
The NMD Pathway Regulates GABARAPL1 mRNA during the EMT


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies, Vectors, Chemicals and siRNAs
2.2. Cell Culture and Transfection
2.3. Western Blotting
2.4. RNA Extraction and RT-qPCR
2.5. RNA-FISH
2.6. Immunohistochemistry and Immunostaining Assessment
2.7. RNA-Immunoprecipitation (RIP)
2.8. Statistical Analysis
3. Results
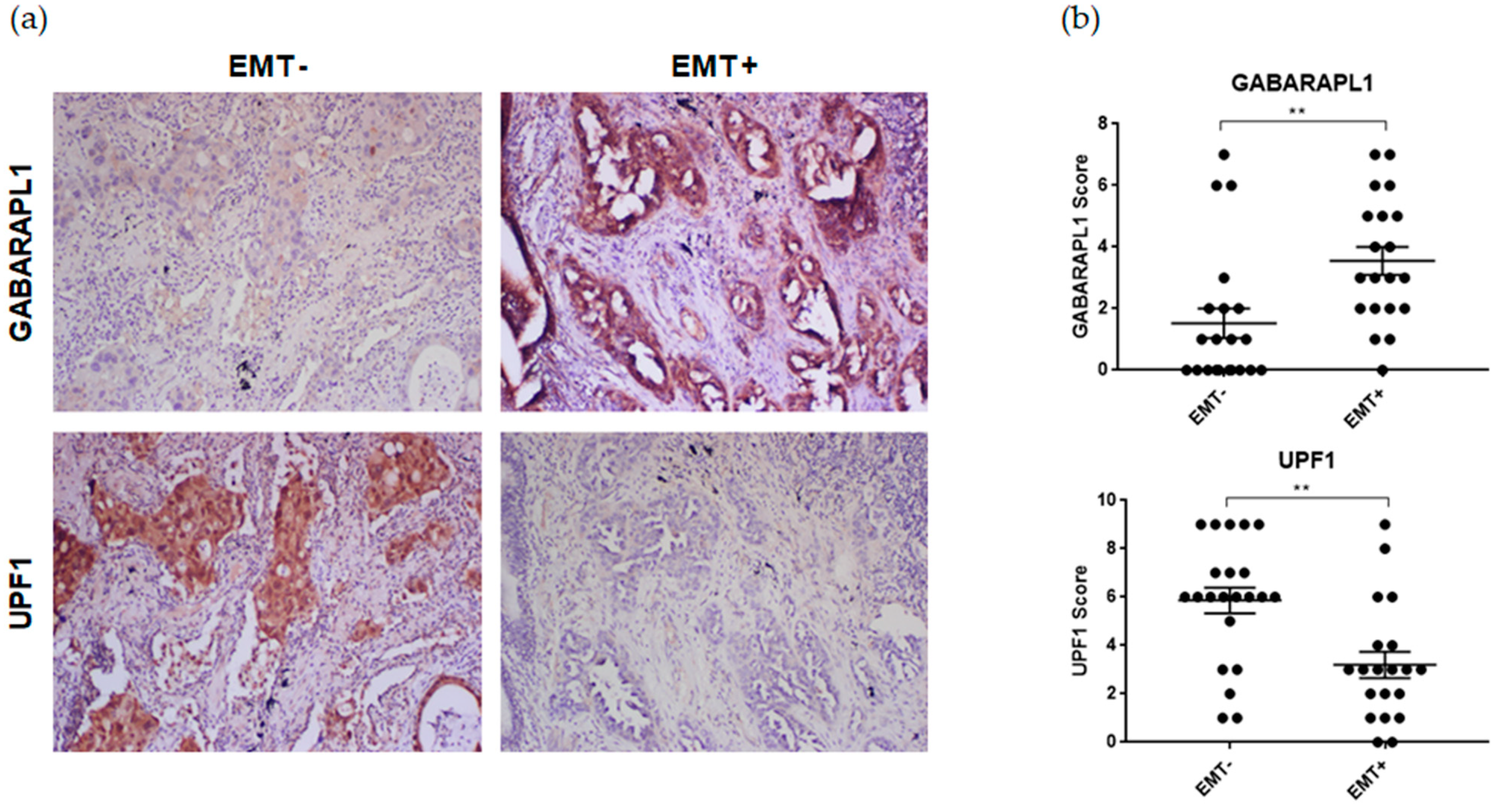
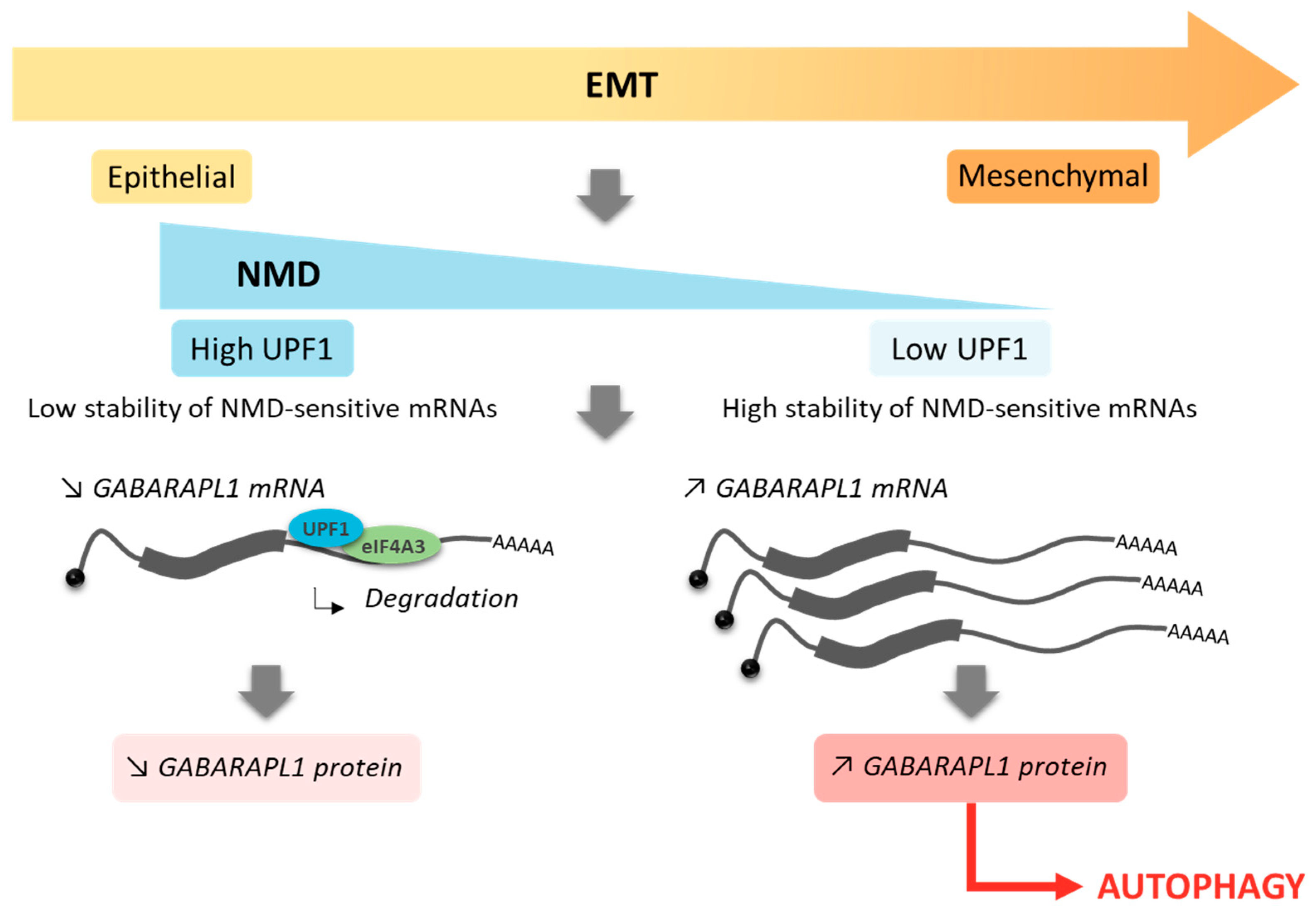
3.1. The EMT was Associated with A Loss of UPF1 Expression as well as GABARAPL1 Overexpression
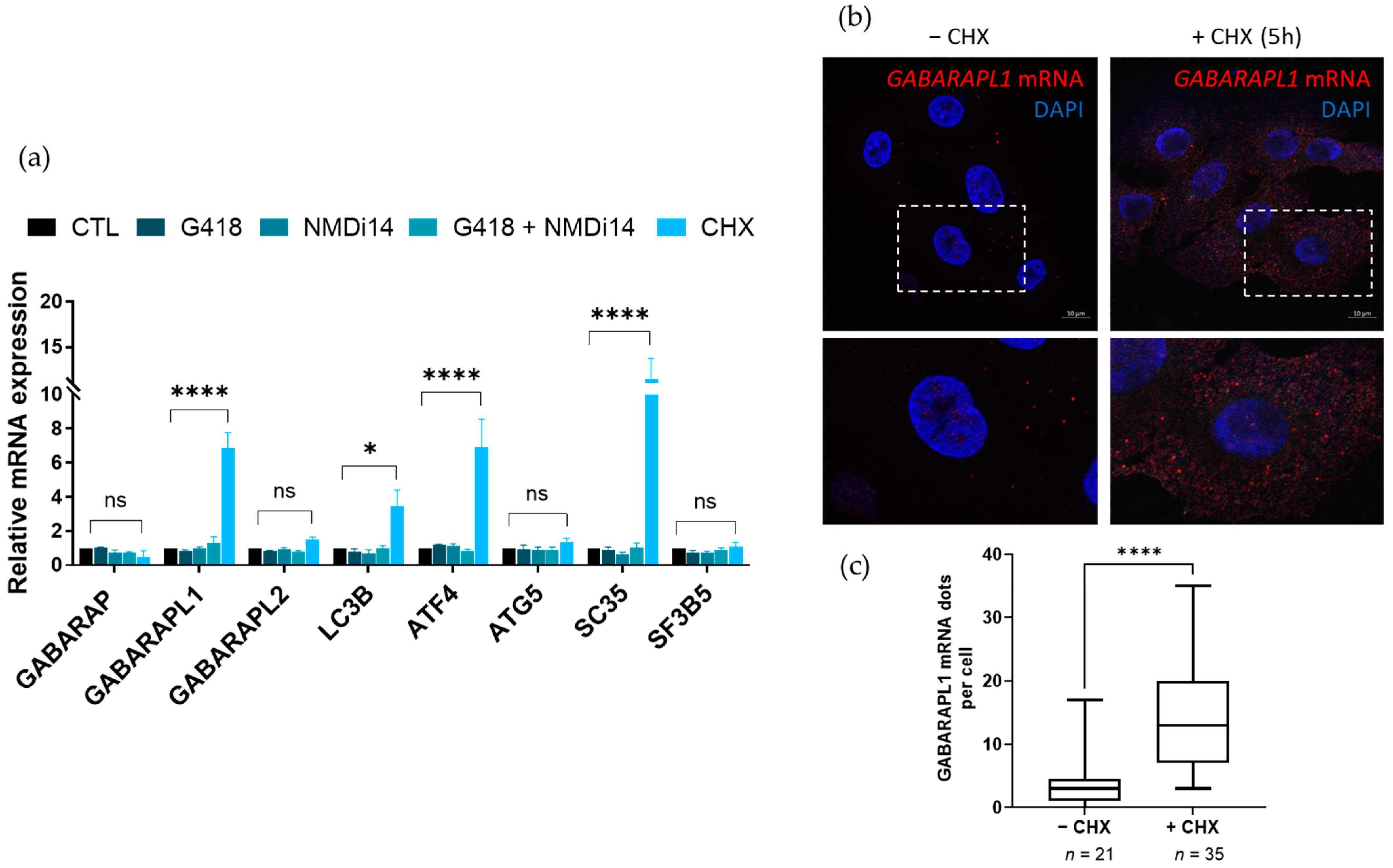
3.2. GABARAPL1 Transcript Increased upon NMD Inhibition
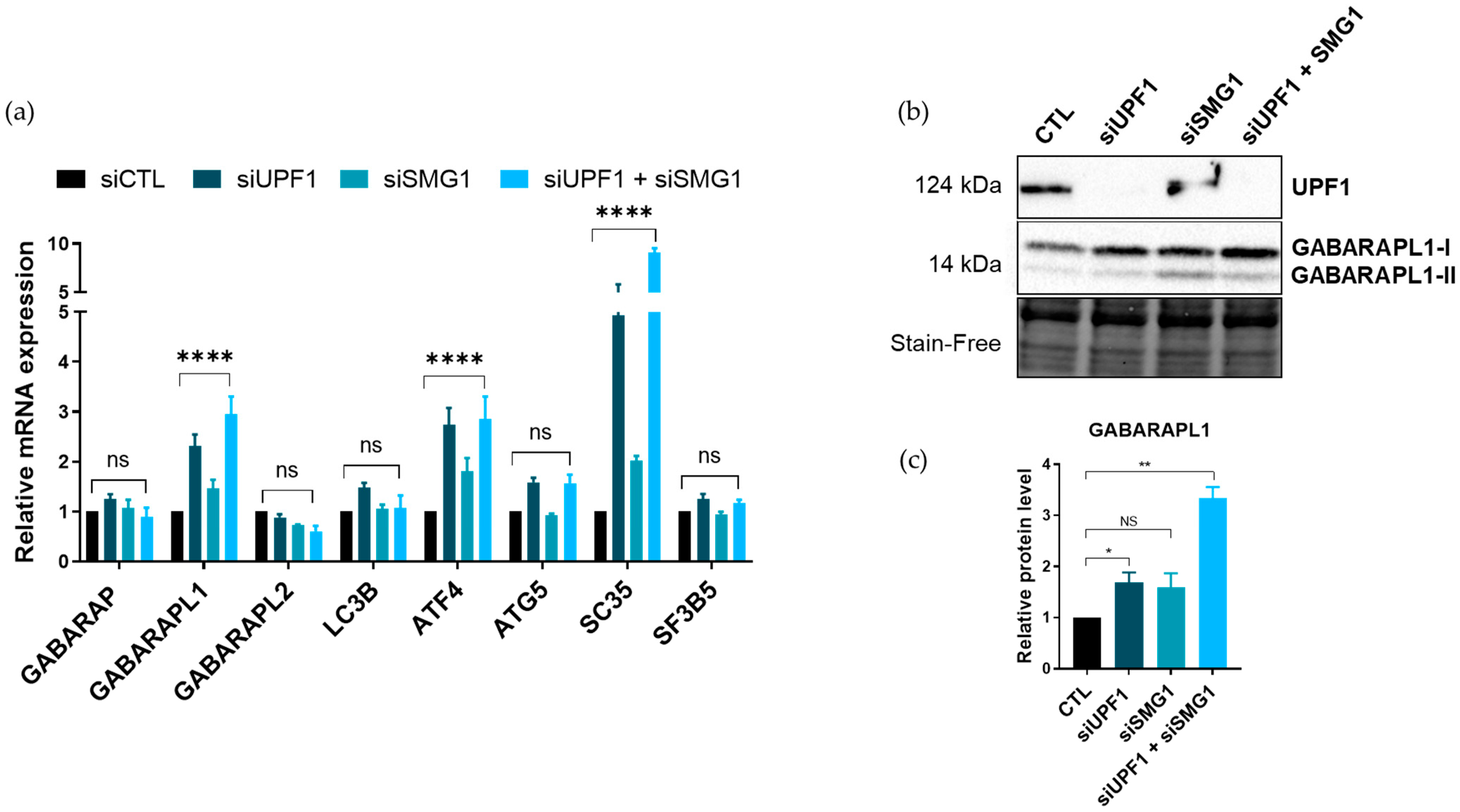
3.3. UPF1 Was Associated with GABARAPL1 mRNAs
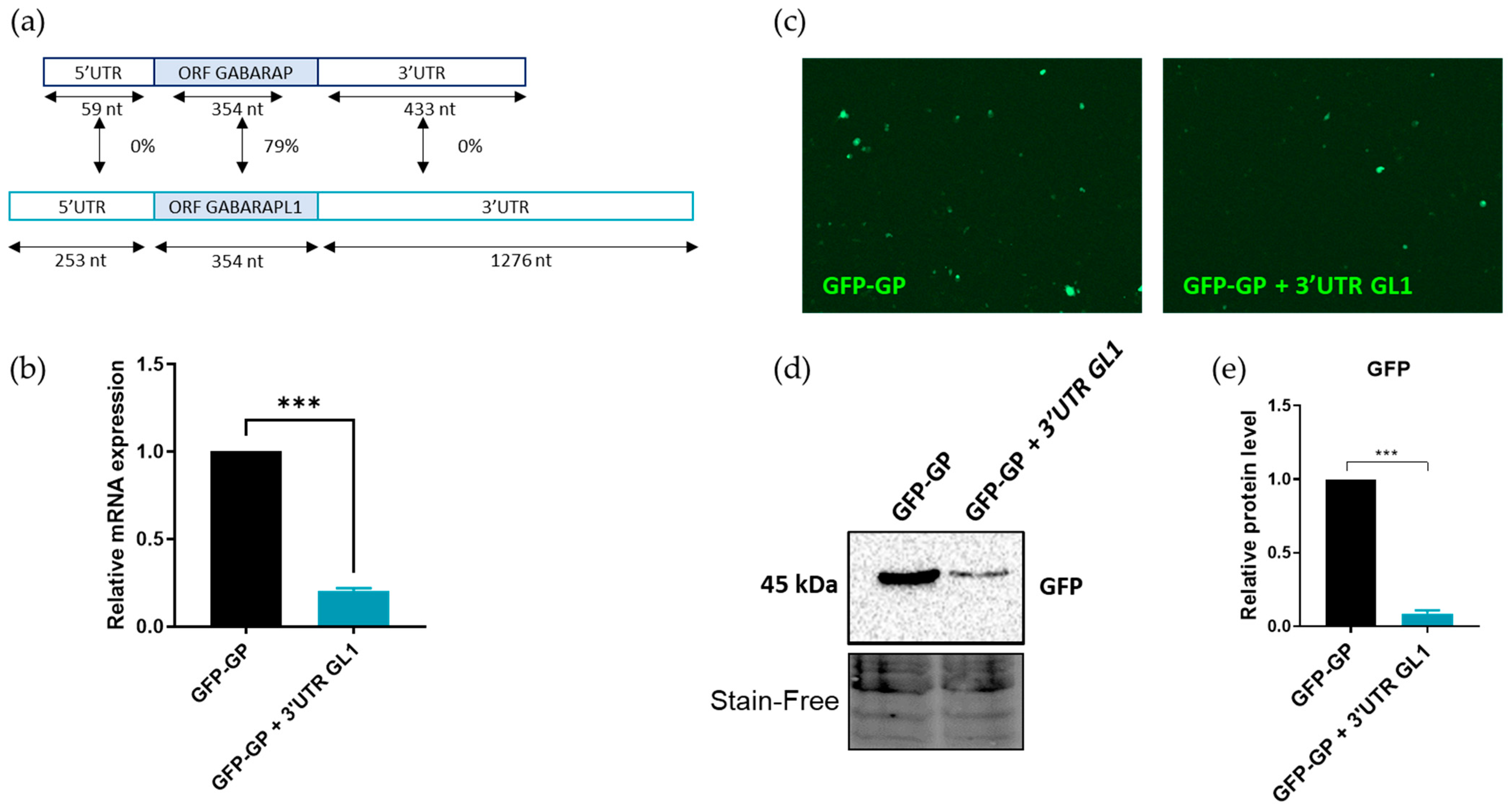
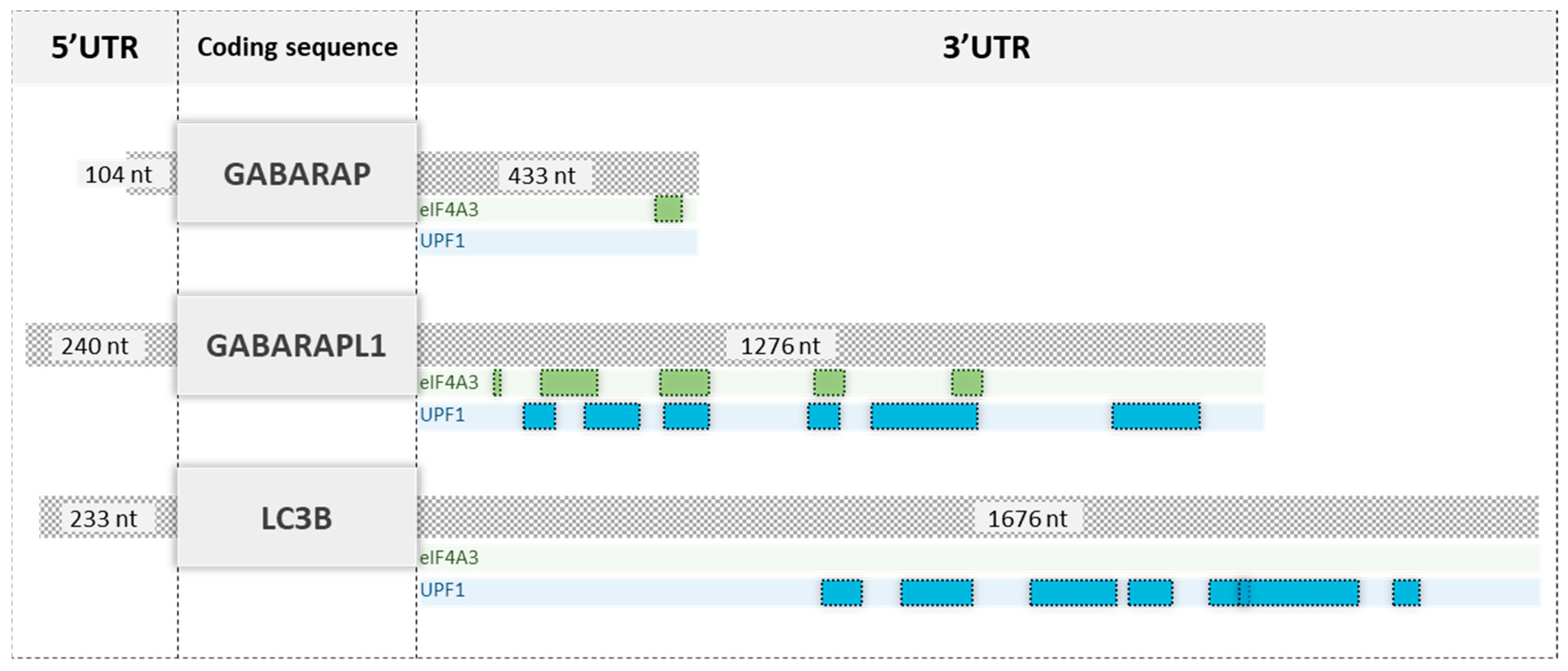
3.4. The 3′UTR of GABARAPL1 mRNAs Contributed to Its Degradation by The NMD
3.5. eIF4A3 Regulated GABARAPL1 mRNA Degradation via Its 3′UTR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachat, C.; Peixoto, P.; Hervouet, E. Epithelial to Mesenchymal Transition History: Embryonic Development Cancers. Biomolecules 2021, 11, 782. [Google Scholar] [CrossRef]
- Nemos, C.; Mansuy, V.; Vernier-Magnin, S.; Fraichard, A.; Jouvenot, M.; Delage-Mourroux, R. Expression of gec1/GABARAPL1 versus GABARAP mRNAs in human: Predominance of gec1/GABARAPL1 in the central nervous system. Mol. Brain Res. 2003, 119, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Rouschop, K.M.A.; van den Beucken, T.; Dubois, L.; Niessen, H.; Bussink, J.; Savelkouls, K.; Keulers, T.; Mujcic, H.; Landuyt, W.; Voncken, J.W.; et al. The unfolded protein response protects human tumor cells during hypoxia through regulation of the autophagy genes MAP1LC3B and ATG5. J. Clin. Investig. 2010, 120, 127–141. [Google Scholar] [CrossRef]
- B’chir, W.; Maurin, A.-C.; Carraro, V.; Averous, J.; Jousse, C.; Muranishi, Y.; Parry, L.; Stepien, G.; Fafournoux, P.; Bruhat, A. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res. 2013, 41, 7683–7699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinniburgh, A.J.; Maquat, L.E.; Schedl, T.; Rachmilewitz, E.; Ross, J. mRNA-deficient beta o-thalassemia results from a single nucleotide deletion. Nucleic Acids Res. 1982, 10, 5421–5427. [Google Scholar] [CrossRef]
- Maquat, L.E.; Kinniburgh, A.J.; Rachmilewitz, E.A.; Ross, J. Unstable β-globin mRNA in mRNA-deficient β0 thalassemia. Cell 1981, 27, 543–553. [Google Scholar] [CrossRef]
- Kim, Y.K.; Maquat, L.E. UPFront and center in RNA decay: UPF1 in nonsense-mediated mRNA decay and beyond. RNA 2019, 25, 407–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Hir, H.; Izaurralde, E.; Maquat, L.E.; Moore, M.J. The spliceosome deposits multiple proteins 20–24 nucleotides upstream of mRNA exon-exon junctions. EMBO J. 2000, 19, 6860–6869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Hir, H.; Moore, M.J.; Maquat, L.E. Pre-mRNA splicing alters mRNP composition: Evidence for stable association of proteins at exon–exon junctions. Genes Dev. 2000, 14, 1098–1108. [Google Scholar]
- Saulière, J.; Murigneux, V.; Wang, Z.; Marquenet, E.; Barbosa, I.; Le Tonquèze, O.; Audic, Y.; Paillard, L.; Crollius, H.R.; Le Hir, H. CLIP-seq of eIF4AIII reveals transcriptome-wide mapping of the human exon junction complex. Nat. Struct. Mol. Biol. 2012, 19, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Le Hir, H.; Andersen, G.R. Structural insights into the exon junction complex. Curr. Opin. Struct. Biol. 2008, 18, 112–119. [Google Scholar] [CrossRef]
- Le Hir, H. The exon-exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-mediated mRNA decay. EMBO J. 2001, 20, 4987–4997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tange, T.Ø.; Nott, A.; Moore, M.J. The ever-increasing complexities of the exon junction complex. Curr. Opin. Cell Biol. 2004, 16, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Metze, S.; Herzog, V.A.; Ruepp, M.-D.; Muhlemann, O. Comparison of EJC-enhanced and EJC-independent NMD in human cells reveals two partially redundant degradation pathways. RNA 2013, 19, 1432–1448. [Google Scholar] [CrossRef] [Green Version]
- Bühler, M.; Steiner, S.; Mohn, F.; Paillusson, A.; Mühlemann, O. EJC-independent degradation of nonsense immunoglobulin-mu mRNA depends on 3′UTR length. Nat. Struct Mol. Biol. 2006, 13, 462–464. [Google Scholar] [CrossRef]
- Kurosaki, T.; Maquat, L.E. Nonsense-mediated mRNA decay in humans at a glance. J. Cell Sci. 2016, 129, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Chakrama, F.Z.; Seguin-Py, S.; Le Grand, J.N.; Fraichard, A.; Delage-Mourroux, R.; Despouy, G.; Perez, V.; Jouvenot, M.; Boyer-Guittaut, M. GABARAPL1 (GEC1) associates with autophagic vesicles. Autophagy 2010, 6, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruyere, D.; Monnien, F.; Colpart, P.; Roncarati, P.; Vuitton, L.; Hendrick, E.; Lepinoy, A.; Luquain, A.; Pilard, C.; Lerho, T.; et al. Treatment algorithm and prognostic factors for patients with stage I-III carcinoma of the anal canal: A 20-year multicenter study. Mod. Pathol. 2021, 34, 116–130. [Google Scholar] [CrossRef]
- Hubert, P.; Herman, L.; Roncarati, P.; Maillard, C.; Renoux, V.; Demoulin, S.; Erpicum, C.; Foidart, J.-M.; Boniver, J.; Noël, A.; et al. Altered α-defensin 5 expression in cervical squamocolumnar junction: Implication in the formation of a viral/tumour-permissive microenvironment. J. Pathol. 2014, 234, 464–477. [Google Scholar] [CrossRef]
- Cao, L.; Qi, L.; Zhang, L.; Song, W.; Yu, Y.; Xu, C.; Li, L.; Guo, Y.; Yang, L.; Liu, C.; et al. Human nonsense-mediated RNA decay regulates EMT by targeting the TGF-ß signaling pathway in lung adenocarcinoma. Cancer Lett. 2017, 403, 246–259. [Google Scholar] [CrossRef]
- Peixoto, P.; Etcheverry, A.; Aubry, M.; Missey, A.; Lachat, C.; Perrard, J.; Hendrick, E.; Delage-Mourroux, R.; Mosser, J.; Borg, C.; et al. EMT is associated with an epigenetic signature of ECM remodeling genes. Cell Death Dis. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawata, M.; Koinuma, D.; Ogami, T.; Umezawa, K.; Iwata, C.; Watabe, T.; Miyazono, K. TGF-β-induced epithelial-mesenchymal transition of A549 lung adenocarcinoma cells is enhanced by pro-inflammatory cytokines derived from RAW 264.7 macrophage cells. J. Biochem. 2012, 151, 205–216. [Google Scholar] [CrossRef]
- Martin, L.; Grigoryan, A.; Wang, D.; Wang, J.; Breda, L.; Rivella, S.; Cardozo, T.; Gardner, L.B. Identification and characterization of small molecules that inhibit nonsense-mediated RNA decay and suppress nonsense p53 mutations. Cancer Res. 2014, 74, 3104–3113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, J.F.; Mogg, A.E. Suppression of a nonsense mutation in mammalian cells in vivo by the aminoglycoside antibiotics G-418 and paromomycin. Nucleic Acids Res. 1985, 13, 6265–6272. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, F.; Breveglieri, G.; Zuccato, C.; Finotti, A.; Bianchi, N.; Borgatti, M.; Feriotto, G.; Destro, F.; Canella, A.; Brognara, E.; et al. Production of beta-globin and adult hemoglobin following G418 treatment of erythroid precursor cells from homozygous beta(0)39 thalassemia patients. Am. J. Hematol. 2009, 84, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Mendell, J.T.; Ap Rhys, C.M.J.; Dietz, H.C. Separable roles for rent1/hUpf1 in altered splicing and decay of nonsense transcripts. Science 2002, 298, 419–422. [Google Scholar] [CrossRef]
- Dassi, E.; Re, A.; Leo, S.; Tebaldi, T.; Pasini, L.; Peroni, D.; Quattrone, A. AURA 2: Empowering discovery of post-transcriptional networks. Translation 2014, 2, e27738. [Google Scholar] [CrossRef]
- Le Grand, J.N.; Chakrama, F.Z.; Seguin-Py, S.; Fraichard, A.; Delage-Mourroux, R.; Jouvenot, M.; Boyer-Guittaut, M. GABARAPL1 (GEC1): Original or copycat? Autophagy 2011, 7, 1098–1107. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Kawamata, T.; Iwasaki, S.; Ohsumi, Y. Selectivity of mRNA degradation by autophagy in yeast. Nat. Commun. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Wengrod, J.; Martin, L.; Wang, D.; Frischmeyer-Guerrerio, P.; Dietz, H.C.; Gardner, L.B. Inhibition of Nonsense-Mediated RNA Decay Activates Autophagy. Mol. Cell. Biol. 2013, 33, 2128–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, L.B. Hypoxic Inhibition of Nonsense-Mediated RNA Decay Regulates Gene Expression and the Integrated Stress Response. Mol. Cell Biol. 2008, 28, 3729–3741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzymski, T.; Milani, M.; Pike, L.; Buffa, F.; Mellor, H.R.; Winchester, L.; Pires, I.; Hammond, E.; Ragoussis, I.; Harris, A.L. Regulation of autophagy by ATF4 in response to severe hypoxia. Oncogene 2010, 29, 4424–4435. [Google Scholar] [CrossRef] [Green Version]
- Toma, K.G.; Rebbapragada, I.; Durand, S.; Lykke-Andersen, J. Identification of elements in human long 3′ UTRs that inhibit nonsense-mediated decay. RNA 2015, 21, 887–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amrani, N.; Ganesan, R.; Kervestin, S.; Mangus, D.A.; Ghosh, S.; Jacobson, A. A faux 3′-UTR promotes aberrant termination and triggers nonsense-mediated mRNA decay. Nature 2004, 432, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Kebaara, B.W.; Atkin, A.L. Long 3′-UTRs target wild-type mRNAs for nonsense-mediated mRNA decay in Saccharomyces cerevisiae. Nucleic Acids Res. 2009, 37, 2771–2778. [Google Scholar] [CrossRef] [Green Version]
- Hauer, C.; Sieber, J.; Schwarzl, T.; Hollerer, I.; Curk, T.; Alleaume, A.-M.; Hentze, M.W.; Kulozik, A.E. Exon Junction Complexes Show a Distributional Bias toward Alternatively Spliced mRNAs and against mRNAs Coding for Ribosomal Proteins. Cell Rep. 2016, 16, 1588–1603. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, T.; Li, W.; Hoque, M.; Popp, M.W.-L.; Ermolenko, D.N.; Tian, B.; Maquat, L.E. A post-translational regulatory switch on UPF1 controls targeted mRNA degradation. Genes Dev. 2014, 28, 1900–1916. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Seo, J.-W.; Ahn, N.; Park, S.; Hwang, J.; Nam, J.-W. UPF1/SMG7-dependent microRNA-mediated gene regulation. Nat. Commun. 2019, 10, 4181. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Target Sequence |
|---|---|
| siUPF1 | 5′-GATGCAGTTCCGCTCCATT-3′ |
| siSMG1 | 5′-GTGTATGTGCGCCAAAGTA-3′ |
| sieIF4A3 | 5′-AGACATGACTAAAGTGGAA-3′ |
| siSTAU1 | 5′-CCUAUAACUACAACAUGAG-3′ |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| 18S | 5′-GTAACCCGTGAACCCCATT-3′ | 5′-CCATCCAATCGGTAGTAGCG-3′ |
| H3B2 | 5′-GCTAGCTGGATGTCTTTTGG-3′ | 5′-GTGGTAAAGCACCCAGGAA-3′ |
| UPF1 | 5′-CGGAAAGCAGCAGTGTACCA-3′ | 5′-GGCAGAAGGGAGTGTGTCTGA-3′ |
| UPF2 | 5′-GAAGATGAGGAAGCCAGCAC-3′ | 5′-CCCGTACCAACTTCTTCCTG-3′ |
| UPF3A | 5′-GAAAAGAGCGACTGGCAAAC-3′ | 5′-TGCCTTGCAGATCCTCCTG-3′ |
| UPF3B | 5′-AAGGGGGAAGATAAGCAGGA-3′ | 5′-GTTCCTGAAGCTGCTCCTTG-3′ |
| SMG1 | 5′-CAAGCGATGTCAGCAGATGT-3′ | 5′-TGCTGACAAAAGCCATTCAG-3′ |
| GABARAPL1 | 5′-CCCTCCCTTGGTTATCATCCA-3′ | 5′-ACTCCCACCCCACAAAATCC-3′ |
| GABARAP | 5′-GCCTTTCCCATCCTGCTGTA-3′ | 5′-GGAAGGGATTGCTGGGTTCT-3′ |
| GABARAPL2 | 5′-TAGTGGCCACAATGACCAGA-3′ | 5′-TGAACACAGCTTTGGTCCAG-3′ |
| LC3B | 5′-CGGAAAGCAGCAGTGTACCA-3′ | 5′-GGCAGAAGGGAGTGTGTCTGA-3′ |
| ATF4 | 5′-CCAAGCACTTCAAACCTCATG-3′ | 5′-ATCCATTTTCTCCAACATCCAATC-3′ |
| ATG5 | 5′-AGGATCAGAATGCAGGGAAC-3′ | 5′-AAGGCCTGACACTGGTTTTG-3′ |
| SC35 | 5′-CGGTGTCCTCTTAAGAAAATGATGTA-3′ | 5′-CTGCTACACAACTGCGCCTTTT-3′ |
| SF3B5 | 5′-ACCGCTACACCATCCATAGC-3′ | 5′-AGGCTGAAGCATCTTTTCCA-3′ |
| GFP | 5′-TCCTGGTCGAGCTGGACGG-3′ | 5′-CAGCTTGCCGTAGGTGGCAT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baudu, T.; Parratte, C.; Perez, V.; Ancion, M.; Millevoi, S.; Hervouet, E.; Peigney, A.; Peixoto, P.; Overs, A.; Herfs, M.; et al. The NMD Pathway Regulates GABARAPL1 mRNA during the EMT. Biomedicines 2021, 9, 1302. https://doi.org/10.3390/biomedicines9101302
Baudu T, Parratte C, Perez V, Ancion M, Millevoi S, Hervouet E, Peigney A, Peixoto P, Overs A, Herfs M, et al. The NMD Pathway Regulates GABARAPL1 mRNA during the EMT. Biomedicines. 2021; 9(10):1302. https://doi.org/10.3390/biomedicines9101302
Chicago/Turabian StyleBaudu, Timothée, Chloé Parratte, Valérie Perez, Marie Ancion, Stefania Millevoi, Eric Hervouet, Anne Peigney, Paul Peixoto, Alexis Overs, Michael Herfs, and et al. 2021. "The NMD Pathway Regulates GABARAPL1 mRNA during the EMT" Biomedicines 9, no. 10: 1302. https://doi.org/10.3390/biomedicines9101302
APA StyleBaudu, T., Parratte, C., Perez, V., Ancion, M., Millevoi, S., Hervouet, E., Peigney, A., Peixoto, P., Overs, A., Herfs, M., Fraichard, A., Guittaut, M., & Baguet, A. (2021). The NMD Pathway Regulates GABARAPL1 mRNA during the EMT. Biomedicines, 9(10), 1302. https://doi.org/10.3390/biomedicines9101302