Astrocytes: News about Brain Health and Diseases


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. The Heterogeneity of Astrocytes
3. Neuroinflammation
4. Traumatic Injuries
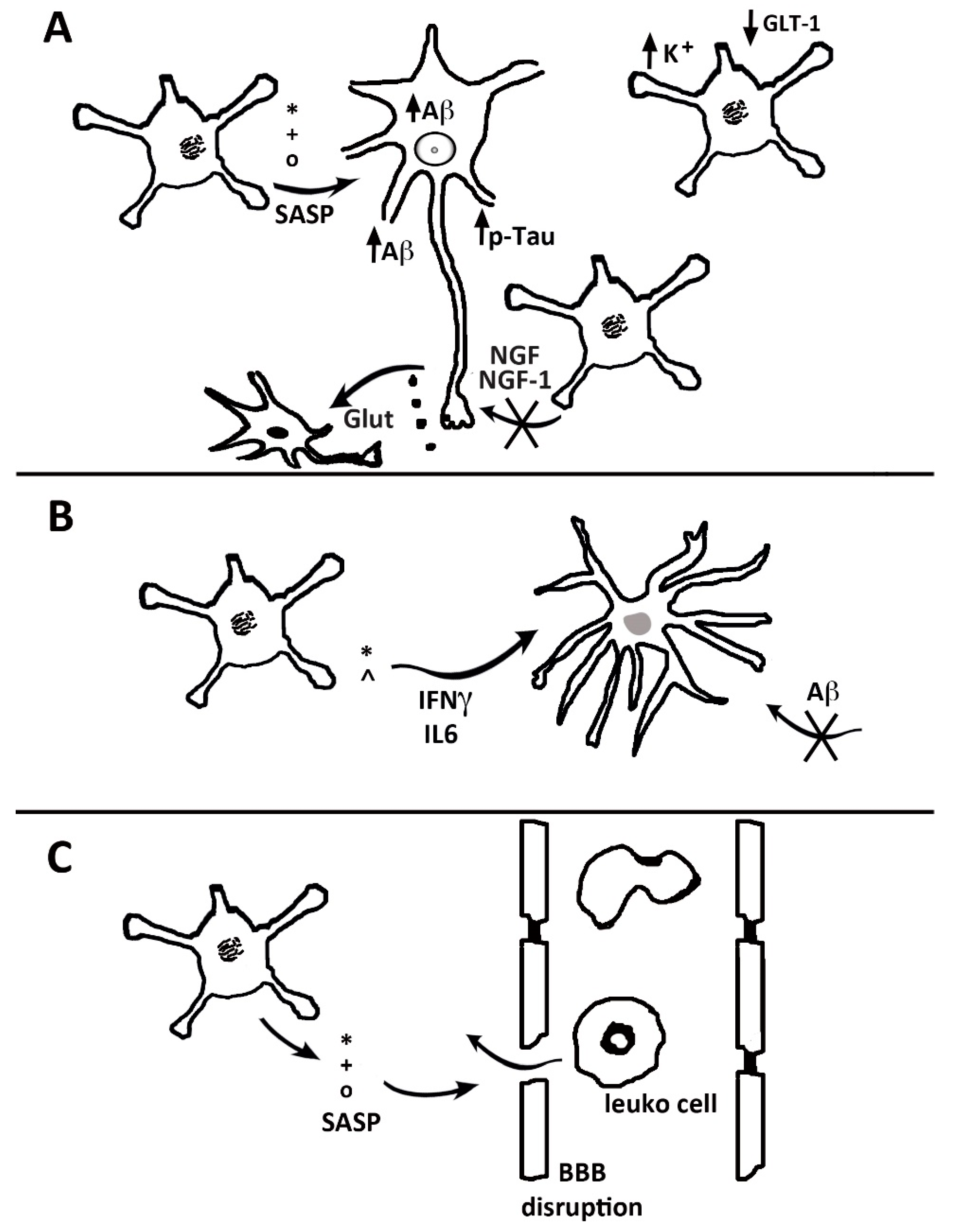
5. Astrocyte Senescence
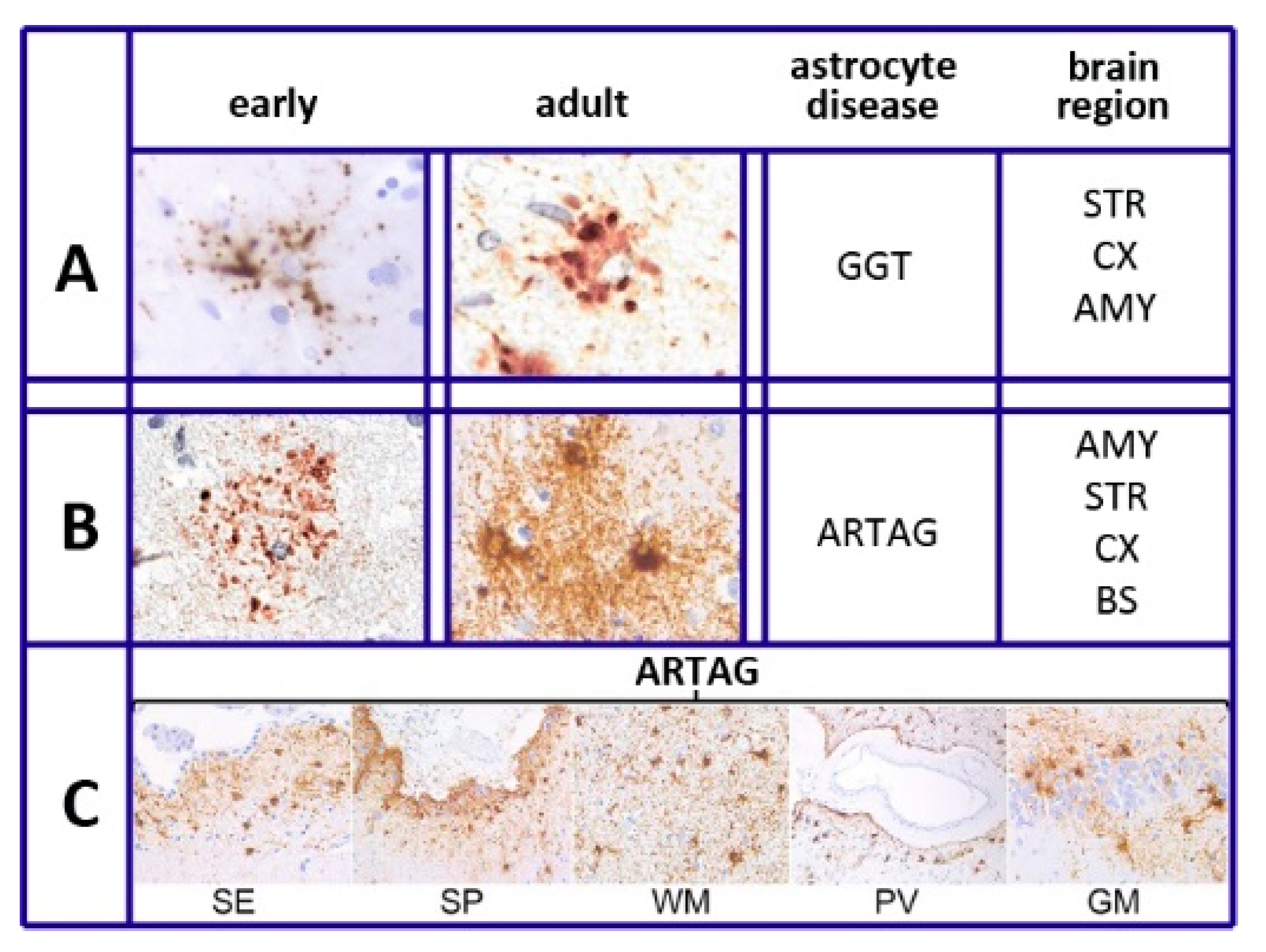
6. Tau and Tauopathies
7. The Role of Astrocytes in Alzheimer’s Disease
8. Conclusions
Funding
Conflicts of Interest
References
- Volterra, A.; Meldolesi, J. Astrocytes, from glue to communication elements: The revolution continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef]
- Allen, N.J.; Eroglu, C. Cell biology of astrocyte-synapse oscillations. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef]
- Bezzi, P.; Gundersen, V.; Galbete, J.S.; Seifert, G.; Steinhauser, C.; Pilati, E.; Volterra, A. Astrocytes contain a vesicular compartment that is competent for regulated exocytosis of glutamate. Nat. Neurosci. 2004, 7, 613–620. [Google Scholar] [CrossRef]
- Pasti, L.; Volterra, A.; Pozzan, T.; Carmignoto, G. Intracellular calcium oscillations in astrocytes: A highly plastic, bidirectional form of communication between neurons and astrocytes in situ. J. Neurosci. 1997, 17, 7817–7830. [Google Scholar] [CrossRef]
- Morita, M.; Higuchi, C.; Moto, T.; Kozuka, N.; Susuki, J.; Itofusa, R.; Yamashita, J.; Kudo, Y. Dual regulation of calcium oscillation in astrocytes by growth factors and pro-inflammatory cytokines via the mitogen-activated protein kinase cascade. J. Neurosci. 2003, 23, 10944–10952. [Google Scholar] [CrossRef]
- Piet, R.; Vargova, L.; Sykova, E.; Poulain, D.A.; Oliet, S.H. Physiological contribution of the astrocytic environment of neurons to inter-synaptic crosstalk. Proc. Natl. Acad. Sci. USA 2004, 101, 2151–2155. [Google Scholar] [CrossRef] [PubMed]
- Middleldorn, J.; Hol, E.M. GFAP in health and disease. Prog. Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.C.; Bak, L.K.; Waagenpetersen, H.S.; Schousboe, A.; Noremberg, M.D. Primary cultures of astrocytes: Their value in understanding astrocytes in health and disease. Neurochem. Res. 2012, 37, 2569–2588. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Fusco, R.; Cuzzocrea, S. Astrocytes: Role and functions in brain pathologies. Front. Pharmacol. 2019, 10, 114. [Google Scholar] [CrossRef] [PubMed]
- Cragnolini, A.B.; Lampitella, G.; Virtuoso, A.; Viscovo, I.; Panetson, F.; Papa, M.; Cirillo, G. Regonal brain susceptibility to neurodegeneration: What is the role of glial cells? Neural Regen. Res. 2020, 15, 838–842. [Google Scholar] [PubMed]
- Zhang, Y.; Barres, B.A. Astrocyte heterogeneity: An underappreciated topic in neurobiology. Curr. Opin. Neurobiol. 2010, 20, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gonzalo, M.; Martin-Fernandez, M.; Martinez-Murillo, R.; Maderos, S.; Hernandez-Vivanco, A.; Jamieson, S.; Fernandez, A.P.; Serrano, J.; Calero, P.; Futch, H.S.; et al. Neuron-astrocyte signaling is preserved in the aging brain. Glia 2017, 65, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; Deneen, B. The emerging nature of astrocyte diversity. Ann. Rev. Neurosci. 2019, 42, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Munch, A.E.; Heiman, M.; Barres, B.A. Normal aging induces A1-like astrocyte reactivity. Proc. Natl. Acad. Sci. USA 2018, 115, E1896–E1905. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.L.; Ousman, S.S. Astrocytes and aging. Front. Aging Neurosci. 2018, 10, 337. [Google Scholar] [CrossRef]
- Mathias, I.; Morgado, J.; Gomes, F.C.A. Astrocyte heterogeneity: Input to brain aging and disease. Front. Aging Neurosci. 2019, 11, 59. [Google Scholar] [CrossRef]
- Miki, T.; Yokota, O.; Haraguchi, T.; Ishizu, H.; Hasegawa, M.; Ishihara, T.; Ueno, S.I.; Takenoshita, S.; Terada, S.; Tamada, N. Factors associated with development and distribution of granular/fuzzy astrocytes in neurodegenerative diseases. Brain Pathol. 2020, 30, 811–830. [Google Scholar] [CrossRef]
- Trias, E.; Barbeito, L.; Yamanaka, K. Phenotypic heterogeneity of astrocytes in motor neuron disease. Clin. Exp. Neuroimmunol. 2018, 9, 225–234. [Google Scholar] [CrossRef]
- Pan, J.; Maa, N.; Yu, B.; Zhang, W.; Wan, J. Transcriptomic profiling of microglia and astrocytes throughout aging. J. Neuroinflammation 2020, 17, 97. [Google Scholar] [CrossRef]
- Boisvert, M.M.; Erikson, G.A.; Shokhivert, M.N.; Allen, N.J. The aging astrocyte transcriptome: From multiple regions of the mouse brain. Cell Rep. 2018, 22, 269–285. [Google Scholar] [CrossRef]
- Ben Haim, L.; Rowitch, D.H. Functional diversity of astrocytes in neural circuits. Nat. Rev. Neurosci. 2017, 18, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G. Astroglia and tau: New perspectives. Front. Aging Neurosci. 2020, 12, 96. [Google Scholar] [CrossRef] [PubMed]
- Bronzuoli, M.R.; Facchinetti, R.; Valenza, M.; Cassano, T.; Steardo, L.; Scuderi, C. Astrocyte function is affected by aging and not Alzheimer’s disease: A preliminary investigation in hippocampi of 3xTg-AD mice. Front. Pharmacol. 2019, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, F.; Quintana, F.J. The role of astrocytes in CNS inflammation. Trends Immunol. 2020, 41, 805–819. [Google Scholar] [CrossRef]
- Gimsa, U.; Michison, N.A.; Brunner-Weinzieri, M.C. Immune privilege as an intrinsic CNS property: Astrocytes protect the CNS against T-cell-mediated neuroinflammation. Mediators Inflamm. 2013, 2013, 320519. [Google Scholar] [CrossRef]
- Schafer, S.; Calas, A.G.; Vergouts, M.; Hermans, E. Immuno-modulatory influence of bone marrow-derived mesenchymal stem cells on neuroinflammation in astrocyte cultures. J. Neuroimmunol. 2012, 249, 40–48. [Google Scholar] [CrossRef]
- Colombo, E.; Farina, C. Astrocytes: Key regulators of neuroinflammation. Trends Immunol. 2016, 37, 606–620. [Google Scholar] [CrossRef]
- Stephenson, J.; Nutma, E.; Van der Valk, P.; Amor, S. Inflammation of CNS neurodegenerative diseases. Immunology 2018, 154, 204–2018. [Google Scholar] [CrossRef]
- Sochoka, M.; Diniz, B.S.; Leszek, J. Inflammatory responses in the CNS: Fried or foe? Mol. Neurobiol. 2017, 54, 8071–8089. [Google Scholar] [CrossRef]
- Alsegiani, A.S.; Shah, Z.A. The role of colfilin in age-related neuroinflammation. Rev. Neural Regen. Res. 2020, 15, 1451–1459. [Google Scholar]
- Lana, D.; Invino, L.; Nosi, D.; Wenk, G.L.; Giovannino, M.G. The neuro-astrocyte-microglia triad involvement in neuroinflammating mechanisms in the CA3 hippocampus of memory impaired aged rats. Exp. Gerontol. 2016, 83, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, H.A.; Clarke, L.E.; Bennett, F.C.; Bolen, C.J.; Schirmer, L.; Bennett, L.; Munch, A.E.; Chung, W.S.; Petersen, C.P.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Santos, C.L.; Bobermin, L.D.; Souza, D.O.; Quincozes-Santos, A. Leptin stimulates the release of pro-inflammatory cytokines in hypothalamic astrocyte cultures from adult and aged rats. Metab. Brain Dis. 2018, 33, 2059–2063. [Google Scholar] [CrossRef] [PubMed]
- Leng, L.; Zhuang, K.; Liu, Z.; Huang, C.; Yuehong, G.; Guimiao, C.; Lin, H.; Hu, Y.; Wu, D.; Shi, M.; et al. Menin deficiency leads to depressive-like behaviors in mice by modulating astrocyte-mediated neuro-inflammation. Neuron 2018, 100, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Sompol, P.; Norris, C.M. Ca2+, astrocyte activation and calcineurin/NFAT signaling in age-related neurodegenerative diseases. Front. Aging Neurosci. 2018, 10, 199. [Google Scholar] [CrossRef]
- Wang, J.L.; Xu, C.J. Astrocyte autophagy in aging and neurodegenerative disorders. Biomed. Pharmacother. 2020, 122, 109691. [Google Scholar] [CrossRef]
- Qju, Y.; Pan, J.; Li, Y.; Li, X.; Li, M.; Abukhousa, I.; Wang, Y. Relationship between activated astrocytes and hypoxic cerebral tissue in a rat model of cerebral ischemia/reperfusion. Int. J. Neurosci. 2011, 121, 1–7. [Google Scholar]
- Barreto, G.; White, R.E.; Ouyang, Y.; Xu, L.; Giffard, R.G. Astrocytes: Targets for neuroprotection in stroke. Cent. Nerv. Syst. Agents Med. Chem. 2011, 11, 164–173. [Google Scholar] [CrossRef]
- Sutherland, T.C.; Geoffroy, C.G. The influence of neuron-extrinsic factors and aging on injury progression and axonal repair in the central nervous system. Front. Cell Dev. Biol. 2020, 8, 190. [Google Scholar] [CrossRef]
- Duan, C.L.; Ciu, C.W.; Shen, S.W.; Zhang, Y.; Mo, J.L.; Chen, X.H.; Sun, F.Y. Striatal astrocytes transdifferentiate into functional mature neurons following ischemic brain injury. Glia 2015, 63, 1660–1670. [Google Scholar] [CrossRef]
- Shen, S.W.; Duan, C.L.; Chen, X.H.; Wang, Y.Q.; Sun, X.; Zhang, Q.W.; Cui, H.R.; Sun, S.Y. Neurogenic effects of VEGF are related to increase of astrocytes transdifferentiaton into new mature neurons in rat brain after stroke. Neuropharmacology 2016, 108, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.L.; Liu, O.; Kou, Z.W.; Wu, K.W.; Yang, P.; Chen, X.H.; Sin, F.Y. MicroRNA-365 modulates astrocyte conversion into neuron in adult rat brain after stroke by targeting Pax6. Glia 2018, 66, 1346–1362. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, B.B.; Buthani, A.; Stary, C.M. Adult neurogenesis from reprogrammed astrocytes. Rev. Neural Regen. Res. 2020, 15, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Michelucci, A.; Bithell, A.; Burney, M.J.; Johnston, C.E.; Wong, K.T.; Teng, S.W.; Desai, J.; Teng, S.W.; Desai, J.; Gumbleton, N.; et al. The neurogenic potential of astrocytes is regulated by inflammatory signals. Mol. Neurobiol. 2016, 53, 3724–3739. [Google Scholar] [CrossRef] [PubMed]
- Burda, J.E.; Bernstein, A.M.; Sofroniew, M.J. Astrocyte roles in traumatic brain injury. Exp. Neurol. 2016, 275, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, G.R.; Ding, S. Reactive astrocytes and therapeutic potential in focal ischemic stroke. Neurobiol. Dis. 2016, 85, 234–244. [Google Scholar] [CrossRef]
- Pivokova, H.; Anderova, M. Altered homeostatic functions in reactive astrocytes and their potential as a therapeutic target after brain ischemic injury. Curr. Pharm. Des. 2017, 23, 5056–5074. [Google Scholar]
- Revuelta, M.; Elicegui, A.; Moreno-Cugnon, L.; Buhrer, C.; Mautheu, A.; Schmitz, T. Ischemic stroke in neonatal and adult astrocytes. Mech. Aging Dev. 2019, 183, 111147. [Google Scholar] [CrossRef]
- Early, A.N.; Gorman, A.A.; Van Eldik, L.J.; Bachstetter, A.D.; Morganti, J.M. Effects of advanced age upon astrocyte-specific responses to acute traumatic brain injury in mice. J. Neuro-Inflamm. 2020, 17, 113. [Google Scholar] [CrossRef]
- Pereira Diniz, L.; Mathia, I.; Siqueira, M.; Stipursky, J.; Gomes, F.C.A. Astrocytes and TGF-β1 pathway in the healthy and diseased brain: A double-edged sword. Mol. Neurobiol. 2019, 56, 4653–4679. [Google Scholar] [CrossRef]
- Das, M.M.; Avalos, P.; Suezaki, P.; Godoy, M.; Garcia, L.; Chang, C.D.; Vit, J.P.; Schelley, B.; Gowing, G.; Svendsen, C.N. Human neural progenitors differentiate into astrocytes and protect motor neurons in aging rats. Exp. Neurol. 2016, 280, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, W.; Wang, S.; Xie, W.; Li, H.; Ning, B. MicroRNA-21 regulates astrocyte reaction post-acute phase of spinal cord injury through modulating TGF-β signaling. Aging 2018, 10, 1474–1488. [Google Scholar] [CrossRef] [PubMed]
- Raihan, O.; Brishti, A.; Molla, M.R.; Li, W.; Ahang, Q.; Peng, X.; Khan, M.I.; Zhang, J.; Liu, Q. The age-dependent elevation of miR335-3p leads to refused cholesterol and impaired memory in brain. Neuroscience 2018, 390, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Ojala, A.; Kaarninarant, K.; Haapsalo, A.; Hiltunen, A.; Soininen, H. Astrocytes in the aging brain express characteristics of senescence-activated secretory phenotype. Eur. J. Neurosci. 2011, 34, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Lye, I.I.; Latorre, E.; Lee, B.P.; Bandinelli, S.; Holley, J.E.; Gutowski, N.J.; Ferrucci, L.; Harries, L.W. Astrocyte senescence may drive alterations of GFAPa, CDKN2A, p14ARF, and tau3 transcript expression and contribute to cognitive decline. Geroscience 2019, 41, 561–573. [Google Scholar] [CrossRef]
- Santoro, A.; Spinelli, C.C.; Martucciello, S.; Nori, S.L.; Capunzo, M.; Puca, A.A.; Ciaglia, E. Innate immunity and cellular senescence: The good and bad in the developmental and aged brain. J. Leukoc. Biol. 2018, 103, 509–524. [Google Scholar] [CrossRef]
- Bussian, T.J.; Aziz, A.; Leyer, C.F.; Swenson, B.L.; van Deursen, J.M.; Baker, D.J. Clearance of senescent glial cells prevents tau-dependent pathology and cognitive decline. Nature 2018, 562, 578–582. [Google Scholar] [CrossRef]
- Crowe, E.P.; Tuzer, F.; Gregory, B.D.; Donahue, G.; Gosay, S.J.; Cohen, J.; Leung, Y.Y.; Yetkin, E.; Nativio, R.; Wang, L.S.; et al. Changes of the transcriptome in human astrocytes accompanying oxidative stress-induced senescence. Front. Aging Neurosci. 2016, 8, 208. [Google Scholar] [CrossRef]
- Baker, D.J.; Petersen, R.C. Cellular senescence in brain aging and neurodegenerative diseases: Evidence and perspectives. J. Clin. Investig. 2018, 128, 1208–1216. [Google Scholar] [CrossRef]
- Limbad, C.; Oron, T.R.; Almirah, F.; Davalos, A.R.; Tracy, T.E.; Gen, L.; Desprez, P.Y.; Campisi, J. Astrocyte senescence promotes glutamate toxicity in cortical neurons. PLoS ONE 2020, 15, e0227887. [Google Scholar] [CrossRef]
- Hou, I.; Cui, C.; Kim, S.; Sung, C.; Choi, C. Ginsenoside F1 suppresses astrocytic senescence-associated secretory phenotype. Chem. Biol. Interact. 2018, 283, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Vande Vrede, L.; Liubenkov, P.A.; Rojas, J.C.; Welch, A.E.; Boxer, A.L. Four-repeat tauopathies: Current management and future treatments. Neurotherapeutics 2020. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Doherty, K.M.; Silveira-Moriyama, L.; Bamdopadyay, R.; Leshley, T.; Mamais, A.; Hondhamuni, G.; Wray, S.; Newcombe, J.; O’Sullivan, S.S.; et al. Globular glial tauopathies (GGT) presenting with motor neuron disease of frontotemporal dementia: An emerging group of 4-repeat tauopathies. Acta Neuropathol. 2011, 122, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Clavaguera, F.; Duyckaertis, C.; Haik, S. Prion-like properties of tau assemblies. Curr. Opin. Neurobiol. 2020, 61, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Santpere, G.; Ferrer, I. Delineation of early changes in cases with progressive supranuclear palsy-like pathology. Astrocytes in striatum are primary targets of tau phosphorylation and GFAP oxidation. Brain Pathol. 2009, 19, 177–187. [Google Scholar] [CrossRef]
- Lance, G.; Ince, P.G.; Brayne, C.; Sayva, G.M.; Mattews, F.E.; de Silva, R.; Simpson, J.E.; Wharton, S.B. Mesial temporal astrocyte tau pathology in the MRC-CFAS aging brain cohort. Dement. Geriatr. Cogn. Disord. 2012, 34, 15–24. [Google Scholar]
- Ferrer, I.; Lopez-Gonzalez, J.; Carmona, M.; Arregui, L.; Dalfo, E.; Torrejon-Escribano, B.; Diehl, R.; Kovacs, G.G. Glial and neuronal tau pathology in tauopathies: Characterization of disease-specific phenotypes and tau pathology progression. J. Neuropathol. Exp. Neurol. 2014, 73, 81–97. [Google Scholar] [CrossRef]
- Sidoryk-Wegrzynowicz, M.; Gerber, Y.N.; Ries, M.; Sastre, M.; Tolkovsky, A.M.; Spillantini, M.G. Astrocytes in mouse models of tauopathies acquire early deficits and lose neurosupportive functions. Acta Neuropathol. Commun. 2017, 5, 89. [Google Scholar] [CrossRef]
- Narasimhan, S.N.; Changolkar, L.; Riddle, D.M.; Kats, A.; Stieber, A.; Weitzman, S.A.; Zhang, B.; Li, Z.; Robertson, E.D.; Trojanowski, J.Q.; et al. Human tau pathology transmits glial tau aggregates in the absence of neuronal tau. J. Exp. Med. 2020, 217, e20190783. [Google Scholar] [CrossRef]
- Kovacs, G.G.; Robinson, J.L.; Xie, S.X.; Lee, E.B.; Grossman, M.; Wolk, D.A.; Irwin, D.J.; Weintraub, D.; Kim, C.F.; Schuck, T.; et al. Evaluating the patterns of aging-related tau astrogliopathy unravels novel insights into brain aging and neurodegenerative diseases. J. Neuropathol. Exp. Neurol. 2017, 76, 270–288. [Google Scholar] [CrossRef]
- Ferrer, I.; Garcia, M.A.; Lopez-Gonzalez, I.; Diaz-Lucerna, D.; Roig-Villalonga, A.; Carmona-Tech, M.; Lorens, F.; Garcia-Esparcia, P.; Martinez-Maldonado, A.; Frau Mendez, M.; et al. Aging-related tau astrogliopathy (ARTAG): Not only tau phosphorylation in astrocytes. Brain Pathol. 2018, 28, 965–985. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G.; Xie, S.X.; Robinson, J.L.; Lee, E.B.; Smith, D.H.; Schuck, T.; Lee, V.M.; Trojanowski, J.Q. Sequential stages and distribution patterns of aging-related tau astrogliopathy (ARTAG) in the human brain. Acta Neuropathol. Commun. 2018, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Amari, M.; Fukuda, T.; Suzuki, K.; Takatama, M. Astrocytic tau pathologies in aged human brain. Neuropathology 2019, 39, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Nolan, A.; Resende, E.P.F.; Petersen, C.; Neylan, K.; Spina, S.; Huang, E.; Seeley, W.; Miller, Z.; Greenberg, T. Astrocyte tau deposition is frequent in typical and atypical Alzheimer’s disease presentations. J. Neuropathol. Exp. Neurol. 2019, 78, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Yan, P.; Ma, X.; Liu, H.; Perez, R.; Zhu, A.; Gonzalez, E.; Burkett, J.M.; Schuler, D.R.; Cirrito, J.R.; et al. Enhancing astrocyte lysosome biogenesis facilitates Aβ clearance and attenuates amyloid plaque pathogenesis. J. Neurosci. 2014, 34, 9607–9620. [Google Scholar] [CrossRef]
- Martini-Stoica, H.; Cole, A.L.; Swartzlander, D.B.; Chen, F.; Wan, F.; Bajaj, L.; Bader, D.A.; Lee, V.M.Y.; Trojanowski, J.Q.; Liu, Z.; et al. TFEB enhances astroglial uptake of extracellular tau species and reduces tau spreading. J. Exp. Med. 2018, 215, 2355–2377. [Google Scholar] [CrossRef]
- De Sampayo-Spohr, T.C.; Martinez, R.; De Silva, E.F.; Neto, V.M.; Gomes, F.C. Neuro-glia interaction effects on GFAP gene: A novel role for transforming growth factor-β1. Eur. J. Neurosci. 2002, 16, 2059–2069. [Google Scholar] [CrossRef]
- Simpson, J.E.; Ince, P.G.; Forster, L.G.; Shaw, P.J.; Matthews, F.; Savva, G.; Brayne, C.; Wharton, S.B. Astrocyte phenotype in relation to Alzheimer-type pathology in the aging brain. Neurobiol. Aging 2010, 31, 578–590. [Google Scholar] [CrossRef]
- Allaman, I.; Gavillet, M.; Bélanger, M.; Laroche, T.; Vertl, D.; Laschuel, H.A.; Magistretti, P.J. Amyloid-β aggregates cause alterations of astrocyte metabolic phenotype: Impact on neuronal viability. J. Neurosci. 2010, 30, 3326–3338. [Google Scholar] [CrossRef]
- Habib, N.; McCabe, C.; Medina, S.; Varshavsky, M.; Kitsberg, D.; Dvir-Sternfeld, R.; Green, G.; Dionne, D.; Nguyen, L.; Marshall, J.L.; et al. Disease-associated astrocytes in Alzheimer’s disease and aging. Nat. Neurosci. 2020, 23, 701–706. [Google Scholar] [CrossRef]
- Zorec, R.; Parpura, V.; Vardjan, N.; Verkhratsky, A. Astrocytic face of Alzheimer’s disease. Behav. Brain. Res. 2017, 322, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Arranz, A.M.; De Strooper, B. The role of astroglia in Alzheimer’s disease: Pathophysiology and clinical implications. Lancet Neurol. 2019, 18, 406–414. [Google Scholar] [CrossRef]
- Liang, Y.; Raven, F.; Ward, J.F.; Zhen, S.; Zhang, S.; Sun, H.; Miller, S.J.; Choi, S.H.; Tanzi, R.E.; Zhang, C. Up-regulation of Alzheimer’s disease amyloid-β protein precursor in astrocytes both in vitro and in vivo. J. Alzheimer’s Dis. 2020, 76, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Perez-Nievas, B.G.; Serrano-Pozo, A. Deciphering the astrocyte reaction in Alzheimer’s disease. Front. Aging Neurosci. 2018, 10, 114. [Google Scholar] [CrossRef]
- Li, K.; Li, J.; Zheng, J.; Quin, S. Reactive astrocytes in neuro-degenerative diseases. Aging Dis. 2019, 10, 664–675. [Google Scholar] [CrossRef]
- Ceyzériat, K.; Haim, L.B.; Denizot, A.; Pommier, D.; Matos, M.; Guillemaud, O.; Palomares, M.-A.; Abjean, L.; Petit, F.; Gipchtein, P.; et al. Modulation of astrocyte reactivity improves functional deficits in mouse models of Alzheimer’s disease. Acta Neuropathol. Commun. 2018, 6, 104. [Google Scholar] [CrossRef]
- Guillemaud, O.; Ceyzériat, K.; Saint-Georges, T.; Cambon, K.; Petit, F.; Haim, B.; Carillo-de Sauvage, M.A.; Guillermier, M.; Bernier, S.; Herald, H.S.; et al. Complex roles for reactive astrocytes in the triple transgenic mouse model of Alzheimer’s disease. Neurobiol. Aging 2020, 90, 135–146. [Google Scholar] [CrossRef]
- Zhao, W.; Fan, J.; Kulic, I.; Koh, C.; Clark, A.; Mueller, J.; Engwist, O.; Barichievy, S.; Raynoschek, C.; Hicks, R.; et al. Axl receptor tyrosine kinase is a regulator of apolipoprotein E. Mol. Brain 2020, 13, 66. [Google Scholar] [CrossRef]
- Shentu, Y.P.; Hu, W.T.; Zhang, Q.; Huo, Y.; Liang, J.W.; Liuyang, Z.Y.; Zhu, H.; Wei, H.; Ke, D.; Wang, X.C.; et al. CIP2A-promoted astrogliosis, induces AD-like synaptic degeneration and cognitive deficits. Neurobiol. Aging 2019, 75, 198–208. [Google Scholar] [CrossRef]
- Son, S.M.; Nam, D.W.; Cha, M.Y.; Kim, K.H.; Byun, J.; Ryu, H.; Mook-Jung, I. Thrombospondin-1 prevents amyloid-β-mediated synaptic pathology in Alzheimer’s disease. Neurobiol. Aging 2015, 36, 3214–3227. [Google Scholar] [CrossRef]
- Diniz, L.P.; Tortelli, V.; Mahias, I.; Morgado, J.; Bergamo Araujo, A.P.; Melo, H.M.; Seixas da Silva, G.S.; Alves-Leon, S.V.; de Souza, J.M.; Ferreira, S.T.; et al. Astrocyte transforming growth factor-β1 protects synapses against Aβ oligomers in Alzheimer’s disease model. J. Neurosci. 2017, 37, 6797–6809. [Google Scholar] [CrossRef] [PubMed]
- Trenblay, M.E.M.; Cookson, M.R.; Civiero, L. Glial phagocytic clearance in Parkinson’s disease. Mol. Neurodegener. 2019, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Ries, M.; Sastre, M. Mechanisms of clearance and degradation by glial cells. Front. Aging Neurosci. 2016, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kazim, S.F.; Larson, C.S.; Ramakrushnan, A.; Gray, J.D.; McEwen, B.S.; Rosemberg, B.A.; Shen, L.; Pereira, A.C. Divergent roles of astrocytic versus neuronal EAAT2 deficiency on cognitive and overlap with aging and Alzheimer’s molecular signatures. Proc. Natl. Acad. Sci. USA 2019, 116, 21800–21811. [Google Scholar] [CrossRef]
- Fruhbeis, C.; Frohlich, D.; Luo, W.P.; Kramer-Albers, E.M. Extracellular vesicles as mediators of neuron-glia communication. Front. Cell Neurosci. 2013, 7, 182. [Google Scholar] [CrossRef]
- Goetzl, E.J.; Schwartz, J.B.; Abner, E.L.; Jicha, G.A.; Kapogiannis, D. High complement levels in astrocyte-derived exosomes of Alzheimer disease. Ann. Neurol. 2018, 83, 544–552. [Google Scholar] [CrossRef]
- Elsharbini, A.; Kirov, A.S.; Dinkins, M.B.; Wang, G.; Quin, H.; Zhu, Z.; Tripathi, P.; Crivelli, S.M.; Bieberich, E. Association of Aβ with ceramide-enriched astrosomes mediates Aβ nerotoxicity. Acta Neuropathol. Commun. 2020, 8, 60. [Google Scholar] [CrossRef]
- Li, K.Y.; Gong, P.F.; Li, J.T.; Xu, N.J.; Qin, S. Morphological and molecular alterations of reactive astrocytes without proliferation in cerebral cortex of an APP/PS1 transgenic mouse model and Alzheimer’s patients. Glia 2020, 68, 2361–2376. [Google Scholar]
- Brancaggio, M.; Edwards, M.D.; Patton, A.P.; Smyllie, N.J.; Maywood, E.S.; Hasting, M.H. Cell autonomous clock of astrocytes drives circadian behavior in mammals. Science 2019, 363, 187–192. [Google Scholar] [CrossRef]
- McKee, C.A.; Lananna, B.V.; Musiel, E.S. Circadian regulation of astrocyte function: Implications for Alzheimer’s disease. Cell Mol. Life Sci. 2020, 77, 1049–1058. [Google Scholar] [CrossRef]



© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meldolesi, J. Astrocytes: News about Brain Health and Diseases. Biomedicines 2020, 8, 394. https://doi.org/10.3390/biomedicines8100394
Meldolesi J. Astrocytes: News about Brain Health and Diseases. Biomedicines. 2020; 8(10):394. https://doi.org/10.3390/biomedicines8100394
Chicago/Turabian StyleMeldolesi, Jacopo. 2020. "Astrocytes: News about Brain Health and Diseases" Biomedicines 8, no. 10: 394. https://doi.org/10.3390/biomedicines8100394
APA StyleMeldolesi, J. (2020). Astrocytes: News about Brain Health and Diseases. Biomedicines, 8(10), 394. https://doi.org/10.3390/biomedicines8100394