

Cimetidine Attenuates Therapeutic Effect of Anti-PD-1 and Anti-PD-L1 and Modulates Tumor Microenvironment in Colon Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. MTT Assay
2.3. Clonogenic Assay
2.4. Cell Cycle Analysis
2.5. H2R, PD-1, and PD-L1 Expression Profile
2.6. Syngeneic Tumor Implantation Model
2.7. Evaluation of Tumor Volume and Toxicity
2.8. Flow Cytometry Analysis of the Immune Cells
2.9. Statistical Analysis
3. Results
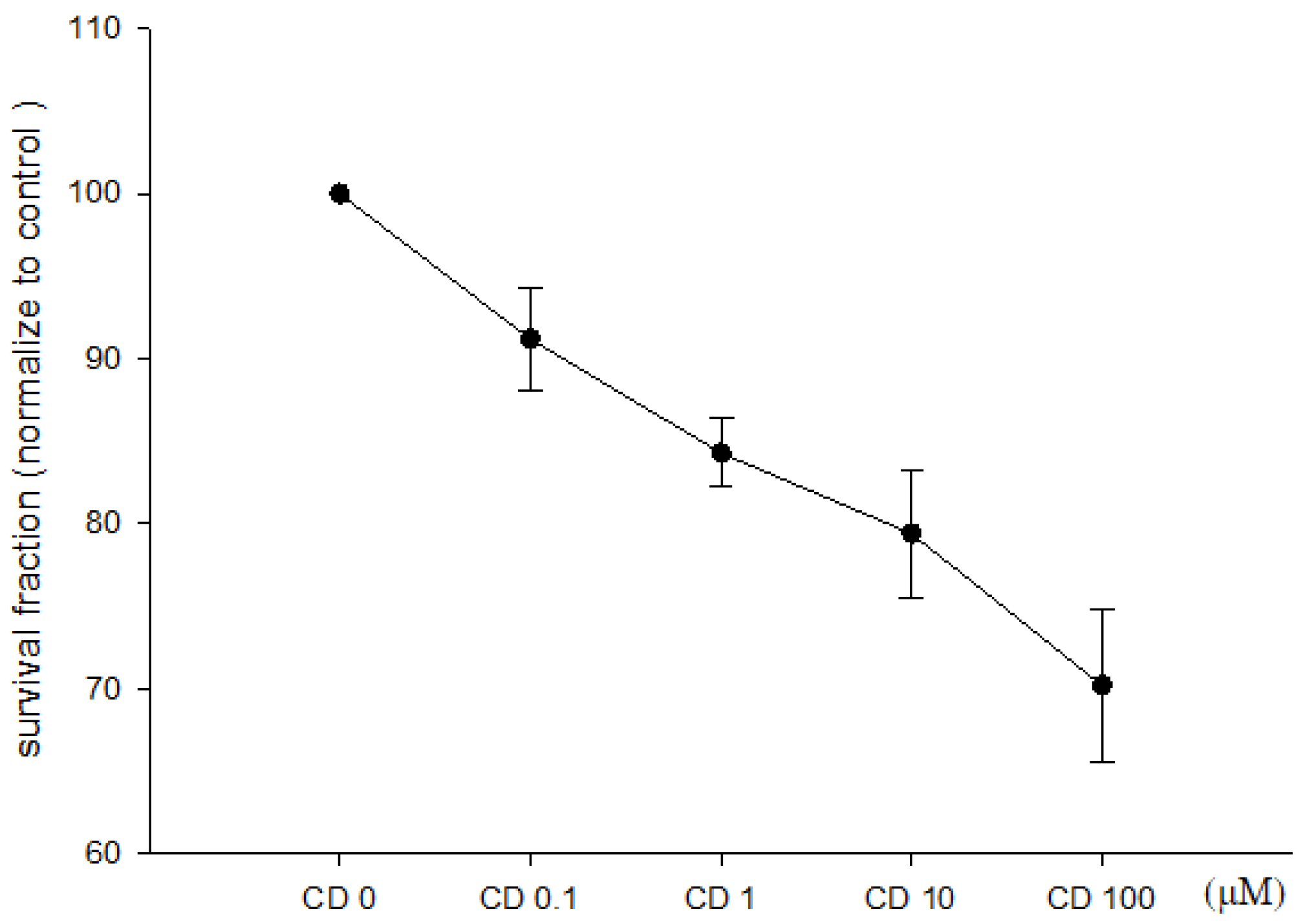
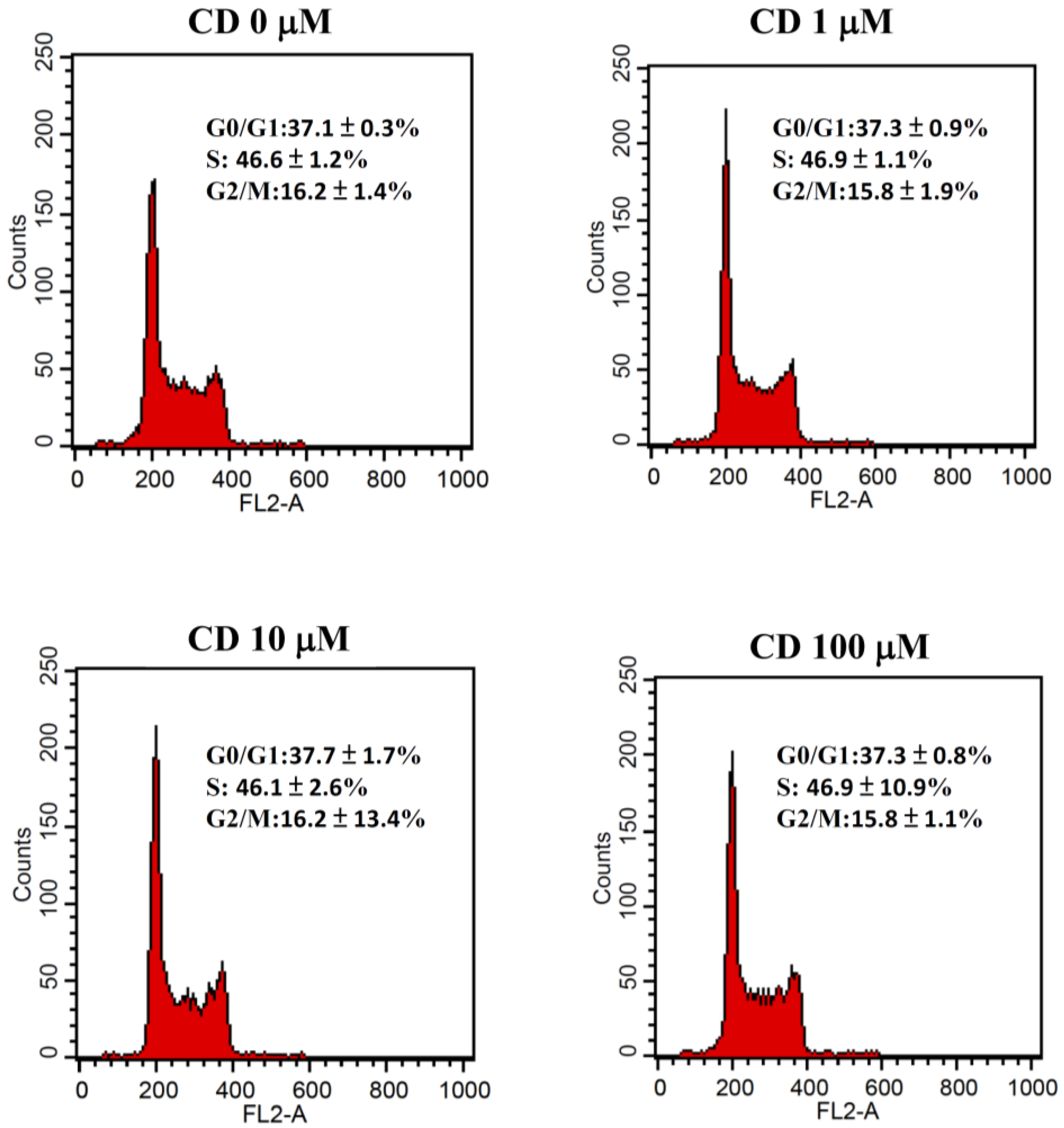
3.1. Effect of Cimetidine on Cell Viability, Clonogenicity, and Cell Cycle Distribution
3.2. Effect of Cimetidine on Expression of Surface H2R and PD-L1
3.3. Effect of Cimetidine and Anti-PD-1/Anti-PD-L1 on CT26 Tumor Growth and Toxicity
3.4. Effect of Cimetidine and Anti-PD-1/Anti-PD-L1 on Immune Cells Expression in Spleen and Tumor Microenvironment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Wagle, N.S.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 233–254. [Google Scholar] [CrossRef]
- Lochhead, P.; Kuchiba, A.; Imamura, Y.; Liao, X.; Yamauchi, M.; Nishihara, R.; Qian, Z.R.; Morikawa, T.; Shen, J.; Meyerhardt, J.A.; et al. Microsatellite instability and BRAF mutation testing in colorectal cancer prognostication. J. Natl. Cancer Inst. 2013, 105, 1151–1156. [Google Scholar] [CrossRef]
- Evrard, C.; Tachon, G.; Randrian, V.; Karayan-Tapon, L.; Tougeron, D. Microsatellite instability: Diagnosis, heterogeneity, discordance, and clinical impact in colorectal cancer. Cancers 2019, 11, 1567. [Google Scholar] [CrossRef]
- Overman, M.J.; Lonardi, S.; Wong, K.Y.M.; Lenz, H.J.; Gelsomino, F.; Aglietta, M.; Morse, M.A.; Van Cutsem, E.; McDermott, R.; Hill, A.; et al. Durable clinical benefit with nivolumab plus ipilimumab in DNA mismatch repair-deficient/microsatellite instability-high metastatic colorectal cancer. J. Clin. Oncol. 2018, 36, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Kim, T.W.; Van Cutsem, E.; Geva, R.; Jäger, D.; Hara, H.; Burge, M.; O’Neil, B.; Kavan, P.; Yoshino, T.; et al. Phase II open-label study of pembrolizumab in treatment-refractory, microsatellite instability-high/mismatch repair-deficient metastatic colorectal cancer: KEYNOTE-164. J. Clin. Oncol. 2020, 38, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Uram, J.N.; Wang, H.; Bartlett, B.R.; Kemberling, H.; Eyring, A.D.; Skora, A.D.; Luber, B.S.; Azad, N.S.; Laheru, D.; et al. PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. N. Engl. J. Med. 2015, 372, 2509–2520. [Google Scholar] [CrossRef]
- Mandal, R.; Samstein, R.M.; Lee, K.W.; Havel, J.J.; Wang, H.; Krishna, C.; Sabio, E.Y.; Makarov, V.; Kuo, F.; Blecua, P.; et al. Genetic diversity of tumors with mismatch repair deficiency influences anti-PD-1 immunotherapy response. Science 2019, 364, 485–491. [Google Scholar] [CrossRef] [PubMed]
- de Visser, K.E.; Joyce, J.A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell 2023, 41, 374–403. [Google Scholar] [CrossRef] [PubMed]
- Tie, Y.; Tang, F.; Wei, Y.Q.; Wei, X.W. Immunosuppressive cells in cancer: Mechanisms and potential therapeutic targets. J. Hematol. Oncol. 2022, 15, 61. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Liu, Y.; Ali, N.M.; Zhang, B.; Cui, X. The role of innate immune cells in the tumor microenvironment and research progress in anti-tumor therapy. Front. Immunol. 2022, 13, 1039260. [Google Scholar] [CrossRef]
- Feldman, M.; Burton, M.E. Histamine2-receptor antagonists. Standard therapy for acid-peptic diseases. Part II. N. Engl. J. Med. 1990, 323, 1749–1755. [Google Scholar] [PubMed]
- Feldman, M.; Burton, M.E. Histamine2-receptor antagonists. Standard therapy for acid-peptic diseases. Part I. N. Engl. J. Med. 1990, 323, 1672–1680. [Google Scholar] [PubMed]
- Kelly, M.D.; King, J.; Cherian, M.; Dwerryhouse, S.J.; Finlay, I.G.; Adams, W.J.; King, D.W.; Lubowski, D.Z.; Morris, D.L. Randomized trial of preoperative cimetidine in patients with colorectal carcinoma with quantitative assessment of tumor-associated lymphocytes. Cancer 1999, 85, 1658–1663. [Google Scholar] [CrossRef]
- Tønnesen, H.; Knigge, U.; Bülow, S.; Damm, P.; Fischerman, K.; Hesselfeldt, P.; Hjortrup, A.; Pedersen, I.K.; Pedersen, V.M.; Siemssen, O.J. Effect of cimetidine on survival after gastric cancer. Lancet 1988, 2, 990–992. [Google Scholar] [CrossRef]
- Dexeus, F.H.; Logothetis, C.J.; Sella, A.; Fitz, K.; Amato, R.; Reuben, J.M.; Dozier, N. Phase II study of coumarin and cimetidine in patients with metastatic renal cell carcinoma. J. Clin. Oncol. 1990, 8, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.F.; Creagan, E.T.; Cullinan, S.A.; Mailliard, J.A.; Ebbert, L.; Veeder, M.H.; Chang, M. Phase II studies of single-agent cimetidine and the combination N-phosphonacetyl-L-aspartate (NSC-224131) plus L-alanosine (NSC-153353) in advanced malignant melanoma. J. Clin. Oncol. 1987, 5, 1078–1082. [Google Scholar] [CrossRef]
- Aogi, K.; Takeuchi, H.; Saeki, T.; Aiba, K.; Tamura, K.; Iino, K.; Imamura, C.K.; Okita, K.; Kagami, Y.; Tanaka, R.; et al. Optimizing antiemetic treatment for chemotherapy-induced nausea and vomiting in Japan: Update summary of the 2015 Japan Society of Clinical Oncology Clinical Practice Guidelines for antiemesis. Int. J. Clin. Oncol. 2021, 26, 1–17. [Google Scholar] [CrossRef]
- Li, Y.; Yang, G.L.; Yuan, H.Y.; Bai, D.J.; Wang, K.; Lin, C.R.; Hu, M.B.; Feng, M.H. Effects of perioperative cimetidine administration on peripheral blood lymphocytes and tumor infiltrating lymphocytes in patients with gastrointestinal cancer: Results of a randomized controlled clinical trial. Hepato-Gastroenterol. 2005, 52, 504–508. [Google Scholar]
- O’Mahony, L.; Akdis, M.; Akdis, C.A. Regulation of the immune response and inflammation by histamine and histamine receptors. J. Allergy Clin. Immunol. 2011, 128, 1153–1162. [Google Scholar] [CrossRef]
- Van der Leek, A.P.; Yanishevsky, Y.; Kozyrskyj, A.L. The kynurenine pathway as a novel link between allergy and the gut microbiome. Front. Immunol. 2017, 8, 1374. [Google Scholar] [CrossRef]
- Vaziri, P.; Dang, K.; Anderson, G.H. Evidence for histamine involvement in the effect of histidine loads on food and water intake in rats. J. Nutr. 1997, 127, 1519–1526. [Google Scholar] [CrossRef]
- Zeitzer, J.M. Control of sleep and wakefulness in health and disease. Prog. Mol. Biol. Transl. Sci. 2013, 119, 137–154. [Google Scholar]
- Savitz, J. The kynurenine pathway: A finger in every pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef]
- Adams, W.J.; Lawson, J.A.; Morris, D.L. Cimetidine inhibits in vivo growth of human colon cancer and reverses histamine stimulated in vitro and in vivo growth. Gut 1994, 35, 1632–1636. [Google Scholar] [CrossRef]
- Garcia-Caballero, M.; Neugebauer, E.; Rodriguez, F.; Nuñez de Castro, I.; Vara-Thorbeck, C. Histamine synthesis and content in benign and malignant breast tumours. Its effects on other host tissues. Surg. Oncol. 1994, 3, 167–173. [Google Scholar] [CrossRef]
- Kobayashi, K.; Matsumoto, S.; Morishima, T.; Kawabe, T.; Okamoto, T. Cimetidine inhibits cancer cell adhesion to endothelial cells and prevents metastasis by blocking E-selectin expression. Cancer Res. 2000, 60, 3978–3984. [Google Scholar]
- Tomita, K.; Izumi, K.; Okabe, S. Roxatidine- and cimetidine-induced angiogenesis inhibition suppresses growth of colon cancer implants in syngeneic mice. J. Pharmacol. Sci. 2003, 93, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Chihara, Y.; Fujimoto, K.; Miyake, M.; Hiasa, Y.; Hirao, Y. Anti-tumor effect of cimetidine via inhibiting angiogenesis factors in N-butyl-N-(4-hydroxybutyl) nitrosamine-induced mouse and rat bladder carcinogenesis. Oncol. Rep. 2009, 22, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Sabesin, S.M. Safety issues relating to long-term treatment with histamine H2-receptor antagonists. Aliment. Pharmacol. Ther. 1993, 7 (Suppl. 2), 35–40. [Google Scholar] [CrossRef]
- Kubota, T.; Fujiwara, H.; Ueda, Y.; Itoh, T.; Yamashita, T.; Yoshimura, T.; Okugawa, K.; Yamamoto, Y.; Yano, Y.; Yamagishi, H. Cimetidine modulates the antigen presenting capacity of dendritic cells from colorectal cancer patients. Br. J. Cancer 2002, 86, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Katoh, J.; Tsuchiya, K.; Osawa, H.; Sato, W.; Matsumura, G.; Iida, Y.; Suzuki, S.; Hosaka, S.; Yoshii, S.; Tada, Y. Cimetidine reduces impairment of cellular immunity after cardiac operations with cardiopulmonary bypass. J. Thorac. Cardiovasc. Surg. 1998, 116, 312–318. [Google Scholar] [CrossRef][Green Version]
- Li, B.; Cao, F.; Zhu, Q.; Li, B.; Gan, M.; Wang, D. Perioperative cimetidine administration improves systematic immune response and tumor infiltrating lymphocytes in patients with colorectal cancer. Hepato-Gastroenterol. 2013, 60, 244–247. [Google Scholar] [CrossRef]
- Martin, R.K.; Saleem, S.J.; Folgosa, L.; Zellner, H.B.; Damle, S.R.; Nguyen, G.K.; Ryan, J.J.; Bear, H.D.; Irani, A.M.; Conrad, D.H. Mast cell histamine promotes the immunoregulatory activity of myeloid-derived suppressor cells. J. Leukoc. Biol. 2014, 96, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Yang, G.; Yuan, H.; Li, Y.; Wang, K.; Shao, H. Perioperative cimetidine application modulates natural killer cells in patients with colorectal cancer: A randomized clinical study. J. Tongji Med. Univ. 1999, 19, 300–303. [Google Scholar] [PubMed]
- Takahashi, K.; Tanaka, S.; Ichikawa, A. Effect of cimetidine on intratumoral cytokine expression in an experimental tumor. Biochem. Biophys. Res. Commun. 2001, 281, 1113–1119. [Google Scholar] [CrossRef]
- Tanaka, T.; Kochi, T.; Shirakami, Y.; Mori, T.; Kurata, A.; Watanabe, N.; Moriwaki, H.; Shimizu, M. Cimetidine and clobenpropit attenuate inflammation-associated colorectal carcinogenesis in male ICR mice. Cancers 2016, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Castle, J.C.; Loewer, M.; Boegel, S.; de Graaf, J.; Bender, C.; Tadmor, A.D.; Boisguerin, V.; Bukur, T.; Sorn, P.; Paret, C.; et al. Immunomic, genomic and transcriptomic characterization of CT26 colorectal carcinoma. BMC Genom. 2014, 15, 190. [Google Scholar] [CrossRef]
- Wen, Y.; Ye, S.; Li, Z.; Zhang, X.; Liu, C.; Wu, Y.; Zheng, R.; Xu, C.; Tian, J.; Shu, L.; et al. HDAC6 inhibitor ACY-1215 enhances STAT1 acetylation to block PD-L1 for colorectal cancer immunotherapy. Cancer Immunol. Immunother. 2024, 73, 7. [Google Scholar] [CrossRef]
- Sharma, P.; Hu-Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, adaptive, and acquired resistance to cancer immunotherapy. Cell 2017, 168, 707–723. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, F.-C.; Lai, J.C.-Y.; Shieh, H.-R.; Liou, W.-Z.; Bair, M.-J.; Chen, Y.-J. Cimetidine Attenuates Therapeutic Effect of Anti-PD-1 and Anti-PD-L1 and Modulates Tumor Microenvironment in Colon Cancer. Biomedicines 2024, 12, 697. https://doi.org/10.3390/biomedicines12030697
Kuo F-C, Lai JC-Y, Shieh H-R, Liou W-Z, Bair M-J, Chen Y-J. Cimetidine Attenuates Therapeutic Effect of Anti-PD-1 and Anti-PD-L1 and Modulates Tumor Microenvironment in Colon Cancer. Biomedicines. 2024; 12(3):697. https://doi.org/10.3390/biomedicines12030697
Chicago/Turabian StyleKuo, Feng-Chi, Jerry Cheng-Yen Lai, Hui-Ru Shieh, Wan-Zu Liou, Ming-Jong Bair, and Yu-Jen Chen. 2024. "Cimetidine Attenuates Therapeutic Effect of Anti-PD-1 and Anti-PD-L1 and Modulates Tumor Microenvironment in Colon Cancer" Biomedicines 12, no. 3: 697. https://doi.org/10.3390/biomedicines12030697
APA StyleKuo, F.-C., Lai, J. C.-Y., Shieh, H.-R., Liou, W.-Z., Bair, M.-J., & Chen, Y.-J. (2024). Cimetidine Attenuates Therapeutic Effect of Anti-PD-1 and Anti-PD-L1 and Modulates Tumor Microenvironment in Colon Cancer. Biomedicines, 12(3), 697. https://doi.org/10.3390/biomedicines12030697