

Neurobehavioral and Oxidative Stress Effects of SiO2 Nanoparticles in Zebrafish and the Protective Role of N-Acetylcysteine

 , , ,
, , ,  ,
, 
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
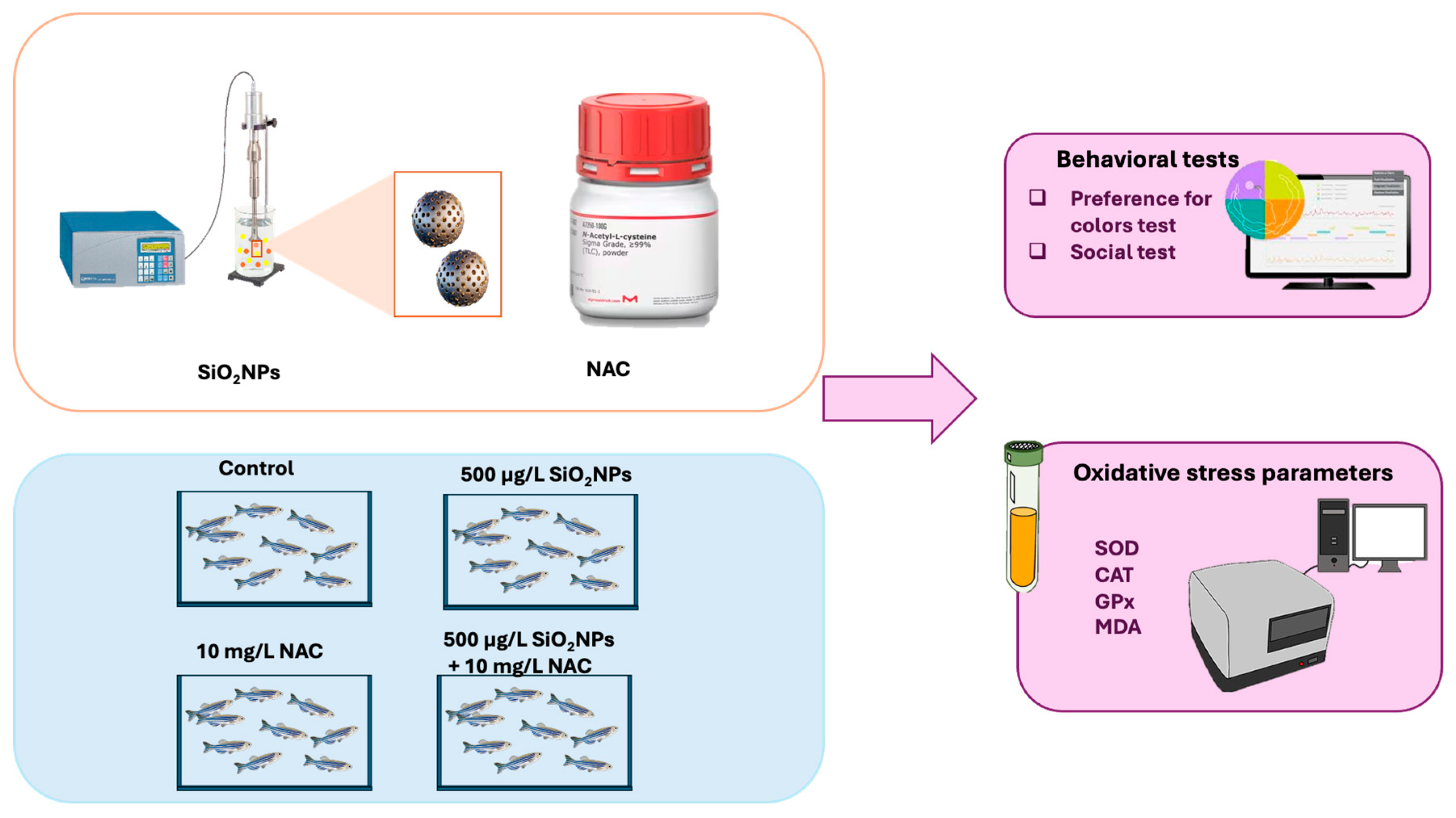
2. Materials and Methods
2.1. Zebrafish
2.2. Materials
2.3. Zebrafish Exposed to SiO2NPs and NAC
2.3.1. Experimental Setup for Behavioral Testing
2.3.2. Acclimatization for Preference Tests
2.3.3. Behavioral Parameters for Color Preference
2.3.4. Behavioral Parameters for Social Test
2.4. Preparation of Homogenates and Biochemical Parameter Analysis
2.4.1. Superoxide Dismutase Activity Determination
2.4.2. Catalase Activity Determination
2.4.3. Glutathione Peroxidase Activity Determination
2.4.4. Malondialdehyde Level Determination
2.5. Statistical Analysis
3. Results
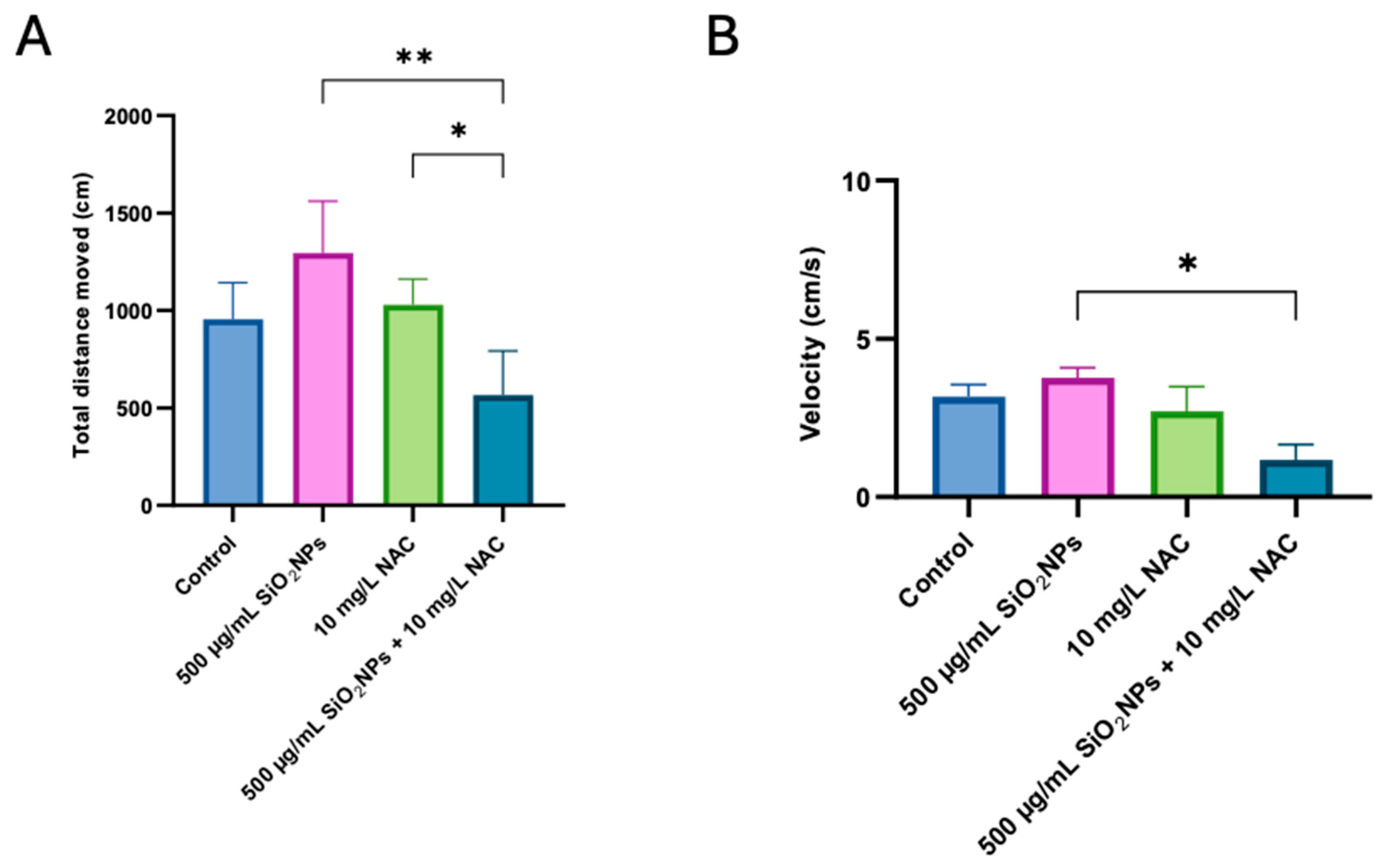
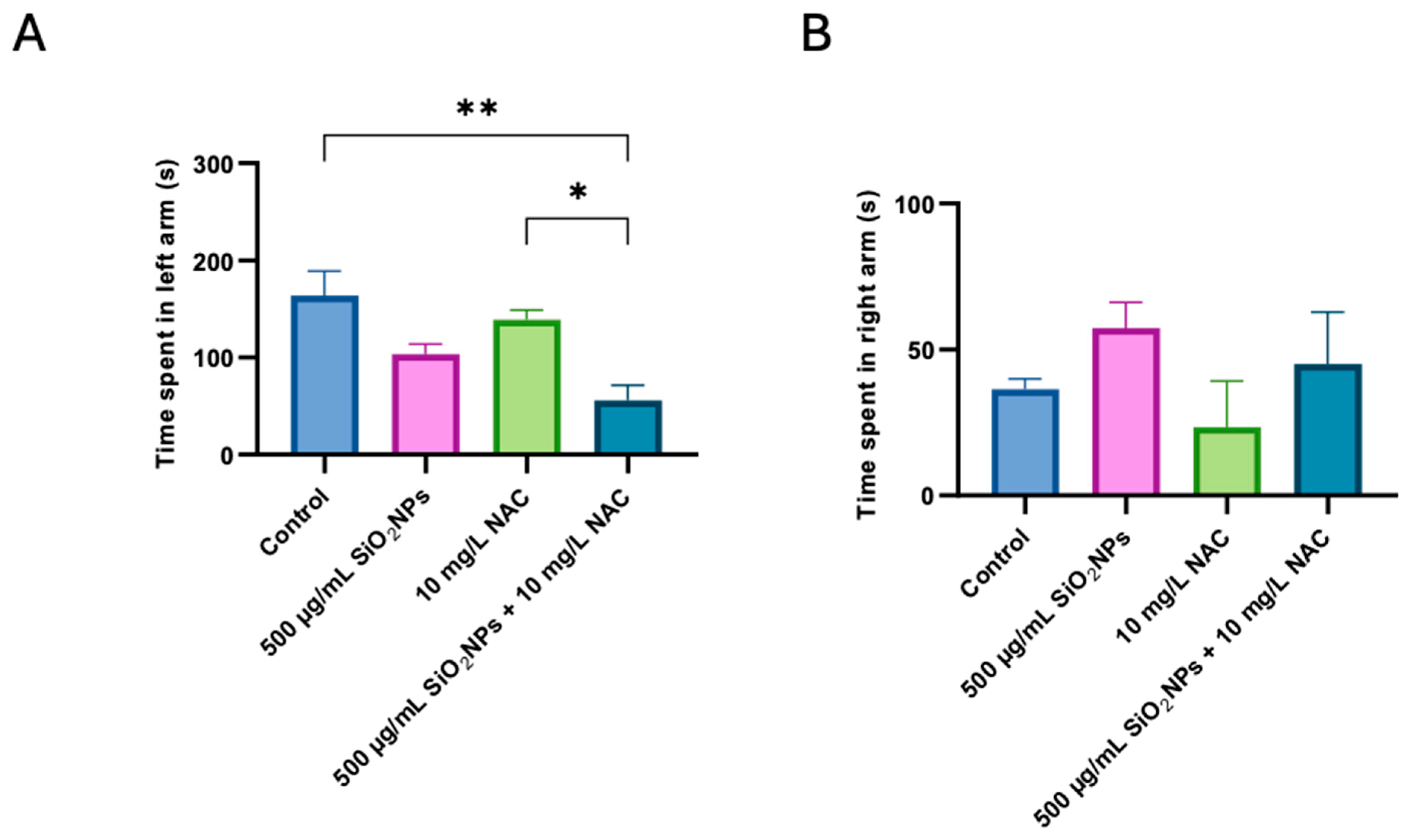
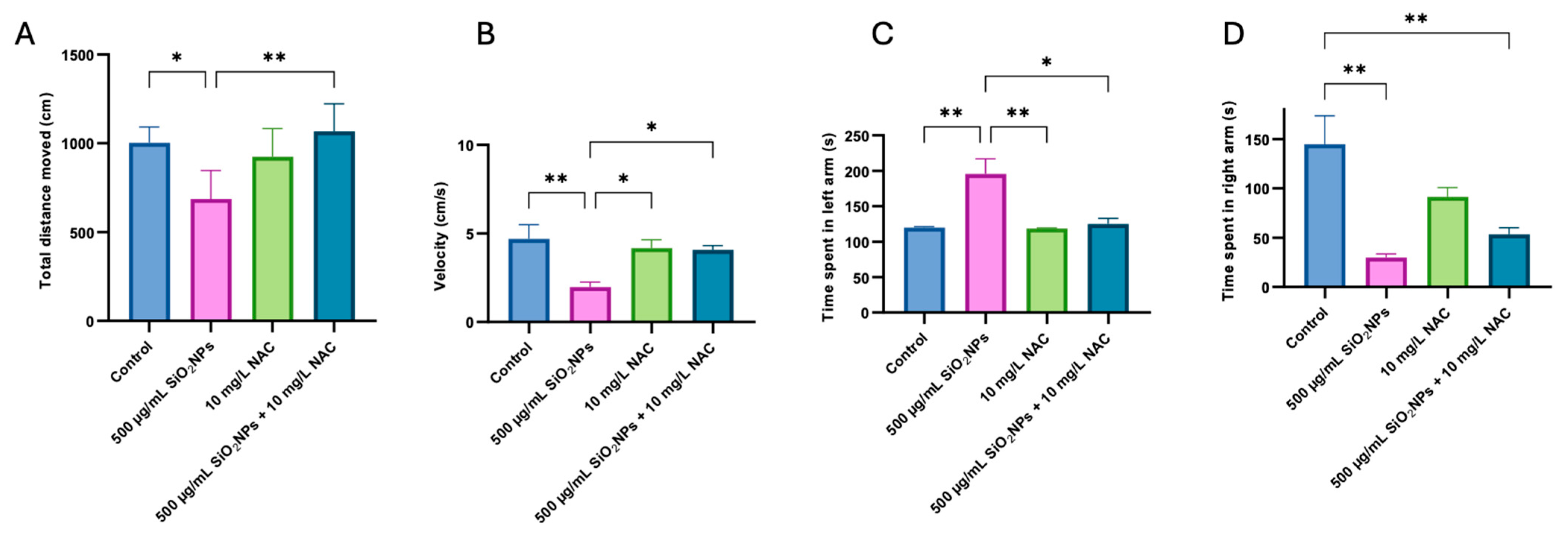
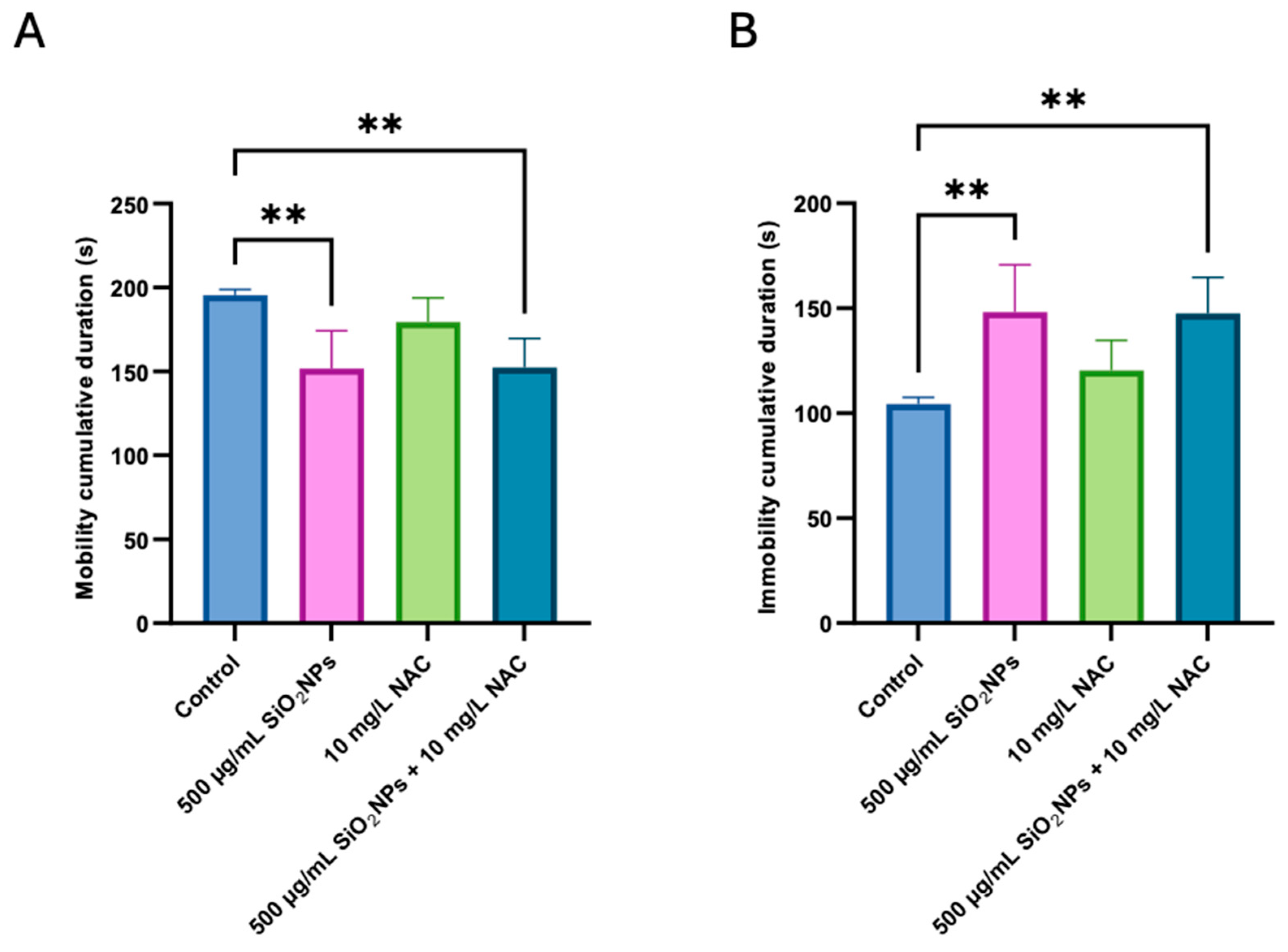
3.1. Social Test
3.2. Preference for Colors Test
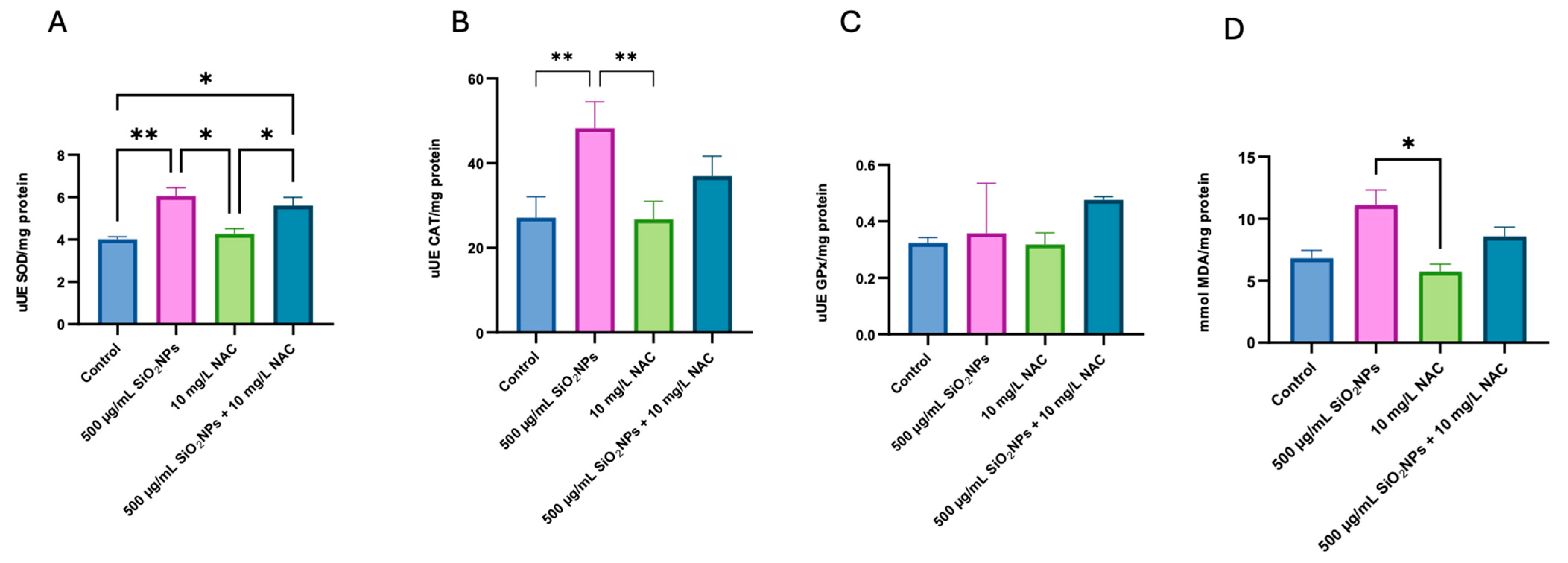
3.3. Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.; Li, H.; Xiao, K. Distribution and Biological Effects of Nanoparticles in the Reproductive System. Curr. Drug Metab. 2016, 17, 478–496. [Google Scholar] [CrossRef] [PubMed]
- Moraes, G.; Zambom, C.; Siqueira, W.L. Nanoparticles in Dentistry: A Comprehensive Review. Pharmaceuticals 2021, 14, 752. [Google Scholar] [CrossRef] [PubMed]
- Dubey, R.S.; Rajesh, Y.B.R.D.; More, M.A. Synthesis and Characterization of SiO2 Nanoparticles via Sol-Gel Method for Industrial Applications. Mater. Today Proc. 2015, 2, 3575–3579. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, L.; Yang, W. Synthesis of Mesoporous Silica Using the Sol–Gel Approach: Adjusting Architecture and Composition for Novel Applications. Nanomaterials 2024, 14, 903. [Google Scholar] [CrossRef] [PubMed]
- Palanimuthu, V.; Periakaruppan, R.; Romanovski, V.; Bharathi, A.; Vijai Selvaraj, K.S.; Anukeerthana, S.; Nishanthi, R.; Vanajadevi, G. Synthesis and Structural Characterization of SiO2 Nanoparticles Using Extract of Gracilaria crassa via Green Chemistry Approach. ChemistryOpen 2025, 14, e202400356. [Google Scholar] [CrossRef] [PubMed]
- Rahimzadeh, C.Y.; Barzinjy, A.A.; Mohammed, A.S.; Hamad, S.M. Green Synthesis of SiO2 Nanoparticles from Rhus coriaria L. Extract: Comparison with Chemically Synthesized SiO2 Nanoparticles. PLoS ONE 2022, 17, e0268184. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Sayes, C.M. A Toxicological Profile of Silica Nanoparticles. Toxicol. Res. 2022, 11, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.Y.; Holt, M.G.; Hoet, P.H.M.; Ghosh, M. Neurotoxicity of Four Frequently Used Nanoparticles: A Systematic Review to Reveal the Missing Data. Arch. Toxicol. 2022, 96, 1141–1212. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Son, M.Y.; Choi, M.S.; Kim, S.; Choi, A.Y.; Lee, H.A.; Kim, K.S.; Kim, J.; Song, C.W.; Yoon, S. Integrative Analysis of Genes and miRNA Alterations in Human Embryonic Stem Cells-Derived Neural Cells after Exposure to Silver Nanoparticles. Toxicol. Appl. Pharmacol. 2016, 299, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Zeinabad, H.A.; Zarrabian, A.; Saboury, A.A.; Alizadeh, A.M.O.; Falahati, M. Interaction of Single and Multi Wall Carbon Nanotubes with the Biological Systems: Tau Protein and PC12 Cells as Targets. Sci. Rep. 2016, 6, 26508. [Google Scholar] [CrossRef] [PubMed]
- Kumah, E.A.; Fopa, R.D.; Harati, S.; Boadu, P.; Zohoori, F.V.; Pak, T. Human and Environmental Impacts of Nanoparticles: A Scoping Review of the Current Literature. BMC Public Health 2023, 23, 1059. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Yang, C.C.; Lee, I.T.; Wu, W.B.; Lin, C.C.; Hsiao, L.D.; Yang, C.M. Reactive Oxygen Species-Dependent Activation of EGFR/Akt/P38 Mitogen-Activated Protein Kinase and JNK1/2/FoxO1 and AP-1 Pathways in Human Pulmonary Alveolar Epithelial Cells Leads to Up-Regulation of COX-2/PGE2 Induced by Silica Nanoparticles. Biomedicines 2023, 11, 2628. [Google Scholar] [CrossRef]
- Kundu, J.; Kim, D.H.; Chae, I.G.; Lee, J.K.; Lee, S.; Jeong, C.H.; Chun, K.S. Silicon Dioxide Nanoparticles Induce COX-2 Expression through Activation of STAT3 Signaling Pathway in HaCaT Cells. Toxicol. Vitr. 2018, 52, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Reis, C.G.; Mocelin, R.; Benvenutti, R.; Marcon, M.; Sachett, A.; Herrmann, A.P.; Elisabetsky, E.; Piato, A. Effects of N-Acetylcysteine Amide on Anxiety and Stress Behavior in Zebrafish. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Mocelin, R.; Marcon, M.; D’ambros, S.; Herrmann, A.P.; da Rosa Araujo, A.S.; Piato, A. Behavioral and Biochemical Effects of N-Acetylcysteine in Zebrafish Acutely Exposed to Ethanol. Neurochem. Res. 2018, 43, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Mocelin, R.; Herrmann, A.P.; Marcon, M.; Rambo, C.L.; Rohden, A.; Bevilaqua, F.; De Abreu, M.S.; Zanatta, L.; Elisabetsky, E.; Barcellos, L.J.G.; et al. N-Acetylcysteine Prevents Stress-Induced Anxiety Behavior in Zebrafish. Pharmacol. Biochem. Behav. 2015, 139, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Mocelin, R.; Marcon, M.; da Rosa Araujo, A.S.; Herrmann, A.P.; Piato, A. Withdrawal Effects Following Repeated Ethanol Exposure Are Prevented by N-Acetylcysteine in Zebrafish. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 93, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Mocelin, R.; Marcon, M.; D’ambros, S.; Mattos, J.; Sachett, A.; Siebel, A.M.; Herrmann, A.P.; Piato, A. N-Acetylcysteine Reverses Anxiety and Oxidative Damage Induced by Unpredictable Chronic Stress in Zebrafish. Mol. Neurobiol. 2019, 56, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book. In A Guide for the Laboratory Use of Zebrafish (Danio rerio), 5th ed.; University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- The European Parliament; The Council Of The European Union. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Off. J. Eur. Union 2010, L276, 33–79. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The Arrive Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, J. AVMA Guidelines for the Euthanasia of Animals. Anim. Welf. 2013, 22, 412. [Google Scholar] [CrossRef]
- Li, X.; Liu, B.; Li, X.L.; Li, Y.X.; Sun, M.Z.; Chen, D.Y.; Zhao, X.; Feng, X.Z. SiO2 Nanoparticles Change Colour Preference and Cause Parkinson’s-like Behaviour in Zebrafish. Sci. Rep. 2014, 4, 3810. [Google Scholar] [CrossRef]
- Chitolina, R.; Reis, C.G.; Stahlhofer-Buss, T.; Linazzi, A.; Benvenutti, R.; Marcon, M.; Herrmann, A.P.; Piato, A. Effects of N-Acetylcysteine and Acetyl-l-Carnitine on Acute PTZ-Induced Seizures in Larval and Adult Zebrafish. Pharmacol. Rep. 2023, 75, 1544–1555. [Google Scholar] [CrossRef] [PubMed]
- Avdesh, A.; Martin-Iverson, M.T.; Mondal, A.; Chen, M.; Askraba, S.; Morgan, N.; Lardelli, M.; Groth, D.M.; Verdile, G.; Martins, R.N. Evaluation of Color Preference in Zebrafish for Learning and Memory. J. Alzheimers Dis. 2012, 28, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Bault, Z.A.; Peterson, S.M.; Freeman, J.L. Directional and Color Preference in Adult Zebrafish: Implications in Behavioral and Learning Assays in Neurotoxicology Studies. J. Appl. Toxicol. 2015, 35, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Roy, T.; Suriyampola, P.S.; Flores, J.; López, M.; Hickey, C.; Bhat, A.; Martins, E.P. Color Preferences Affect Learning in Zebrafish, Danio rerio. Sci. Rep. 2019, 9, 14531. [Google Scholar] [CrossRef] [PubMed]
- Jorge, S.; Félix, L.; Costas, B.; Valentim, A.M. Housing Conditions Affect Adult Zebrafish (Danio rerio) Behavior but Not Their Physiological Status. Animals 2023, 13, 1120. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hawkins, R.E.; Brian, M.; Carrell, R.W. The Estimation of Red Cell Superoxide Dismutase Activity. J. Lab. Clin. Med. 1975, 85, 337–341. [Google Scholar] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K. Colorimetric Assay of Catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, K.; Tokumurai, A. Glutathione Peroxidase Activity in Tissues of Vitamin E-Deficient Mice. J. Nutr. Sci. Vitaminol. 1976, 22, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Tagkalidou, N.; Multisanti, C.R.; Bleda, M.J.; Bedrossiantz, J.; Prats, E.; Faggio, C.; Barata, C.; Raldúa, D. Analyzing the Effects of Age, Time of Day, and Experiment on the Basal Locomotor Activity and Light-Off Visual Motor Response Assays in Zebrafish Larvae. Toxics 2024, 12, 349. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Siregar, P.; Chen, J.R.; Lai, Y.H.; Huang, J.C.; Hsiao, C.D. Systematical Exploration of the Common Solvent Toxicity at Whole Organism Level by Behavioral Phenomics in Adult Zebrafish. Environ. Pollut. 2020, 266 Pt 1, 115239. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.T.; Lai, Y.H.; Hsiao, C.D. A Versatile Setup for Measuring Multiple Behavior Endpoints in Zebrafish. Inventions 2018, 3, 75. [Google Scholar] [CrossRef]
- Shen, P.Y.; Wu, J.; Pu, G.; Huang, K.; Lin, Q. Altered Locomotion and Anxiety after Exposure to SiO2 Nanoparticles in Larval Zebrafish. Sci. Rep. 2025, 15, 18229. [Google Scholar] [CrossRef]
- Guo, Y.; Fu, Y.; Sun, W. 50 Hz Magnetic Field Exposure Inhibited Spontaneous Movement of Zebrafish Larvae through ROS-Mediated syn2a Expression. Int. J. Mol. Sci. 2023, 24, 7576. [Google Scholar] [CrossRef] [PubMed]
- Rashidian, G.; Mohammadi-Aloucheh, R.; Hosseinzadeh-Otaghvari, F.; Chupani, L.; Stejskal, V.; Samadikhah, H.; Zamanlui, S.; Multisanti, C.R.; Faggio, C. Long-Term Exposure to Small-Sized Silica Nanoparticles (SiO2-NPs) Induces Oxidative Stress and Impairs Reproductive Performance in Adult Zebrafish (Danio rerio). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2023, 273, 109715. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Cha, C.J. Antibiotic Resistome from the One-Health Perspective: Understanding and Controlling Antimicrobial Resistance Transmission. Exp. Mol. Med. 2021, 53, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.; Smith, C. Innate and Learned Colour Preference in the Zebrafish, Danio rerio. Ethology 2008, 114, 582–588. [Google Scholar] [CrossRef]
- Li, X.; Ji, X.; Wang, R.; Zhao, J.; Dang, J.; Gao, Y.; Jin, M. Zebrafish Behavioral Phenomics Employed for Characterizing Behavioral Neurotoxicity Caused by Silica Nanoparticles. Chemosphere 2020, 240, 124937. [Google Scholar] [CrossRef] [PubMed]
- Book, F.; Backhaus, T. Aquatic Ecotoxicity of Manufactured Silica Nanoparticles: A Systematic Review and Meta-Analysis. Sci. Total Environ. 2022, 806 Pt 4, 150893. [Google Scholar] [CrossRef] [PubMed]
- Kovrižnych, J.A.; Sotníková, R.; Zeljenková, D.; Rollerová, E.; Szabová, E.; Wimmerová, S. Acute Toxicity of 31 Different Nanoparticles to Zebrafish (Danio rerio) Tested in Adulthood and in Early Life Stages: A Comparative Study. Interdiscip. Toxicol. 2013, 6, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, L.; Ma, Y.; Liu, J.; Huang, Y.; Fu, X.; Peng, S.; Wang, X.; Yang, Y.; Zhang, X.; et al. Mechanistic Study of Silica Nanoparticles on the Size-Dependent Retinal Toxicity in Vitro and in Vivo. J. Nanobiotechnol. 2022, 20, 146. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, F.; Climent, M.; Malvindi, M.A.; Pompa, P.P.; Bonetti, P.; Nicassio, F. Delivery of Biologically Active MiR-34a in Normal and Cancer Mammary Epithelial Cells by Synthetic Nanoparticles. Nanomedicine 2019, 19, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Flores-Contreras, E.A.; González-González, R.B.; Pablo Pizaña-Aranda, J.J.; Parra-Arroyo, L.; Rodríguez-Aguayo, A.A.; Iñiguez-Moreno, M.; González-Meza, G.M.; Araújo, R.G.; Ramírez-Gamboa, D.; Parra-Saldívar, R.; et al. Agricultural Waste as a Sustainable Source for Nanoparticle Synthesis and Their Antimicrobial Properties for Food Preservation. Front. Nanotechnol. 2024, 6, 1346069. [Google Scholar] [CrossRef]
- Valadares, L.P.A.; Lima, L.C.O.; Saboia-Morais, S.M.T.; Arantes, T.M.; Cristovan, F.H.; da Silva, N.M.; Andrade, A.B.; Ribeiro, S.A.B.; Alves, B.G.; Virote, B.D.C.R.; et al. Embryotoxicity of Silica Nanoparticles in the Drug Delivery of Domperidone in Zebrafish. Aquat. Toxicol. 2023, 258, 106454. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; He, X.; Yang, X.; Shi, H. Functionalized Silica Nanoparticles: A Platform for Fluorescence Imaging at the Cell and Small Animal Levels. Acc. Chem. Res. 2013, 46, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Son, T.; Kim, M.; Choi, M.; Nam, S.H.; Yoo, A.; Lee, H.; Han, E.H.; Hong, K.S.; Park, H.S. Advancing Fluorescence Imaging: Enhanced Control of Cyanine Dye-Doped Silica Nanoparticles. J. Nanobiotechnol. 2024, 22, 347. [Google Scholar] [CrossRef] [PubMed]
- Ehrhorn, E.G.; Muilenburg, K.M.; Mohs, A.M. Targeting MUC16 for Fluorescence-Guided Surgery in Pancreatic Cancer. In Proceedings of the Visualizing and Quantifying Drug Distribution in Tissue IX, San Francisco, CA, USA, 25–31 January 2025; Evans, C.L., Chan, K.F., Eds.; SPIE: Bellingham, WA, USA, 2025; p. 7. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rarinca, V.; Gurzu, I.-L.; Nicoara, M.N.; Ciobica, A.; Visternicu, M.; Ionescu, C.; Balmus, I.M.; Plavan, G.-I.; Todirascu-Ciornea, E.; Gurzu, B. Neurobehavioral and Oxidative Stress Effects of SiO2 Nanoparticles in Zebrafish and the Protective Role of N-Acetylcysteine. Biomedicines 2025, 13, 1762. https://doi.org/10.3390/biomedicines13071762
Rarinca V, Gurzu I-L, Nicoara MN, Ciobica A, Visternicu M, Ionescu C, Balmus IM, Plavan G-I, Todirascu-Ciornea E, Gurzu B. Neurobehavioral and Oxidative Stress Effects of SiO2 Nanoparticles in Zebrafish and the Protective Role of N-Acetylcysteine. Biomedicines. 2025; 13(7):1762. https://doi.org/10.3390/biomedicines13071762
Chicago/Turabian StyleRarinca, Viorica, Irina-Luciana Gurzu, Mircea Nicusor Nicoara, Alin Ciobica, Malina Visternicu, Catalina Ionescu, Ioana Miruna Balmus, Gabriel-Ionut Plavan, Elena Todirascu-Ciornea, and Bogdan Gurzu. 2025. "Neurobehavioral and Oxidative Stress Effects of SiO2 Nanoparticles in Zebrafish and the Protective Role of N-Acetylcysteine" Biomedicines 13, no. 7: 1762. https://doi.org/10.3390/biomedicines13071762
APA StyleRarinca, V., Gurzu, I.-L., Nicoara, M. N., Ciobica, A., Visternicu, M., Ionescu, C., Balmus, I. M., Plavan, G.-I., Todirascu-Ciornea, E., & Gurzu, B. (2025). Neurobehavioral and Oxidative Stress Effects of SiO2 Nanoparticles in Zebrafish and the Protective Role of N-Acetylcysteine. Biomedicines, 13(7), 1762. https://doi.org/10.3390/biomedicines13071762