
miRNAs’ Cross-Involvement in Skin Allergies: A New Horizon for the Pathogenesis, Diagnosis and Therapy of Atopic Dermatitis, Allergic Contact Dermatitis and Chronic Spontaneous Urticaria

 ,
,  , ,
, ,
Abstract
1. Introduction
1.1. Pathogenesis of Atopic Dermatitis
1.2. Pathogenesis of Allergic Contact Dermatitis
1.3. Pathogenesis of Chronic Spontaneous Urticaria
1.4. MicroRNAs
2. miRNA in AD, ACD and CSU: Pathogenetic Role and Therapeutic Strategies
2.1. Pro-Inflammatory and Anti-Inflammatory miRNA in AD
2.2. Pro-Inflammatory and Anti-Inflammatory miRNA in ACD
2.3. Pro-Inflammatory and Anti-Inflammatory miRNA in CSU
3. Therapeutic Perspectives
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic Dermatitis. Nat. Rev. Dis. Primers 2018, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Nettis, E.; Patella, V.; Brancaccio, R.; Detoraki, C.; Di Leo, E.; Incorvaia, C.; Macchia, L.; Pellacani, G.; Bonzano, L. Efficacy of Dupilumab in Concomitant Atopic Dermatitis and Chronic Rhinosinusitis With Nasal Polyps: A Preliminary Study. Allergy Asthma Immunol. Res. 2021, 13, 347–349. [Google Scholar] [CrossRef]
- Megna, M.; Patruno, C.; Balato, A.; Rongioletti, F.; Stingeni, L.; Balato, N.; Ayala, F.; Brambilla, L.; Congedo, M.; Corazza, M.; et al. An Italian Multicentre Study on Adult Atopic Dermatitis: Persistent versus Adult-Onset Disease. Arch. Derm. Res. 2017, 309, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Kraft, M.; Worm, M. Dupilumab in the Treatment of Moderate-to-Severe Atopic Dermatitis. Expert Rev. Clin. Immunol. 2017, 13, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Feldman, S.R. Management of Atopic Dermatitis. Adherence in Atopic Dermatitis. Introduction. Adv. Exp. Med. Biol. 2017, 1027, 139–159. [Google Scholar] [CrossRef]
- Elias, P.M.; Steinhoff, M. “Outside-to-inside” (and Now Back to “Outside”) Pathogenic Mechanisms in Atopic Dermatitis. J. Investig. Dermatol. 2008, 128, 1067–1070. [Google Scholar] [CrossRef]
- Bertino, L.; Guarneri, F.; Cannavò, S.P.; Casciaro, M.; Pioggia, G.; Gangemi, S. Oxidative Stress and Atopic Dermatitis. Antioxidants 2020, 9, 196. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Reszka, E.; Gutowska-Owsiak, D.; Trzeciak, M.; Lange, M.; Jarczak, J.; Niedoszytko, M.; Jablonska, E.; Romantowski, J.; Strapagiel, D.; et al. Genetic and Epigenetic Aspects of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 6484. [Google Scholar] [CrossRef]
- Zaniboni, M.C.; Samorano, L.P.; Orfali, R.L.; Aoki, V. Skin Barrier in Atopic Dermatitis: Beyond Filaggrin. Bras Derm. 2016, 91, 472–478. [Google Scholar] [CrossRef]
- Katsunuma, T.; Kawahara, H.; Yuki, K.; Akasawa, A.; Saito, H. Impaired Interferon-Gamma Production in a Subset Population of Severe Atopic Dermatitis. Int. Arch. Allergy Immunol. 2004, 134, 240–247. [Google Scholar] [CrossRef]
- Malik, K.; Heitmiller, K.D.; Czarnowicki, T. An Update on the Pathophysiology of Atopic Dermatitis. Derm. Clin. 2017, 35, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D.; et al. Common Loss-of-Function Variants of the Epidermal Barrier Protein Filaggrin Are a Major Predisposing Factor for Atopic Dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Salimi, M.; Barlow, J.L.; Saunders, S.P.; Xue, L.; Gutowska-Owsiak, D.; Wang, X.; Huang, L.C.; Johnson, D.; Scanlon, S.T.; McKenzie, A.N.J.; et al. A Role for IL-25 and IL-33-Driven Type-2 Innate Lymphoid Cells in Atopic Dermatitis. J. Exp. Med. 2013, 210, 2939–2950. [Google Scholar] [CrossRef]
- Ito, T.; Wang, Y.H.; Duramad, O.; Hori, T.; Delespesse, G.J.; Watanabe, N.; Qin, F.X.F.; Yao, Z.; Cao, W.; Liu, Y.J. TSLP-Activated Dendritic Cells Induce an Inflammatory T Helper Type 2 Cell Response through OX40 Ligand. J. Exp. Med. 2005, 202, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.A.; Bennett, B.L.; Graham, N.M.H.; Pirozzi, G.; Stahl, N.; Yancopoulos, G.D. Targeting Key Proximal Drivers of Type 2 Inflammation in Disease. Nat. Rev. Drug Discov. 2016, 15, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.A.; Luster, A.D. T Cell Homing to Epithelial Barriers in Allergic Disease. Nat. Med. 2012, 18, 705–715. [Google Scholar] [CrossRef]
- Liu, B.; Tai, Y.; Achanta, S.; Kaelberer, M.M.; Caceres, A.I.; Shao, X.; Fang, J.; Jordt, S.E. IL-33/ST2 Signaling Excites Sensory Neurons and Mediates Itch Response in a Mouse Model of Poison Ivy Contact Allergy. Proc. Natl. Acad. Sci. USA 2016, 113, E7572–E7579. [Google Scholar] [CrossRef]
- Nassau, S.; Fonacier, L. Allergic Contact Dermatitis. Med. Clin. N. Am. 2020, 104, 61–76. [Google Scholar] [CrossRef]
- Zack, B.; Arrandale, V.H.; Holness, D.L. Preventing Occupational Skin Disease: A Review of Training Programs. Dermatitis 2017, 28, 169–182. [Google Scholar] [CrossRef]
- Rustemeyer, T.; Van Hoogstraten, I.M.W.; Von Blomberg, B.M.E.; Gibbs, S.; Scheper, R.J. Mechanisms of Irritant and Allergic Contact Dermatitis. Contact Dermat. 2011, 43–90. [Google Scholar] [CrossRef]
- Adelman, D.C.; Casale, T.B.; Corren, J.; Ovid Technologies, Inc. Manual of Allergy and Immunology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012; p. 494. [Google Scholar]
- Zuberbier, T.; Abdul Latiff, A.H.; Abuzakouk, M.; Aquilina, S.; Asero, R.; Baker, D.; Ballmer-Weber, B.; Bangert, C.; Ben-Shoshan, M.; Bernstein, J.A.; et al. The International EAACI/GA2LEN/EuroGuiDerm/APAAACI Guideline for the Definition, Classification, Diagnosis, and Management of Urticaria. Allergy Eur. J. Allergy Clin. Immunol. 2022, 77, 734–766. [Google Scholar] [CrossRef]
- Nettis, E.; Cegolon, L.; Di Leo, E.; Lodi Rizzini, F.; Detoraki, A.; Canonica, G.W. Omalizumab in Chronic Spontaneous Urticaria: Efficacy, Safety, Predictors of Treatment Outcome, and Time to Response. Ann. Allergy Asthma Immunol. 2018, 121, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, C.; Toubi, E.; Maurer, M.; Triggiani, M.; Ballmer-Weber, B.; Marsland, A.; Ferrer, M.; Knulst, A.; Giménez-Arnau, A. Treatment of Chronic Spontaneous Urticaria with an Inadequate Response to H1-Antihistamines: An Expert Opinion. Eur. J. Dermatol. 2017, 27, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Bracken, S.J.; Abraham, S.; MacLeod, A.S. Autoimmune Theories of Chronic Spontaneous Urticaria. Front. Immunol. 2019, 10, 627. [Google Scholar] [CrossRef]
- Grattan, C.E.H.; Dawn, G.; Gibbs, S.; Francis, D.M. Blood Basophil Numbers in Chronic Ordinary Urticaria and Healthy Controls: Diurnal Variation, Influence of Loratadine and Prednisolone and Relationship to Disease Activity. Clin. Exp. Allergy 2003, 33, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Kern, F.; Lichtenstein, L.M. Defective Histamine Release in Chronic Urticaria. J. Clin. Investig. 1976, 57, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.S.; Sui, J.F.; Chen, X.H.; Ran, X.Z.; Yang, Z.F.; Guan, W.D.; Yang, T. Detection of CD4+ CD25+ FOXP3+ Regulatory T Cells in Peripheral Blood of Patients with Chronic Autoimmune Urticaria. Australas J. Dermatol. 2011, 52, e15–e18. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, Y.; Ming, B.; Wu, X.; Chen, Y.; Dong, L. Regulatory T-Cell Levels in Systemic Lupus Erythematosus Patients: A Meta-Analysis. Lupus 2019, 28, 445–454. [Google Scholar] [CrossRef]
- Morita, T.; Shima, Y.; Wing, J.B.; Sakaguchi, S.; Ogata, A.; Kumanogoh, A. The Proportion of Regulatory T Cells in Patients with Rheumatoid Arthritis: A Meta-Analysis. PLoS ONE 2016, 11, e0162306. [Google Scholar] [CrossRef]
- Vasagar, K.; Vonakis, B.M.; Gober, L.M.; Viksman, A.; Gibbons, S.P.; Saini, S.S. Evidence of in Vivo Basophil Activation in Chronic Idiopathic Urticaria. Clin. Exp. Allergy 2006, 36, 770–776. [Google Scholar] [CrossRef]
- Ulambayar, B.; Chen, Y.H.; Ban, G.Y.; Lee, J.H.; Jung, C.G.; Yang, E.M.; Park, H.S.; Ye, Y.M. Detection of Circulating IgG Autoantibody to FcεRIα in Sera from Chronic Spontaneous Urticaria Patients. J. Microbiol. Immunol. Infect. 2020, 53, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Qi, R.; Yang, Y.; Gao, X.; Chen, H.; Xiao, T. Serum MiR-125a-5p and CCL17 Upregulated in Chronic Spontaneous Urticaria and Correlated with Treatment Response. Acta Derm. Venereol. 2019, 99, 571–578. [Google Scholar] [CrossRef]
- Ambros, V.; Lee, R.C.; Lavanway, A.; Williams, P.T.; Jewell, D. MicroRNAs and Other Tiny Endogenous RNAs in C. Elegans. Curr. Biol. 2003, 13, 807–818. [Google Scholar] [CrossRef]
- Setoyama, T.; Ling, H.; Natsugoe, S.; Calin, G.A. Non-Coding RNAs for Medical Practice in Oncology. Keio J. Med. 2011, 60, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA Polymerase III Transcribes Human MicroRNAs. Nat. Struct Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Müller, P.; et al. Conservation of the Sequence and Temporal Expression of Let-7 Heterochronic Regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Akiyama, Y.; Yuasa, Y. Multiple-to-multiple relationships between microRNAs and target genes in gastric cancer. PLoS ONE 2013, 8, e62589. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Annotating High Confidence MicroRNAs Using Deep Sequencing Data. Nucleic. Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef]
- Sayed, D.; Abdellatif, M. MicroRNAs in Development and Disease. Physiol. Rev. 2011, 91, 827–887. [Google Scholar] [CrossRef]
- Hutvágner, G.; Zamore, P.D. A MicroRNA in a Multiple-Turnover RNAi Enzyme Complex. Science 2002, 297, 2056–2060. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef]
- Hu, H.Y.; Yan, Z.; Xu, Y.; Hu, H.; Menzel, C.; Zhou, Y.H.; Chen, W.; Khaitovich, P. Sequence Features Associated with MicroRNA Strand Selection in Humans and Flies. BMC Genom. 2009, 10, 413. [Google Scholar] [CrossRef]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of MRNA Translation and Stability by MicroRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 14, 172. [Google Scholar] [CrossRef]
- Tong, K.L.; Tan, K.E.; Lim, Y.Y.; Tien, X.Y.; Wong, P.F. CircRNA-miRNA interactions in atherogenesis. Mol. Cell. Biochem. 2022, 477, 2703–2733. [Google Scholar] [CrossRef]
- Huntzinger, E.; Izaurralde, E. Gene Silencing by MicroRNAs: Contributions of Translational Repression and MRNA Decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef]
- Ambros, V. The Functions of Animal MicroRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Fiorucci, G.; Chiantore, M.V.; Mangino, G.; Percario, Z.A.; Affabris, E.; Romeo, G. Cancer Regulator MicroRNA: Potential Relevance in Diagnosis, Prognosis and Treatment of Cancer. Curr. Med. Chem. 2012, 19, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Shelburne, C.P.; Ryan, J.J. The Role of Th2 Cytokines in Mast Cell Homeostasis. Immunol. Rev. 2001, 179, 82–93. [Google Scholar] [CrossRef]
- Banerjee, J.; Sen, C.K. MicroRNAs in skin and wound healing. Methods Mol. Biol. 2013, 936, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Frankel, H.C.; Qureshi, A.A. Comparative Effectiveness of Topical Calcineurin Inhibitors in Adult Patients with Atopic Dermatitis. Am. J. Clin. Dermatol. 2012, 13, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Janson, P.; Majuri, M.L.; Savinko, T.; Fyhrquist, N.; Eidsmo, L.; Xu, N.; Meisgen, F.; Wei, T.; Bradley, M.; et al. MiR-155 Is Overexpressed in Patients with Atopic Dermatitis and Modulates T-Cell Proliferative Responses by Targeting Cytotoxic T Lymphocyte-Associated Antigen 4. J. Allergy Clin. Immunol. 2010, 126, 581–589.e20. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Pivarcsi, A. Advances in MicroRNAs: Implications for Immunity and Inflammatory Diseases. J. Cell Mol. Med. 2009, 13, 24–38. [Google Scholar] [CrossRef]
- Rodriguez, A.; Vigorito, E.; Clare, S.; Warren, M.V.; Couttet, P.; Soond, D.R.; Van Dongen, S.; Grocock, R.J.; Das, P.P.; Miska, E.A.; et al. Requirement of Bic/MicroRNA-155 for Normal Immune Function. Science 2007, 316, 608–611. [Google Scholar] [CrossRef]
- Thai, T.H.; Calado, D.P.; Casola, S.; Ansel, K.M.; Xiao, C.; Xue, Y.; Murphy, A.; Frendewey, D.; Valenzuela, D.; Kutok, J.L.; et al. Regulation of the Germinal Center Response by MicroRNA-155. Science 2007, 316, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Rebane, A.; Runnel, T.; Aab, A.; Maslovskaja, J.; Rückert, B.; Zimmermann, M.; Plaas, M.; Kärner, J.; Treis, A.; Pihlap, M.; et al. MicroRNA-146a Alleviates Chronic Skin Inflammation in Atopic Dermatitis through Suppression of Innate Immune Responses in Keratinocytes. J. Allergy Clin. Immunol. 2014, 134, 836–847.e11. [Google Scholar] [CrossRef]
- West, C.; McDermott, M.F. Effects of MicroRNA-146a on the Proliferation and Apoptosis of Human Osteochondrocytes by Targeting TRAF6 through the NF- ΚB Signalling Pathway. Biosci. Rep. 2017, 37, 180. [Google Scholar] [CrossRef]
- Park, H.; Huang, X.; Lu, C.; Cairo, M.S.; Zhou, X. MicroRNA-146a and MicroRNA-146b Regulate Human Dendritic Cell Apoptosis and Cytokine Production by Targeting TRAF6 and IRAK1 Proteins. J. Biol. Chem. 2015, 290, 2831–2841. [Google Scholar] [CrossRef]
- Lindner, J.M.; Kayo, H.; Hedlund, S.; Fukuda, Y.; Fukao, T.; Nielsen, P.J. Cutting Edge: The Transcription Factor Bob1 Counteracts B Cell Activation and Regulates MiR-146a in B Cells. J. Immunol. 2014, 192, 4483–4486. [Google Scholar] [CrossRef]
- Williams, A.E.; Perry, M.M.; Moschos, S.A.; Larner-Svensson, H.M.; Lindsay, M.A. Role of MiRNA-146a in the Regulation of the Innate Immune Response and Cancer. Biochem. Soc. Trans. 2008, 36, 1211–1215. [Google Scholar] [CrossRef]
- Chen, X.F.; Zhang, L.J.; Zhang, J.; Dou, X.; Shao, Y.; Jia, X.J.; Zhang, W.; Yu, B. MiR-151a Is Involved in the Pathogenesis of Atopic Dermatitis by Regulating Interleukin-12 Receptor Β2. Exp. Dermatol. 2018, 27, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Park, C.D.; Lee, A.Y. Administration of Poly(I:C) Improved Dermatophagoides Farinae-Induced Atopic Dermatitis-like Skin Lesions in NC/Nga Mice by the Regulation of Th1/Th2 Balance. Vaccine 2012, 30, 2405–2410. [Google Scholar] [CrossRef]
- Guo, H.W.; Yun, C.X.; Hou, G.H.; Du, J.; Huang, X.; Lu, Y.; Keller, E.T.; Zhang, J.; Deng, J.G. Mangiferin Attenuates TH1/TH2 Cytokine Imbalance in an Ovalbumin-Induced Asthmatic Mouse Model. PLoS ONE 2014, 9, e100394. [Google Scholar] [CrossRef]
- Jia, H.Z.; Liu, S.L.; Zou, Y.F.; Chen, X.F.; Yu, L.; Wan, J.; Zhang, H.Y.; Chen, Q.; Xiong, Y.; Yu, B.; et al. MicroRNA-223 Is Involved in the Pathogenesis of Atopic Dermatitis by Affecting Histamine-N-Methyltransferase. Cell Mol. Biol. 2018, 64, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Allantaz, F.; Cheng, D.T.; Bergauer, T.; Ravindran, P.; Rossier, M.F.; Ebeling, M.; Badi, L.; Reis, B.; Bitter, H.; D’Asaro, M.; et al. Expression Profiling of Human Immune Cell Subsets Identifies MiRNA-MRNA Regulatory Relationships Correlated with Cell Type Specific Expression. PLoS ONE 2012, 7, e29979. [Google Scholar] [CrossRef] [PubMed]
- Herberth, G.; Bauer, M.; Gasch, M.; Hinz, D.; Röder, S.; Olek, S.; Kohajda, T.; Rolle-Kampczyk, U.; Von Bergen, M.; Sack, U.; et al. Maternal and Cord Blood MiR-223 Expression Associates with Prenatal Tobacco Smoke Exposure and Low Regulatory T-Cell Numbers. J. Allergy Clin. Immunol. 2014, 133, 543–550.e4. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Qi, R.; Xu, J.; Di, Z.; Zheng, H.; Huo, W.; Zhang, L.; Chen, H.; Gao, X. Profiling of Serum and Urinary MicroRNAs in Children with Atopic Dermatitis. PLoS ONE 2014, 9, e115448. [Google Scholar] [CrossRef] [PubMed]
- Freire De Carvalho, J.; Machado Ribeiro, F. The Potential Role of MicroRNAs as Biomarkers in Atopic Dermatitis: A Systematic Review. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11801–11803. [Google Scholar] [CrossRef]
- Vaher, H.; Runnel, T.; Urgard, E.; Aab, A.; Carreras Badosa, G.; Maslovskaja, J.; Abram, K.; Raam, L.; Kaldvee, B.; Annilo, T.; et al. MiR-10a-5p Is Increased in Atopic Dermatitis and Has Capacity to Inhibit Keratinocyte Proliferation. Allergy 2019, 74, 2146–2156. [Google Scholar] [CrossRef]
- Chaoying Gu; Yin Li; Jinfeng Wu; Jinhua Xu IFN-γ-Induced MicroRNA-29b up-Regulation Contributes Tokeratinocyte Apoptosis in Atopic Dermatitis through Inhibiting Bcl2L2—PubMed. Int. J. Clin. Exp. Pathol. 2017, 10, 10117–10126.
- Ralfkiaer, U.; Lindahl, L.M.; Litman, T.; Gjerdrum, L.M.; Ahler, C.B.; Gniadecki, R.; Marstrand, T.; Fredholm, S.; Iversen, L.; Wasik, M.A.; et al. MicroRNA Expression in Early Mycosis Fungoides Is Distinctly Different from Atopic Dermatitis and Advanced Cutaneous T-Cell Lymphoma. Anticancer. Res. 2014, 34, 7207–7217. [Google Scholar] [PubMed]
- Sonkoly, E.; Wei, T.; Janson, P.C.J.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A.; et al. MicroRNAs: Novel Regulators Involved in the Pathogenesis of Psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.P.; Nguyen, G.H.; Jin, H.Z. MicroRNA-143 Inhibits IL-13-Induced Dysregulation of the Epidermal Barrier-Related Proteins in Skin Keratinocytes via Targeting to IL-13Rα1. Mol. Cell Biochem. 2016, 416, 63–70. [Google Scholar] [CrossRef]
- Jeong, D.; Kim, J.; Nam, J.; Sun, H.; Lee, Y.H.; Lee, T.J.; Aguiar, R.C.T.; Kim, S.W. MicroRNA-124 Links P53 to the NF-ΚB Pathway in B-Cell Lymphomas. Leukemia 2015, 29, 1868–1874. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, B.; Wang, C.; Wang, H.; Huang, P.; Pan, Y. MicroRNA-124 Alleviates Chronic Skin Inflammation in Atopic Eczema via Suppressing Innate Immune Responses in Keratinocytes. Cell Immunol. 2017, 319, 53–60. [Google Scholar] [CrossRef]
- Vennegaard, M.T.; Bonefeld, C.M.; Hagedorn, P.H.; Bangsgaard, N.; Løvendorf, M.B.; Odum, N.; Woetmann, A.; Geisler, C.; Skov, L. Allergic Contact Dermatitis Induces Upregulation of Identical MicroRNAs in Humans and Mice. Contact Dermat. 2012, 67, 298–305. [Google Scholar] [CrossRef]
- Gulati, N.; Løvendorf, M.B.; Zibert, J.R.; Akat, K.M.; Renwick, N.; Tuschl, T.; Krueger, J.G. Unique MicroRNAs Appear at Different Times during the Course of a Delayed-Type Hypersensitivity Reaction in Human Skin. Exp. Dermatol. 2015, 24, 953–957. [Google Scholar] [CrossRef]
- Anderson, S.E.; Beezhold, K.; Lukomska, E.; Richardson, J.; Long, C.; Anderson, K.; Franko, J.; Meade, B.J.; Beezhold, D.H. Expression Kinetics of MiRNA Involved in Dermal Toluene 2,4-Diisocyanate Sensitization. J. Immunotoxicol. 2014, 11, 250–259. [Google Scholar] [CrossRef]
- Werner, P.; Wisgrill, L.; Riskumäki, M.; Jalonen, E.; Vendelin, J.; Suomela, S.; Lauerma, A.; Alenius, H.; Fyhrquist, N. Identification of Novel MiRNA-MRNA Regulatory Networks in Contact Dermatitis by Integrated Microarray Analysis. Allergy 2021, 76, 1257–1261. [Google Scholar] [CrossRef]
- Lin, C.K.E.; Kaptein, J.S.; Sheikh, J. Differential Expression of MicroRNAs and Their Possible Roles in Patients with Chronic Idiopathic Urticaria and Active Hives. Allergy Rhinol. 2017, 8, e67–e80. [Google Scholar] [CrossRef]
- Yoon, W.S.; Lee, S.S.; Chae, Y.S.; Park, Y.K. Therapeutic Effects of Recombinant Salmonella Typhimurium Harboring CCL22 MiRNA on Atopic Dermatitis-like Skin in Mice. Exp. Mol. Med. 2011, 43, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhang, T.; Zhang, S.; Tang, Q.; Yan, Y.; Feng, H. Integrated Bioinformatics and Validation Reveal IL1B and Its Related Molecules as Potential Biomarkers in Chronic Spontaneous Urticaria. Front. Immunol. 2022, 13, 850993. [Google Scholar] [CrossRef] [PubMed]
- Innao, V.; Allegra, A.; Pulvirenti, N.; Allegra, A.G.; Musolino, C. Therapeutic Potential of AntagomiRs in Haematological and Oncological Neoplasms. Eur. J. Cancer Care 2020, 29, e13208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lian, Y.; Xie, N.; Cheng, X.; Chen, C.; Xu, H.; Zheng, Y. Antagomirs Targeting MiR-142-5p Attenuate Pilocarpine-Induced Status Epilepticus in Mice. Exp. Cell Res. 2020, 393, 112089. [Google Scholar] [CrossRef]
- Reschke, C.R.; Silva, L.F.A.; Vangoor, V.R.; Rosso, M.; David, B.; Cavanagh, B.L.; Connolly, N.M.C.; Brennan, G.P.; Sanz-Rodriguez, A.; Mooney, C.; et al. Systemic Delivery of Antagomirs during Blood-Brain Barrier Disruption Is Disease-Modifying in Experimental Epilepsy. Mol. Ther. 2021, 29, 2041–2052. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study | miRNA | Study Population | Samples | Results | Molecular Pathways/Mechanism of Action |
|---|---|---|---|---|---|
| Sonkoly, E., et al. [54] 2010 | miR-155 | Humans (18) | Serum | Upregulated | Overexpression of miR-155 decreases CTLA-4 levels and increases proliferation in T helper cells, promoting chronic skin inflammation. |
| Rebane, A., et al. [58] 2014 | miR-146a | Humans/mice | Skin | Upregulated | miR-146a decreases the expression of IFN-γ-inducible genes CCL5, CCL8 and ubiquitin D (UBD) in keratinocytes and in a mouse model of AD by targeting the upstream mediators of NF-κB signaling—IRAK1 and CARD10. |
| Chen, X.F., et al. [63] 2014 | miR-151a | Humans (500) | Plasma | Overexpressed | miR-151a targets the IL-12 receptor β2 (IL12RB2), a subunit of the IL-12 receptor. |
| Jia, H.Z., et al. [66] 2018 | miR-223 | N/A | Serum | Upregulated | miR-223 expression is correlated with lower Treg cell numbers, a decreased number of which at birth correlates with an increased risk of AD. |
| Lv, Y., et al. [49] 2014 | miR-483-5p | Humans (30) | Serum and urine | Upregulated | miR-483-5p modulates fibrogenesis through the regulation of collagen homeostasis. |
| Vaher, H., et al. [51] 2019 | miR-10a-5p | Humans (10) | Skin (lesional and non-lesional) | Upregulated | miR-10a-5p is a direct target of HAS3, a damage-associated positive regulator of keratinocyte proliferation and migration. Upregulation of miR-10a-5p affects keratinocyte proliferation, thus impairing normal skin barrier function. |
| Gu, C., et al. [52] 2017 | miR-29b | Humans (21) | Lesional skin and serum | Upregulated | miR-29b triggers IFN-γ-mediated apoptosis of keratinocytes by targeting BCL2L. |
| Lv, Y., et al. [69] 2014 | miR-203 | Humans (30) | Serum | Upregulated | The miR-203 target gene is the regulator of cytokine production SOCS-3 (suppressor of cytokine signaling 3). |
| Skin | Upregulated | ||||
| Urine | Downregulated | ||||
| Zeng, Y.P., et al. [75] 2018 | miR-143 | N/A | Skin | Downregulated | miRNA-143 decreases IL-13 activity and inflammatory reaction by inhibiting IL-13 receptor-alpha1 (IL-13Ra1) in epidermal keratinocytes. |
| Yang, Z., et al. [77] 2017 | miR-124 | Humans (37) | Serum | Downregulated | miR-124 inhibits the p65 subunit of NF-kB and downregulates CCL5 and CCL8, thereby regulating inflammatory responses of keratinocytes and chronic skin inflammation in AD. |
| Study | miRNA | Study Population | Samples | Results | Molecular Pathways/Mechanism of Action |
|---|---|---|---|---|---|
| Werner et al. [81] 2020 | miR-142-3p, miR-142-5p, miR-146b-5p, miR-155-5p | Humans (nickel sulfate, epoxy resin (EP) and methylochloroisothia zolinone (MCI); n = 5 for each), irritants (sodium lauryl sulfate (SLS, n = 9) and nonanoic acid (NO, n = 5)) and from non-affected skin (baseline, n = 5). | Skin | Upregulated | miR-155-5p: enrichment of biological processes for axon guidance, smooth muscle cell migration and leukocyte/T cell apoptotic process. |
| Werner et al. [81] 2020 | miR-497-5p | Humans (nickel sulfate, epoxy resin (EP) and methylochloroisothia zolinone (MCI); n = 5 for each), irritants (sodium lauryl sulfate (SLS, n = 9) and nonanoic acid (NO, n = 5)) and from non-affected skin (baseline, n = 5). | Skin (patch tests with MCI) | Upregulated | T cell activation, cell–cell adhesion, cytokine and chemokine regulation pathways and a role in TGF-β-pathways via the regulation of SMAD3. |
| Werner et al. [81] 2020 | miR-23b-3p, miR-99a-5p, miR-193b-3p, miR-199a-3p | Humans (nickel sulfate, epoxy resin (EP) and methylochloroisothia zolinone (MCI); n = 5 for each), irritants (sodium lauryl sulfate (SLS, n = 9) and nonanoic acid (NO, n = 5)) and from non-affected skin (baseline, n = 5). | Skin (Patch Tests with MCI) | Upregulated | miR23b-3p and miR-99a-5p: skin homeostasis and development in vitro via TGIF1 and IGFR1. miR-193b-3p and miR-199a-3: leukocyte proliferation and keratinocyte/epidermis differentiation. |
| Vennegaard et al. [78] 2012 | miR-21, miR-223, miR-142-3p, miR-142-5p | Humans (nickel sulfate, epoxy resin (EP) and methylochloroisothia zolinone (MCI); n = 5 for each), irritants (sodium lauryl sulfate (SLS, n = 9) and nonanoic acid (NO, n = 5)) and from non-affected skin (baseline, n = 5). | Skin | Upregulated | T cells, T cell activation and skin inflammation. |
| Gulati et al. [79] 2015 | miR-21, miR-223, miR-142-3p, miR-142-5p | Humans (7) (DPCP at day 3, day 14 and day 120) | Skin | Upregulated | T cells, T cell activation and skin inflammation. |
| Anderson et al. [80] 2014 | miR-21, miR-22, miR-155, miR-126, miR-27b, miR-210, miR-31, miR-301a | Murine (toluene 2,4-diisocyanate (TDI)). | Skin | Upregulated | T cells, T cell activation and skin inflammation. |
| Study | miRNA | Study Population | Samples | Results | Molecular Pathways/Mechanism of Action |
|---|---|---|---|---|---|
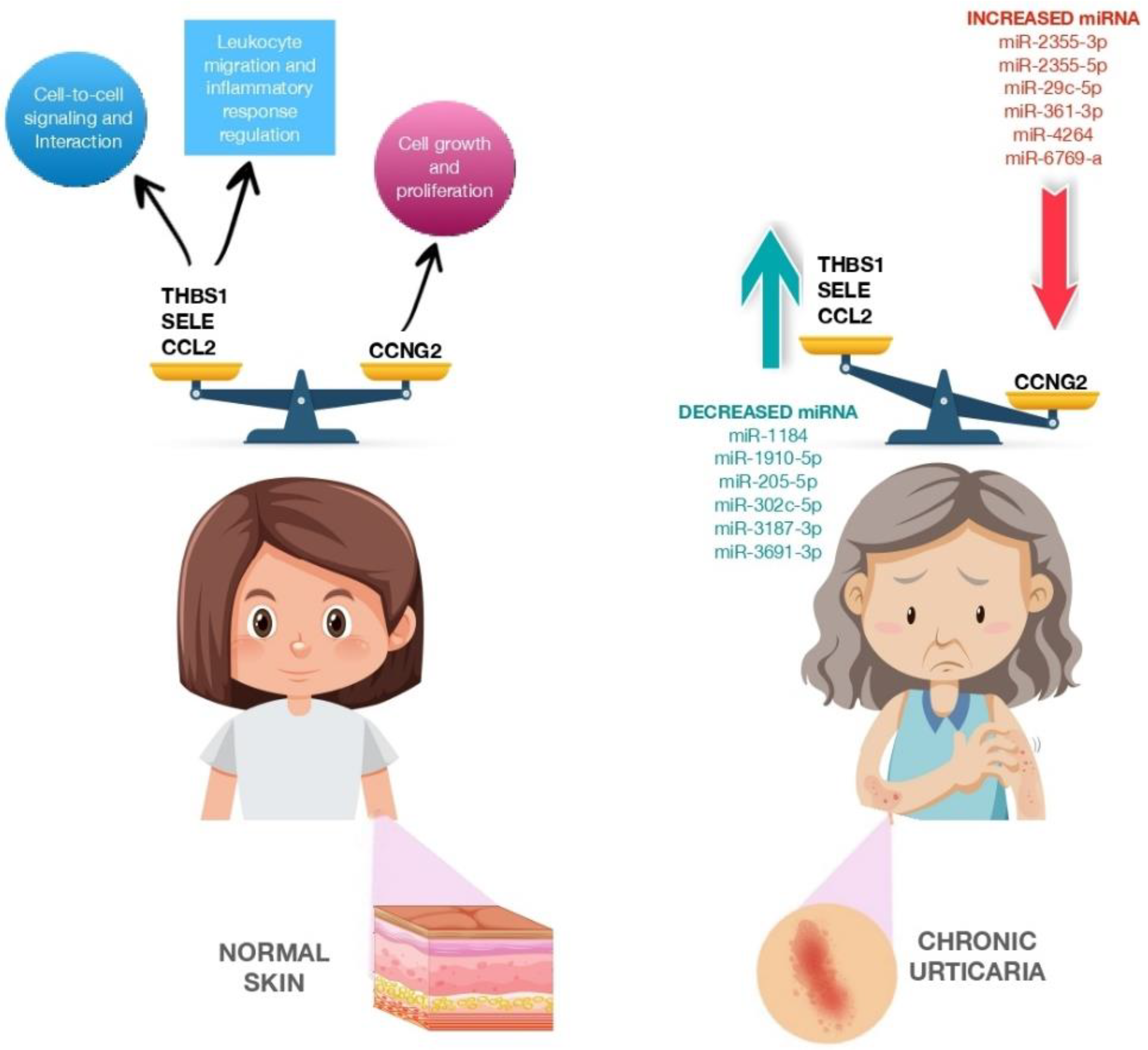
| Lin et al. [82] 2017 | miR-2355-3p miR-2355-5p miR-4264 miR-29c-5p miR-361-3p miR-6769a-5p | Humans (12) | Serum | Upregulated | Cell growth and proliferation |
| Lin et al. [82] 2017 | miR-1184 miR-1910-5p miR-205-5p miR-302c-5p miR-3187-3p miR-3691-3p miR-4649-5p miR-4733-5p miR-6799-3p miR-6800-3p | Humans (12) | Serum | Downregulated | Cell-to-cell signaling and interaction, cellular movement, regulation of leukocyte migration, tissue development immune cell trafficking, regulation of inflammatory response |
| Zhang et al. [33] 2019 | miR-125a-5p | Humans (20 active CIU patients and 20 healthy controls) | Serum | Upregulated | BLC2, STAT3, TGF-β and CCL17 |
| Key Points |
|---|
|
|
|
|
|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brancaccio, R.; Murdaca, G.; Casella, R.; Loverre, T.; Bonzano, L.; Nettis, E.; Gangemi, S. miRNAs’ Cross-Involvement in Skin Allergies: A New Horizon for the Pathogenesis, Diagnosis and Therapy of Atopic Dermatitis, Allergic Contact Dermatitis and Chronic Spontaneous Urticaria. Biomedicines 2023, 11, 1266. https://doi.org/10.3390/biomedicines11051266
Brancaccio R, Murdaca G, Casella R, Loverre T, Bonzano L, Nettis E, Gangemi S. miRNAs’ Cross-Involvement in Skin Allergies: A New Horizon for the Pathogenesis, Diagnosis and Therapy of Atopic Dermatitis, Allergic Contact Dermatitis and Chronic Spontaneous Urticaria. Biomedicines. 2023; 11(5):1266. https://doi.org/10.3390/biomedicines11051266
Chicago/Turabian StyleBrancaccio, Raffaele, Giuseppe Murdaca, Rossella Casella, Teresa Loverre, Laura Bonzano, Eustachio Nettis, and Sebastiano Gangemi. 2023. "miRNAs’ Cross-Involvement in Skin Allergies: A New Horizon for the Pathogenesis, Diagnosis and Therapy of Atopic Dermatitis, Allergic Contact Dermatitis and Chronic Spontaneous Urticaria" Biomedicines 11, no. 5: 1266. https://doi.org/10.3390/biomedicines11051266
APA StyleBrancaccio, R., Murdaca, G., Casella, R., Loverre, T., Bonzano, L., Nettis, E., & Gangemi, S. (2023). miRNAs’ Cross-Involvement in Skin Allergies: A New Horizon for the Pathogenesis, Diagnosis and Therapy of Atopic Dermatitis, Allergic Contact Dermatitis and Chronic Spontaneous Urticaria. Biomedicines, 11(5), 1266. https://doi.org/10.3390/biomedicines11051266