1. Introduction
In addressing cancer, a paramount health threat, it’s essential to comprehend the mechanisms of cell death [
1]. A recent paper has identified a novel cell death pathway known as “disulfidptosis”, which is divergent from other conventional mechanisms such as apoptosis, necroptosis, and oncosis. For cancer cells experiencing glucose starvation, elevated expression of cystine transporter solute carrier family 7 member 11 (SLC7A11; also known as xCT), a cystine transporter, leads to NADPH depletion and abnormal disulfide bonding, which ultimately results in actin network collapse and subsequent cell death [
2]. A visual representation of the specific mechanism can be found in
Supplementary Figure S1. As a tumor metabolism-related pathway, disulfide bond formation is found in cancer-related proteins, meaning this metastable bond can be a target for future therapies [
3]. In addition, disulfidptosis has been demonstrated to be associated with immune response and prognosis in certain cancers, such as hepatocellular carcinoma (HCC), esophageal squamous cell carcinoma (ESCC), and bladder cancer (BCa) [
4,
5,
6]. Prior research established disulfidptosis as a predictive biomarker for immune characteristics and drug responses, suggesting SLC7A11-mediated disulfidptosis as a novel therapeutic strategy in cancer.
The current consensus is that chronic inflammation, characterized by immune cell infiltration, can promote the initiation or progression of malignancies, including colorectal tumors, renal cell carcinoma, and HCC [
7]. Recent evidence reveals that certain cell death modalities can intensify inflammation rather than just result from it. These processes vary in their capacity to provoke inflammation, influenced by the differential release of mediators, affecting dead cell clearance. Subsequent increases in cytokines, chemokines, and reactive oxygen species escalate the inflammatory response [
8]. Additionally, targeted therapy induces significant modifications in the transcriptional dynamics within cancer cells and the adjacent tumor milieu, which are pivotal in determining the emergent tumor phenotypes associated with pharmacological sensitivity or resistance [
9]. In the treatment of HCC, traditional chemotherapy agents like doxorubicin and fluorouracil have shown limited efficacy and are associated with substantial adverse effects [
10]. Recent studies have witnessed groundbreaking randomized controlled trials in advanced hepatocellular carcinoma, heralding significant shifts in clinical practice [
11]. These include the introduction of lenvatinib as a first-line alternative to sorafenib and the second-line use of regorafenib, cabozantinib, and ramucirumab following sorafenib. Among them, regorafenib and cabozantinib were suggested to be chosen as second-line therapies [
12]. Moreover, the efficacy of immunotherapy has been cemented in HCC treatment through the combination of the anti-PD-L1 agent atezolizumab and the anti-vascular endothelial growth factor (VEGF) agent bevacizumab, which gradually becomes a first-line choice [
13]. Prior studies show that SCL7A11-mediated ferroptosis in tumor-associated macrophages significantly alters the hepatocellular carcinoma microenvironment. This underscores the potential of targeting SCL7A11 in HCC treatment, suggesting that combining SCL7A11 with immunotherapy could enhance therapeutic efficacy clinically [
14]. The mechanisms by which SLC7A11-mediated disulfideptosis operates and influences the immune microenvironment and inflammation remain nascent. Consequently, additional research is needed to clarify disulfideptosis’s role within the tumor microenvironment.
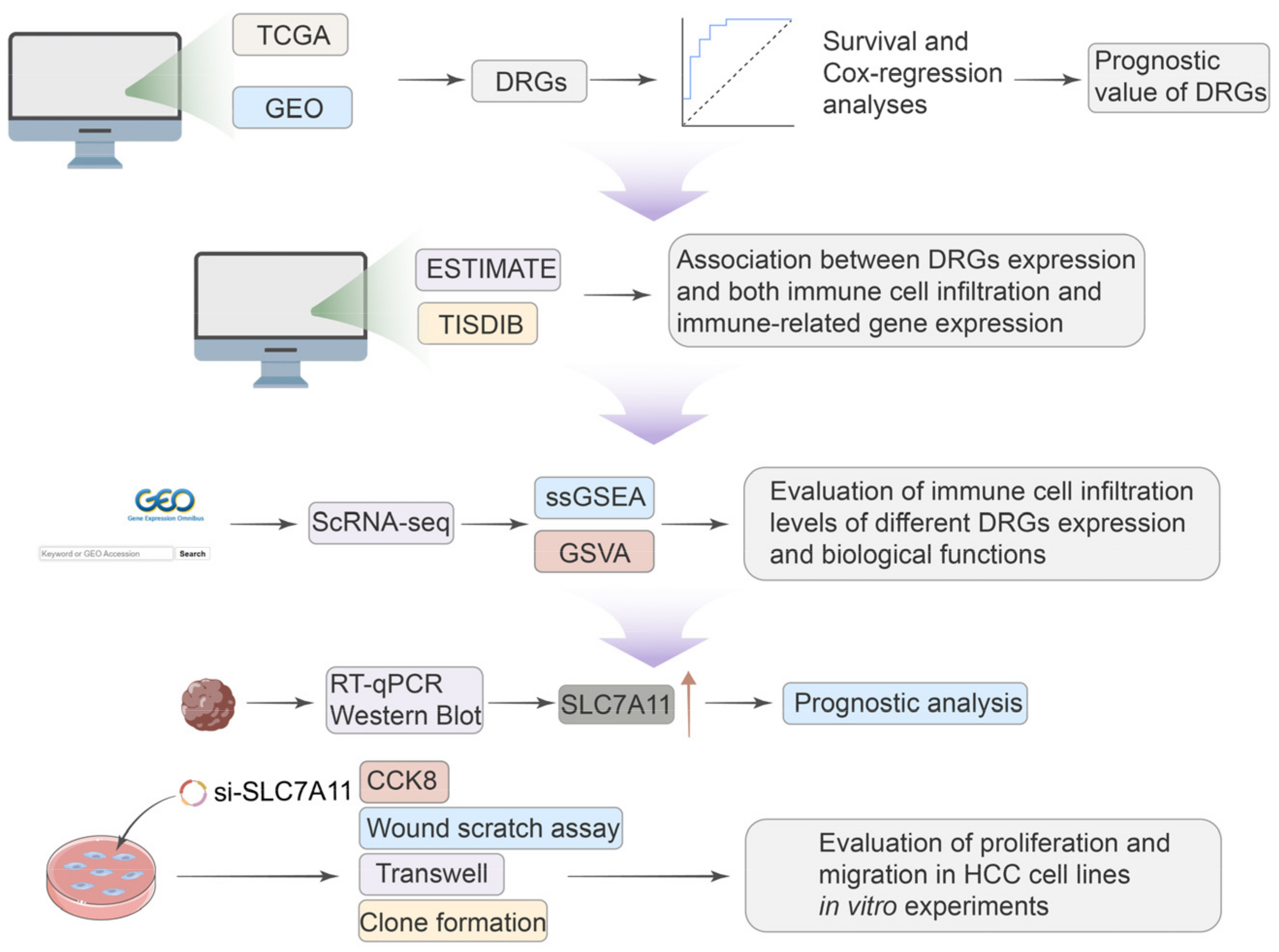
Given the significant role of disulfidoptosis in cancer, we analyzed pan-cancer data across multiple databases. We aimed to investigate the differential expression, prognostic significance, and biological implications of disulfidoptosis-related genes (DRGs) in various tumors. Critical insights from single-cell sequencing further illuminated the intricate cellular heterogeneity of HCC, often hidden in traditional bulk RNA sequencing approaches, emphasizing the nuanced roles that DRGs play within this cellular mosaic [
15]. We observed different immune statuses and biological functions in HCC with differential DRG expression, underlining the pivotal role disulfidptosis has in shaping TME characteristics in HCC. The expression of SCL7A11 was confirmed at both mRNA and protein levels using qRT-PCR and Western blotting in clinical samples and cell lines. And the knockdown of SLC7A11 resulted in diminished proliferation and migration in HCC cell lines. Our research methodology is delineated in
Figure 1. Our findings highlight disulfidoptosis as a promising biomarker and therapeutic avenue for malignancies, notably hepatocellular carcinoma.
2. Materials and Methods
2.1. Data Collection and Processing
According to the study of Xiaoguang Liu, a total of 10 genes were identified as key genes most associated with disulfidoptosis, which were annotated as DRGs [
2]. The genomic, clinicopathological, and somatic mutation data of 33 TCGA GDC pan cancers were gathered from the University of California Santa Cruz (UCSC) Xena (
https://xenabrowser.net/, accessed on 6 June 2023). The fragment per kilobase million (FPKM) value was normalized before comparison. The abbreviations of various cancers are provided in
Supplementary Table S1.
The GSE242889 dataset encompassed single-cell RNA sequencing details of five HCC specimens based on the 10X Genomics platform. The dataset was constructed on the platform GPL24676 via the Illumina NovaSeq 6000 system. To meticulously scrutinize the scRNA-seq dataset, a stepwise analytical framework was invoked. As a starting point, data preprocessing was accomplished through the Seurat toolkit. This stage paved the way for a comprehensive series of analyses. Utilizing the PercentageFeatureSet function, we delineated the proportion of mitochondrial gene presence. Additionally, we performed correlational evaluations to probe relationships among sequencing depth, mitochondrial gene content, and overall intracellular transcriptomes.
To bolster the precision of our analysis, only genes expressed in a minimum of five cells were considered. Cells were stringently curated based on explicit benchmarks: gene expression counts lying between 300 and 5000, a mitochondrial constitution below 10%, and an essential UMI threshold of 1000 per cell. This rigorous curation was geared towards ensuring cellular data fidelity. After the data filtering process, we normalized the scRNA-seq data using the LogNormalize method, thereby enhancing the accuracy of downstream analysis and interpretation.
For downstream analysis, the top 20 principal components (PCs) were subjected to Seurat’s Elbow plot program. With a resolution of 0.7, primary cell clusters were identified by Seurat’s Find Clusters tool, which were subsequently visualized via two-dimensional UMAP plots. Cells were categorized into previously recognized biological cell types using conventional markers established in former studies.
2.2. Survival Analysis
Based on the median expression levels of DRGs, patients were stratified into low- and high-expression cohorts. We employed univariate Cox regression analysis on TCGA datasets, utilizing the R packages “survival” and “forestplot”, to assess the prognostic significance of DRGs regarding overall survival (OS). The derived log-rank p-values and hazard ratios (HRs), complemented with 95% confidence intervals, are illustrated in forest plots to provide a comprehensive representation of the survival analysis. Additionally, KM survival analysis was executed to compare OS between the two defined expression cohorts, facilitated by the “survminer” and “survival” packages in R.
2.3. Immune Subtypes and Stemness Score Analysis
The immune subtype analysis of pan cancer was performed utilizing the “limma”, “ggplot2”, and “reshape2” packages in R. The p-value was set at 0.05 as a threshold for statistical significance.
2.4. DNA Promoter Methylation Analysis
The GSCA database (
http://bioinfo.life.hust.edu.cn/GSCA, accessed on 6 June 2023) was employed to scrutinize the promoter methylation distribution of DRGs and the correlation between DNA promoter methylation levels and DRG mRNA expression, as well as survival outcomes in diverse cancers. Correlations are depicted through detailed scatter diagrams, and Spearman’s correlation coefficient was applied to calculate correlation values (Cor) and false discovery rates (FDR), offering a robust quantitative analysis of the interdependencies.
2.5. Immune Infiltration Analysis
The ESTIMATE algorithm, which is a computational method designed to infer the fraction of immune and stromal cells within tumor samples by analyzing signatures of gene expression, has been used to calculate the ImmuneScore (Immune Component Ratio), StromalScore (Matrix Component Ratio), and ESTIMATEScore (Sum of ImmuneScore and StromalScore) for tumor purity prediction [
16]. Spearman correlation was used to calculate the correlation between these scores and DRG expression, and the results are presented as scatter plots with the
p-value and Cor.
TME cell infiltration was evaluated using a single-sample gene set enrichment analysis. Utilizing the “GSVA” R package, we conducted a single-sample gene set enrichment analysis (ssGSEA) to gauge the infiltration rates of various immune cell populations. The ssGSEA technique interrogates individual oncological specimens based on gene profiles characteristic of specific immune cells [
17]. We conducted correlation analysis using the Spearman method to assess the relationship between essential genes and 24 immune cell types. Adopting a deconvolution methodology, we assessed the presence of 24 distinct immune cell types, which encompassed activated B cells, CD4+, and CD8+ T cells in their activated states, dendritic cells (both activated and immature forms), natural killer cells characterized by CD56 bright and CD56 dim phenotypes, eosinophils, gamma delta T cells, immature B cells, myeloid-derived suppressor cells (MDSCs), macrophages, mast cells, monocytes, NK-T cells, neutrophils, plasmacytoid dendritic cells, regulatory T cells, T follicular helper cells, and the T helper cell subsets—Th1, Th2, and Th17 [
18].
2.6. Single-Sample Gene Set Enrichment Analysis (ssGSEA) and Gene Set Enrichment Analysis
In biomedical analysis, ssGSEA is essential for calculating gene set enrichment scores within a single sample. The ssGSEA score serves as an indicator, elucidating the extent of systematic upregulation or downregulation of a designated gene set within a sample [
19]. We derived DRG scores for each sample using ssGSEA via the R “GSVA” package. We employed the “GSVA” package in R to execute a gene set variation analysis, delving into the biological functionalities and potential pathways linked to the expression of DRGs. Prior to this analysis, the gene set designated as “c2.cp.kegg.v7.5.1.symbols” was extracted.
2.7. Cell Culture
Human osteoblast cells (hFOB1.19) and human osteosarcoma cells (U2OS) were gifts from Dr. HJ. Lu, Guangdong Pharmaceutical University. Human renal cancer cells (786-0) and human kidney cells (HK-2) were gifts from the research group of Prof. JH. Luo, Department of Urology, The First Affiliated Hospital of Sun Yat-Sen University. Human hepatocellular carcinoma cells (SNU449 and Huh7) and human normal hepatic cells (LO2) were acquired from the Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences (ATCC, Shanghai, China). Human U2OS and hFOB1.19 cell lines were maintained in DMEM media (Gibco, Thermo Fisher Scientific, Suzhou, China) enriched with 10% fetal bovine serum (ABW, AB-FBS-1050S, Uruguay) and 1% penicillin–streptomycin (Invitrogen, Carlsbad, CA, USA). The 786-0, HK-2, SNU449, Huh7, and LO2 cell lines were cultured in RPMI 1640 media (Gibco, Thermo Fisher Scientific, Suzhou, China) supplemented identically. Cells were cultured at 37 °C in a 5% CO2 atmosphere at a constant humidity of 80%. Cells in the logarithmic growth stage, approaching 80% confluence, were selected for subsequent experiments.
2.8. Clinical Samples Collection
Samples of hepatocellular carcinomas and adjacent normal tissues were collected from patients admitted to the First Affiliated Hospital of Sun Yat-sen University inbetween 2019 and 2020. The patients range in age from 18 to 80. The patients did not undergo neoadjuvant chemotherapy or radiotherapy prior to surgery. All samples were obtained through surgical resection and were processed and stored in liquid nitrogen containers within half an hour post-surgery. A review and approval of this study were obtained from the Institutional Ethics Committee for Clinical Research of the First Affiliated Hospital of Sun Yat-sen University ((2021)170). We conducted all experimental procedures in accordance with the Helsinki Declaration and obtained written informed consent from all patients.
2.9. RNA Isolation and Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Analysis
The results of this study were verified by quantitative real-time PCR (qRT-PCR) after RNA was isolated from 10 different cell lines, including hFOB1.19, U2OS, 786-0, KH-2, SNU449, Huh7, LO2, and si-SLC7A11-transfected SNU449 or Huh7, as well as 8 pairs of HCC tissues and adjacent normal hepatic tissues. As part of the RT-qPCR process, primer sequences for the relevant genes are found in
Supplementary Table S2 and are synthesized by RiboBio (Guangzhou, China). As stated in
Supplementary Table S2, each primer has its own melting temperature (Tm) and annealing temperature (Tm-5 °C). The total RNA was extracted by the EZ-press RNA Purification Kit (EZBioscience, ZScience Biotechnology Corporation Limited, Roseville, MN, USA), and the purity and concentration of the total RNA were assessed using a NanoDrop 2000 spectrometer. As specified by the manufacturer, the RT-qPCR reaction system and conditions were conducted using the 4×Reverse Transcription Master Mix (EZBioscience, ZScience Biotechnology Corporation Limited, Roseville, MN, USA) and 2×SYBR Green qPCR Master Mix (EZBioscience, ZScience Biotechnology Corporation Limited, Roseville, MN, USA) on the Applied Biosystems™ QuantStudio™ 5 Real-Time PCR System. Drawing the figure was performed using Graphpad Prism 9.0 software (GraphPad Prism 9.0,
https://www.graphpad-prism.cn/, accessed on 23 February 2022, China). In order to analyze the RT-qPCR data, we used GAPDH as an internal reference, and the 2-ΔΔCt method was used to normalize the expression of the genes targeted. In the studies, cells were used when the confluence rate reached approximately 80% while they were in the logarithmic growth stage.
2.10. Western Blotting and Immunohistochemical
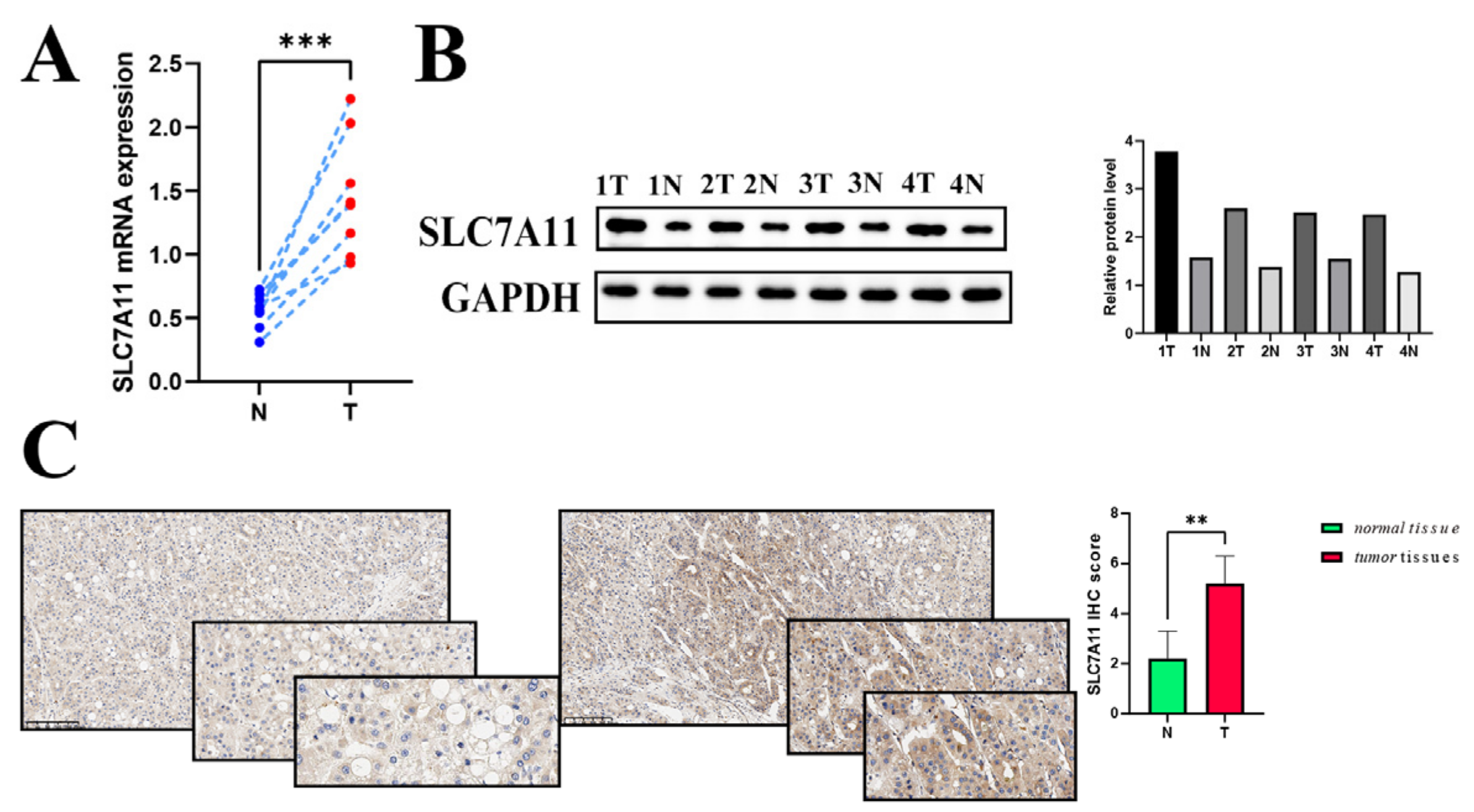
We lysed cell lines and four pairs of hepatocellular carcinoma tissues using RIPA lysis buffer (Beyotime, Shanghai, China) containing a Protease Inhibitor Cocktail (CoWin Biosciences, Jiangsu, China) (1:100). A bicinchoninic acid (BCA) protein assay kit (Thermo Scientific, Guangzhou, China) was utilized to quantify total protein. Following protein quantification, 10 ug of every protein sample was loaded onto a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). A 0.45 μm polyvinylidene difluoride membrane was used to transfer total proteins after electrophoresis. Further blocking of the membranes was conducted in a protein-free rapid blocking buffer (Epizyme Biotech, PS108) for 10–15 min. An ECL chromogenic kit (Thermo Fisher Scientific, Guangzhou, China) was used to detect antibody binding after sequential incubation with primary antibodies (overnight at 4 °C) and secondary antibodies (1 h at room temperature). The imaging systems using chemiluminescence (Beijing, China) and the Fusion FX5 image analyzer (Vilber Lourmat, Marne La Vallée, France) were used to visualize antibody binding. The following primary antibodies were as follows: SLC7A11 (Proteintech, Wuhan, China), GAPDH (CST, NewYork, NY, USA), and β-Actin (CST).
For immunohistochemical (IHC) analysis, sections derived from human hepatocellular carcinoma and matched normal liver tissues were used. IHC staining was performed at Wuhan Servicebio Technology. Anti-CD206 Rabbit pAb (GB113497-100; Servicebio, Wuhan, China), anti-SLC7A11/xCT Rabbit pAb (GB115276-100), and anti-FOXP3 Rabbit pAb (GB112325-100; Servicebio) were used to assess macrophages and Treg cells on paraffin-embedded tumor tissue and matched normal tissue slides. Images were captured with the OCUS portable digital scanning microscopic imaging system (APG Bio, Shanghai, China). Following a quick examination of slide quality, the IHC results were evaluated by two independent pathologists. The intensity of IHC staining and the proportion of positively stained tumor cells were distinctly assessed, as detailed below. The percentage of stained tumor cells was recorded as follows: 0%: 0; <1%: 1; 1–10%: 2; 11–33%: 3; 34–66%: 4; and >66%: 5. The degree of staining intensity was categorized as: 0; weak staining: 1; moderate staining: 2; strong staining: 3. IHC staining score = the percentage of stained tumor cells + staining intensity. These IHC results were grouped according to IHC staining score as: negative (IHC staining score: <3), weakly positive (IHC staining score: 4–6), and strongly positive (IHC score: 7–8).
2.11. siRNA Interference Assay and CCK-8 Assay
Two SLC7A11-siRNAs were designed by HanYi Biosciences Inc., Guangzhou, China (
Supplementary Table S3). Transfecting SNU449 or Huh7 with siRNAs was performed using jetPRIME (Polyplus, Illkirch, French) with the manufacturer’s instructions. A functional assay was conducted 48 h after transfection, and proteins and RNA were harvested.
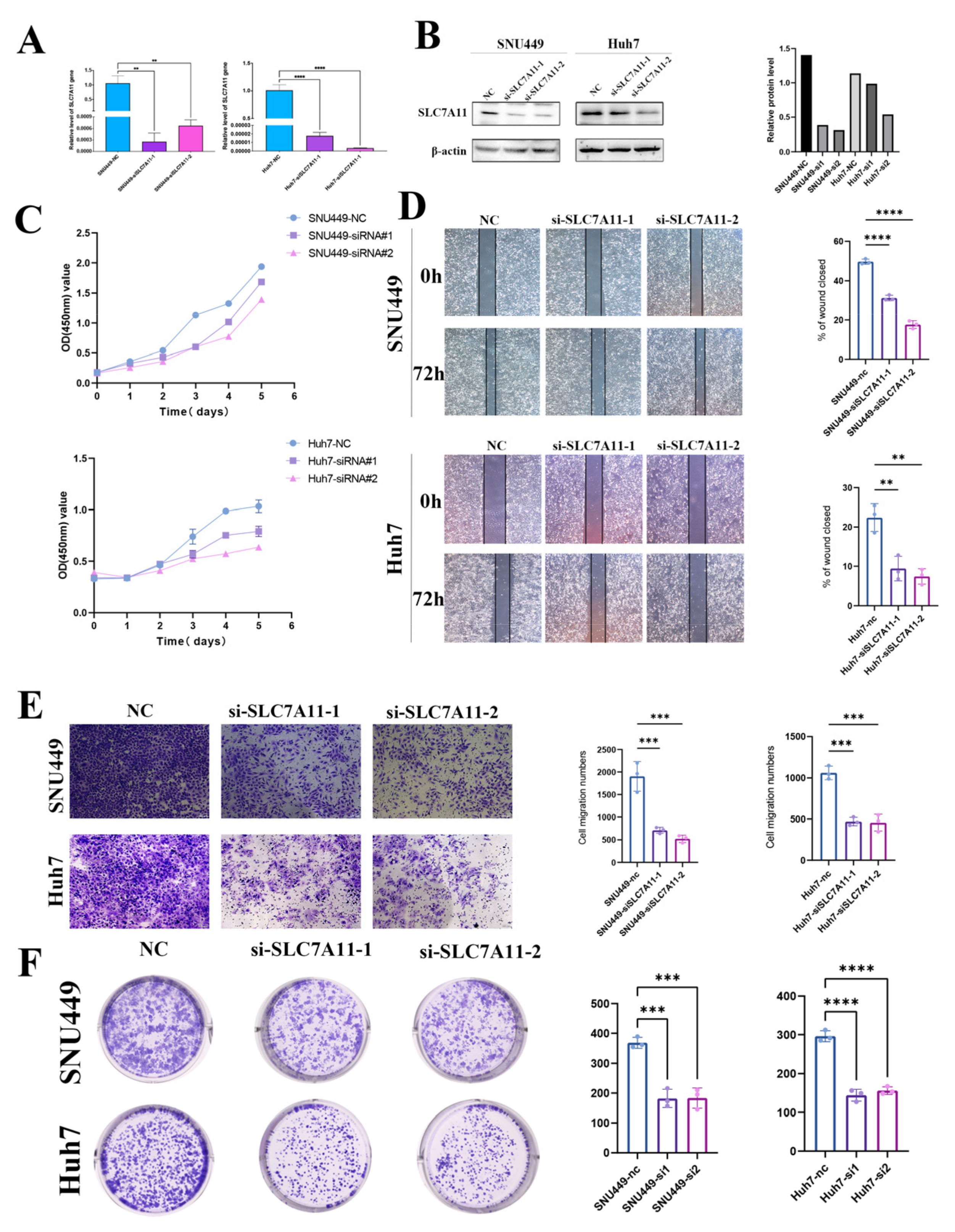
After transfection with SLC7A11-siRNA, cell proliferation was assessed using the Cell Counting Kit-8 (Dojindo, Japan, CK04-5). A 96-well plate was used to seed the cells. A microplate reader (Bio-Rad Laboratories, Hercules, CA, USA) was used to measure cell viability at 1, 2, 3, 4, and 5 days.
2.12. Wound Healing, Cell Migration Assays, Colony Formation Test
A pipette was used to scratch the cells after they had been merged into the six-well plate. After scratching, we took photos at 0 h and 76 h. SNU449 or Huh7 were starved in serum-free RPMI 1640 medium for 8 h in order to evaluate their migration capability. Next, 5 × 104 cells in 100 μL of serum-free RPMI 1640 medium were added to transwell inserts (Corning, NY, USA). As a nutritional attractant, serum-free RPMI 1640 medium with 10% FBS was used as a base for transwell assays. After 8 h, lower surface cells were fixed with 4% polyformaldehyde (Beyotime) for 30 min and stained with 0.4% crystal violet (Beyotime). The upper surface of the slide was wiped clean using a cotton swab, and the lower surface was counted under the microscope. Huh7 or SNU449 transfected with SLC7A11-siRNA were incubated in RPMI-1640 medium containing 10% FBS, maintained in RPMI-1640 medium containing 10% FBS, and incubated at 37 °C with 5% CO2. Within 24 h of transfection, we inoculated 1000 Huh7 or SNU449 cells into the six-well plates, which were then cultured in RPMI-1640 medium with 10% FBS for two weeks, and finally we counted and analyzed the colonies.
2.13. Statistical Analysis
The Kruskal–Wallis test and Wilcoxon rank-sum analysis were used to evaluate differences in gene expression levels between tumors and normal tissues. We used two-tailed tests and then set a significance threshold of
p < 0.05 for all parametric analyses in this study. The statistical analyses were conducted using the R software (version 4.1.3, manufacture, city, if any state, country R Foundation for Statistical Computing), which was obtained from the CRAN mirror hosted by Tsinghua University, Beijing, China (
https://www.r-project.org/), on 4 March 2023. The Fisher’s exact test and the
t-test, assuming equal variances, were used to evaluate group comparisons for categorical and continuous variables unless explicitly specified. We evaluated the diagnostic precision of gene expression levels in predicting preeclampsia, and we conducted an ROC curve analysis and calculated the AUC values. Statistical significance was set at
p < 0.05, unless otherwise noted.
4. Discussion
Cell proliferation and programmed cell death always play critical roles in the development of tumors [
23]. Previous research has established that most (perhaps even all) types of cancer cells are insensitive to apoptosis; therefore, inducing tumor cell non-programed necrosis is an effective way to treat the tumor [
24]. Emerging studies have shown that cuproptosis, ferroptosis, and proptosis play crucial roles in the occurrence and treatment of tumors [
25,
26]. Given that previous studies have shown that inflammation can augment cell death and increase cellular turnover, promoting tumorigenesis [
7]. In this context, apoptosis, characterized as a form of cell death that is devoid of inflammatory responses, emerges as a preferred clinical approach, especially in procedures like tumor ablation [
27]. Consequently, a deeper exploration of cell death promises insights into the foundational mechanisms of tumorigenesis, providing avenues for devising more potent antitumor therapeutic strategies.
Disulfidoptosis, a novel form of cell death distinct from necroptosis and apoptosis, highlights the role of disulfide and glucose metabolism in tumor cell death [
2]. Extracellular cystine transporter SLC7A11 promotes cysteine synthesis, maintaining intracellular glutathione levels to prevent cell death from abnormal disulfide bonds and reactive oxygen accumulation [
25]. ROS accumulation is a common feature in disulfidoptosis, ferroptosis, and cuproptosis, influencing inflammation-related diseases like cancer. ROS have a dual role in tumors, promoting cell death at high levels while stimulating tumor growth [
28]. The intricate relationship between disulfidoptosis and energy metabolism warrants further investigation, particularly in HCC, for potential therapeutic insights.
HCC ranks as the sixth most common cancer globally, characterized by a poor prognosis and short survival rates [
29]. Sorafenib, having gained approval from the US Food and Drug Administration for HCC treatment, is extensively utilized based on its efficacy in hindering tumor cell proliferation and angiogenesis [
30]. However, as the current first-line systemic drug, sorafenib faces potential discontinuation due to its limitations associated with multiple adverse events [
31]. And the standard second-line treatments for HCC are scarce. Recent studies have outlined the safety and efficacy profiles of capecitabine, regorafenib, cabozantinib, and ramucirumab, revealing their potential roles as second-line treatments for hepatocellular carcinoma [
13]. Four clinical trials have showcased the effectiveness of three different drugs in treating patients with HCC who are unresponsive to sorafenib. These trials include RESORCE (testing regorafenib), CELESTIAL (testing cabozantinib), and both REACH and REACH-2 (testing ramucirumab) [
12]. However, HCC exists within a complex immunological microenvironment, which results in lower remission and survival rates when treated with a single immunotherapy method or immunotherapies alone. Consequently, the emphasis of future development should be on multitarget combination therapy [
32,
33]. Identifying and developing new targeted agents is of paramount importance for the advancement of targeted therapy.
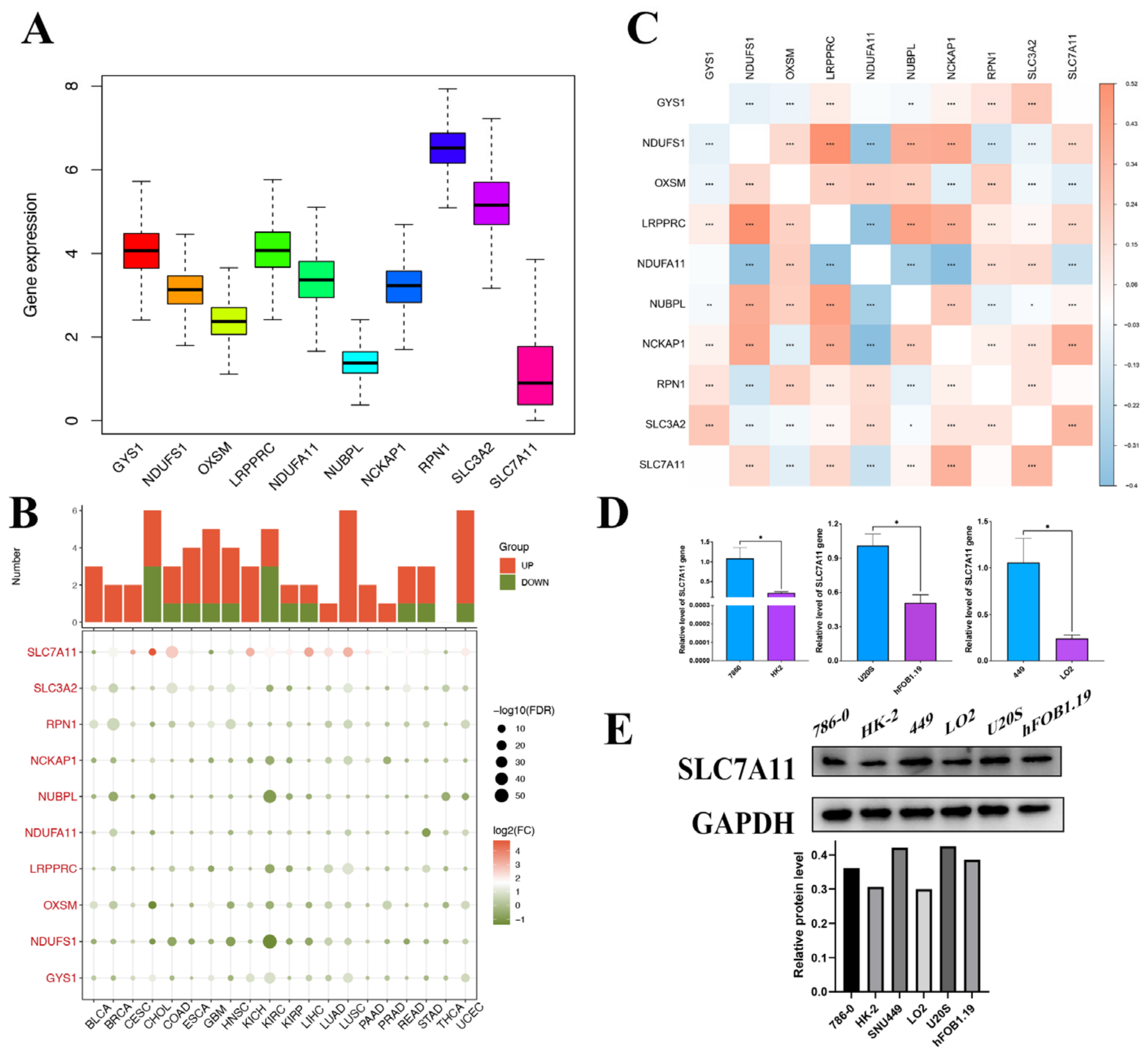
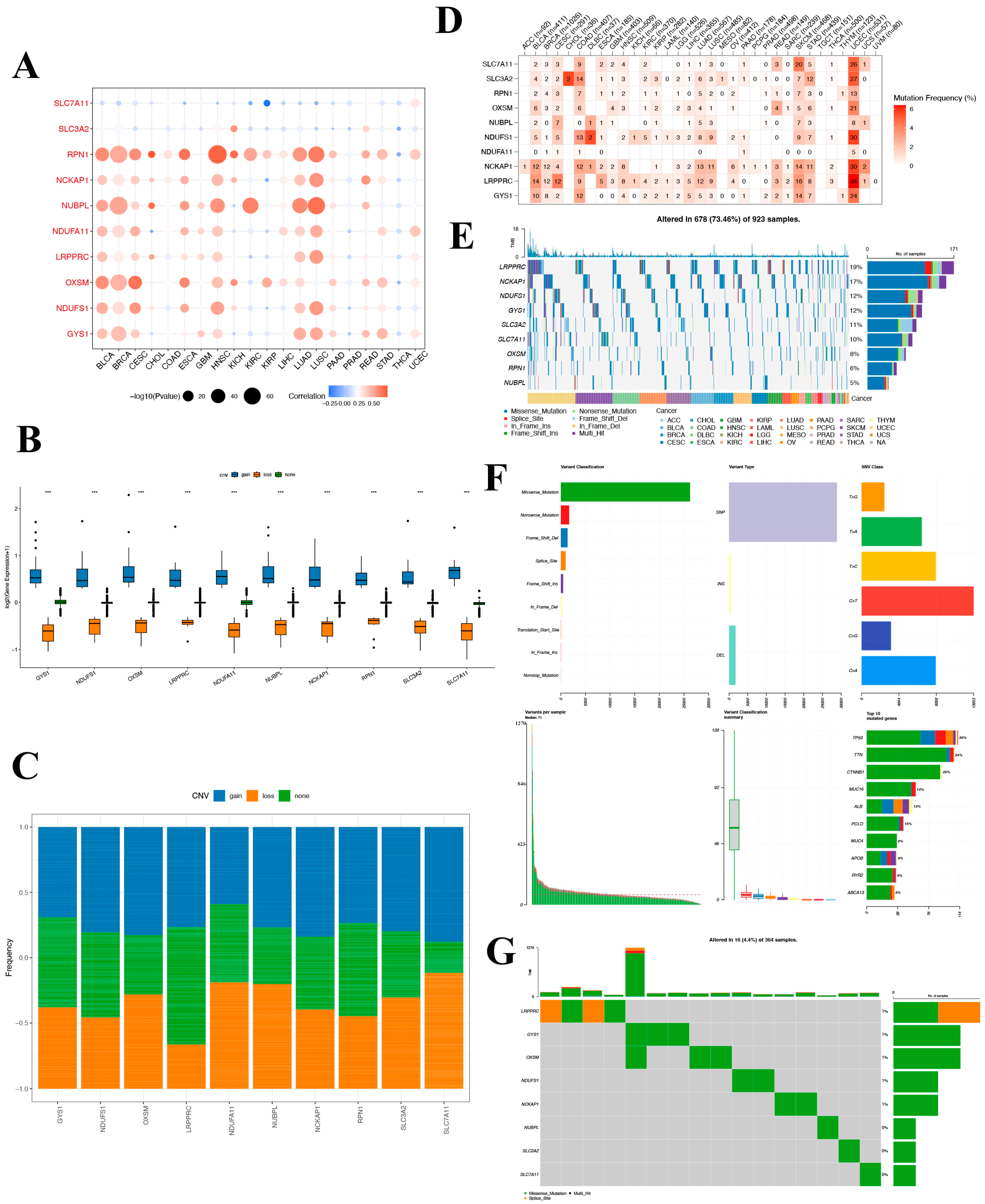
A significant difference in DRG expression was observed in most tumors, despite a lack of normal comparison in certain cancer types. Of note, both SLC7A11 and SLC3A exhibited marked differential expression across almost all cancers. To further elucidate the potential roles of DRGs in tumorigenesis and the mechanism for their aberrant expression, we conducted a thorough analysis of methylation and mutations across pan-cancer settings. In the pan-cancer mutation landscape, DRGs consistently exhibit a high prevalence of CNV and SNV across diverse tumorigenic contexts. However, our study reveals that SLC7A11 infrequently presents with CNV and SNV, especially in hepatocellular carcinoma. DRGs exhibit a negative correlation with DNA promoter methylation across diverse tumor classifications. Particularly, the methylation status of SLC7A11 consistently correlates with a less favorable prognosis pronounced in LIHC. In conjunction with the established framework of disulfidoptosis, the mentioned results emphasize the pivotal function of SLC7A11 and hint at its underlying mechanism. Grounded on the findings of SLC7A11, we suggest a potential therapeutic strategy that might improve clinical outcomes for oncology patients, involving manipulating the methylation profile and modulating disulfidoptosis-driven tumorigenic activities.
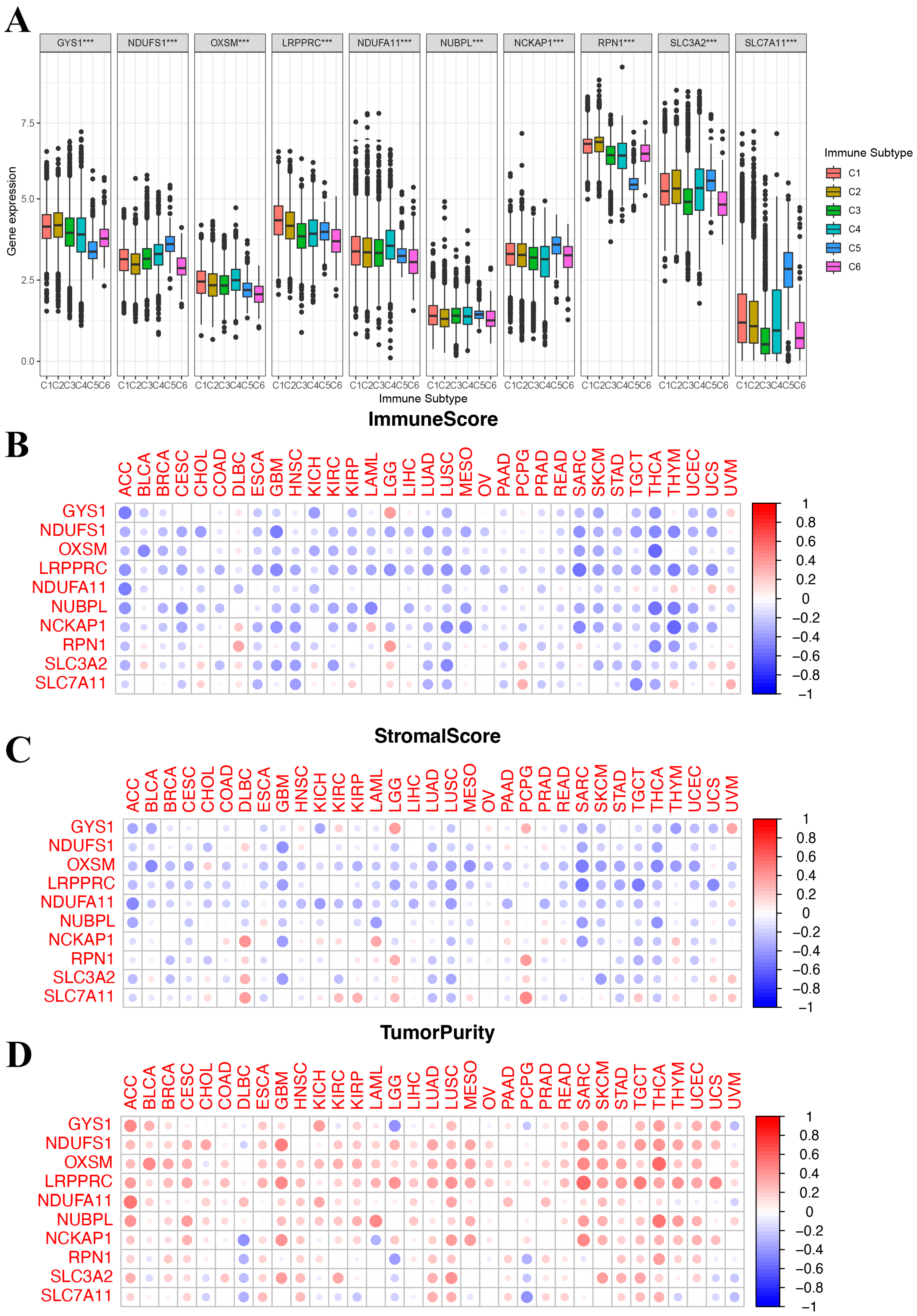
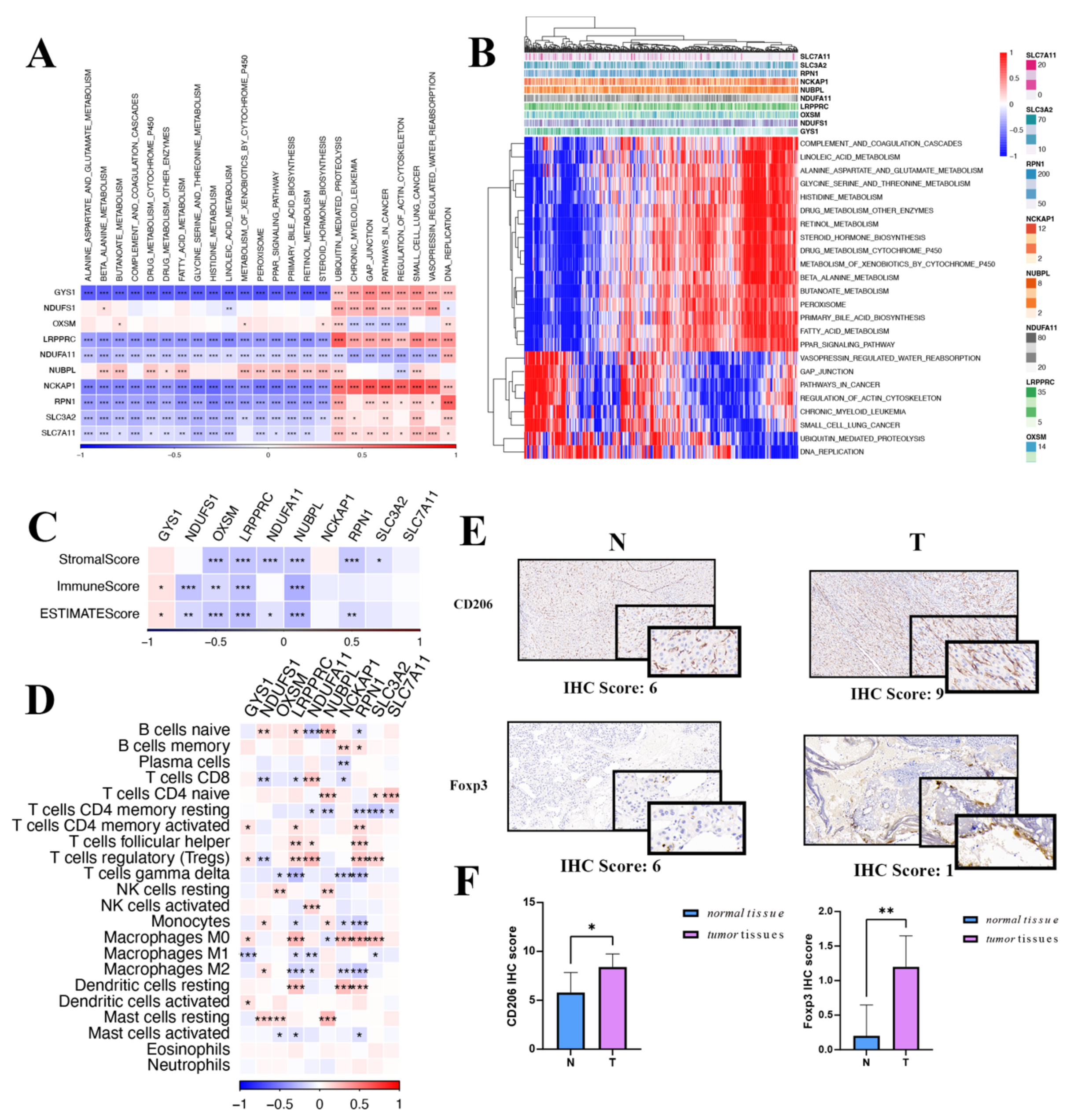
While exploring the relationship between DRGs and the TME, evident variances were found across diverse immune subtypes. The expression of DRGs was inversely correlated with both ImmuneScore and StromalScore and positively correlated with tumor purity, as illustrated by the TME analyses. The correlation analysis between disulfidoptosis and TME, as well as immune infiltration patterns, may unveil a potential scenario where patients with heightened DRG expression might be in an immunologically desert TME state. These findings mentioned above not only elucidate the oncogenic properties of DRGs but also emphasize their prospective significance as therapeutic targets.
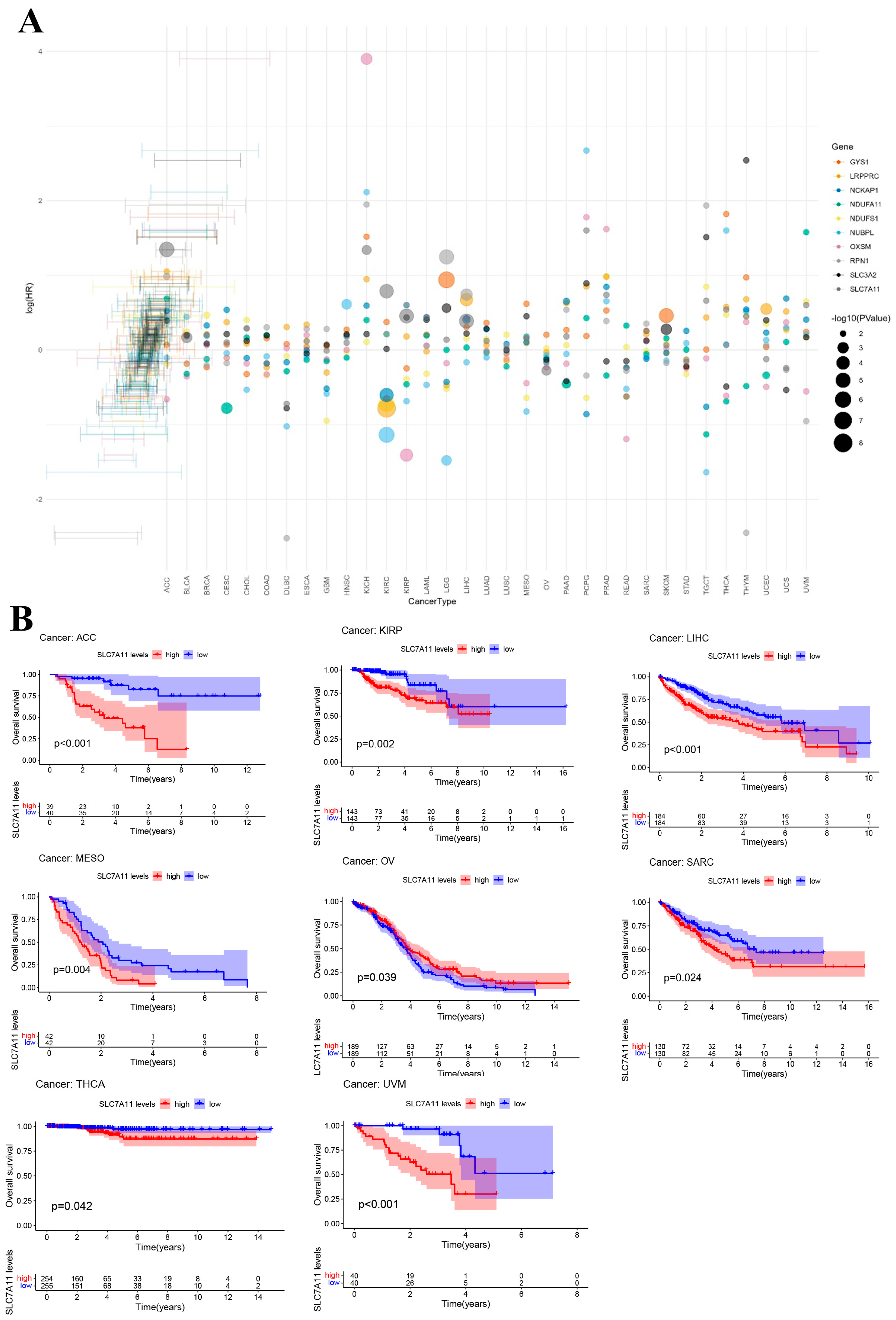
Based on the results from the Cox analysis and KM survival studies, subsequent research primarily focused on LIHC. Among our investigations into hepatocellular carcinoma, single-cell sequencing was notably significant. This advanced approach allowed us to dissect the cellular milieu of HCC and unveil the complicated cellular heterogeneity that often remains concealed in bulk RNA sequencing [
34].
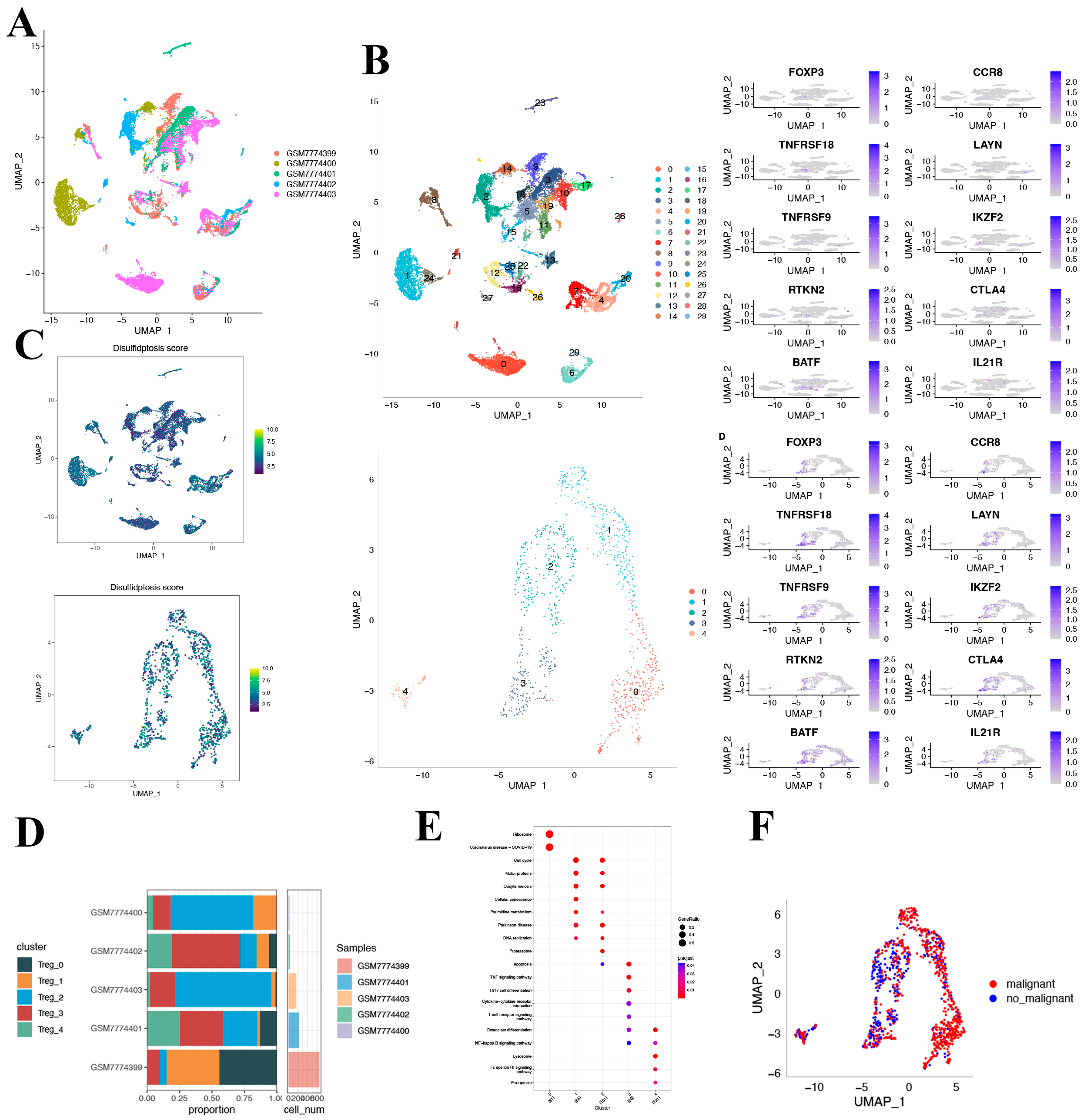
Through log-normalization and dimensionality reduction, our single-cell landscape revealed distinct subgroups within HCC. Each of these subclusters represented a unique cellular identity, characterized by its own molecular signature, which may potentially be associated with their specific functions within the tumor microenvironment.
The Treg cell populations we focused on, especially those expressing marker genes like
FOXP3,
CCR8,
TNFRSF8,
LAYN,
TNFRSF9,
IKZF2,
RTKN2,
CTLA4,
BATF, and
IL21R, played a pivotal role in maintaining immune balance [
21]. Alternatively, Treg cells have an anti-tumor immune role by reducing the immunity of tumor-associated antigen-specific T cells [
35]. Considering both aforementioned points, the expression patterns of these marker genes across different clusters provided not only a snapshot of the T-cell repertoire within HCC but also hinted at the dynamic interplay of immune activation and suppression.
The role of DRGs in this cellular tapestry was another pivotal revelation. Given their known impacts on the survival and death of tumor cells, their differential expression across cellular subpopulations may suggest nuanced roles in determining cellular outcomes. For instance, the distinction of cellular clusters between high and low DRG expression may hint at differing metabolic states, proliferative capacities, and even influences on therapeutic interventions.
Next, we investigated the immune and biological functions of DRGs. Using the GSVA method, we assessed the biological processes associated with the 10 DRGs. The expression of DRGs is primarily positively correlated with several tumoral pathways, DNA replication, and the cell cycle, while it is negatively correlated with tumor metabolism. These findings mentioned that cellular proliferation and metabolism were related to the expression of DRGs, which could potentially act as a prognostic marker for tumor cell progression in HCC. Then, the analysis of TME cell infiltration revealed that the expression of DRGs was negatively correlated with immune-, stromal-, and ESTIMATE scores. Furthermore, it indicated a significant positive correlation between the expression of DRGs and naive CD4 T- and B-cells, Treg, M0 macrophages, resting mast cells, and dendritic cells. Surprisingly, conversely, there was a negative correlation between DRGs and M1 macrophages, M2 macrophages, activated mast cells, and dendritic cells. This observation may provide a potential explanation for the result of the aforementioned immune infiltration analysis. These analyses examining the correlation between disulfidoptosis and TME, as well as immune infiltration patterns, suggest a potential scenario wherein patients exhibiting higher DRG expression may present an immunologically quiescent TME state. These results not only elucidate the oncogenic characteristics of DRGs but also emphasize their prospective significance as therapeutic targets.
HCC, characterized by its distinct molecular pathogenesis and a complex TME, advances rapidly, significantly limiting the efficacy of conventional therapies [
36]. Prior studies have revealed the significantly upregulated expression of SLC7A11 in tumor-associated macrophages (TAM). And knocking out SLC7A11 in these macrophages significantly diminished the infiltration of TAMs and hindered the shift to an M2-like phenotype in HCC tissues, which in turn attenuated tumor growth and metastasis [
37]. Prior investigations have concentrated on the role of ferroptosis in tumor cells, highlighting how SLC7A11-mediated ferroptosis and phenotypic alterations in TAMs profoundly modify the HCC tumor microenvironment and promote tumor proliferation [
38]. Studies have revealed that the silence of SLC7A11 impairs macrophage recruitment and polarization, primarily by inhibiting the secretion of M2 phenotype inducers and cytokines and disrupting the IL-4-driven SOCS3-STAT6-PPAR-γ signaling axis [
14]. Furthermore, ferroptosis in macrophages, driven by SLC7A11, significantly increases PD-L1 expression. The therapeutic efficacy is notably enhanced when SLC7A11-specific knockout in macrophages is combined with anti-PD-L1 therapy, surpassing the outcomes of either approach used independently [
39]. This suggests that a combination of SLC7A11-targeted therapies and immunotherapies may offer superior therapeutic benefits clinically. Our research aims to delve deeper into SLC7A11-mediated disulfidoptosis, underpinning its viability as a target for HCC treatment.
Finally, utilizing CCK8, colony formation, and Transwell assays, we demonstrated that SL7CA11 acts as a driving factor for proliferation, migration, and invasion in hepatocellular carcinoma cell lines.
Our study presented several inherent limitations. Firstly, the data utilized in this article were derived exclusively from the TCGA database alone, without inclusion from our center or external validation in other public databases or centers. Secondly, further explorations in vivo and in vitro are essential to comprehensively understanding the role of DRGs in HCC. And our conclusions also need further animal experiments and subsequent clinical data to solidify our findings. Additionally, experiments using immune checkpoint inhibitors in immunotherapy and second-line therapy are crucial to confirm the potential of DRGs as viable targets in such treatments.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}