

Extracellular Vesicles from Lung Adenocarcinoma Cells Induce Activation of Different Cancer-Associated Fibroblast Subtypes

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture Conditions
2.2. Extracellular Vesicle Collection and Isolation
2.3. Nanoparticle Tracking Analysis (NTA) and Western Blot for EV Characterization
2.4. Coculture of Fibroblasts with EVs
2.5. Western Blot for CAF Activation
2.6. RNA Extraction and RNA Sequencing
2.7. Data Analysis of RNA Sequencing Data
2.8. Quantitative PCR
2.9. ELISA for EMT Candidates
3. Results
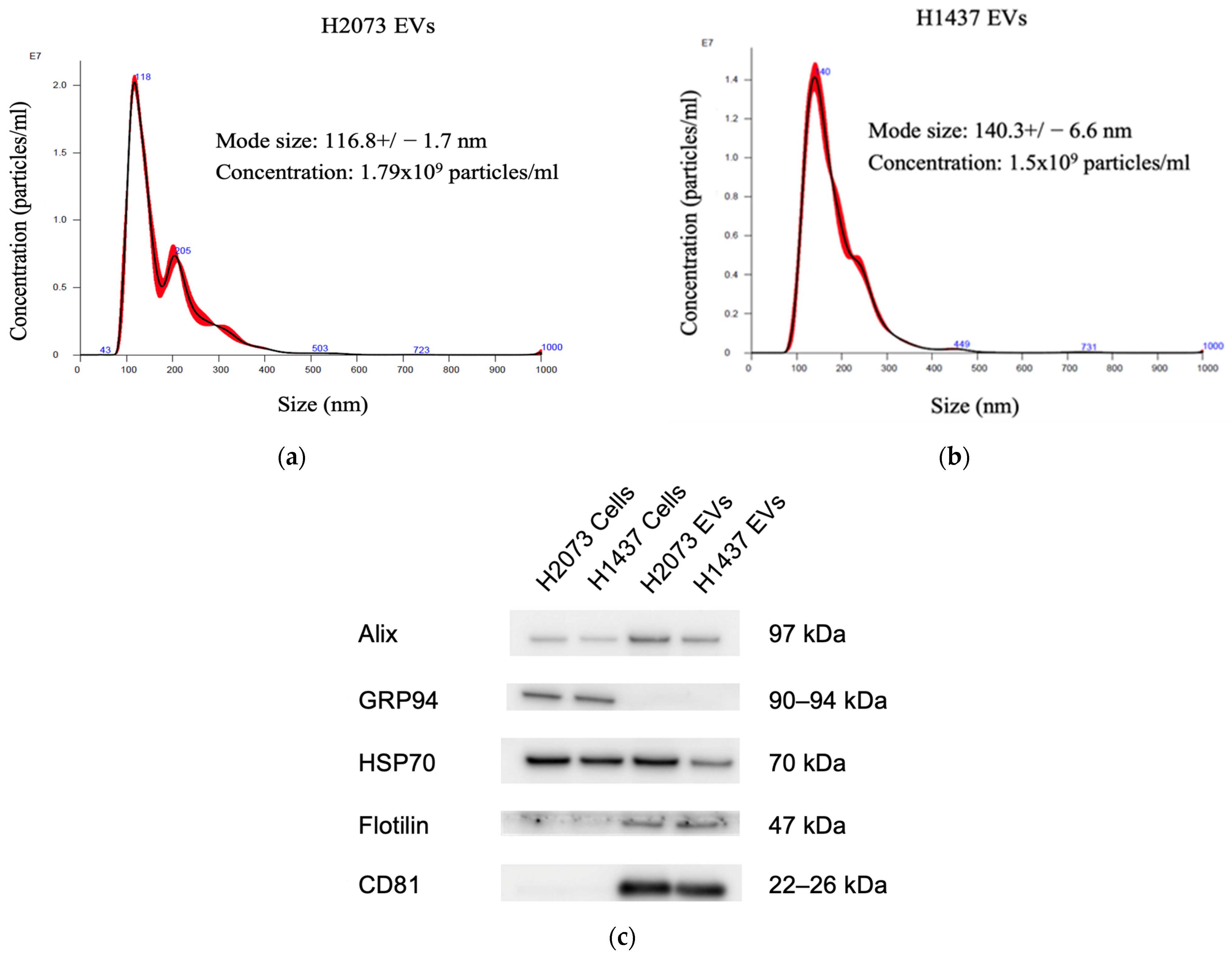
3.1. EVs Isolated from LUAD Cells Are Mainly Small Extracellular Vesicles
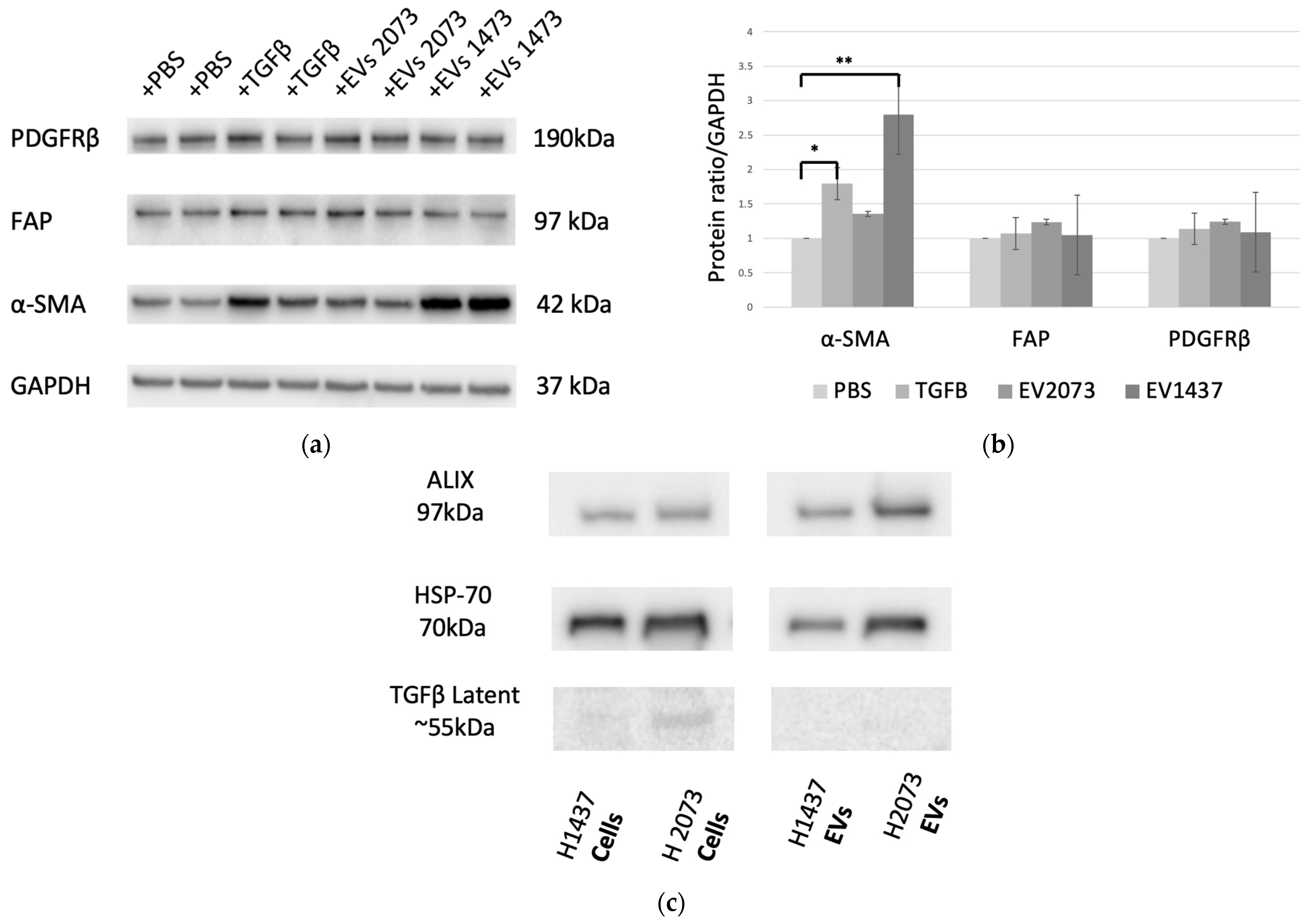
3.2. EVs from LUAD Cells Activate Fibroblasts into Cancer-Associated Fibroblasts (CAFs)
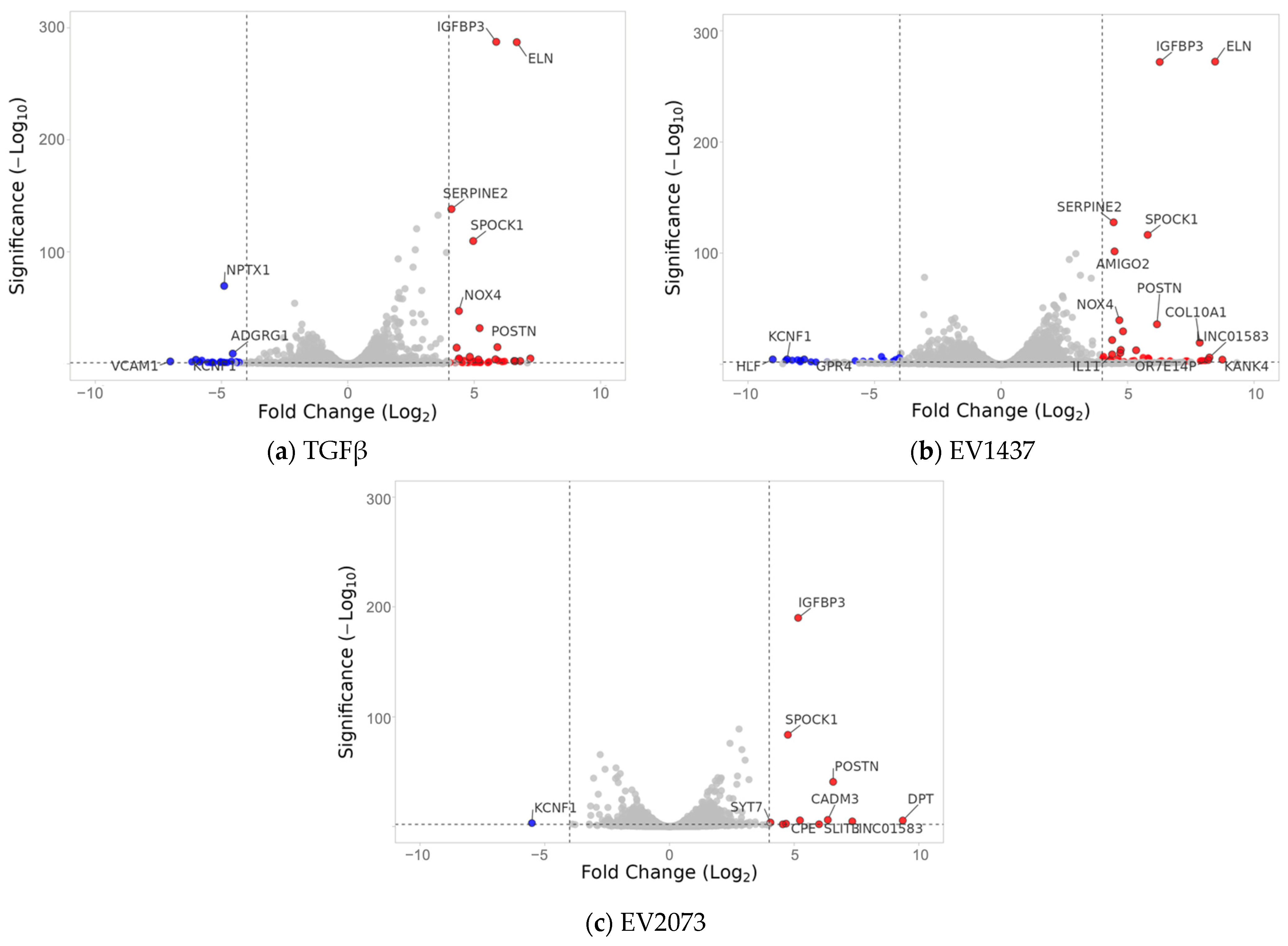
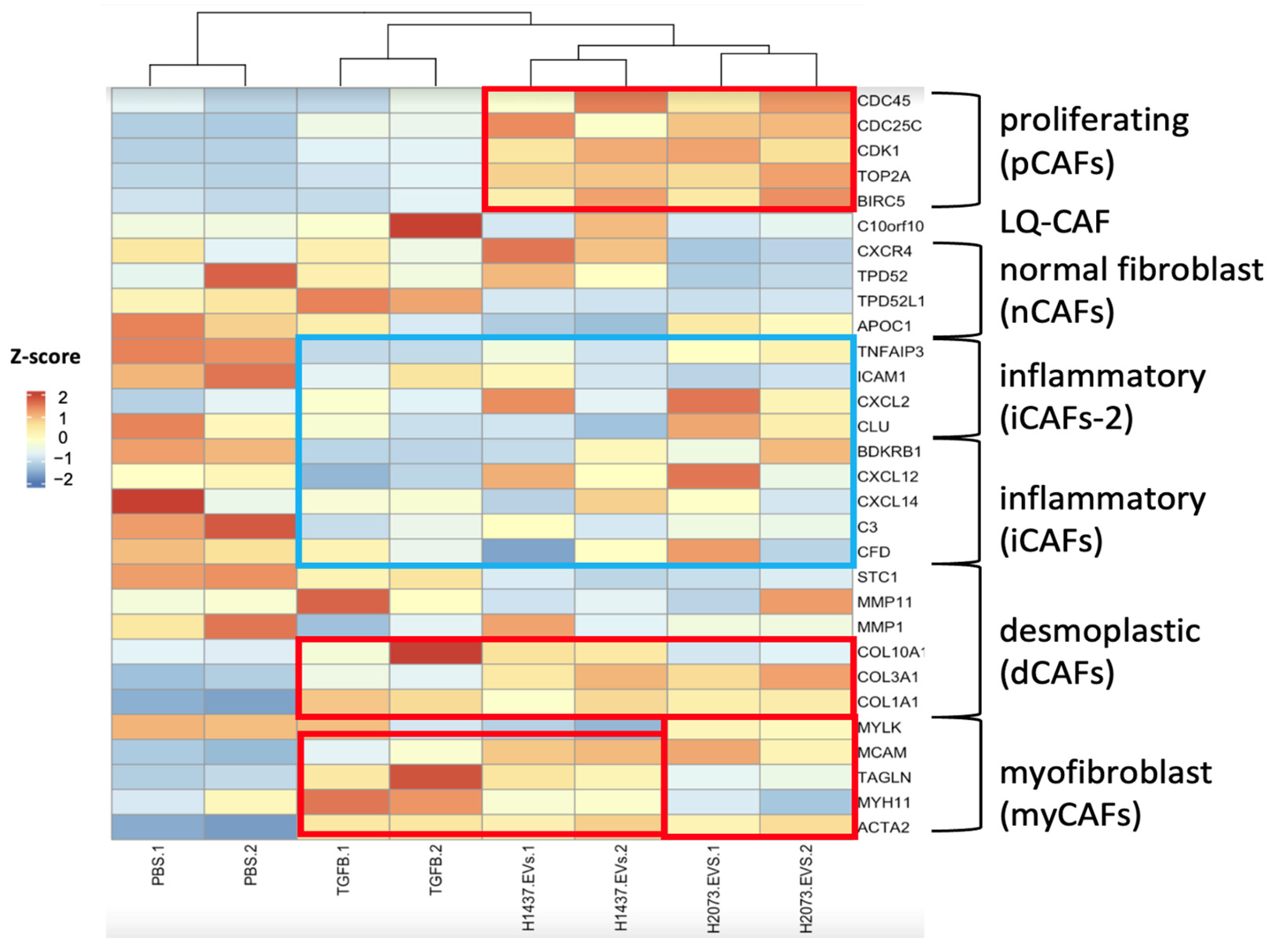
3.3. EVs from LUAD Cells Induce CAFs with Shared and Distinct Phenotypes Compared to TGFβ
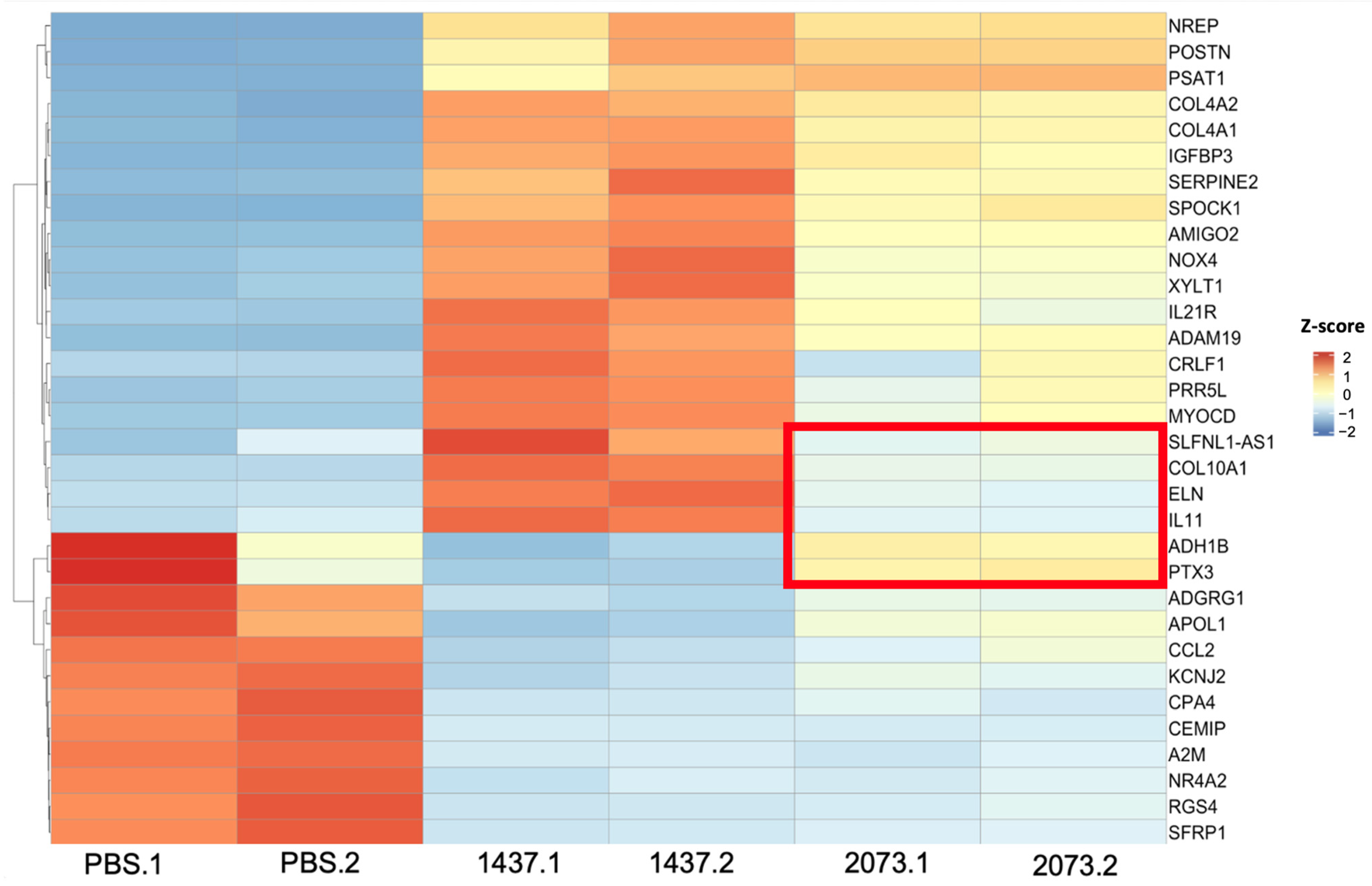
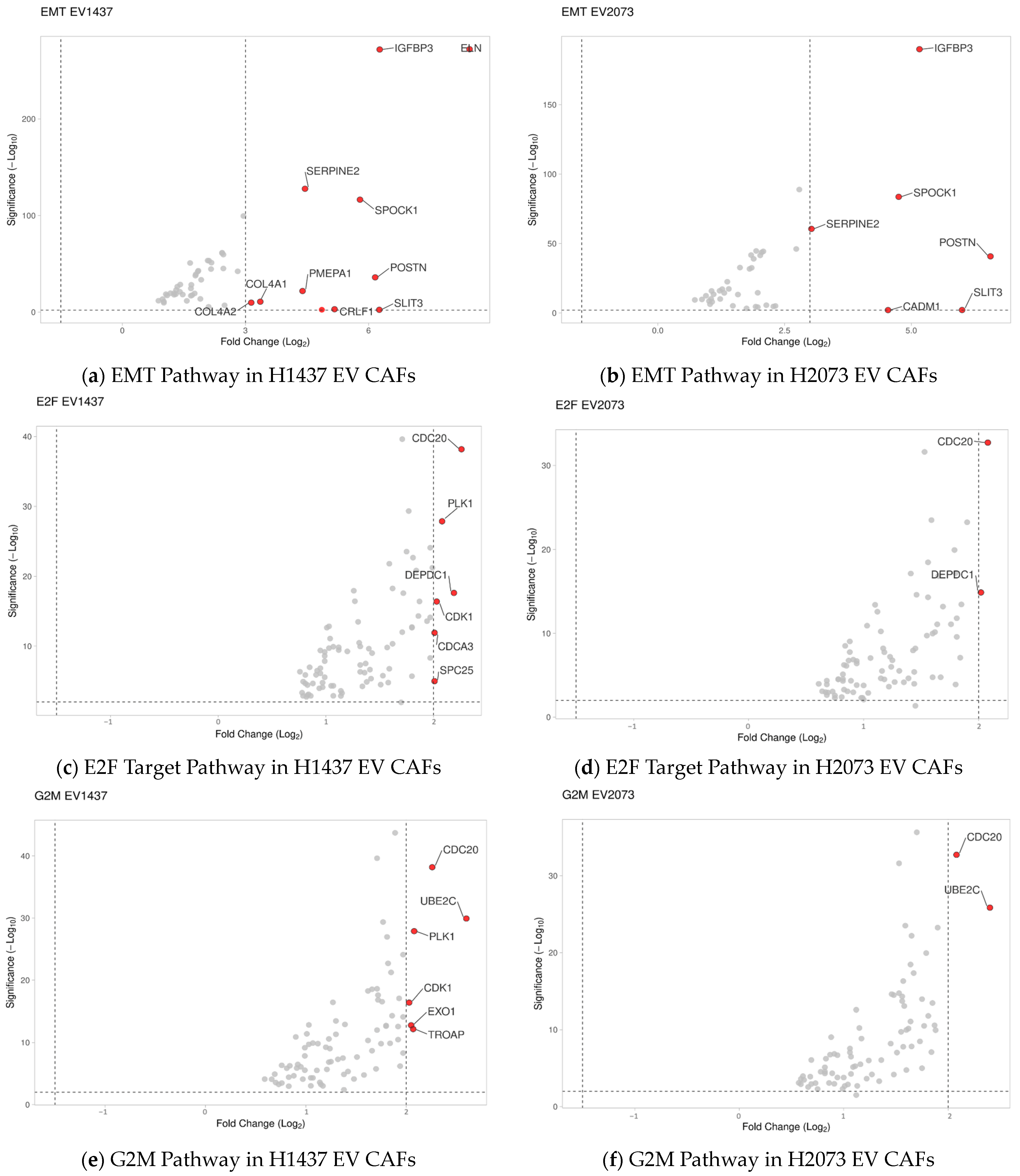
3.4. LUAD EVs Induce Different CAF Subtypes and Upregulation of Proliferation, ECM Remodeling, and Mesenchymal Pathways
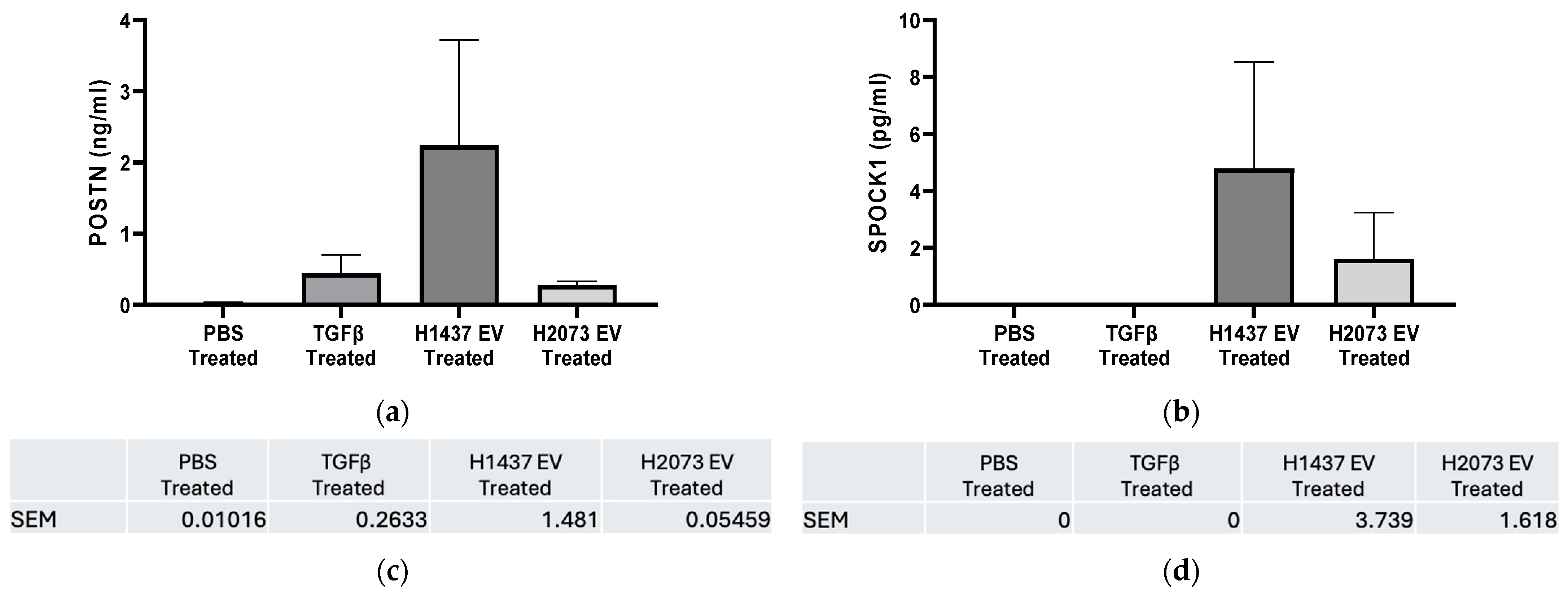
3.5. Highly Upregulated Mesenchymal-Related Genes Are Secreted by CAFs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- PDQ Adult Treatment Editorial Board. Non-Small Cell Lung Cancer Treatment (PDQ®): Health Professional Version. In PDQ Cancer Information Summaries; National Cancer Institute: Bethesda, MD, USA, 2002. [Google Scholar]
- Jin, R.; Wang, X.; Zang, R.; Liu, C.; Zheng, S.; Li, H.; Sun, N.; He, J. Desmoglein-2 modulates tumor progression and osimertinib drug resistance through the EGFR/Src/PAK1 pathway in lung adenocarcinoma. Cancer Lett. 2020, 483, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, X.; Zhou, L.; Ni, J.; Yan, W.; Ma, R.; Wu, J.; Feng, J.; Chen, P. LncRNA HOXA11-AS drives cisplatin resistance of human LUAD cells via modulating miR-454-3p/Stat3. Cancer Sci. 2018, 109, 3068–3079. [Google Scholar] [CrossRef]
- Ball, M.; Hossain, M.; Padalia, D. Anatomy, Airway. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Kratzer, T.B.; Bandi, P.; Freedman, N.D.; Smith, R.A.; Travis, W.D.; Jemal, A.; Siegel, R.L. Lung cancer statistics, 2023. Cancer 2024, 130, 1330–1348. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- Patel, H.; Nilendu, P.; Jahagirdar, D.; Pal, J.K.; Sharma, N.K. Modulating secreted components of tumor microenvironment: A masterstroke in tumor therapeutics. Cancer Biol. Ther. 2018, 19, 3–12. [Google Scholar] [CrossRef]
- Tang, L.; Chen, Z.; Yang, J.; Li, Q.; Wang, S.; Mo, T.; Zeng, W.; Ding, H.; Pan, S. Single-cell and Bulk RNA-Seq reveal angiogenic heterogeneity and microenvironmental features to evaluate prognosis and therapeutic response in lung adenocarcinoma. Front. Immunol. 2024, 15, 1352893. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Ma, Z.; Yang, F.; Zhao, X.; Jiang, W.; Pan, C.; Li, Z.; Pan, X.; He, Z.; Xu, J.; et al. M2 macrophage-derived exosomes promote lung adenocarcinoma progression by delivering miR-942. Cancer Lett. 2022, 526, 205–216. [Google Scholar] [CrossRef]
- Gunaydin, G. CAFs Interacting with TAMs in Tumor Microenvironment to Enhance Tumorigenesis and Immune Evasion. Front. Oncol. 2021, 11, 668349. [Google Scholar] [CrossRef]
- Shi, X.; Young, C.D.; Zhou, H.; Wang, X. Transforming Growth Factor-β Signaling in Fibrotic Diseases and Cancer-Associated Fibroblasts. Biomolecules 2020, 10, 1666. [Google Scholar] [CrossRef]
- Galbo, P.M.; Zang, X.; Zheng, D. Molecular Features of Cancer-Associated Fibroblast Subtypes and their Implication on Cancer Pathogenesis, Prognosis, and Immunotherapy Resistance. Clin. Cancer Res. 2021, 27, 2636–2647. [Google Scholar] [CrossRef]
- Yang, S.-S.; Ma, S.; Dou, H.; Liu, F.; Zhang, S.-Y.; Jiang, C.; Xiao, M.; Huang, Y.-X. Breast cancer-derived exosomes regulate cell invasion and metastasis in breast cancer via miR-146a to activate cancer associated fibroblasts in tumor microenvironment. Exp. Cell Res. 2020, 391, 111983. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Liu, X.; Xiang, J.; Xu, J.; Yin, N.; Wang, L.; Liu, C.; Liu, Y.; Zhao, T.; Zhao, Z.; et al. Exosomal ITGB6 from dormant lung adenocarcinoma cells activates cancer-associated fibroblasts by KLF10 positive feedback loop and the TGF-β pathway. Transl. Lung Cancer Res. 2023, 12, 2520–2537. [Google Scholar] [CrossRef] [PubMed]
- Clancy, J.; D’Souza-Schorey, C. Extracellular Vesicles in Cancer: Purpose and Promise. Cancer J. 2018, 24, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Li, L.; Luo, Y.; Yang, W.; Hu, J.; Zhao, Z.; Cheng, C.; Zhang, T.; Zhang, Y.; Liu, L.; et al. Tumor Cell Derived Lnc-FSD2-31:1 Contributes to Cancer-Associated Fibroblasts Activation in Pancreatic Ductal Adenocarcinoma Progression Through Extracellular Vesicles Cargo MiR-4736. Adv. Sci. 2023, 10, e2203324. [Google Scholar] [CrossRef]
- Arebro, J.; Towle, R.; Lee, C.-M.; Bennewith, K.L.; Garnis, C. Extracellular vesicles promote activation of pro-inflammatory cancer-associated fibroblasts in oral cancer. Front. Cell Dev. Biol. 2023, 11, 1240159. [Google Scholar] [CrossRef]
- SBME Seq Bulk mRNA Seq. Available online: https://bme.ubc.ca/wp-content/uploads/2023/08/SBME-Seq-Bulk-mRNA-Seq-methods-for-publication.pdf (accessed on 6 February 2024).
- Venny 2.1.0. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 2 March 2024).
- Goedhart, J.; Luijsterburg, M.S. VolcaNoseR is a web app for creating, exploring, labeling and sharing volcano plots. Sci. Rep. 2020, 10, 20560. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell. Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef]
- Koncz, A.; Turiák, L.; Németh, K.; Lenzinger, D.; Bárkai, T.; Lőrincz, P.; Zelenyánszki, H.; Vukman, K.V.; Buzás, E.I.; Visnovitz, T. Endoplasmin Is a Hypoxia-Inducible Endoplasmic Reticulum-Derived Cargo of Extracellular Vesicles Released by Cardiac Cell Lines. Membranes 2023, 13, 431. [Google Scholar] [CrossRef]
- Marzec, M.; Eletto, D.; Argon, Y. GRP94: An HSP90-like protein specialized for protein folding and quality control in the endoplasmic reticulum. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2012, 1823, 774–787. [Google Scholar] [CrossRef]
- Ekström, K.; Crescitelli, R.; Pétursson, H.I.; Johansson, J.; Lässer, C.; Olofsson Bagge, R. Characterization of surface markers on extracellular vesicles isolated from lymphatic exudate from patients with breast cancer. BMC Cancer 2022, 22, 50. [Google Scholar] [CrossRef]
- Trejo Vazquez, J.A. Extracellular Vesicles from Lung Adenocarcinoma Cells Induce Activation of Different CAF Subtypes. Master’s Thesis, University of British Columbia, Vancouver, BC, Canada, 2024. [Google Scholar] [CrossRef]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, J.; Qian, H.; Zhuang, Q. Cancer-associated fibroblasts: From basic science to anticancer therapy. Exp. Mol. Med. 2023, 55, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Shoucair, I.; Weber Mello, F.; Jabalee, J.; Maleki, S.; Garnis, C. The Role of Cancer-Associated Fibroblasts and Extracellular Vesicles in Tumorigenesis. Int. J. Mol. Sci. 2020, 21, 6837. [Google Scholar] [CrossRef] [PubMed]
- Marsh, T.; Pietras, K.; McAllister, S.S. Fibroblasts as architects of cancer pathogenesis. Biochim. Biophys. Acta 2013, 1832, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef]
- Nurmik, M.; Ullmann, P.; Rodriguez, F.; Haan, S.; Letellier, E. In search of definitions: Cancer-associated fibroblasts and their markers. Int. J. Cancer 2020, 146, 895–905. [Google Scholar] [CrossRef]
- Dwivedi, N.; Shukla, N.; Prathima, K.M.; Das, M.; Dhar, S.K. Novel CAF-identifiers via transcriptomic and protein level analysis in HNSC patients. Sci. Rep. 2023, 13, 13899. [Google Scholar] [CrossRef]
- Burbidge, K.; Zwikelmaier, V.; Cook, B.; Long, M.M.; Balva, B.; Lonigro, M.; Ispas, G.; Rademacher, D.J.; Campbell, E.M. Cargo and cell-specific differences in extracellular vesicle populations identified by multiplexed immunofluorescent analysis. J. Extracell. Vesicles 2020, 9, 1789326. [Google Scholar] [CrossRef]
- Erdogan, B.; Webb, D.J. Cancer-associated fibroblasts modulate growth factor signaling and extracellular matrix remodeling to regulate tumor metastasis. Biochem. Soc. Trans. 2017, 45, 229–236. [Google Scholar] [CrossRef]
- Szabo, P.M.; Vajdi, A.; Kumar, N.; Tolstorukov, M.Y.; Chen, B.J.; Edwards, R.; Ligon, K.L.; Chasalow, S.D.; Chow, K.-H.; Shetty, A.; et al. Cancer-associated fibroblasts are the main contributors to epithelial-to-mesenchymal signatures in the tumor microenvironment. Sci. Rep. 2023, 13, 3051. [Google Scholar] [CrossRef]
- Baulida, J. Epithelial-to-mesenchymal transition transcription factors in cancer-associated fibroblasts. Mol. Oncol. 2017, 11, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Váncza, L.; Horváth, A.; Seungyeon, L.; Rókusz, A.; Dezső, K.; Reszegi, A.; Petővári, G.; Götte, M.; Kovalszky, I.; Baghy, K. SPOCK1 Overexpression Suggests Poor Prognosis of Ovarian Cancer. Cancers 2023, 15, 2037. [Google Scholar] [CrossRef] [PubMed]
- Dorafshan, S.; Razmi, M.; Safaei, S.; Gentilin, E.; Madjd, Z.; Ghods, R. Periostin: Biology and function in cancer. Cancer Cell Int. 2022, 22, 315. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, D.W. Functional role of periostin in development and wound repair: Implications for connective tissue disease. J. Cell Commun. Signal. 2008, 2, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Katada, K.; Okato, A.; Arai, T.; Idichi, T.; Osako, Y.; Okamoto, Y.; Seki, N. Antitumor miR-150-5p and miR-150-3p inhibit cancer cell aggressiveness by targeting SPOCK1 in head and neck squamous cell carcinoma. Auris Nasus Larynx 2018, 45, 854–865. [Google Scholar] [CrossRef]
- Ye, Z.; Chen, J.; Hu, X.; Yang, S.; Xuan, Z.; Lu, X.; Zhao, Q. SPOCK1: A multi-domain proteoglycan at the crossroads of extracellular matrix remodeling and cancer development. Am. J. Cancer Res. 2020, 10, 3127–3137. [Google Scholar]
- Coenye, T. Do results obtained with RNA-sequencing require independent verification? Biofilm 2021, 3, 100043. [Google Scholar] [CrossRef]
- Everaert, C.; Luypaert, M.; Maag, J.L.V.; Cheng, Q.X.; Dinger, M.E.; Hellemans, J.; Mestdagh, P. Benchmarking of RNA-sequencing analysis workflows using whole-transcriptome RT-qPCR expression data. Sci. Rep. 2017, 7, 1559. [Google Scholar] [CrossRef]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef]
- Glabman, R.A.; Choyke, P.L.; Sato, N. Cancer-Associated Fibroblasts: Tumorigenicity and Targeting for Cancer Therapy. Cancers 2022, 14, 3906. [Google Scholar] [CrossRef]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed]
- Tatischeff, I. Extracellular Vesicle-DNA: The Next Liquid Biopsy Biomarker for Early Cancer Diagnosis? Cancers 2023, 15, 1456. [Google Scholar] [CrossRef] [PubMed]
- Valcz, G.; Újvári, B.; Buzás, E.I.; Krenács, T.; Spisák, S.; Kittel, Á.; Tulassay, Z.; Igaz, P.; Takács, I.; Molnár, B. Small extracellular vesicle DNA-mediated horizontal gene transfer as a driving force for tumor evolution: Facts and riddles. Front. Oncol. 2022, 12, 945376. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.T.; Chen, H.; Nguyen, P.H.D.; Jayasinghe, M.K.; Le, A.H.; Le, M.T. Endosomal escape of nucleic acids from extracellular vesicles mediates functional therapeutic delivery. Pharmacol. Res. 2023, 188, 106665. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jia, Y.; Mao, C.; Liu, S. Small extracellular vesicles: Non-negligible vesicles in tumor progression, diagnosis, and therapy. Cancer Lett. 2024, 580, 216481. [Google Scholar] [CrossRef]
- Zhao, Z.; Wijerathne, H.; Godwin, A.K.; Soper, S.A. Isolation and analysis methods of extracellular vesicles (EVs). Extracell. Vesicles Circ. Nucleic Acids 2021, 2, 80–103. [Google Scholar] [CrossRef]
- Aheget, H.; Mazini, L.; Martin, F.; Belqat, B.; Marchal, J.A.; Benabdellah, K. Exosomes: Their Role in Pathogenesis, Diagnosis and Treatment of Diseases. Cancers 2020, 13, 84. [Google Scholar] [CrossRef]
- Fonseka, P.; Mathivanan, S. Extracellular Vesicles Biogenesis, Cargo Sorting and Implications in Disease Conditions. Cells 2023, 12, 280. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, T.; Sun, L.; Yuan, Y.; Zhu, Y. Potential mechanisms of cancer-associated fibroblasts in therapeutic resistance. Biomed. Pharmacother. 2023, 166, 115425. [Google Scholar] [CrossRef]
- Welcome to ToppGene. Available online: https://toppgene.cchmc.org/ (accessed on 7 February 2024).
- Lawson, J.; Dickman, C.; MacLellan, S.; Towle, R.; Jabalee, J.; Lam, S.; Garnis, C. Selective secretion of microRNAs from lung cancer cells via extracellular vesicles promotes CAMK1D-mediated tube formation in endothelial cells. Oncotarget 2017, 8, 83913–83924. [Google Scholar] [CrossRef]
- Friedman, G.; Levi-Galibov, O.; David, E.; Bornstein, C.; Giladi, A.; Dadiani, M.; Mayo, A.; Halperin, C.; Pevsner-Fischer, M.; Lavon, H.; et al. Cancer-associated fibroblast compositions change with breast cancer progression linking the ratio of S100A4+ and PDPN+ CAFs to clinical outcome. Nat. Cancer 2020, 1, 692–708. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, D.; Wauters, E.; Boeckx, B.; Aibar, S.; Nittner, D.; Burton, O.; Bassez, A.; Decaluwé, H.; Pircher, A.; Van Den Eynde, K.; et al. Phenotype molding of stromal cells in the lung tumor microenvironment. Nat. Med. 2018, 24, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Menezes, S.; Okail, M.H.; Jalil, S.M.A.; Kocher, H.M.; Cameron, A.J.M. Cancer-associated fibroblasts in pancreatic cancer: New subtypes, new markers, new targets. J. Pathol. 2022, 257, 526–544. [Google Scholar] [CrossRef]
- Geng, X.; Chen, H.; Zhao, L.; Hu, J.; Yang, W.; Li, G.; Cheng, C.; Zhao, Z.; Zhang, T.; Li, L.; et al. Cancer-Associated Fibroblast (CAF) Heterogeneity and Targeting Therapy of CAFs in Pancreatic Cancer. Front. Cell Dev. Biol. 2021, 9, 655152. [Google Scholar] [CrossRef]
- Knipper, K.; Lyu, S.I.; Quaas, A.; Bruns, C.J.; Schmidt, T. Cancer-Associated Fibroblast Heterogeneity and Its Influence on the Extracellular Matrix and the Tumor Microenvironment. Int. J. Mol. Sci. 2023, 24, 13482. [Google Scholar] [CrossRef]
- Wan, S.; Liang, Z.; Yao, S.; Wang, A.; Liu, X.; He, H.; Cai, H.; Gao, Y.; Wang, H. Integrated transcriptomics identifies prognostic significance and therapeutic response of cancer-associated fibroblast subpopulations in ovarian cancer. Aging 2024. [Google Scholar] [CrossRef]
- Cords, L.; Engler, S.; Haberecker, M.; Rüschoff, J.H.; Moch, H.; de Souza, N.; Bodenmiller, B. Cancer-associated fibroblast phenotypes are associated with patient outcome in non-small cell lung cancer. Cancer Cell 2024, 42, 396–412.e5. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Xu, J. Cancer-associated fibroblasts: A versatile mediator in tumor progression, metastasis, and targeted therapy. Cancer Metastasis Rev. 2024, 43, 1095–1116. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Yan, M.; Zhang, J.; Wang, X.; Shen, Z.; Lv, Z.; Li, Z.; Wei, W.; Chen, W. TGFβ3-mediated induction of Periostin facilitates head and neck cancer growth and is associated with metastasis. Sci. Rep. 2016, 6, 20587. [Google Scholar] [CrossRef]
- Ouanouki, A.; Lamy, S.; Annabi, B. Periostin, a signal transduction intermediate in TGF-β-induced EMT in U-87MG human glioblastoma cells, and its inhibition by anthocyanidins. Oncotarget 2018, 9, 22023–22037. [Google Scholar] [CrossRef]
- Váncza, L.; Tátrai, P.; Reszegi, A.; Baghy, K.; Kovalszky, I. SPOCK1 with unexpected function. The start of a new career. Am. J. Physiol. Cell Physiol. 2022, 322, C688–C693. [Google Scholar] [CrossRef] [PubMed]
- Yamato, H.; Kimura, K.; Fukui, E.; Kanou, T.; Ose, N.; Funaki, S.; Minami, M.; Shintani, Y. Periostin secreted by activated fibroblasts in idiopathic pulmonary fibrosis promotes tumorigenesis of non-small cell lung cancer. Sci. Rep. 2021, 11, 21114. [Google Scholar] [CrossRef] [PubMed]
- Takatsu, F.; Suzawa, K.; Tomida, S.; Thu, Y.M.; Sakaguchi, M.; Toji, T.; Ohki, M.; Tsudaka, S.; Date, K.; Matsuda, N.; et al. Periostin secreted by cancer-associated fibroblasts promotes cancer progression and drug resistance in non-small cell lung cancer. J. Mol. Med. 2023, 101, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Veenstra, V.L.; Damhofer, H.; Waasdorp, C.; Steins, A.; Kocher, H.M.; Medema, J.P.; van Laarhoven, H.W.; Bijlsma, M.F. Stromal SPOCK1 supports invasive pancreatic cancer growth. Mol. Oncol. 2017, 11, 1050–1064. [Google Scholar] [CrossRef]
- Fan, L.-C.; Jeng, Y.-M.; Lu, Y.-T.; Lien, H.-C. SPOCK1 Is a Novel Transforming Growth Factor-β–Induced Myoepithelial Marker That Enhances Invasion and Correlates with Poor Prognosis in Breast Cancer. PLoS ONE 2016, 11, e0162933. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | +TGFβ | +H1437 | +H2073 |
|---|---|---|---|
| COL1A1 | 1.98 | 2.27 | 2.41 |
| COL1A2 | 1.06 | 2.61 | 2.58 |
| COL3A1 | 0.44 | 1.36 | 1.26 |
| COL4A1 | 2.54 | 3.52 | 2.30 |
| COL4A2 | 2.24 | 2.20 | 1.63 |
| COL5A1 | 1.76 | 2.61 | 3.04 |
| COL6A1 | 0.06 | 1.30 | 1.15 |
| COL7A1 | 1.48 | 1.92 | 1.40 |
| COL8A1 | 0.97 | 1.04 | 0.26 |
| COL10A1 | 5.92 | 7.85 | 3.44 |
| COL11A1 | 1.72 | 1.51 | 1.75 |
| COL13A1 | −0.45 | −0.88 | −1.06 |
| COL16A1 | 0.81 | 1.30 | 1.22 |
| TAGLN | 1.08 | 1.26 | 0.39 |
| ELN | 6.69 | 8.46 | 2.73 |
| BGN | 2.67 | 2.95 | 2.79 |
| DCN | −0.20 | 0.31 | 0.36 |
| LUM | 1.02 | 1.90 | 1.97 |
| MMP1 | −0.77 | −0.02 | −0.48 |
| MMP2 | 1.42 | 1.79 | 1.37 |
| MMP3 | −1.37 | −0.32 | −0.83 |
| MMP10 | 2.17 | −0.57 | 2.08 |
| MMP11 | 0.64 | −0.47 | 0.58 |
| MMP14 | 0.42 | 0.56 | 0.13 |
| MMP19 | 0.80 | 0.50 | 0.80 |
| SERPINE1 | 1.06 | 1.38 | 0.40 |
| CTHRC1 | 2.06 | 2.17 | 1.89 |
| THBS2 | 0.72 | 1.01 | 0.03 |
| SULF1 | 0.50 | 0.56 | −0.11 |
| COMP | 3.61 | 2.20 | 3.14 |
| INHBA | 1.54 | 1.78 | 0.11 |
| TGFBI | 1.56 | 2.43 | 2.08 |
| PDGFA | 1.33 | 1.31 | 0.06 |
| PDGFC | 1.14 | 1.01 | 0.73 |
| VEGFA | 0.41 | 0.27 | 0.50 |
| ANGPT2 | 0.07 | −1.28 | −1.47 |
| ACTA2 | 1.52 | 1.85 | 1.62 |
| FAP | 2.03 | 2.08 | 1.84 |
| POSTN | 5.21 | 6.16 | 6.56 |
| PDGFRβ | 0.55 | 0.17 | 0.19 |
| MYL6 | 0.11 | 0.35 | −0.27 |
| MYH9 | 0.43 | 0.19 | 0.21 |
| MYH11 | 0.94 | 0.72 | −0.42 |
| TPM1 | 2.01 | 2.46 | 1.07 |
| SORBS2 | 5.37 | 6.14 | 3.38 |
| RRAS | −0.35 | −0.20 | −0.19 |
| CXCL12 | −1.98 | 0.51 | 0.20 |
| CXCL14 | −2.17 | −1.14 | 0.55 |
| CXCL2 | 0.29 | 1.37 | 1.32 |
| CXCL3 | −0.98 | 0.81 | 0.31 |
| CCL2 | −2.52 | −3.97 | −1.27 |
| IL6 | −0.44 | 0.78 | −1.28 |
| PDPN | 1.22 | 2.18 | 2.19 |
| CD29 | 0.53 | 0.62 | −0.11 |
| CD90 | 0.47 | −0.31 | 0.62 |
| Vimentin | 0.19 | 0.20 | 0.08 |
| GPR77 | 0.01 | 1.05 | −1.10 |
| CD10 | −0.12 | 0.69 | 0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trejo Vazquez, J.A.; Towle, R.; Farnsworth, D.A.; Sarafan, M.; Lockwood, W.W.; Garnis, C. Extracellular Vesicles from Lung Adenocarcinoma Cells Induce Activation of Different Cancer-Associated Fibroblast Subtypes. Biomedicines 2024, 12, 2523. https://doi.org/10.3390/biomedicines12112523
Trejo Vazquez JA, Towle R, Farnsworth DA, Sarafan M, Lockwood WW, Garnis C. Extracellular Vesicles from Lung Adenocarcinoma Cells Induce Activation of Different Cancer-Associated Fibroblast Subtypes. Biomedicines. 2024; 12(11):2523. https://doi.org/10.3390/biomedicines12112523
Chicago/Turabian StyleTrejo Vazquez, Jessica Angelina, Rebecca Towle, Dylan Andrew Farnsworth, Masih Sarafan, William Wallace Lockwood, and Cathie Garnis. 2024. "Extracellular Vesicles from Lung Adenocarcinoma Cells Induce Activation of Different Cancer-Associated Fibroblast Subtypes" Biomedicines 12, no. 11: 2523. https://doi.org/10.3390/biomedicines12112523
APA StyleTrejo Vazquez, J. A., Towle, R., Farnsworth, D. A., Sarafan, M., Lockwood, W. W., & Garnis, C. (2024). Extracellular Vesicles from Lung Adenocarcinoma Cells Induce Activation of Different Cancer-Associated Fibroblast Subtypes. Biomedicines, 12(11), 2523. https://doi.org/10.3390/biomedicines12112523