options for the management of pancreatic cancer. World J. Gastroenterol. 2014, 20, 11142–11159. [Google Scholar] [CrossRef]
- Yue, Q.; Gao, G.; Zou, G.; Yu, H.; Zheng, X. Natural Products as Adjunctive Treatment for Pancreatic Cancer: Recent Trends and Advancements. BioMed Res. Int. 2017, 2017, 8412508. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Khan, S.; Gupta, S.C.; Kashyap, V.K.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. MUC13 contributes to rewiring of glucose metabolism in pancreatic cancer. Oncogenesis 2018, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Sikander, M.; Malik, S.; Chauhan, N.; Khan, P.; Kumari, S.; Kashyap, V.K.; Khan, S.; Ganju, A.; Halaweish, F.T.; Yallapu, M.M.; et al. Cucurbitacin D Reprograms Glucose Metabolic Network in Prostate Cancer. Cancers 2019, 11, 364. [Google Scholar] [CrossRef] [Green Version]
- Sikander, M.; Hafeez, B.B.; Malik, S.; Alsayari, A.; Halaweish, F.T.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. Cucurbitacin D exhibits potent anti-cancer activity in cervical cancer. Sci. Rep. 2016, 6, 36594. [Google Scholar] [CrossRef]
- Hafeez, B.B.; Ganju, A.; Sikander, M.; Kashyap, V.K.; Hafeez, Z.B.; Chauhan, N.; Malik, S.; Massey, A.E.; Tripathi, M.K.; Halaweish, F.T.; et al. Ormeloxifene Suppresses Prostate Tumor Growth and Metastatic Phenotypes via Inhibition of Oncogenic β-catenin Signaling and EMT Progression. Mol. Cancer 2017, 16, 2267–2280. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Ebeling, M.C.; Chauhan, N.; Thompson, P.A.; Gara, R.K.; Ganju, A.; Yallapu, M.M.; Behrman, S.W.; Zhao, H.; Zafar, N.; et al. Ormeloxifene suppresses desmoplasia and enhances sensitivity of gemcitabine in pancreatic cancer. Cancer Res. 2015, 75, 2292–2304. [Google Scholar] [CrossRef] [Green Version]
- Zaman, M.S.; Chauhan, N.; Yallapu, M.M.; Gara, R.K.; Maher, D.M.; Kumari, S.; Sikander, M.; Khan, S.; Zafar, N.; Jaggi, M.; et al. Curcumin Nanoformulation for Cervical Cancer Treatment. Sci. Rep. 2016, 6, 20051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Tong, X.; Ye, X. Cyclin D1 promotes cell cycle progression through enhancing NDR1/2 kinase activity independent of cyclin-dependent kinase 4. J. Biol. Chem. 2013, 288, 26678–26687. [Google Scholar] [CrossRef] [Green Version]
- Kaczanowski, S. Apoptosis: Its origin, history, maintenance and the medical implications for cancer and aging. Phys. Biol. 2016, 13, 031001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.; Watari, H.; AbuAlmaaty, A.; Ohba, Y.; Sakuragi, N. Apoptosis and molecular targeting therapy in cancer. BioMed Res. Int. 2014, 2014, 150845. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.U.; Ur Rahman, M.S.; Jia, Z.; Jiang, C. Eukaryotic translation initiation factors and cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2017, 39, 1010428317709805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingras, A.C.; Raught, B.; Sonenberg, N. eIF4 initiation factors: Effectors of mRNA recruitment to ribosomes and regulators of translation. Annu. Rev. Biochem. 1999, 68, 913–963. [Google Scholar] [CrossRef]
- Levine, A.J.; Puzio-Kuter, A.M. The control of the metabolic switch in cancers by oncogenes and tumor suppressor genes. Science 2010, 330, 1340–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Blum, R.; Kloog, Y. Metabolism addiction in pancreatic cancer. Cell Death Dis. 2014, 5, e1065. [Google Scholar] [CrossRef] [Green Version]
- Burris, H.A., III; Moore, M.J.; Andersen, J.; Green, M.R.; Rothenberg, M.L.; Modiano, M.R.; Cripps, M.C.; Portenoy, R.K.; Storniolo, A.M.; Tarassoff, P.; et al. Improvements in survival and clinical benefit with gemcitabine as first-line therapy for patients with advanced pancreas cancer: A randomized trial. J. Clin. Oncol. 1997, 15, 2403–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.N.; Summy, J.M.; Zhang, J.; Park, S.I.; Parikh, N.U.; Gallick, G.E. Development and characterization of gemcitabine-resistant pancreatic tumor cells. Ann. Surg. Oncol. 2007, 14, 3629–3637. [Google Scholar] [CrossRef] [PubMed]
- Sikander, M.; Malik, S.; Khan, S.; Kumari, S.; Chauhan, N.; Khan, P.; Halaweish, F.T.; Chauhan, B.; Yallapu, M.M.; Jaggi, M.; et al. Novel Mechanistic Insight into the Anticancer Activity of Cucurbitacin D against Pancreatic Cancer (Cuc D Attenuates Pancreatic Cancer). Cells 2019, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoxhaj, G.; Manning, B.D. The PI3K-AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Spilka, R.; Ernst, C.; Mehta, A.K.; Haybaeck, J. Eukaryotic translation initiation factors in cancer development and progression. Cancer Lett. 2013, 340, 9–21. [Google Scholar] [CrossRef]
- D’Abronzo, L.S.; Ghosh, P.M. eIF4E Phosphorylation in Prostate Cancer. Neoplasia (N. Y.) 2018, 20, 563–573. [Google Scholar] [CrossRef]
- Panagiotou, C.; Mihailidou, C.; Brauhli, G.; Katsarou, O.; Moutsatsou, P. Effect of steviol, steviol glycosides and stevia extract on glucocorticoid receptor signaling in normal and cancer blood cells. Mol. Cell. Endocrinol. 2018, 460, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Bundgaard Anker, C.C.; Rafiq, S.; Jeppesen, P.B. Effect of Steviol Glycosides on Human Health with Emphasis on Type 2 Diabetic Biomarkers: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2019, 11, 1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Xia, Y.; Sui, X.; Peng, Q.; Zhang, T.; Li, J.; Zhang, J. Steviol, a natural product inhibits proliferation of the gastrointestinal cancer cells intensively. Oncotarget 2018, 9, 26299–26308. [Google Scholar] [CrossRef]
- Clemens, M.J. Targets and mechanisms for the regulation of translation in malignant transformation. Oncogene 2004, 23, 3180–3188. [Google Scholar] [CrossRef] [Green Version]
- Mamane, Y.; Petroulakis, E.; Rong, L.; Yoshida, K.; Ler, L.W.; Sonenberg, N. eIF4E--from translation to transformation. Oncogene 2004, 23, 3172–3179. [Google Scholar] [CrossRef] [Green Version]
- De Benedetti, A.; Harris, A.L. eIF4E expression in tumors: Its possible role in progression of malignancies. Int. J. Biochem. Cell Biol. 1999, 31, 59–72. [Google Scholar] [CrossRef]
- Graff, J.R.; Boghaert, E.R.; De Benedetti, A.; Tudor, D.L.; Zimmer, C.C.; Chan, S.K.; Zimmer, S.G. Reduction of translation initiation factor 4E decreases the malignancy of ras-transformed cloned rat embryo fibroblasts. Int. J. Cancer 1995, 60, 255–263. [Google Scholar] [CrossRef]
- Cohen, N.; Sharma, M.; Kentsis, A.; Perez, J.M.; Strudwick, S.; Borden, K.L. PML RING suppresses oncogenic transformation by reducing the affinity of eIF4E for mRNA. EMBO J. 2001, 20, 4547–4559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, J.; Graff, J.; Ruggero, D.; Sonenberg, N. Targeting the eIF4F translation initiation complex: A critical nexus for cancer development. Cancer Res. 2015, 75, 250–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yu, T.; Cao, J.; Liu, L.; Liu, Y.; Kong, H.W.; Zhu, M.X.; Lin, H.C.; Chu, D.D.; Yao, M.; et al. MicroRNA-148a Suppresses Invasion and Metastasis of Human Non-Small-Cell Lung Cancer. Cell Physiol. Biochem. 2015, 37, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Konicek, B.W.; Dumstorf, C.A.; Graff, J.R. Targeting the eIF4F translation initiation complex for cancer therapy. Cell Cycle 2008, 7, 2466–2471. [Google Scholar] [CrossRef] [Green Version]
- Lazaris-Karatzas, A.; Montine, K.S.; Sonenberg, N. Malignant transformation by a eukaryotic initiation factor subunit that binds to mRNA 5’ cap. Nature 1990, 345, 544–547. [Google Scholar] [CrossRef]
- Ruggero, D.; Montanaro, L.; Ma, L.; Xu, W.; Londei, P.; Cordon-Cardo, C.; Pandolfi, P.P. The translation factor eIF-4E promotes tumor formation and cooperates with c-Myc in lymphomagenesis. Nat. Med. 2004, 10, 484–486. [Google Scholar] [CrossRef]
- Chen, J.M.; Ding, L.; Sui, X.C.; Xia, Y.M.; Wan, H.D.; Lu, T. Production of a bioactive sweetener steviolbioside via specific hydrolyzing ester linkage of stevioside with a β-galactosidase. Food Chem. 2016, 196, 155–160. [Google Scholar] [CrossRef]
- Pasqualli, T.; Chaves, P.E.E.; Pereira, C.; Serpa, É.A.; Oliveira, L.F.S.; Machado, M.M. Steviol, the active principle of the stevia sweetener, causes a reduction of the cells of the immunological system even consumed in low concentrations. Immunopharmacol. Immunotoxicol. 2020, 42, 504–508. [Google Scholar] [CrossRef]
- Dooley, J.; Lagou, V.; Dresselaers, T.; van Dongen, K.A.; Himmelreich, U.; Liston, A. No Effect of Dietary Aspartame or Stevia on Pancreatic Acinar Carcinoma Development, Growth, or Induced Mortality in a Murine Model. Front. Oncol. 2017, 7, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumari, S.; Sikander, M.; Malik, S.; Tripathi, M.K.; Hafeez, B.B.; Yallapu, M.M.; Chauhan, S.C.; Khan, S.; Jaggi, M. Steviol Represses Glucose Metabolism and Translation Initiation in Pancreatic Cancer Cells. Biomedicines 2021, 9, 1814. https://doi.org/10.3390/biomedicines9121814
Kumari S, Sikander M, Malik S, Tripathi MK, Hafeez BB, Yallapu MM, Chauhan SC, Khan S, Jaggi M. Steviol Represses Glucose Metabolism and Translation Initiation in Pancreatic Cancer Cells. Biomedicines. 2021; 9(12):1814. https://doi.org/10.3390/biomedicines9121814
Chicago/Turabian StyleKumari, Sonam, Mohammed Sikander, Shabnam Malik, Manish K. Tripathi, Bilal B. Hafeez, Murali M. Yallapu, Subhash C. Chauhan, Sheema Khan, and Meena Jaggi. 2021. "Steviol Represses Glucose Metabolism and Translation Initiation in Pancreatic Cancer Cells" Biomedicines 9, no. 12: 1814. https://doi.org/10.3390/biomedicines9121814
APA StyleKumari, S., Sikander, M., Malik, S., Tripathi, M. K., Hafeez, B. B., Yallapu, M. M., Chauhan, S. C., Khan, S., & Jaggi, M. (2021). Steviol Represses Glucose Metabolism and Translation Initiation in Pancreatic Cancer Cells. Biomedicines, 9(12), 1814. https://doi.org/10.3390/biomedicines9121814