
Poly-Gamma-Glutamic Acid Nanopolymer Effect against Bacterial Biofilms: In Vitro and In Vivo Study

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Antibacterial Activity of Ɣ-PGA NPs
2.3. MBIC Technique
2.4. MBEC Technique
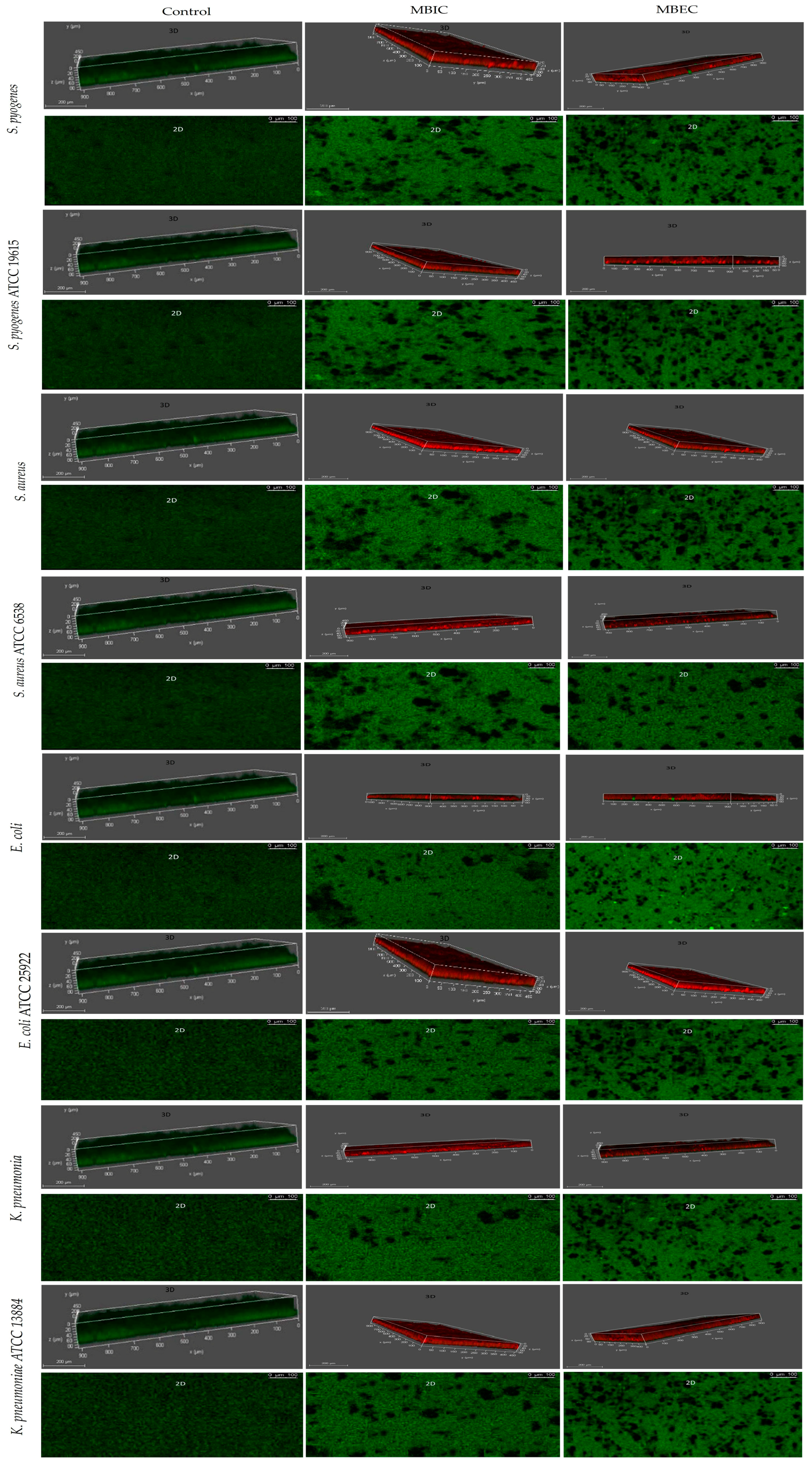
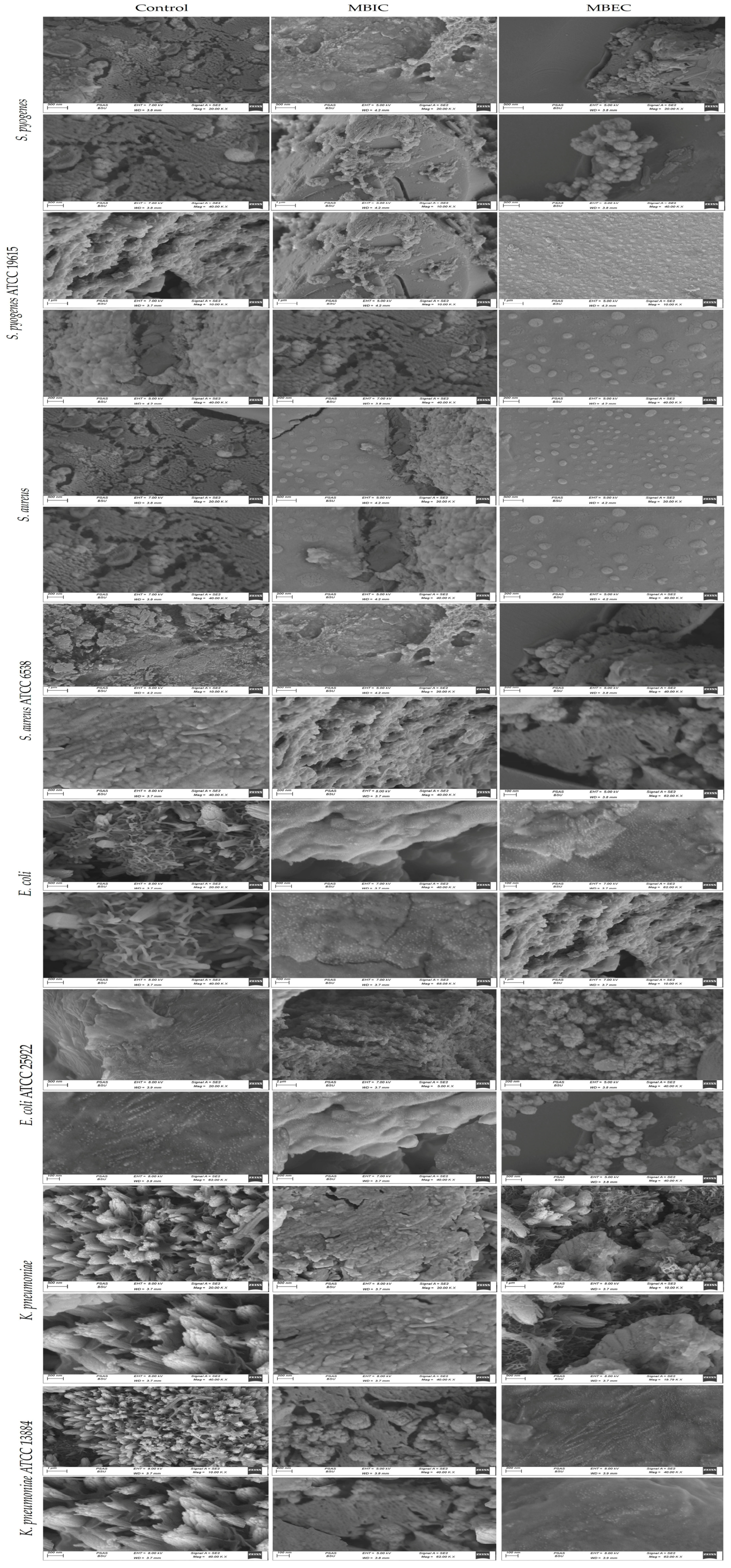
2.5. CLSM and SEM Characterization of the Biofilms
2.6. Image Analysis
2.7. Quantification of Adhered Biofilm Cells on Microtiter Plates (CFU)
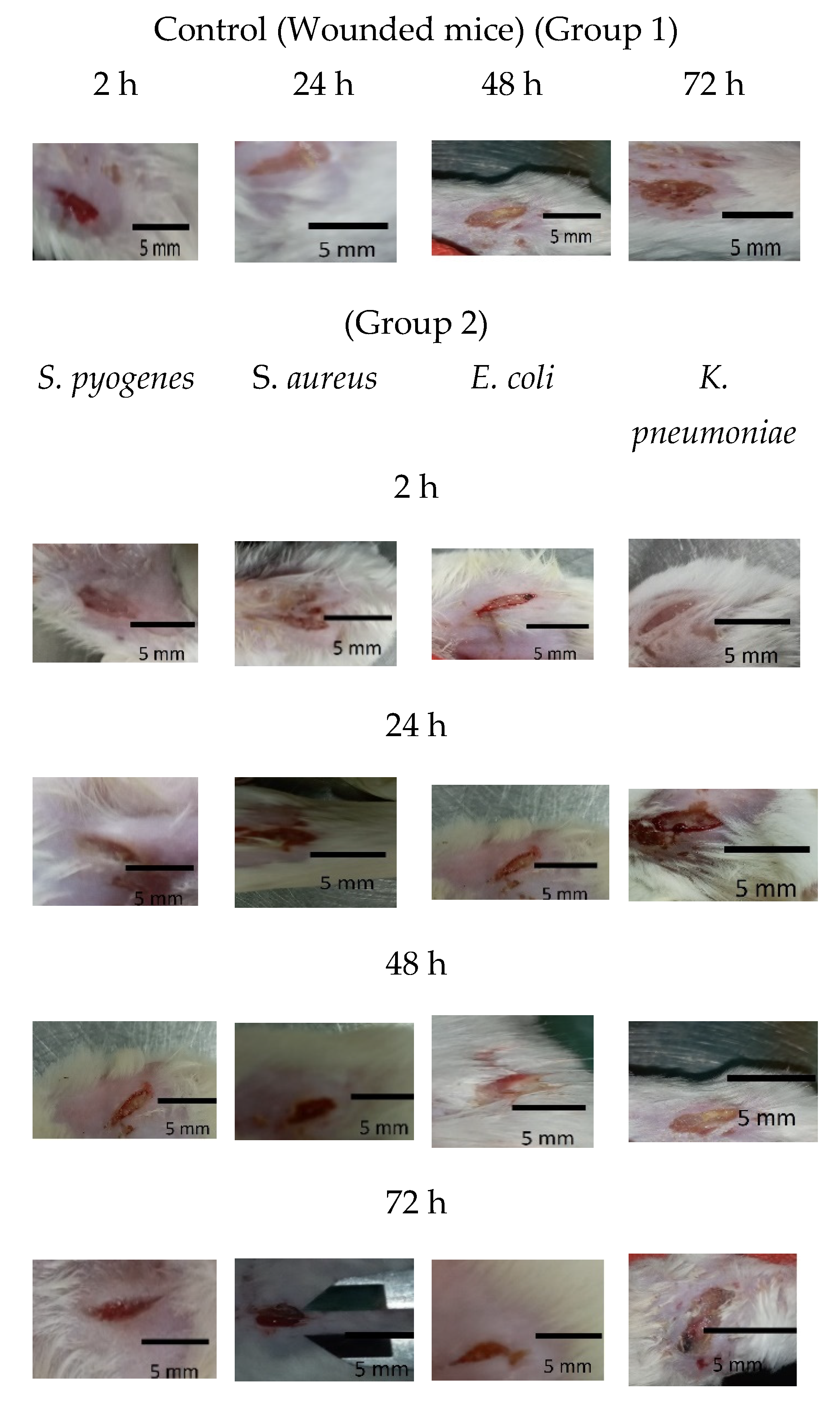
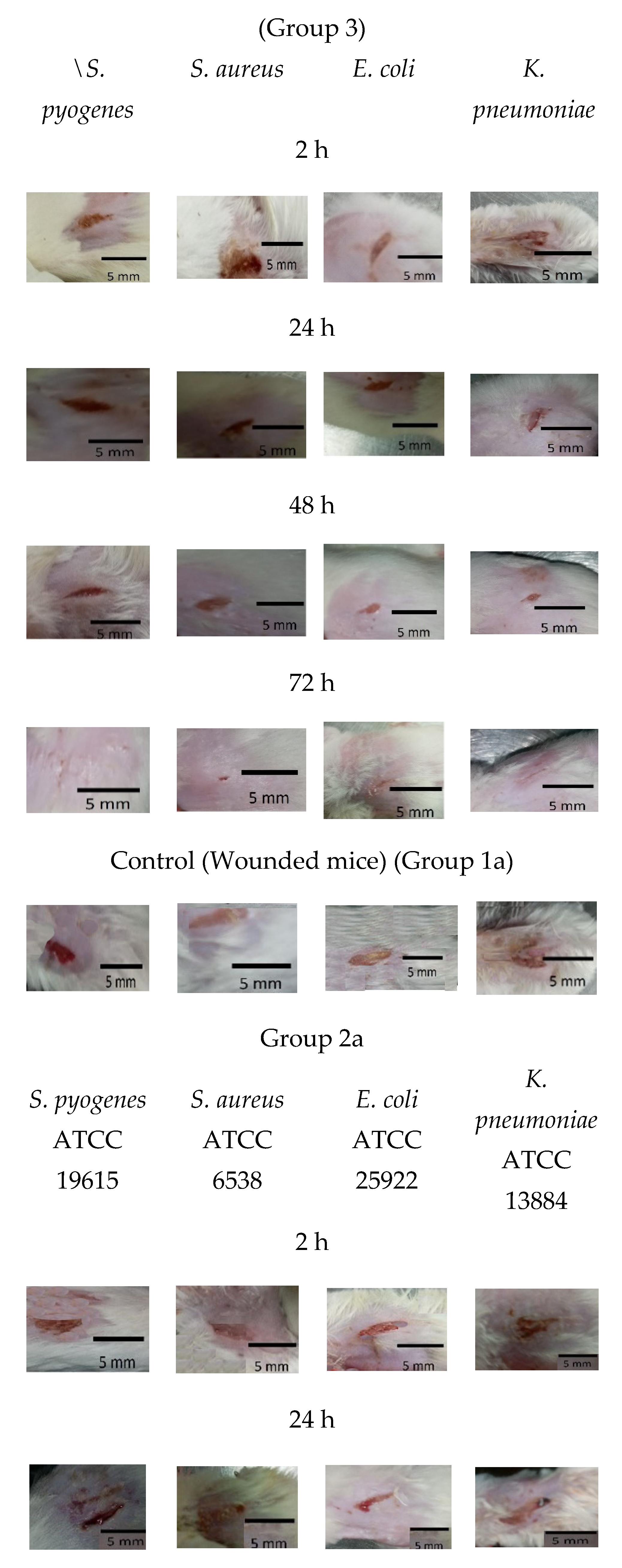
2.8. Antibiofilm Effect of Ɣ-PGA NP against Mice-Infected Wounds
2.9. Statistical Analysis
3. Results and Discussion
3.1. MIC of Ɣ-PGA NPs
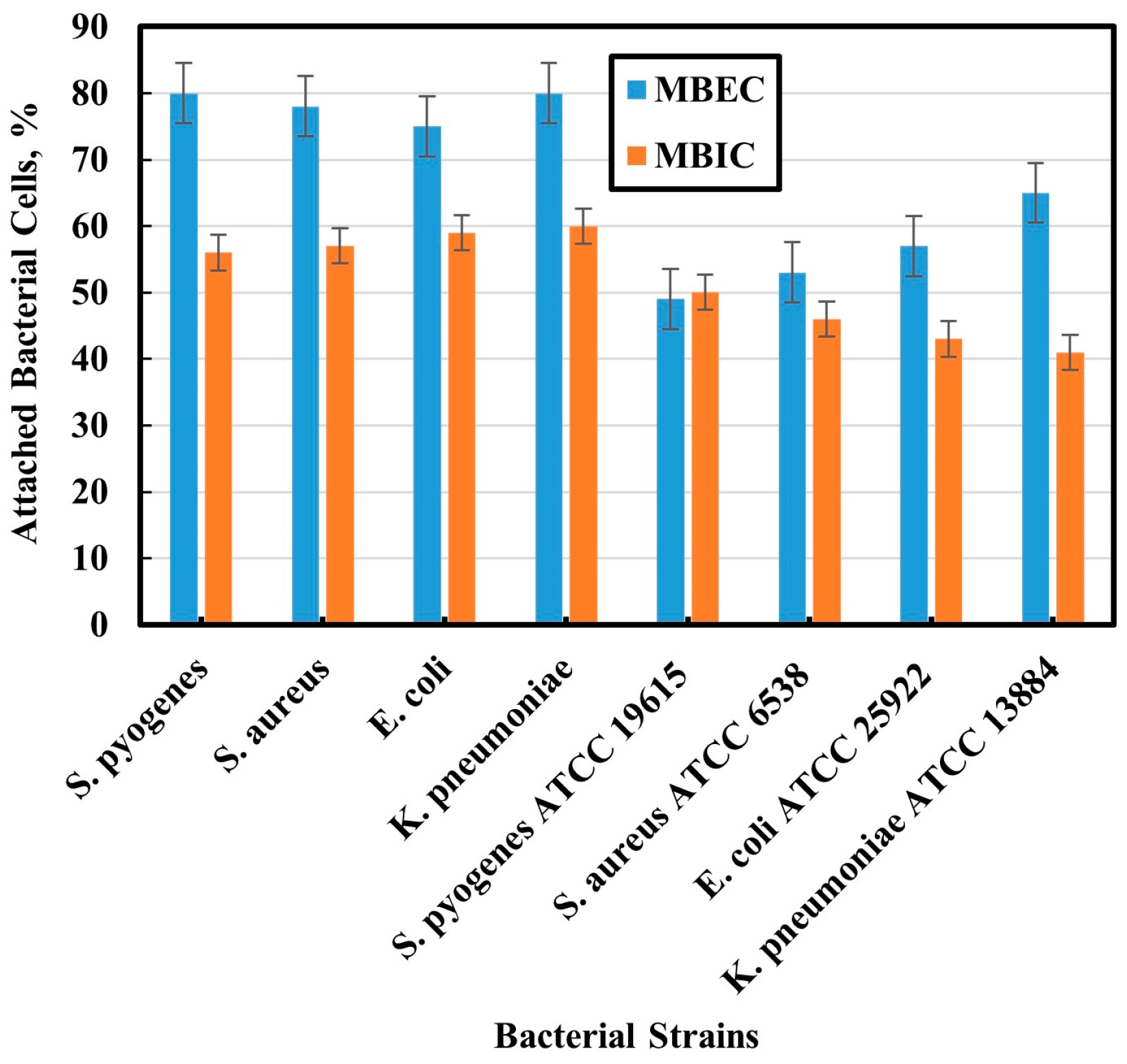
3.2. Biofilm Inhibition and Eradication Assays on Microtiter Plates
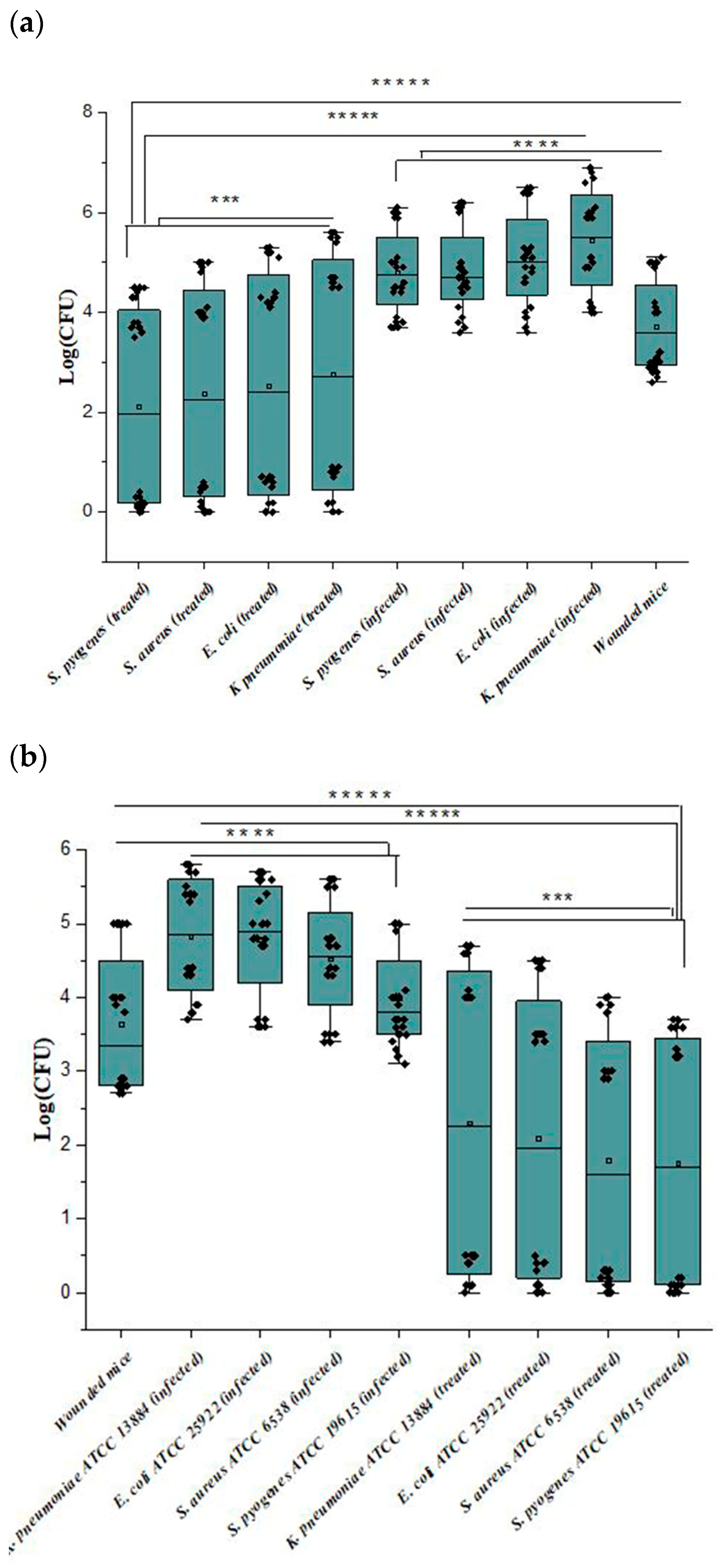
3.3. Assessment of Antibiofilm by Cell Counting
3.4. Analysis of Biofilm Vitality and Thickness by CLSM
3.5. ImageJ Counting
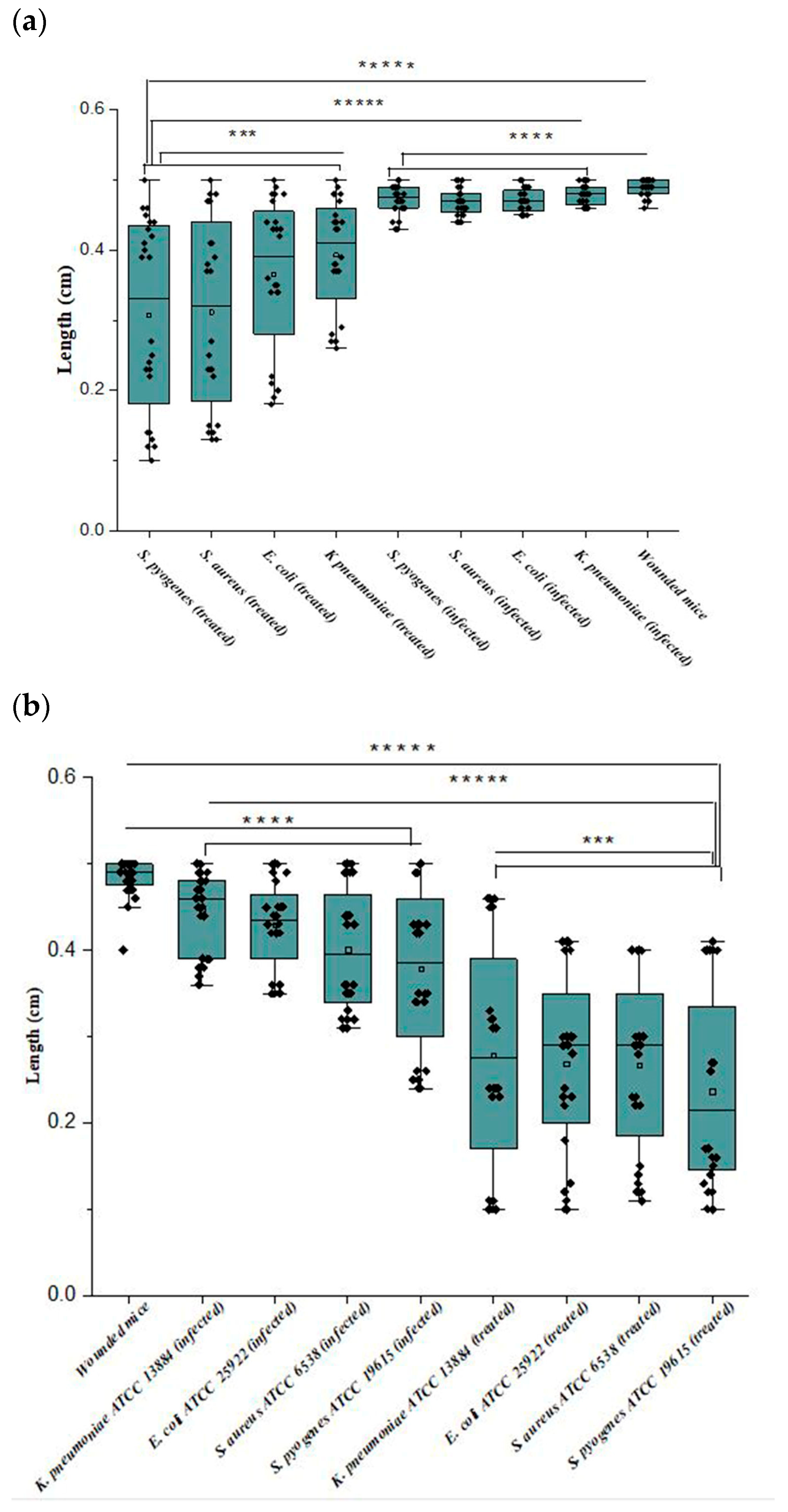
3.6. In Vivo Anti-Biofilm Efficiency of Ɣ-PGA NPs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Worthington, R.J.; Richards, J.J.; Melander, C. Small molecule control of bacterial biofilms. Org. Biomol. Chem. 2012, 10, 7457–7474. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural Anti-biofilm Agents: Strategies to Control Biofilm-Forming Pathogens. Front. Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Thieme, L.; Hartung, A.; Tramm, K.; Klinger-Strobel, M.; Jandt, K.D.; Makarewicz, O.; Pletz, M.W. MBEC versus MBIC: The lack of differentiation between biofilm reducing and inhibitory effects as a current problem in biofilm methodology. Biol. Proced. Online 2019, 21, 18. [Google Scholar] [CrossRef] [PubMed]
- Kulshrestha, S.; Khan, S.; Hasan, S.; Khan, M.E.; Misba, L.; Khan, A.U. Calcium fluoride nanoparticles induced suppression of Streptococcus mutans biofilm: An in vitro and in vivo approach. Appl. Microbiol. Biotechnol. 2016, 100, 1901–1914. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo, J.L.; Patel, R. The challenge of treating biofilm-associated bacterial infections. Clin. Pharmacol. Ther. 2007, 82, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Weerasekera, M.M.; Wijesinghe, G.K.; Jayarathna, T.A.; Gunasekara, C.P.; Fernando, N.; Kottegoda, N.; Samaranayake, L.P. Culture media profoundly affect Candida albicans and Candida tropicalis growth, adhesion and biofilm development. Memórias do Inst. Oswaldo Cruz 2016, 111, 697–702. [Google Scholar] [CrossRef]
- Wolcott, R.D.; Rhoads, D.D.; Bennett, M.E.; Wolcott, B.M.; Gogokhia, L.; Costerton, J.W.; Dowd, S.E. Chronic wounds and the medical biofilm paradigm. J. Wound Care 2010, 19, 45–53. [Google Scholar] [CrossRef]
- Cruz, D.; Shah, S.; Tammela, P. Defining conditions for biofilm inhibition and eradication assays for Gram-positive clinical reference strains. BMC Microbiol. 2018, 18, 173. [Google Scholar] [CrossRef]
- Vaso, O.; Bila, N.M.; Pandolfi, F.; De Vita, D.; Bortolami, M.; Bonatti, J.L.C.; de Moraes Silva, R.A.; Gonçalves, L.N.C.; Tudino, V.; Costi, R.; et al. Evaluation of the Anti-Histoplasma capsulatum Activity of Indole and Nitrofuran Derivatives and Their Pharmacological Safety in Three-Dimensional Cell Cultures. Pharmaceutics 2022, 14, 1043. [Google Scholar] [CrossRef]
- Hughes, G.; Webber, M.A. Novel approaches to the treatment of bacterial biofilm infections. Br. J. Pharmacol. 2017, 174, 2237–2246. [Google Scholar] [CrossRef]
- Oniciuc, E.A.; Cerca, N.; Nicolau, A.I. Compositional analysis of biofilms formed by Staphylococcus aureus isolated from food sources. Front. Microbiol. 2016, 7, 2014–2017. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Tarar, S.M.; Gul, I.; Nawaz, U.; Arshad, M. Challenges of antibiotic resistance biofilms and potential combating strategies: A review. 3 Biotech 2021, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.R. Going beyond the control of quorum-sensing to combat biofilm infections. Antibiotics 2016, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Hasan, N.; Cao, J.; Lee, J.; Hlaing, S.P.; Yoo, J.W. Chitosan-based nitric oxide-releasing dressing for anti-biofilm and in vivo healing activities in MRSA biofilm-infected wounds. Int. J. Biol. Macromol. 2020, 142, 680–692. [Google Scholar] [CrossRef]
- Guo, S.; DiPietro, L.A. Critical review in oral biology & medicine: Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Moran, K.A.; Murray, C.K.; Anderson, E.L. Bacteriology of Blood, Wound, and Sputum Cultures from Non-US Casualties Treated in a Combat Support Hospital in Iraq. Infect. Control Hosp. Epidemiol. 2008, 29, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Kirketerp-Møller, K.; Jensen, P.Ø.; Madsen, K.G.; Phipps, R.; Krogfelt, K.; Høiby, N.; Givskov, M. Why chronic wounds will not heal: A novel hypothesis. Wound Repair Regen. 2008, 16, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef]
- Di Conza, J.A.; Badaracco, A.; Ayala, J.; Rodriguez, C.; Famiglietti, Á.; Gutkind, G.O. β-lactamasas producidas por enterobacterias resistentes a amoxicilina-ácido clavulánico aisladas en Buenos Aires, Argentina: Un nuevo gen blaTEM. Rev. Argent. Microbiol. 2014, 46, 210–217. [Google Scholar] [CrossRef]
- Thompson, M.G.; Truong-Le, V.; Alamneh, Y.A.; Black, C.C.; Anderl, J.; Honnold, C.L.; Pavlicek, R.L.; Abu-Taleb, R.; Wise, M.C.; Hall, E.R.; et al. Evaluation of gallium citrate formulations against a multidrug-resistant strain of Klebsiella pneumoniae in a murine wound model of infection. Antimicrob. Agents Chemother. 2015, 59, 6484–6493. [Google Scholar] [CrossRef]
- Alharbi, N.S.; Khaled, J.M.; Kadaikunnan, S.; Alobaidi, A.S.; Sharafaddin, A.H.; Alyahya, S.A.; Almanaa, T.N.; Alsughayier, M.A.; Shehu, M.R. Prevalence of Escherichia coli strains resistance to antibiotics in wound infections and raw milk. Saudi J. Biol. Sci. 2019, 26, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Kožár, M.; Hamilton, H.; Koščová, J. Types of Wounds and the Prevalence of Bacterial Contamination of Wounds in the Clinical Practice of Small Animals. Folia Vet. 2018, 62, 39–47. [Google Scholar] [CrossRef]
- Vila, J.; Sáez-López, E.; Johnson, J.R.; Römling, U.; Dobrindt, U.; Cantón, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martínez-Medina, M.; et al. Escherichia coli: An old friend with new tidings. FEMS Microbiol. Rev. 2016, 40, 437–463. [Google Scholar] [CrossRef]
- Augustine, R. Skin bioprinting: A novel approach for creating artificial skin from synthetic and natural building blocks. Prog. Biomater. 2018, 7, 77–92. [Google Scholar] [CrossRef]
- Augustine, R.; Rehman, S.R.U.; Ahmed, R.; Zahid, A.A.; Sharifi, M.; Falahati, M.; Hasan, A. Electrospun chitosan membranes containing bioactive and therapeutic agents for enhanced wound healing. Int. J. Biol. Macromol. 2020, 156, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Halim, A.S.; Khoo, T.L.; Shah, S.J. Biologic and synthetic skin substitutes: An overview. Indian J. Plast. Surg. 2010, 43, S23–S28. [Google Scholar] [CrossRef]
- Groeber, F.; Holeiter, M.; Hampel, M.; Hinderer, S.; Schenke-Layland, K. Skin tissue engineering—In vivo and in vitro applications. Adv. Drug Deliv. Rev. 2011, 63, 352–366. [Google Scholar] [CrossRef]
- Doppalapudi, S.; Jain, A.; Khan, W.; Domb, A.J. Biodegradable polymers-an overview. Polym. Adv. Technol. 2014, 25, 427–435. [Google Scholar] [CrossRef]
- Ahmed, M.K.; Zayed, M.A.; El-dek, S.I.; Hady, M.A.; El Sherbiny, D.H.; Uskoković, V. Nanofibrous ε-polycaprolactone scaffolds containing Ag-doped magnetite nanoparticles: Physicochemical characterization and biological testing for wound dressing applications in vitro and in vivo. Bioact. Mater. 2021, 6, 2070–2088. [Google Scholar] [CrossRef]
- Wentao, W.; Tao, Z.; Bulei, S.; Tongchang, Z.; Qicheng, Z.; Fan, W.; Ninglin, Z.; Jian, S.; Ming, Z.; Yi, S. Functionalization of polyvinyl alcohol composite film wrapped in am-ZnO@CuO@Au nanoparticles for antibacterial application and wound healing. Appl. Mater. Today. 2019, 17, 36–44. [Google Scholar] [CrossRef]
- Uskoković, V. Entering the era of nanoscience: Time to be so small. J. Biomed. Nanotechnol. 2013, 9, 1441–1470. [Google Scholar] [CrossRef]
- Hu, P.Y.; Zhao, Y.T.; Zhang, J.; Yu, S.X.; Yan, J.S.; Wang, X.X.; Hu, M.Z.; Xiang, H.F.; Long, Y.Z. In situ melt electrospun polycaprolactone/Fe3O4 nanofibers for magnetic hyperthermia. Mater. Sci. Eng. C 2019, 110, 110708. [Google Scholar] [CrossRef]
- Hao, D.; Zhang, G.; Gong, Y.; Ma, Z. Development and biological evaluation of cerium oxide loaded polycaprolactone dressing on cutaneous wound healing in nursing care. Mater. Lett. 2020, 265, 127401. [Google Scholar] [CrossRef]
- Nuzhina, J.V.; Shtil, A.A.; Prilepskii, A.Y.; Vinogradov, V.V. Preclinical Evaluation and Clinical Translation of Magnetite-Based Nanomedicines. J. Drug Deliv. Sci. Technol. 2019, 54, 101282. [Google Scholar] [CrossRef]
- Moradi, A.; Kheirollahkhani, Y.; Fatahi, P.; Abdollahifar, M.A.; Amini, A.; Naserzadeh, P.; Ashtari, K.; Ghoreishi, S.K.; Chien, S.; Rezaei, F.; et al. An improvement in acute wound healing in mice by the combined application of photobiomodulation and curcumin-loaded iron particles. Lasers Med. Sci. 2019, 34, 779–791. [Google Scholar] [CrossRef] [PubMed]
- El-Dib, F.I.; Mohamed, D.E.; El-Shamy, O.A.A.; Mishrif, M.R. Study the adsorption properties of magnetite nanoparticles in the presence of different synthesized surfactants for heavy metal ions removal. Egypt. J. Pet. 2020, 29, 1–7. [Google Scholar] [CrossRef]
- Cui, H.; Dai, Y.; Lin, L. Enhancing antibacterial efficacy of nisin in pork by poly-γ-glutamic acid/poly-l-lysine nanoparticles encapsulation. J. Food Saf. 2018, 38, e12475. [Google Scholar] [CrossRef]
- Inbaraj, B.S.; Kao, T.H.; Tsai, T.Y.; Chiu, C.P.; Kumar, R.; Chen, B.H. The synthesis and characterization of poly(γ-glutamic acid)-coated magnetite nanoparticles and their effects on antibacterial activity and cytotoxicity. Nanotechnology 2011, 22, 075101. [Google Scholar] [CrossRef]
- Wang, Y.; Dou, C.; He, G.; Ban, L.; Huang, L.; Li, Z.; Gong, J.; Zhang, J.; Yu, P. Biomedical potential of ultrafine Ag nanoparticles coated on poly (gamma-glutamic acid) hydrogel with special reference to wound healing. Nanomaterials 2018, 8, 324. [Google Scholar] [CrossRef]
- Elsayed, E.M.; Farghali, A.A.; Zanaty, M.I.; Abdel-Fattah, M.; Alkhalifah, D.H.M.; Hozzein, W.N.; Mahmoud, A.M. In Vitro Cytotoxicity and Hemolysis Effect of Poly-Gamma-Glutamic Acid Nano-Polymer Biosynthesized Using Some Isolates of Bacillus spp. Microbiol. Res. 2023, 14, 1720–1735. [Google Scholar] [CrossRef]
- Elsayed, E.; Salah, T.; Husseiny, S. Evaluation of Morphological and Ultrastructure Effects of Nanoparticles on Some Microbial Cells; Ain-Shams: Cairo, Egypt, 2017. [Google Scholar]
- Christensen, G.D.; Simpson, W.A.; Bisno, A.L.; Beachey, E.H. Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect. Immun. 1982, 37, 318–326. [Google Scholar] [CrossRef]
- Haney, E.F.; Mansour, S.C.; Hilchie, A.L.; De La Fuente-Núñez, C.; Hancock, R.E.W. High throughput screening methods for assessing antibiofilm and immunomodulatory activities of synthetic peptides. Peptides 2015, 71, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Trimble, M.J.; Cheng, J.T.; Vallé, Q.; Hancock, R.E.W. Critical assessment of methods to quantify biofilm growth and evaluate antibiofilm activity of host defence peptides. Biomolecules 2018, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Mountcastle, S.E.; Vyas, N.; Villapun, V.M.; Cox, S.C.; Jabbari, S.; Sammons, R.L.; Shelton, R.M.; Walmsley, A.D.; Kuehne, S.A. Biofilm viability checker: An open-source tool for automated biofilm viability analysis from confocal microscopy images. NPJ Biofilms Microbiomes 2021, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Lu, F.; Ran, L.; Yu, K.; Xiao, Y.; Li, Z.; Dai, F.; Wu, D.; Lan, G. Imidazole-molecule-capped chitosan–gold nanocomposites with enhanced antimicrobial activity for treating biofilm-related infections. J. Colloid Interface Sci. 2018, 531, 269–281. [Google Scholar] [CrossRef]
- Rosenberg, M.; Visnapuu, M.; Vija, H.; Kisand, V.; Kasemets, K.; Kahru, A.; Ivask, A. Selective antibiofilm properties and biocompatibility of nano-ZnO and nano-ZnO/Ag coated surfaces. Sci. Rep. 2020, 10, 13478. [Google Scholar] [CrossRef]
- Hakkimane, S.S.; Shenoy, V.P.; Gaonkar, S.L.; Bairy, I.; Guru, B.R. Antimycobacterial susceptibility evaluation of rifampicin and isoniazid benz-hydrazone in biodegradable polymeric nanoparticles against Mycobacterium tuberculosis H37Rv strain. Int. J. Nanomed. 2018, 13, 4303–4318. [Google Scholar] [CrossRef]
- Saliha Durak, S.D.; Tulin Arasoglu, S.D.; Ates, S.C. Enhanced antibacterial and antiparasitic activity of multifunctional polymeric nanoparticles. Nanotechnology 2020, 31, 175705. [Google Scholar] [CrossRef]
- El-Sayed, E.M.; Salah, T.A.; Husseiny, S. Biosynthesized silver nanoparticles from new bacterial isolate and studying their effect on dermatophyte fungi. J. Sci. Res. Sci. 2017, 34, 123–141. [Google Scholar]
- Mohammadinia, M.; Rahmani, S.; Eslami, G.; Ghassemi-Broumand, M.; Aghazadh Amiri, M.; Aghaie, G.; Tabatabaee, S.M.; Taheri, S.; Behgozin, A. Contact lens disinfecting solutions antibacterial efficacy: Comparison between clinical isolates and the standard ISO ATCC strains of Pseudomonas aeruginosa and Staphylococcus aureus. Eye 2012, 26, 327–330. [Google Scholar] [CrossRef]
- Shariati, A.; Asadian, E.; Fallah, F.; Azimi, T.; Hashemi, A.; Yasbolaghi Sharahi, J.; Taati Moghadam, M. Evaluation of nano-curcumin effects on expression levels of virulence genes and biofilm production of multidrug-resistant Pseudomonas aeruginosa isolated from burn wound infection in Tehran, Iran. Infect. Drug Resist. 2019, 12, 2223–2235. [Google Scholar] [CrossRef] [PubMed]
- Fadwa, A.O.; Alkoblan, D.K.; Mateen, A.; Albarag, A.M. Synergistic effects of zinc oxide nanoparticles and various antibiotics combination against Pseudomonas aeruginosa clinically isolated bacterial strains. Saudi J. Biol. Sci. 2021, 28, 928–935. [Google Scholar] [CrossRef]
- Ghosh, S.; Amariei, G.; Mosquera, M.E.G.; Rosal, R. Conjugated polymer nanostructures displaying highly photoactivated antimicrobial and antibiofilm functionalities. J. Mater. Chem. B 2021, 9, 4390–4399. [Google Scholar] [CrossRef]
- Su, C.H.; Velusamy, P.; Kumar, G.V.; Adhikary, S.; Pandian, K.; Anbu, P. Studies of antibacterial efficacy of different biopolymer protected silver nanoparticles synthesized under reflux condition. J. Mol. Struct. 2017, 1128, 718–723. [Google Scholar] [CrossRef]
- Cheow, W.S.; Chang, M.W.; Hadinoto, K. Antibacterial efficacy of inhalable antibiotic-encapsulated biodegradable polymeric nanoparticles against E. coli biofilm cells. J. Biomed. Nanotechnol. 2010, 6, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Kuaté Tokam, C.R.; Bisso Ndezo, B.; Boulens, N.; Allémann, E.; Delie, F.; Dzoyem, J.P. Antibiofilm Activity and Synergistic Effects of Thymol-Loaded Poly (Lactic-Co-Glycolic Acid) Nanoparticles with Amikacin against Four Salmonella enterica Serovars. Can. J. Infect. Dis. Med. Microbiol. 2023, 2023, 7274309. [Google Scholar] [CrossRef]
- Hemmati, F.; Salehi, R.; Ghotaslou, R.; Kafil, H.S.; Hasani, A.; Gholizadeh, P.; Rezaee, M.A. The assessment of antibiofilm activity of chitosan-zinc oxide-gentamicin nanocomposite on Pseudomonas aeruginosa and Staphylococcus aureus. Int. J. Biol. Macromol. 2020, 163, 2248–2258. [Google Scholar] [CrossRef]
- Tassew, D.; Mechesso, A.; Park, N.; Song, J.; Shur, J.; Park, S. Biofilm formation and determination of minimum biofilm eradication concentration of antibiotics in Mycoplasma hyopneumoniae Dereje. J. Vet. Med. Sci. 2017, 79, 1716–1720. [Google Scholar] [CrossRef] [PubMed]
- Ansarifard, E.; Zareshahrabadi, Z.; Sarafraz, N.; Zomorodian, K. Evaluation of Antimicrobial and Antibiofilm Activities of Copper Oxide Nanoparticles within Soft Denture Liners against Oral Pathogens. Bioinorg. Chem. Appl. 2021, 2021, 9939275. [Google Scholar] [CrossRef]
- Kragh, K.N.; Alhede, M.; Kvich, L.; Bjarnsholt, T. Into the well—A close look at the complex structures of a microtiter biofilm and the crystal violet assay. Biofilm 2019, 1, 100006. [Google Scholar] [CrossRef]
- Costa, E.M.; Silva, S.; Vicente, S.; Neto, C.; Castro, P.M.; Veiga, M.; Madureira, R.; Tavaria, F.; Pintado, M.M. Chitosan nanoparticles as alternative anti-staphylococci agents: Bactericidal, antibiofilm and antiadhesive effects. Mater. Sci. Eng. C 2017, 79, 221–226. [Google Scholar] [CrossRef]
- Santhosh, S.M.; Natarajan, K. Antibiofilm activity of epoxy/Ag-TiO2 polymer nanocomposite coatings against Staphylococcus aureus and Escherichia coli. Coatings 2015, 5, 95–114. [Google Scholar] [CrossRef]
- Ali, S.; Ansari, M.A.; Alzohairy, M.A.; Alomary, M.N.; AlYahya, S.; Jalal, M.; Khan, H.M.; Asiri, S.M.M.; Ahmad, W.; Mahdi, A.A.; et al. Biogenic Gold Nanoparticles as Potent Antibacterial Pseudomonas aeruginosa. Antibiotics 2020, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, S.; Malaikozhundan, B.; Parthasarathy, A.; Saravanakumar, K.; Wang, M.H.; Vaseeharan, B. Nano Biomedical Potential of Biopolymer Chitosan-Capped Silver Nanoparticles with Special Reference to Antibacterial, Antibiofilm, Anticoagulant and Wound Dressing Material. J. Clust. Sci. 2020, 31, 355–366. [Google Scholar] [CrossRef]
- Ansari, M.A.; Khan, H.M.; Khan, A.A.; Cameotra, S.S.; Pal, R. Antibiofilm efficacy of silver nanoparticles against biofilm of extended spectrum β-lactamase isolates of Escherichia coli and Klebsiella pneumoniae. Appl. Nanosci. 2014, 7, 859–868. [Google Scholar] [CrossRef]
- Ali, S.G.; Ansari, M.A.; Khan, H.M.; Jalal, M.; Mahdi, A.A.; Cameotra, S.S. Antibacterial and Antibiofilm Potential of Green Synthesized Silver Nanoparticles against Imipenem Resistant Clinical Isolates of P. aeruginosa. Bionanoscience 2018, 8, 544–553. [Google Scholar] [CrossRef]
- Ansari, M.A.; Khan, H.M.; Khan, A.A.; Cameotra, S.S.; Alzohairy, M.A. Anti-biofilm efficacy of silver nanoparticles against MRSA and MRSE isolated from wounds in a tertiary care hospital. Indian J. Med. Microbiol. 2015, 33, 101–109. [Google Scholar] [CrossRef]
- Nair, N.; James, B.; Devadathan, A.; Johny, M.; Mathew, J.; Jacob, J. Comparative Evaluation of Antibiofilm Efficacy of Chitosan Nanoparticle- and Zinc Oxide Nanoparticle-Incorporated Calcium Hydroxide-Based Sealer: An in vitro Study. Contemp. Clin. Dent. 2018, 9, 434. [Google Scholar] [CrossRef]
- Ma, S.; Moser, D.; Han, F.; Leonhard, M.; Schneider-Stickler, B.; Tan, Y. Preparation and antibiofilm studies of curcumin loaded chitosan nanoparticles against polymicrobial biofilms of Candida albicans and Staphylococcus aureus. Carbohydr. Polym. 2020, 241, 116254. [Google Scholar] [CrossRef]
- Dorgham, R.A.; Abd Al Moaty, M.N.; Chong, K.P.; Elwakil, B.H. Molasses-Silver Nanoparticles: Synthesis, Optimization, Characterization, and Antibiofilm Activity. Int. J. Mol. Sci. 2022, 23, 10243. [Google Scholar] [CrossRef]
- El-Telbany, M.; Mohamed, A.A.; Yahya, G.; Abdelghafar, A.; Abdel-Halim, M.S.; Saber, S.; Alfaleh, M.A.; Mohamed, A.H.; Abdelrahman, F.; Fathey, H.A.; et al. Combination of Meropenem and Zinc Oxide Nanoparticles; Antimicrobial Synergism, Exaggerated Antibiofilm Activity, and Efficient Therapeutic Strategy against Bacterial Keratitis. Antibiotics 2022, 11, 1374. [Google Scholar] [CrossRef]
- Raouf, M.; Essa, S.; El Achy, S.; Essawy, M.; Rafik, S.; Baddour, M. Evaluation of Combined Ciprofloxacin and azithromycin free and nano formulations to control biofilm producing Pseudomonas aeruginosa isolated from burn wounds. Indian J. Med. Microbiol. 2021, 39, 81–87. [Google Scholar] [CrossRef]
- Miglani, S.; Tani-Ishii, N. Biosynthesized selenium nanoparticles: Characterization, antimicrobial, and antibiofilm activity against Enterococcus faecalis. PeerJ 2021, 9, 2021. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Vyas, V.; Patil, R.; Sharma, V.; Scopelliti, P.E.; Bongiorno, G.; Podesta, A.; Lenardi, C.; Gade, W.N.; Milani, P. Quantitative characterization of the influence of the nanoscale morphology of nanostructured surfaces on bacterial adhesion and biofilm formation. PLoS ONE 2011, 6, e25029. [Google Scholar] [CrossRef]
- Raghuwanshi, N.; Kumari, P.; Srivastava, A.K.; Vashisth, P.; Yadav, T.C.; Prasad, R.; Pruthi, V. Synergistic effects of Woodfordia fruticosa gold nanoparticles in preventing microbial adhesion and accelerating wound healing in Wistar albino rats in vivo. Mater. Sci. Eng. C 2017, 80, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Chari, N.; Felix, L.O.; Davoodbasha, M.A.; Sulaiman, A.; Nooruddin, T. In vitro and in vivo antibiofilm effect of copper nanoparticles against aquaculture pathogens. Biocatal. Agric. Biotechnol. 2017, 10, 336–341. [Google Scholar] [CrossRef]
- Muthamil, S.; Devi, V.A.; Balasubramaniam, B.; Balamurugan, K.; Pandian, S.K. Green synthesized silver nanoparticles demonstrating enhanced in vitro and in vivo antibiofilm activity against Candida spp. J. Basic Microbiol. 2018, 58, 343–357. [Google Scholar] [CrossRef]
- Cheng, H.; Li, Y.; Huo, K.; Gao, B.; Xiong, W. Long-lasting in vivo and in vitro antibacterial ability of nanostructured titania coating incorporated with silver nanoparticles. J. Biomed. Mater. Res. Part A 2014, 102, 3488–3499. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | MIC (μg/mL) | |
|---|---|---|
| Ɣ-PGA NPs | Standard (Gentamicin) | |
| K. pneumoniae | 3.2 | 12.8 |
| E. coli | 1.6 | 0.8 |
| S. aureus | 0.4 | 0.4 |
| S. pyogenes | 0.2 | 0.2 |
| K. pneumoniae ATCC 13884 | 1.6 | 3.2 |
| E. coli ATCC 25922 | 0.8 | 1.6 |
| S. aureus ATCC 6538 | 0.2 | 0.4 |
| S. pyogenes ATCC 19615 | 0.2 | 0.4 |
| Strain | Time (h) | MBEC (μg/mL) | CFU/μL | MBIC (μg/mL) | CFU/μL |
|---|---|---|---|---|---|
| K. pneumoniae | 2 | 51.2 ± 0.02 | 220 ± 0.09 | 12.8 ± 0.01 | 160 ± 0.005 |
| 4 | 51.2 ± 0.10 | 130 ± 0.07 | 6.4 ± 0.02 | 110 ± 0.006 | |
| 6 | 12.8 ± 0.01 | 110 ± 0.06 | 1.6 ± 0.022 | 80 ± 0.02 | |
| 8 | 6.4 ± 0.20 | 20 ± 0.03 | 0.8 ± 0.002 | 30 ± 0.008 | |
| 12 | 3.2 ± 0.025 | 2 ± 0.02 | 0.4 ± 0.01 | 2 ± 0.02 | |
| 24 | 1.6 ± 0.012 | 2 ± 0.02 | 0.4 ± 0.01 | 2 ± 0.10 | |
| 48 | 1.6 ± 0.022 | Nil | 0.2 ± 0.02 | Nil | |
| K. pneumoniae ATCC 13884 | 2 | 25.6 ± 0.10 | 200 ± 0.02 | 12.8 ± 0.10 | 130 ± 0.01 |
| 4 | 12.8 ± 0.01 | 190 ± 0.24 | 6.4 ± 0.03 | 110 ± 0.02 | |
| 6 | 12.8 ± 0.02 | 120 ± 0.24 | 0.8 ± 0.02 | 80 ± 0.01 | |
| 8 | 3.2 ± 0.20 | 10 ± 0.02 | 0.8 ± 0.10 | 2 ± 0.01 | |
| 12 | 3.2 ± 0.20 | 2 ± 0.01 | 0.4 ± 0.04 | Nil | |
| 24 | 1.6 ± 0.30 | Nil | 0.2 ± 0.10 | Nil | |
| 48 | 0.8 ± 0.01 | Nil | 0.2 ± 0.025 | Nil | |
| E. coli | 2 | 51.2 ± 0.015 | 90 ± 0.07 | 12.8 ± 0.014 | 150 ± 0.08 |
| 4 | 51.2 ± 0.01 | 30 ± 0.06 | 3.2 ± 0.013 | 100 ± 0.07 | |
| 6 | 12.8 ± 0.02 | 3 ± 0.03 | 1.6 ± 0.021 | 50 ± 0.009 | |
| 8 | 6.4 ± 0.20 | 2 ± 0.03 | 0.8 ± 0.02 | 3 ± 0.11 | |
| 12 | 1.6 ± 0.21 | 2 ± 0.01 | 0.4 ± 0.22 | 2 ± 0.11 | |
| 24 | 0.8 ± 0.016 | Nil | 0.2 ± 0.21 | Nil | |
| 48 | 0.8 ± 0.021 | Nil | 0.2 ± 0.16 | Nil | |
| E. coli ATCC 25922 | 2 | 25.6 ± 0.12 | 80 ± 0.001 | 12.8 ± 0.015 | 130 ± 0.013 |
| 4 | 12.8 ± 0.01 | 30 ± 0.009 | 6.4 ± 0.3 | 110 ± 0.06 | |
| 6 | 3.2 ± 0.01 | 10 ± 0.007 | 0.8 ± 0.03 | 30 ± 0.002 | |
| 8 | 1.6 ± 0.03 | 2 ± 0.005 | 0.4 ± 0.02 | 2 ± 0.0014 | |
| 12 | 1.6 ± 0.02 | 2 ± 0.21 | 0.2 ± 0.01 | Nil | |
| 24 | 0.8 ± 0.10 | Nil | 0.2 ± 0.10 | Nil | |
| 48 | 0.8 ± 0.02 | Nil | 0.1 ± 0.016 | Nil | |
| S. aureus | 2 | 51.2 ± 0.15 | 110 ± 0.04 | 6.4 ± 0.01 | 130 ± 0.19 |
| 4 | 51.2 ± 0.13 | 60 ± 0.04 | 3.2 ± 0.02 | 50 ± 0.10 | |
| 6 | 6.4 ± 0.10 | 20 ± 0.10 | 0.8 ± 0.02 | 3 ± 0.14 | |
| 8 | 3.2 ± 0.01 | 3 ± 0.01 | 0.4 ± 0.24 | 2 ± 0.11 | |
| 12 | 1.6 ± 0.02 | 2 ± 0.03 | 0.4 ± 0.02 | Nil | |
| 24 | 0.8 ± 0.24 | Nil | 0.2 ± 0.01 | Nil | |
| 48 | 0.4 ± 0.02 | Nil | 0.2 ± 0.11 | Nil | |
| S. aureus ATCC 6538 | 2 | 12.8 ± 0.02 | 90 ± 0.003 | 6.4 ± 0.02 | 90 ± 0.10 |
| 4 | 12.8 ± 0.01 | 60 ± 0.002 | 3.2 ± 0.013 | 50 ± 0.20 | |
| 6 | 3.2 ± 0.03 | 10 ± 0.001 | 0.4 ± 0.013 | 2 ± 0.21 | |
| 8 | 3.2 ± 0.02 | 8 ± 0.11 | 0.2 ± 0.02 | 2 ± 0.03 | |
| 12 | 1.6 ± 0.02 | 3 ± 0.01 | 0.2 ± 0.01 | Nil | |
| 24 | 0.4 ± 0.03 | Nil | 0.2 ± 0.03 | Nil | |
| 48 | 0.4 ± 0.02 | Nil | 0.1 ± 0.12 | Nil | |
| S. pyogenes | 2 | 51.2 ± 0.12 | 50 ± 0.06 | 6.4 ± 0.01 | 60 ± 0.12 |
| 4 | 25.6 ± 0.02 | 30 ± 0.05 | 1.6 ± 0.01 | 10 ± 0.01 | |
| 6 | 6.4 ± 0.015 | 3 ± 0.01 | 0.8 ± 0.02 | 2 ± 0.11 | |
| 8 | 3.2 ± 0.014 | 2 ± 19 | 0.4 ± 0.11 | Nil | |
| 12 | 1.6 ± 0.02 | Nil | 0.2 ± 0.12 | Nil | |
| 24 | 0.4 ± 0.20 | Nil | 0.2 ± 0.11 | Nil | |
| 48 | 0.2 ± 0.24 | Nil | 0.1 ± 0.002 | Nil | |
| S. pyogenes ATCC 19615 | 2 | 6.4 ± 0.24 | 50 ± 0.01 | 3.2 ± 0.0012 | 50 ± 0.002 |
| 4 | 6.4 ± 0.20 | 40 ± 0.12 | 1.6 ± 0.11 | 30 ± 0.12 | |
| 6 | 3.2 ± 0.2 | 30 ± 0.001 | 0.8 ± 0.2 | 10 ± 0.11 | |
| 8 | 3.2 ± 0.21 | 2 ± 0.21 | 0.4 ± 0.24 | 2 ± 0.0013 | |
| 12 | 0.8 ± 0.002 | 2 ± 0.11 | 0.2 ± 0.03 | Nil | |
| 24 | 0.4 ± 0.24 | Nil | 0.1 ± 0.2 | Nil | |
| 48 | 0.2 ± 0.22 | Nil | 0.1 ± 0.22 | Nil |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsayed, E.M.; Farghali, A.A.; Zanaty, M.I.; Abdel-Fattah, M.; Alkhalifah, D.H.M.; Hozzein, W.N.; Mahmoud, A.M. Poly-Gamma-Glutamic Acid Nanopolymer Effect against Bacterial Biofilms: In Vitro and In Vivo Study. Biomedicines 2024, 12, 251. https://doi.org/10.3390/biomedicines12020251
Elsayed EM, Farghali AA, Zanaty MI, Abdel-Fattah M, Alkhalifah DHM, Hozzein WN, Mahmoud AM. Poly-Gamma-Glutamic Acid Nanopolymer Effect against Bacterial Biofilms: In Vitro and In Vivo Study. Biomedicines. 2024; 12(2):251. https://doi.org/10.3390/biomedicines12020251
Chicago/Turabian StyleElsayed, Eman M., Ahmed A. Farghali, Mohamed I. Zanaty, Medhat Abdel-Fattah, Dalal Hussien M. Alkhalifah, Wael N. Hozzein, and Ahmed M. Mahmoud. 2024. "Poly-Gamma-Glutamic Acid Nanopolymer Effect against Bacterial Biofilms: In Vitro and In Vivo Study" Biomedicines 12, no. 2: 251. https://doi.org/10.3390/biomedicines12020251
APA StyleElsayed, E. M., Farghali, A. A., Zanaty, M. I., Abdel-Fattah, M., Alkhalifah, D. H. M., Hozzein, W. N., & Mahmoud, A. M. (2024). Poly-Gamma-Glutamic Acid Nanopolymer Effect against Bacterial Biofilms: In Vitro and In Vivo Study. Biomedicines, 12(2), 251. https://doi.org/10.3390/biomedicines12020251