

The Causal Relationship Between Gut Microbiomes, Inflammatory Mediators, and Traumatic Brain Injury in Europeans: Evidence from Genetic Correlation and Functional Mapping Annotation Analyses

 ,
,
Abstract
1. Introduction
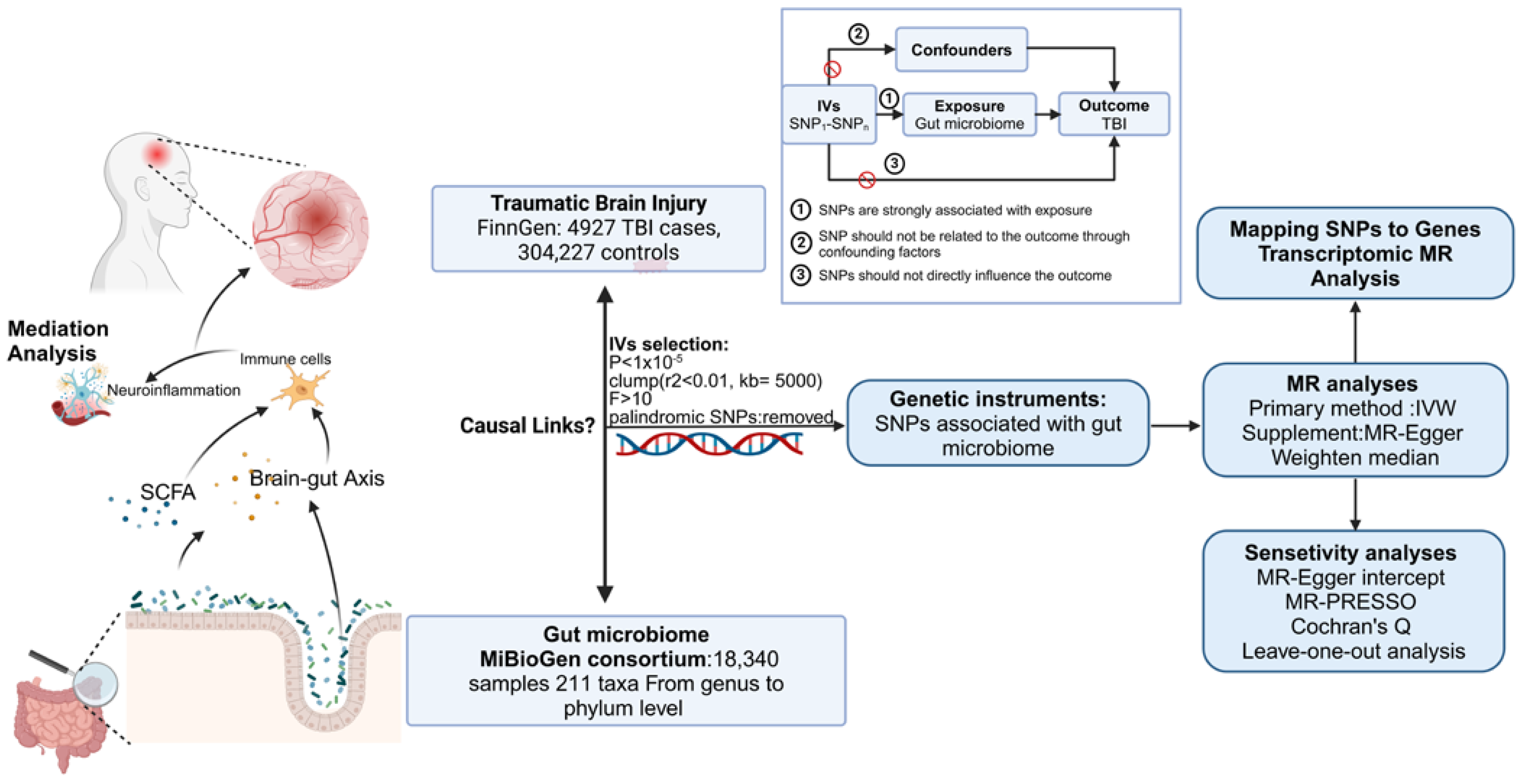
2. Materials and Methods
2.1. Study Design
2.2. Data Sources
2.3. Identification of IVs
2.4. Statistical Analyses
2.5. Mediation Analysis
2.6. Mapping SNPs to Genes and Transcriptomic MR Analysis
2.7. Ethical Approval
3. Results
3.1. Details of GMs
3.2. MR Estimates
3.3. Sensitivity Analysis and Reverse MR Analysis
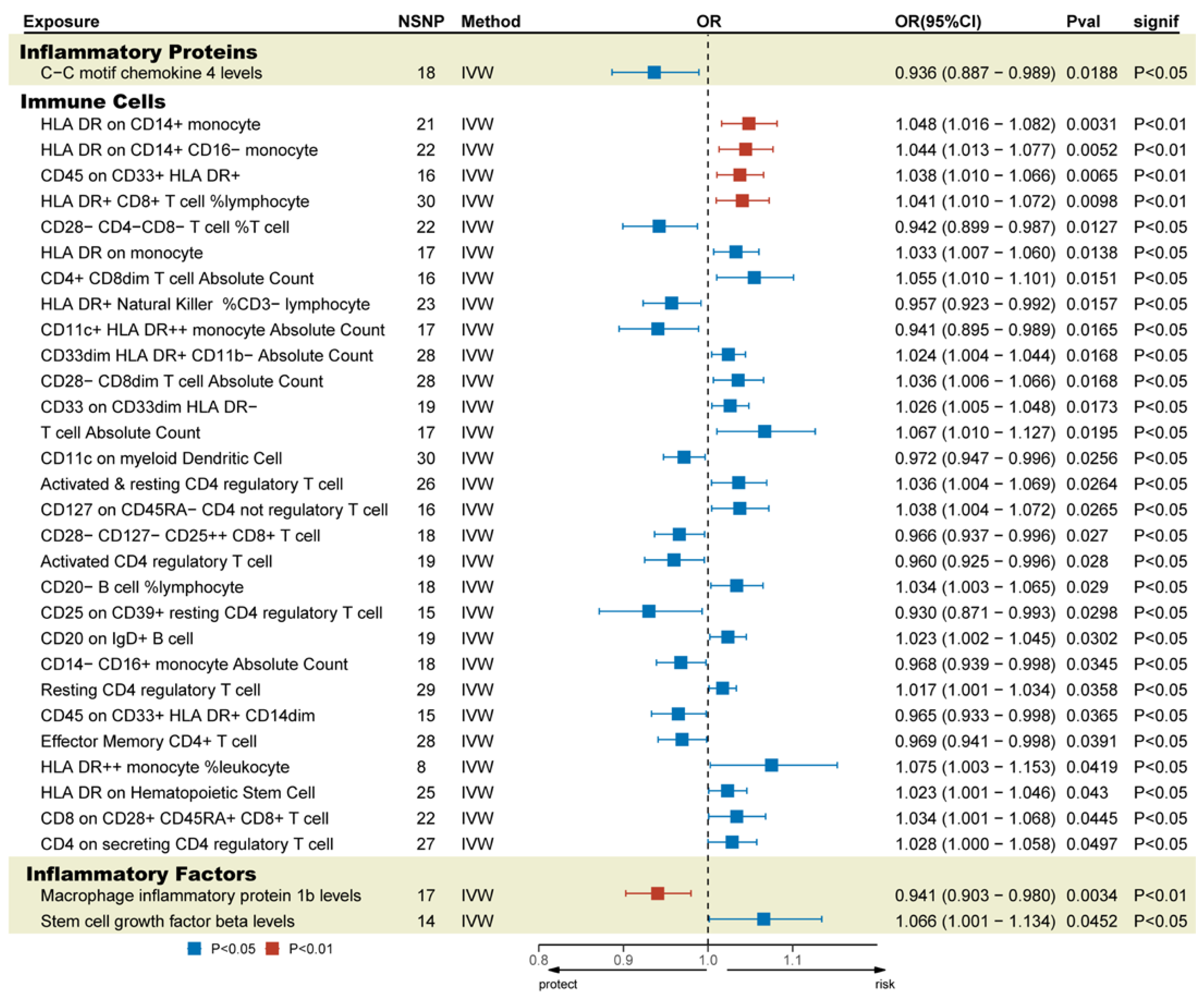
3.4. Mediation Analysis
3.5. Mapping SNPs to Genes and Transcriptomic MR Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TBI | Traumatic brain injury |
| MR | Mendelian randomization |
| GM | Gut microbiome |
| GWAS | Genome-Wide Association Study |
| IVW | Inverse-variance-weighted |
| WM | Weighted median |
| OR | Odds ratios |
| CI | Confidence interval |
| LD | Linkage disequilibrium |
References
- Jiang, J.Y.; Gao, G.Y.; Feng, J.F.; Mao, Q.; Chen, L.G.; Yang, X.F.; Liu, J.F.; Wang, Y.H.; Qiu, B.H.; Huang, X.J. Traumatic brain injury in China. Lancet Neurol. 2019, 18, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Badhiwala, J.H.; Wilson, J.R.; Fehlings, M.G. Global burden of traumatic brain and spinal cord injury. Lancet Neurol. 2019, 18, 24–25. [Google Scholar] [CrossRef] [PubMed]
- Dams-O’Connor, K.; Juengst, S.B.; Bogner, J.; Chiaravalloti, N.D.; Corrigan, J.D.; Giacino, J.T.; Harrison-Felix, C.L.; Hoffman, J.M.; Ketchum, J.M.; Lequerica, A.H.; et al. Traumatic brain injury as a chronic disease: Insights from the United States Traumatic Brain Injury Model Systems Research Program. Lancet Neurol. 2023, 22, 517–528. [Google Scholar] [CrossRef]
- Wilson, L.; Stewart, W.; Dams-O’Connor, K.; Diaz-Arrastia, R.; Horton, L.; Menon, D.K.; Polinder, S. The chronic and evolving neurological consequences of traumatic brain injury. Lancet Neurol. 2017, 16, 813–825. [Google Scholar] [CrossRef]
- Te Ao, B.; Brown, P.; Tobias, M.; Ameratunga, S.; Barker-Collo, S.; Theadom, A.; McPherson, K.; Starkey, N.; Dowell, A.; Jones, K.; et al. Cost of traumatic brain injury in New Zealand: Evidence from a population-based study. Neurology 2014, 83, 1645–1652. [Google Scholar] [CrossRef]
- Bowman, K.; Matney, C.; Berwick, D.M. Improving Traumatic Brain Injury Care and Research: A Report From the National Academies of Sciences, Engineering, and Medicine. JAMA 2022, 327, 419–420. [Google Scholar] [CrossRef]
- Järbrink-Sehgal, E.; Andreasson, A. The gut microbiota and mental health in adults. Curr. Opin. Neurobiol. 2020, 62, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut microbes and the brain: Paradigm shift in neuroscience. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef]
- Hanscom, M.; Loane, D.J.; Shea-Donohue, T. Brain-gut axis dysfunction in the pathogenesis of traumatic brain injury. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Houlden, A.; Goldrick, M.; Brough, D.; Vizi, E.S.; Lénárt, N.; Martinecz, B.; Roberts, I.S.; Denes, A. Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain Behav. Immun. 2016, 57, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Wang, S.; Wu, C.; Cao, Y.; Gu, Q.; Zhu, Y.; Zhang, W.; Hu, W. Gut Microbiota Dysbiosis after Traumatic Brain Injury Contributes to Persistent Microglial Activation Associated with Upregulated Lyz2 and Shifted Tryptophan Metabolic Phenotype. Nutrients 2022, 14, 3467. [Google Scholar] [CrossRef]
- Puntambekar, S.S.; Saber, M.; Lamb, B.T.; Kokiko-Cochran, O.N. Cellular players that shape evolving pathology and neurodegeneration following traumatic brain injury. Brain Behav. Immun. 2018, 71, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhu, J.; Ju, L.; Sun, L.; Tse, L.A.; Kinra, S.; Li, Y. Osteoarthritis & stroke: A bidirectional mendelian randomization study. Osteoarthr. Cartil. 2022, 30, 1390–1397. [Google Scholar] [CrossRef]
- Seyedsalehi, A.; Warrier, V.; Bethlehem, R.A.I.; Perry, B.I.; Burgess, S.; Murray, G.K. Educational attainment, structural brain reserve and Alzheimer’s disease: A Mendelian randomization analysis. Brain 2023, 146, 2059–2074. [Google Scholar] [CrossRef]
- Liu, X.; Miao, Y.; Liu, C.; Lu, W.; Feng, Q.; Zhang, Q. Identification of multiple novel susceptibility genes associated with autoimmune thyroid disease. Front. Immunol. 2023, 14, 1161311. [Google Scholar] [CrossRef]
- Dimou, N.L.; Tsilidis, K.K. A Primer in Mendelian Randomization Methodology with a Focus on Utilizing Published Summary Association Data. Methods Mol. Biol. 2018, 1793, 211–230. [Google Scholar] [CrossRef]
- Skrivankova, V.W.; Richmond, R.C.; Woolf, B.A.R.; Yarmolinsky, J.; Davies, N.M.; Swanson, S.A.; VanderWeele, T.J.; Higgins, J.P.T.; Timpson, N.J.; Dimou, N.; et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: The STROBE-MR Statement. JAMA 2021, 326, 1614–1621. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Raygoza Garay, J.A.; Finnicum, C.T.; Liu, X.; et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef]
- Burgess, S.; Scott, R.A.; Timpson, N.J.; Davey Smith, G.; Thompson, S.G. Using published data in Mendelian randomization: A blueprint for efficient identification of causal risk factors. Eur. J. Epidemiol. 2015, 30, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef]
- Xiang, K.; Wang, P.; Xu, Z.; Hu, Y.Q.; He, Y.S.; Chen, Y.; Feng, Y.T.; Yin, K.J.; Huang, J.X.; Wang, J.; et al. Causal Effects of Gut Microbiome on Systemic Lupus Erythematosus: A Two-Sample Mendelian Randomization Study. Front. Immunol. 2021, 12, 667097. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Taskesen, E.; van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef]
- Võsa, U.; Claringbould, A.; Westra, H.J.; Bonder, M.J.; Deelen, P.; Zeng, B.; Kirsten, H.; Saha, A.; Kreuzhuber, R.; Yazar, S.; et al. Large-scale cis- and trans-eQTL analyses identify thousands of genetic loci and polygenic scores that regulate blood gene expression. Nat. Genet. 2021, 53, 1300–1310. [Google Scholar] [CrossRef] [PubMed]
- Brenner, L.A.; Stearns-Yoder, K.A.; Hoffberg, A.S.; Penzenik, M.E.; Starosta, A.J.; Hernández, T.D.; Hadidi, D.A.; Lowry, C.A. Growing literature but limited evidence: A systematic review regarding prebiotic and probiotic interventions for those with traumatic brain injury and/or posttraumatic stress disorder. Brain Behav. Immun. 2017, 65, 57–67. [Google Scholar] [CrossRef]
- Nicholson, S.E.; Watts, L.T.; Burmeister, D.M.; Merrill, D.; Scroggins, S.; Zou, Y.; Lai, Z.; Grandhi, R.; Lewis, A.M.; Newton, L.M.; et al. Moderate Traumatic Brain Injury Alters the Gastrointestinal Microbiome in a Time-Dependent Manner. Shock 2019, 52, 240–248. [Google Scholar] [CrossRef]
- Wang, S.; Shang, Y.; Pi, Z.; Zhou, Z.; Zhang, X.; Ren, L.; Liang, W.; Guo, Y.; Cai, J.; Zhang, C. Temporal Changes of the Oral and Fecal Microbiota after Mild Traumatic Brain Injury in Rats by 16S rRNA Sequencing. Microorganisms 2023, 11, 1452. [Google Scholar] [CrossRef]
- Urban, R.J.; Pyles, R.B.; Stewart, C.J.; Ajami, N.; Randolph, K.M.; Durham, W.J.; Danesi, C.P.; Dillon, E.L.; Summons, J.R.; Singh, C.K.; et al. Altered Fecal Microbiome Years after Traumatic Brain Injury. J. Neurotrauma 2020, 37, 1037–1051. [Google Scholar] [CrossRef]
- Yuan, B.; Lu, X.J.; Wu, Q. Gut Microbiota and Acute Central Nervous System Injury: A New Target for Therapeutic Intervention. Front. Immunol. 2021, 12, 800796. [Google Scholar] [CrossRef]
- McAllister, T.W. Neurobiological consequences of traumatic brain injury. Dialogues Clin. Neurosci. 2011, 13, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Makinde, H.M.; Cuda, C.M.; Just, T.B.; Perlman, H.R.; Schwulst, S.J. Nonclassical Monocytes Mediate Secondary Injury, Neurocognitive Outcome, and Neutrophil Infiltration after Traumatic Brain Injury. J. Immunol. 2017, 199, 3583–3591. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, J.; Zhang, H.; Li, Y.; Wen, L.; Tan, X.; Cheng, K.; Liu, Y.; Pu, J.; Liu, L.; et al. The gut microbiome modulates the transformation of microglial subtypes. Mol. Psychiatry 2023, 28, 1611–1621. [Google Scholar] [CrossRef]
- Panther, E.J.; Dodd, W.; Clark, A.; Lucke-Wold, B. Gastrointestinal Microbiome and Neurologic Injury. Biomedicines 2022, 10, 500. [Google Scholar] [CrossRef]
- Treangen, T.J.; Wagner, J.; Burns, M.P.; Villapol, S. Traumatic Brain Injury in Mice Induces Acute Bacterial Dysbiosis Within the Fecal Microbiome. Front. Immunol. 2018, 9, 2757. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.B.; Firek, B.; Shi, M.; Yeh, A.; Brower-Sinning, R.; Aveson, V.; Kohl, B.L.; Fabio, A.; Carcillo, J.A.; Morowitz, M.J. Disruption of the microbiota across multiple body sites in critically ill children. Microbiome 2016, 4, 66. [Google Scholar] [CrossRef]
- Opeyemi, O.M.; Rogers, M.B.; Firek, B.A.; Janesko-Feldman, K.; Vagni, V.; Mullett, S.J.; Wendell, S.G.; Nelson, B.P.; New, L.A.; Mariño, E.; et al. Sustained Dysbiosis and Decreased Fecal Short-Chain Fatty Acids after Traumatic Brain Injury and Impact on Neurologic Outcome. J. Neurotrauma 2021, 38, 2610–2621. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Abdel-Haq, R.; Schlachetzki, J.C.M.; Glass, C.K.; Mazmanian, S.K. Microbiome-microglia connections via the gut-brain axis. J. Exp. Med. 2019, 216, 41–59. [Google Scholar] [CrossRef]
- Hoegenauer, C.; Hammer, H.F.; Mahnert, A.; Moissl-Eichinger, C. Methanogenic archaea in the human gastrointestinal tract. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 805–813. [Google Scholar] [CrossRef]
- Kumpitsch, C.; Fischmeister, F.P.S.; Mahnert, A.; Lackner, S.; Wilding, M.; Sturm, C.; Springer, A.; Madl, T.; Holasek, S.; Högenauer, C.; et al. Reduced B12 uptake and increased gastrointestinal formate are associated with archaeome-mediated breath methane emission in humans. Microbiome 2021, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Boros, M.; Ghyczy, M.; Érces, D.; Varga, G.; Tőkés, T.; Kupai, K.; Torday, C.; Kaszaki, J. The anti-inflammatory effects of methane. Crit. Care Med. 2012, 40, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Boros, M.; Keppler, F. Methane Production and Bioactivity-A Link to Oxido-Reductive Stress. Front. Physiol. 2019, 10, 1244. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.; Remes, C.; Shiriaev, D.I.; Liu, Y.; Spåhr, H.; Wibom, R.; Atanassov, I.; Nguyen, M.D.; Cooperman, B.S.; Rorbach, J. Human mitochondria require mtRF1 for translation termination at non-canonical stop codons. Nat. Commun. 2023, 14, 30. [Google Scholar] [CrossRef]
- Chen, J.; Bacanu, S.A.; Yu, H.; Zhao, Z.; Jia, P.; Kendler, K.S.; Kranzler, H.R.; Gelernter, J.; Farrer, L.; Minica, C.; et al. Genetic Relationship between Schizophrenia and Nicotine Dependence. Sci. Rep. 2016, 6, 25671. [Google Scholar] [CrossRef]
- Chambers, J.W.; Cherry, L.; Laughlin, J.D.; Figuera-Losada, M.; Lograsso, P.V. Selective inhibition of mitochondrial JNK signaling achieved using peptide mimicry of the Sab kinase interacting motif-1 (KIM1). ACS Chem. Biol. 2011, 6, 808–818. [Google Scholar] [CrossRef]
- Sodero, A.O.; Rodriguez-Silva, M.; Salio, C.; Sassoè-Pognetto, M.; Chambers, J.W. Sab is differentially expressed in the brain and affects neuronal activity. Brain Res. 2017, 1670, 76–85. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure | Outcome | Method | No. SNP | p-Value | SE | OR (95%CI) | Cochran’s Q Test | |
|---|---|---|---|---|---|---|---|---|
| Q | Q p-Value | |||||||
| class Methanobacteria | TBI | IVW | 10 | <0.001 | 0.045 | 1.175 (1.077, 1.282) | 5.114 | 0.745 |
| family Methanobacteriaceae | TBI | IVW | 10 | <0.001 | 0.045 | 1.175 (1.077, 1.282) | 5.114 | 0.745 |
| genus Dorea | TBI | IVW | 10 | 0.034 | 0.111 | 0.791 (0.637, 0.983) | 11.858 | 0.158 |
| genus Eubacterium fissicatena group | TBI | IVW | 9 | 0.006 | 0.049 | 1.143 (1.038, 1.258) | 3.585 | 0.826 |
| genus Eubacterium hallii group | TBI | IVW | 14 | 0.024 | 0.068 | 0.858 (0.751, 0.980) | 11.152 | 0.516 |
| genus Family XIII AD3011 group | TBI | IVW | 13 | 0.007 | 0.074 | 1.222 (1.057, 1.412) | 11.241 | 0.423 |
| genus Gordonibacter | TBI | IVW | 11 | 0.046 | 0.041 | 0.920 (0.849, 0.998) | 8.750 | 0.461 |
| genus Ruminococcaceae UCG004 | TBI | IVW | 11 | 0.002 | 0.066 | 0.816 (0.718, 0.928) | 4.931 | 0.840 |
| order Methanobacteriales | TBI | IVW | 10 | <0.001 | 0.045 | 1.175 (1.077, 1.282) | 5.114 | 0.745 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, B.; Qiu, Y.; Wang, Z.; Tao, Y.; Wang, M.; Duan, A.; Xie, M.; Yin, Z.; Chen, Z.; Ma, C.; et al. The Causal Relationship Between Gut Microbiomes, Inflammatory Mediators, and Traumatic Brain Injury in Europeans: Evidence from Genetic Correlation and Functional Mapping Annotation Analyses. Biomedicines 2025, 13, 753. https://doi.org/10.3390/biomedicines13030753
Song B, Qiu Y, Wang Z, Tao Y, Wang M, Duan A, Xie M, Yin Z, Chen Z, Ma C, et al. The Causal Relationship Between Gut Microbiomes, Inflammatory Mediators, and Traumatic Brain Injury in Europeans: Evidence from Genetic Correlation and Functional Mapping Annotation Analyses. Biomedicines. 2025; 13(3):753. https://doi.org/10.3390/biomedicines13030753
Chicago/Turabian StyleSong, Bingyi, Youjia Qiu, Zilan Wang, Yuchen Tao, Menghan Wang, Aojie Duan, Minjia Xie, Ziqian Yin, Zhouqing Chen, Chao Ma, and et al. 2025. "The Causal Relationship Between Gut Microbiomes, Inflammatory Mediators, and Traumatic Brain Injury in Europeans: Evidence from Genetic Correlation and Functional Mapping Annotation Analyses" Biomedicines 13, no. 3: 753. https://doi.org/10.3390/biomedicines13030753
APA StyleSong, B., Qiu, Y., Wang, Z., Tao, Y., Wang, M., Duan, A., Xie, M., Yin, Z., Chen, Z., Ma, C., & Wang, Z. (2025). The Causal Relationship Between Gut Microbiomes, Inflammatory Mediators, and Traumatic Brain Injury in Europeans: Evidence from Genetic Correlation and Functional Mapping Annotation Analyses. Biomedicines, 13(3), 753. https://doi.org/10.3390/biomedicines13030753