
Electroacupuncture Reduces Fibromyalgia Pain via Neuronal/Microglial Inactivation and Toll-like Receptor 4 in the Mouse Brain: Precise Interpretation of Chemogenetics



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. FM Model and Bio-Plex ELISA
2.3. EA Treatments
2.4. Pain Behavior Test
2.5. Western Blot Analysis
2.6. Immunofluorescence
2.7. Chemogenetics Operation
2.8. Statistical Analysis
3. Results
3.1. Electroacupuncture Inhibits Cold Stress-Induced FM Pain in Mice
3.2. Inflammatory Mediators Were Increased in the FM Model and Reduced by EA but Not Sham EA
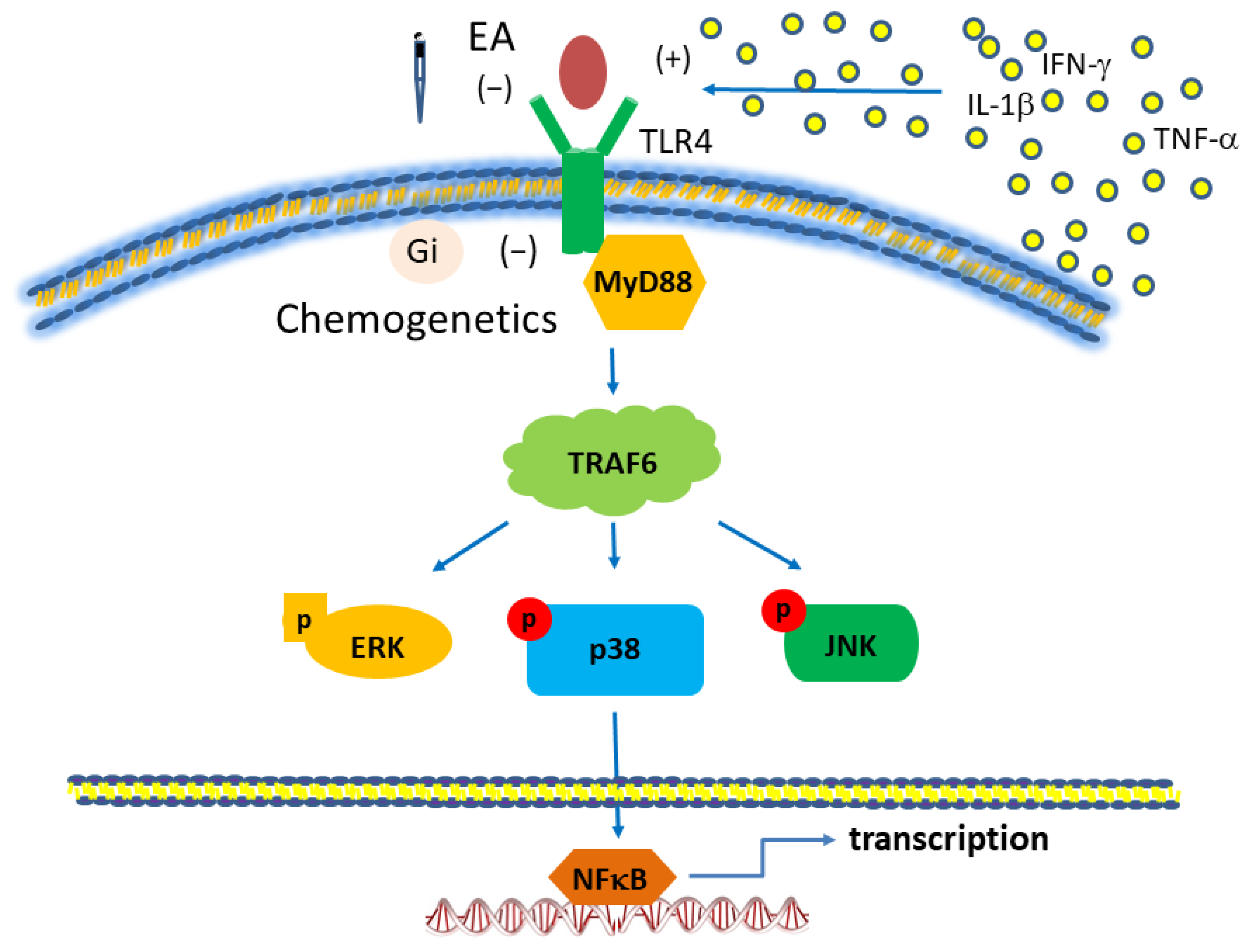
3.3. EA but Not Sham EA Reduced Fibromyalgia Pain through Microglia and TLR4 Signaling Pathways in the Mouse Thalamus
3.4. EA but Not Sham EA Reduced FM through Microglia and TLR4 Signaling Pathways in the Mouse Medial Prefrontal Cortex
3.5. EA but Not Sham EA Reduced FM through Microglia and TLR4 Signaling Pathways in the Mouse Somatosensory Cortex
3.6. EA but Not Sham EA Reduced FM through Microglia and TLR4 Signaling Pathways in the Mouse Amygdala
3.7. Chemogenetics Inhibition of SSC Attenuated Fibromyalgia Pain through the TLR4 Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spaeth, M. Epidemiology, costs, and the economic burden of fibromyalgia. Arthritis Res. Ther. 2009, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, L.P. Worldwide epidemiology of fibromyalgia. Curr. Pain Headache Rep. 2013, 17, 356. [Google Scholar] [CrossRef] [PubMed]
- Knight, T.; Schaefer, C.; Chandran, A.; Zlateva, G.; Winkelmann, A.; Perrot, S. Health-resource use and costs associated with fibromyalgia in France, Germany, and the United States. Clin. Outcomes Res. 2013, 5, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Guymer, E.K.; Littlejohn, G.O.; Brand, C.K.; Kwiatek, R.A. Fibromyalgia onset has a high impact on work ability in Australians. Intern. Med. J. 2016, 46, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Sarzi-Puttini, P.; Giorgi, V.; Marotto, D.; Atzeni, F. Fibromyalgia: An update on clinical characteristics, aetiopathogenesis and treatment. Nat. Rev. Rheumatol. 2020, 16, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.-Y.; Lin, Y.-W. Electroacupuncture reduces cold stress-induced pain through microglial inactivation and transient receptor potential V1 in mice. Chin. Med. 2021, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Dubner, R. Interactions between the immune and nervous systems in pain. Nat. Med. 2010, 16, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Jung, Y.-H.; Lee, D.; Lee, W.J.; Jang, J.H.; Lee, J.-Y.; Choi, S.-H.; Moon, J.Y.; Cheon, G.J.; Kang, D.-H. Abnormal neuroinflammation in fibromyalgia and CRPS using [11C]-(R)-PK11195 PET. PLoS ONE 2021, 16, e0246152. [Google Scholar] [CrossRef]
- Nishiyori, M.; Ueda, H. Prolonged gabapentin analgesia in an experimental mouse model of fibromyalgia. Mol. Pain 2008, 4, 52. [Google Scholar] [CrossRef]
- Lottering, B.; Lin, Y.-W. Functional characterization of nociceptive mechanisms involved in fibromyalgia and electroacupuncture. Brain Res. 2021, 1755, 147260. [Google Scholar] [CrossRef]
- Lin, Y.-W.; Chou, A.I.W.; Su, H.; Su, K.-P. Transient receptor potential V1 (TRPV1) modulates the therapeutic effects for comorbidity of pain and depression: The common molecular implication for electroacupuncture and omega-3 polyunsaturated fatty acids. Brain Behav. Immun. 2020, 89, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Kuzmich, N.N.; Sivak, K.V.; Chubarev, V.N.; Porozov, Y.B.; Savateeva-Lyubimova, T.N.; Peri, F. TLR4 Signaling Pathway Modulators as Potential Therapeutics in Inflammation and Sepsis. Vaccines 2017, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.C.; Hsieh, C.L.; Wu, S.Y.; Lin, Y.W. Toll-like receptor 2 plays an essential role in electroacupuncture analgesia in a mouse model of inflammatory pain. Acupunct. Med. 2019, 37, 356–364. [Google Scholar] [CrossRef]
- Capiralla, H.; Vingtdeux, V.; Zhao, H.; Sankowski, R.; Al-Abed, Y.; Davies, P.; Marambaud, P. Resveratrol mitigates lipopolysaccharide- and Aβ-mediated microglial inflammation by inhibiting the TLR4/NF-κB/STAT signaling cascade. J. Neurochem. 2012, 120, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Facci, L.; Barbierato, M.; Marinelli, C.; Argentini, C.; Skaper, S.D.; Giusti, P. Toll-Like Receptors 2, -3 and -4 Prime Microglia but not Astrocytes Across Central Nervous System Regions for ATP-Dependent Interleukin-1β Release. Sci. Rep. 2014, 4, 6824. [Google Scholar] [CrossRef]
- Ghowsi, M.; Qalekhani, F.; Farzaei, M.H.; Mahmudii, F.; Yousofvand, N.; Joshi, T. Inflammation, oxidative stress, insulin resistance, and hypertension as mediators for adverse effects of obesity on the brain: A review. Biomedicine 2021, 11, 13–22. [Google Scholar] [CrossRef]
- Liao, E.-T.; Lin, Y.-W.; Huang, C.-P.; Tang, N.-Y.; Hsieh, C.-L. Electric Stimulation of Ear Reduces the Effect of Toll-Like Receptor 4 Signaling Pathway on Kainic Acid-Induced Epileptic Seizures in Rats. BioMed Res. Int. 2018, 2018, 5407256. [Google Scholar] [CrossRef]
- Wu, M.K.; Huang, T.L.; Huang, K.W.; Huang, Y.L.; Hung, Y.Y. Association between toll-like receptor 4 expression and symptoms of major depressive disorder. Neuropsychiatr. Dis. Treat. 2015, 11, 1853–1857. [Google Scholar]
- Thakur, K.K.; Saini, J.; Mahajan, K.; Singh, D.; Jayswal, D.P.; Mishra, S.; Bishayee, A.; Sethi, G.; Kunnumakkara, A.B. Therapeutic implications of toll-like receptors in peripheral neuropathic pain. Pharmacol. Res. 2017, 115, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Walter, S.; Letiembre, M.; Liu, Y.; Heine, H.; Penke, B.; Hao, W.; Bode, B.; Manietta, N.; Walter, J.; Schulz-Schüffer, W.; et al. Role of the toll-like receptor 4 in neuroinflammation in Alzheimer’s disease. Cell. Physiol. Biochem. 2007, 20, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Loeches, S.; Pascual-Lucas, M.; Blanco, A.M.; Sanchez-Vera, I.; Guerri, C. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. J. Neurosci. 2010, 30, 8285–8295. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, S.; Herkenham, M. Toll-Like Receptor 4 on Nonhematopoietic Cells Sustains CNS Inflammation during Endotoxemia, Independent of Systemic Cytokines. J. Neurosci. 2005, 25, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Su, K.-P.; Chou, L.-W.; Sun, M.-F.; Lin, J.-G. Acupuncture Treatment in Depression. In Experimental Acupuncturology; Lin, J.-G., Ed.; Springer: Singapore, 2018; pp. 43–66. [Google Scholar]
- Tsai, S.T.; Wei, T.H.; Yang, Y.W.; Lu, M.K.; San, S.; Tsai, C.H.; Lin, Y.W. Transient receptor potential V1 modulates neuroinflammation in Parkinson’s disease dementia: Molecular implications for electroacupuncture and rivastigmine. Iran J. Basic Med. Sci. 2021, 24, 1336–1345. [Google Scholar]
- Lin, S.-K.; Liu, J.-M.; Hsu, R.-J.; Chuang, H.-C.; Wang, Y.-X.; Lin, P.-H. Incidence of iatrogenic pneumothorax following acupuncture treatments in Taiwan. Acupunct. Med. 2019, 37, 332–339. [Google Scholar] [CrossRef] [PubMed]
- NIH Consensus. Conference. Acupuncture. Jama 1998, 280, 1518–1524. [Google Scholar] [CrossRef]
- Deare, J.C.; Zheng, Z.; Xue, C.C.; Liu, J.P.; Shang, J.; Scott, S.W.; Littlejohn, G. Acupuncture for treating fibromyalgia. Cochrane Database Syst. Rev. 2013, 2013, Cd007070. [Google Scholar] [CrossRef]
- Bai, Y.; Guo, Y.; Wang, H.; Chen, B.; Wang, Z.; Liu, Y.; Zhao, X.; Li, Y. Efficacy of acupuncture on fibromyalgia syndrome: A meta-analysis. J. Tradit. Chin. Med. 2014, 34, 381–391. [Google Scholar] [CrossRef]
- Roth, B.L. DREADDs for Neuroscientists. Neuron 2016, 89, 683–694. [Google Scholar] [CrossRef]
- Armbruster, B.N.; Li, X.; Pausch, M.H.; Herlitze, S.; Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 2017, 104, 5163–5168. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y.; Miyakawa, N.; Takuwa, H.; Hori, Y.; Oyama, K.; Ji, B.; Takahashi, M.; Huang, X.-P.; Slocum, S.T.; DiBerto, J.F.; et al. Deschloroclozapine, a potent and selective chemogenetic actuator enables rapid neuronal and behavioral modulations in mice and monkeys. Nat. Neurosci. 2020, 23, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, I.-H.; Yen, C.-M.; Hsu, H.-C.; Liao, H.-Y.; Lin, Y.-W. Chemogenetics Modulation of Electroacupuncture Analgesia in Mice Spared Nerve Injury-Induced Neuropathic Pain through TRPV1 Signaling Pathway. Int. J. Mol. Sci. 2024, 25, 1771. [Google Scholar] [CrossRef]
- Ji, R.-R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef] [PubMed]
- Borchers, A.T.; Gershwin, M.E. Fibromyalgia: A Critical and Comprehensive Review. Clin. Rev. Allergy Immunol. 2015, 49, 100–151. [Google Scholar] [CrossRef] [PubMed]
- Geisser, M.E.; Glass, J.M.; Rajcevska, L.D.; Clauw, D.J.; Williams, D.A.; Kileny, P.R.; Gracely, R.H. A psychophysical study of auditory and pressure sensitivity in patients with fibromyalgia and healthy controls. J. Pain 2008, 9, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-C.; Tsai, S.-T.; Ting, B.; Cheng, Y.-C.; Wang, C.-K.; Chang, J.P.-C.; Su, K.-P. Psychological Outcomes and Quality of Life of Fibromyalgia Patients with Vitamin D Supplementation—A Meta-Analysis. J. Clin. Med. 2023, 12, 2750. [Google Scholar] [CrossRef] [PubMed]
- Martenson, M.E.; Halawa, O.I.; Tonsfeldt, K.J.; Maxwell, C.A.; Hammack, N.; Mist, S.D.; Pennesi, M.E.; Bennett, R.M.; Mauer, K.M.; Jones, K.D.; et al. A possible neural mechanism for photosensitivity in chronic pain. Pain 2016, 157, 868–878. [Google Scholar] [CrossRef]
- Dehghan, M.; Schmidt-Wilcke, T.; Pfleiderer, B.; Eickhoff, S.B.; Petzke, F.; Harris, R.E.; Montoya, P.; Burgmer, M. Coordinate-based (ALE) meta-analysis of brain activation in patients with fibromyalgia. Hum. Brain Mapp. 2016, 37, 1749–1758. [Google Scholar] [CrossRef]
- Murillo-Garcia, A.; Leon-Llamas, J.L.; Villafaina, S.; Gusi, N. Fibromyalgia impact in the prefrontal cortex subfields: An assessment with MRI. Clin. Neurol. Neurosurg. 2022, 219, 107344. [Google Scholar] [CrossRef]
- Simons, L.E.; Moulton, E.A.; Linnman, C.; Carpino, E.; Becerra, L.; Borsook, D. The human amygdala and pain: Evidence from neuroimaging. Hum. Brain Mapp. 2014, 35, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Ichesco, E.; Ratai, E.-M.; Gonzalez, R.G.; Burdo, T.; Loggia, M.L.; Harris, R.E.; Napadow, V. Magnetic resonance imaging of neuroinflammation in chronic pain: A role for astrogliosis? Pain 2020, 161, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, D.S.; Ahmed, S.U.; Kettner, N.W.; Borra, R.J.; Cohen-Adad, J.; Deng, H.; Houle, T.T.; Opalacz, A.; Roth, S.A.; Melo, M.F.V.; et al. Neuroinflammation of the spinal cord and nerve roots in chronic radicular pain patients. Pain 2018, 159, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Ohgidani, M.; Kato, T.A.; Hosoi, M.; Tsuda, M.; Hayakawa, K.; Hayaki, C.; Iwaki, R.; Sagata, N.; Hashimoto, R.; Inoue, K.; et al. Fibromyalgia and microglial TNF-α: Translational research using human blood induced microglia-like cells. Sci. Rep. 2017, 7, 11882. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.A.; Basith, S.; Choi, S. Negative regulatory approaches to the attenuation of Toll-like receptor signaling. Exp. Mol. Med. 2013, 45, e11. [Google Scholar] [CrossRef] [PubMed]
- Lacagnina, M.J.; Watkins, L.R.; Grace, P.M. Toll-like receptors and their role in persistent pain. Pharmacol. Ther. 2018, 184, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, W.; Zhu, C.; Sun, S.; Wu, S.; Wang, L.; Wang, Y.; Ge, Z. Toll-like receptors and their role in neuropathic pain and migraine. Mol. Brain 2022, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Branco, J.C.; Saraiva, F.; Cerinic, M.M.; Zoppi, M.; Martin, N.; Taieb, C.; Myon, E. Fibromyalgia Syndrom: A European Epidemiological Survey. In Annals of the Rheumatic Diseases; BMJ Publishing Group: London, UK; British Medical Association: London, UK, 2005; Volume 64, p. 547. [Google Scholar]
- Itoh, K.; Kitakoji, H. Effects of acupuncture to treat fibromyalgia: A preliminary randomised controlled trial. Chin. Med. 2010, 5, 11. [Google Scholar] [CrossRef]
- Ishizaki, N.; Yano, T.; Kawakita, K. Public status and prevalence of acupuncture in Japan. Evid. Based Complement. Altern. Med. 2010, 7, 493–500. [Google Scholar] [CrossRef]
- Beecher, H.K. The Powerful Placebo. J. Am. Med. Assoc. 1955, 159, 1602–1606. [Google Scholar] [CrossRef]
- Chen, X.; Zou, K.; Abdullah, N.; Whiteside, N.; Sarmanova, A.; Doherty, M.; Zhang, W. The placebo effect and its determinants in fibromyalgia: Meta-analysis of randomised controlled trials. Clin. Rheumatol. 2017, 36, 1623–1630. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Chen, W.-H.; Hsieh, C.-L.; Lin, Y.-W. Abundant expression and functional participation of TRPV1 at Zusanli acupoint (ST36) in mice: Mechanosensitive TRPV1 as an “acupuncture-responding channel”. BMC Complement. Altern. Med. 2014, 14, 96. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, S.-T.; Yang, C.-C.; Liao, H.-Y.; Lin, Y.-W. Electroacupuncture Reduces Fibromyalgia Pain via Neuronal/Microglial Inactivation and Toll-like Receptor 4 in the Mouse Brain: Precise Interpretation of Chemogenetics. Biomedicines 2024, 12, 387. https://doi.org/10.3390/biomedicines12020387
Tsai S-T, Yang C-C, Liao H-Y, Lin Y-W. Electroacupuncture Reduces Fibromyalgia Pain via Neuronal/Microglial Inactivation and Toll-like Receptor 4 in the Mouse Brain: Precise Interpretation of Chemogenetics. Biomedicines. 2024; 12(2):387. https://doi.org/10.3390/biomedicines12020387
Chicago/Turabian StyleTsai, Sheng-Ta, Chia-Chun Yang, Hsien-Yin Liao, and Yi-Wen Lin. 2024. "Electroacupuncture Reduces Fibromyalgia Pain via Neuronal/Microglial Inactivation and Toll-like Receptor 4 in the Mouse Brain: Precise Interpretation of Chemogenetics" Biomedicines 12, no. 2: 387. https://doi.org/10.3390/biomedicines12020387
APA StyleTsai, S.-T., Yang, C.-C., Liao, H.-Y., & Lin, Y.-W. (2024). Electroacupuncture Reduces Fibromyalgia Pain via Neuronal/Microglial Inactivation and Toll-like Receptor 4 in the Mouse Brain: Precise Interpretation of Chemogenetics. Biomedicines, 12(2), 387. https://doi.org/10.3390/biomedicines12020387