



Neuroinflammation, Oxidative Stress, Apoptosis, Microgliosis and Astrogliosis in the Cerebellum of Mice Chronically Exposed to Waterpipe Smoke


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. WPS Exposure
2.3. Cerebellum Collection and Homogenates Preparation
2.4. Measurement of the Concentrations of Tumor Necrosis Factor α (TNFα), Interleukin (IL)-6 and IL-1β
2.5. Quantification of Levels 8-Isoprostane, Thiobarbituric Acid Reactive Substances (TBARS) and Superoxide Dismutase (SOD)
2.6. Assessment of 8-Hydroxy-2′-Deoxyguanosine (8-OHdG), Cytochrome C, Cleaved Caspase-3, Nuclear Factor-κB (NF-κB) and Phosphorylated (Phospho)-NF-κB
2.7. Immunofluorescence Labeling
2.8. Statistical Analysis
3. Results
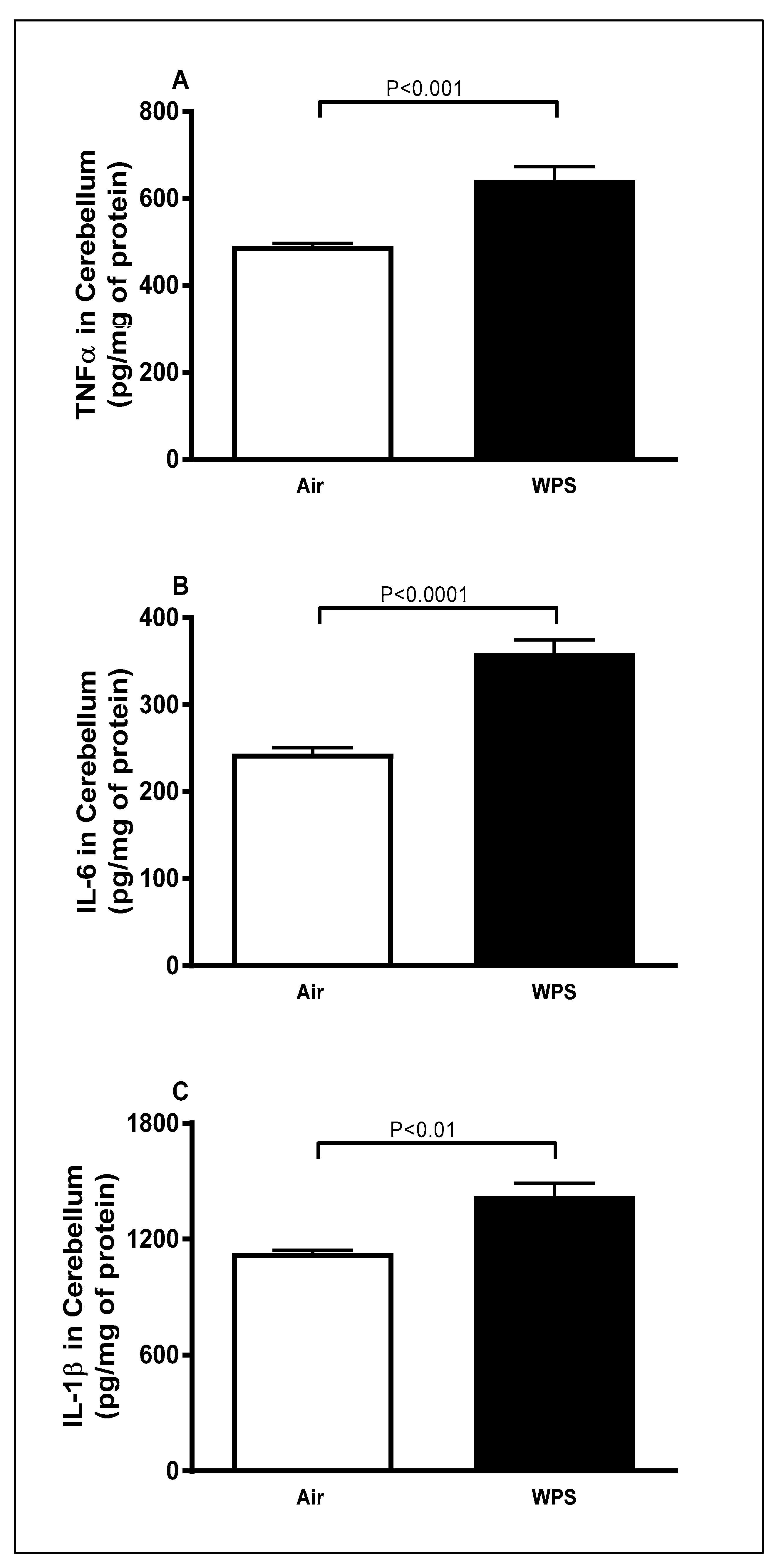
3.1. Effect of Chronic Exposure to WPS on the Concentrations of TNFα, IL-6 and IL-1β in the Cerebellum
3.2. Effect of Chronic Exposure to WPS on the Concentrations of 8-Isoprostane and TBARS and Activity of SOD in the Cerebellum
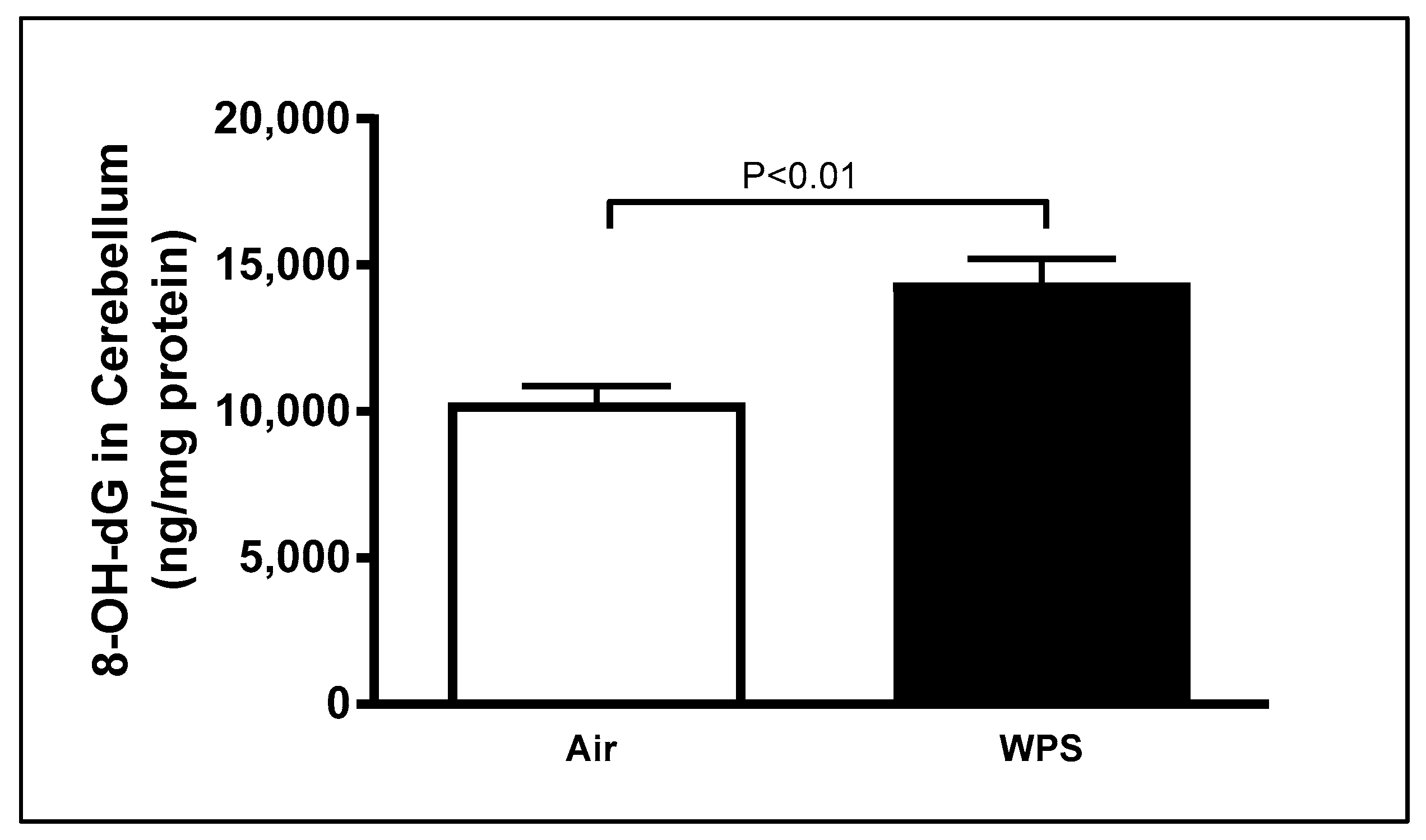
3.3. Effect of Chronic Exposure to WPS on the Concentrations of 8-OHdG in the Cerebellum
3.4. Effect of Chronic Exposure to WPS on the Levels of Cleaved Caspase-3 and Cytochrome C in the Cerebellum
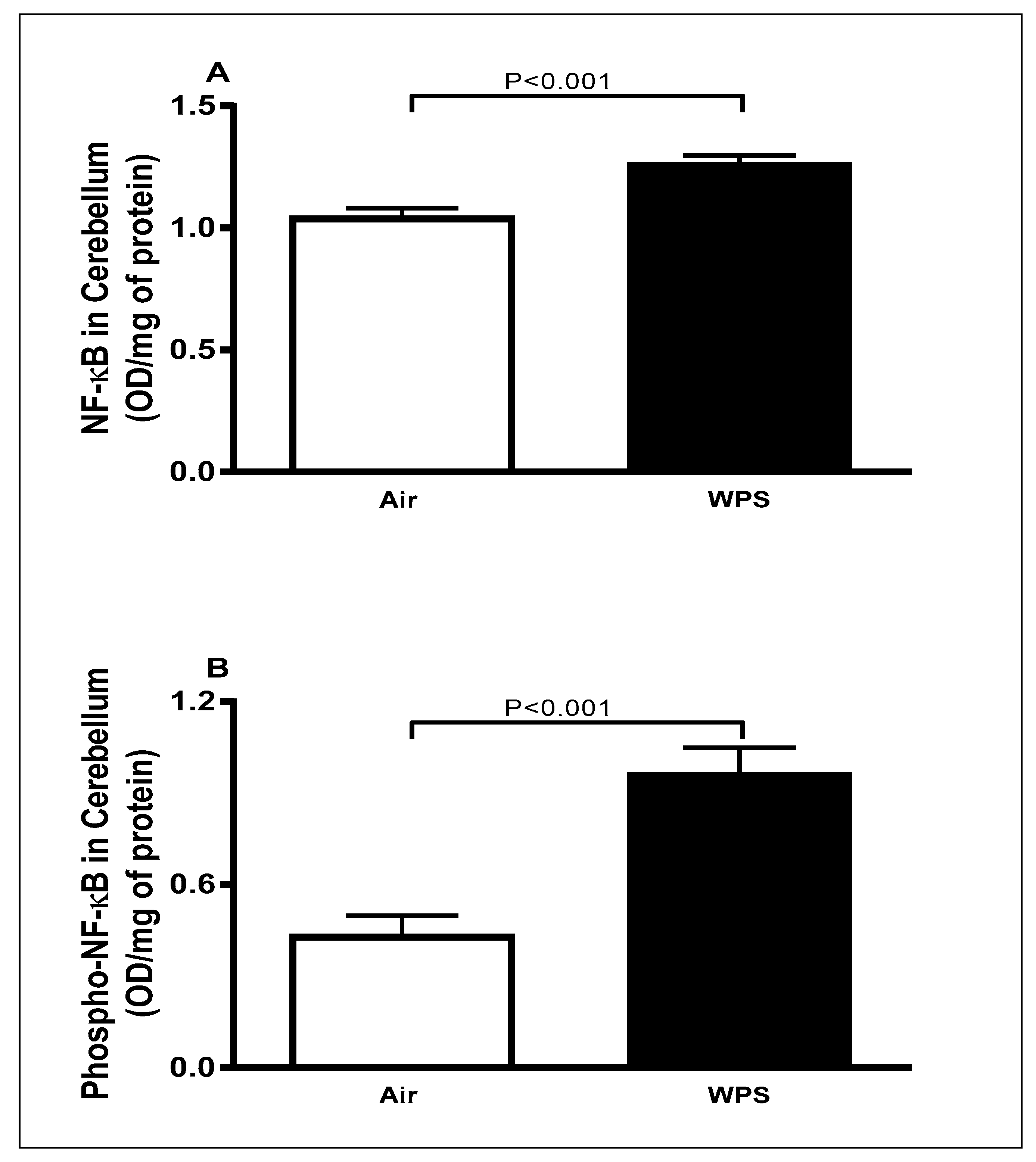
3.5. Effect of Chronic Exposure to WPS on the Expression of NF-κB and Phospho-NF-κB in the Cerebellum
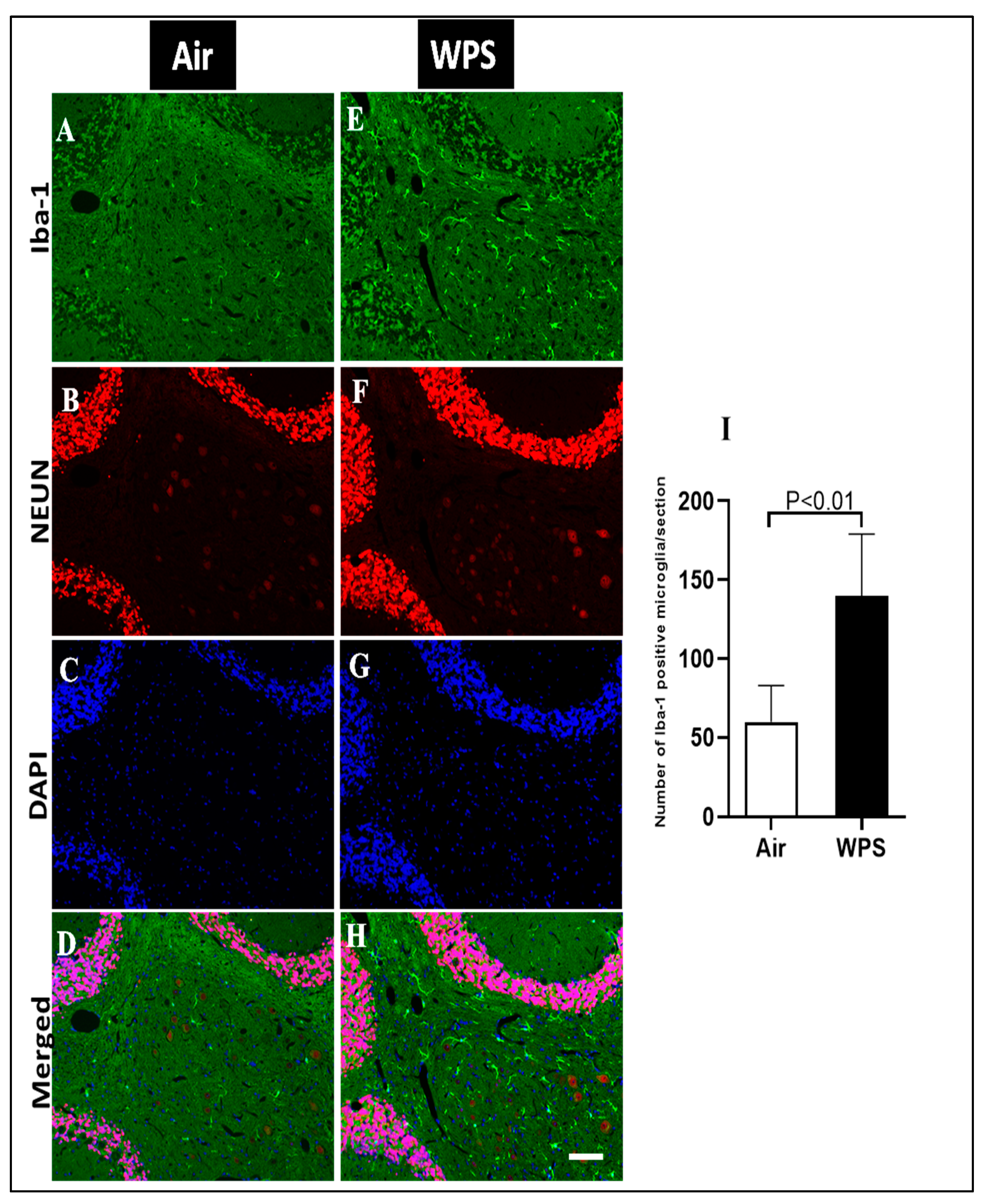
3.6. Effect of Chronic Exposure to WPS on Iba-1 Labeling and Quantification of Iba-1-Positive Microglia in the Cerebellum
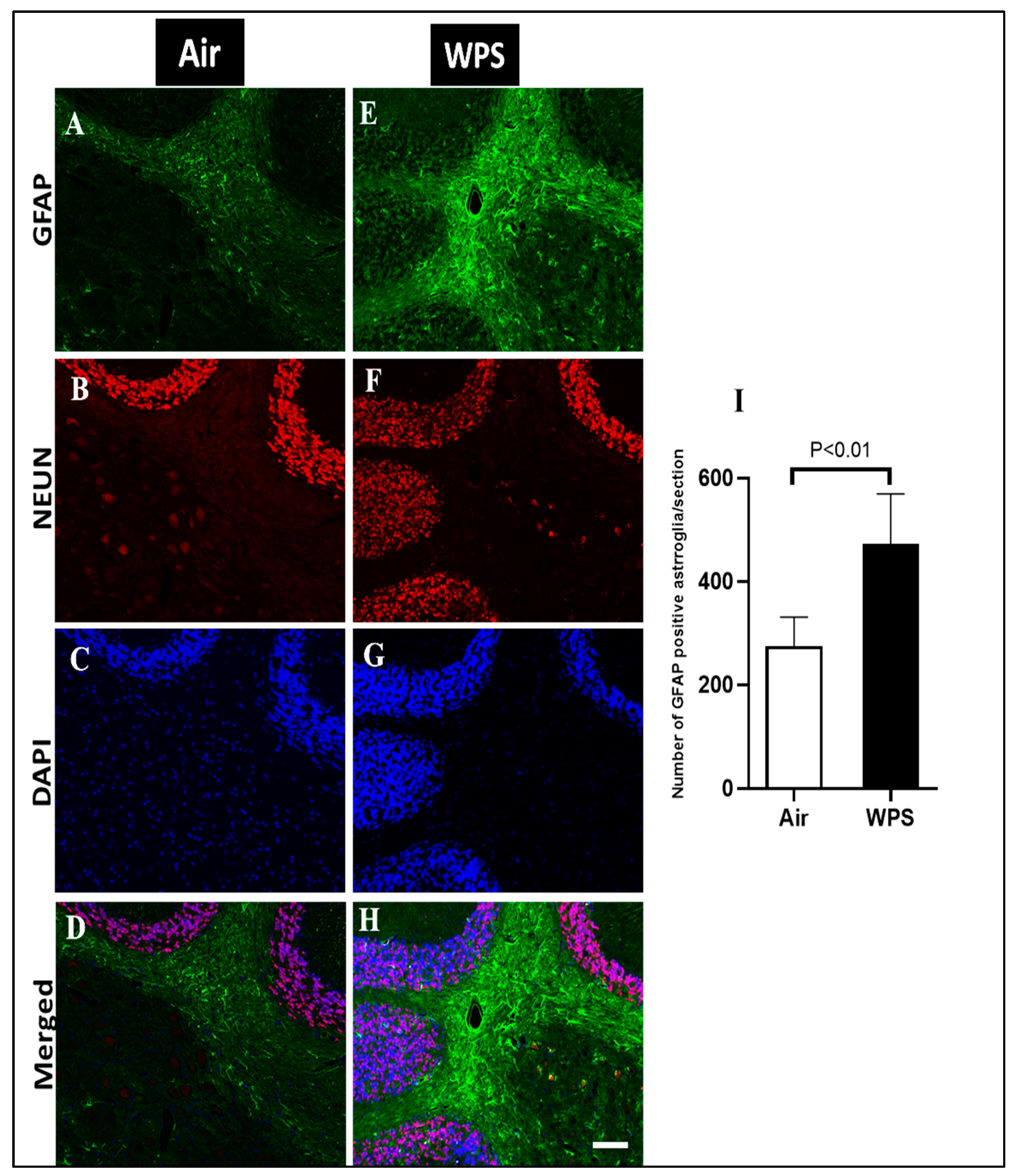
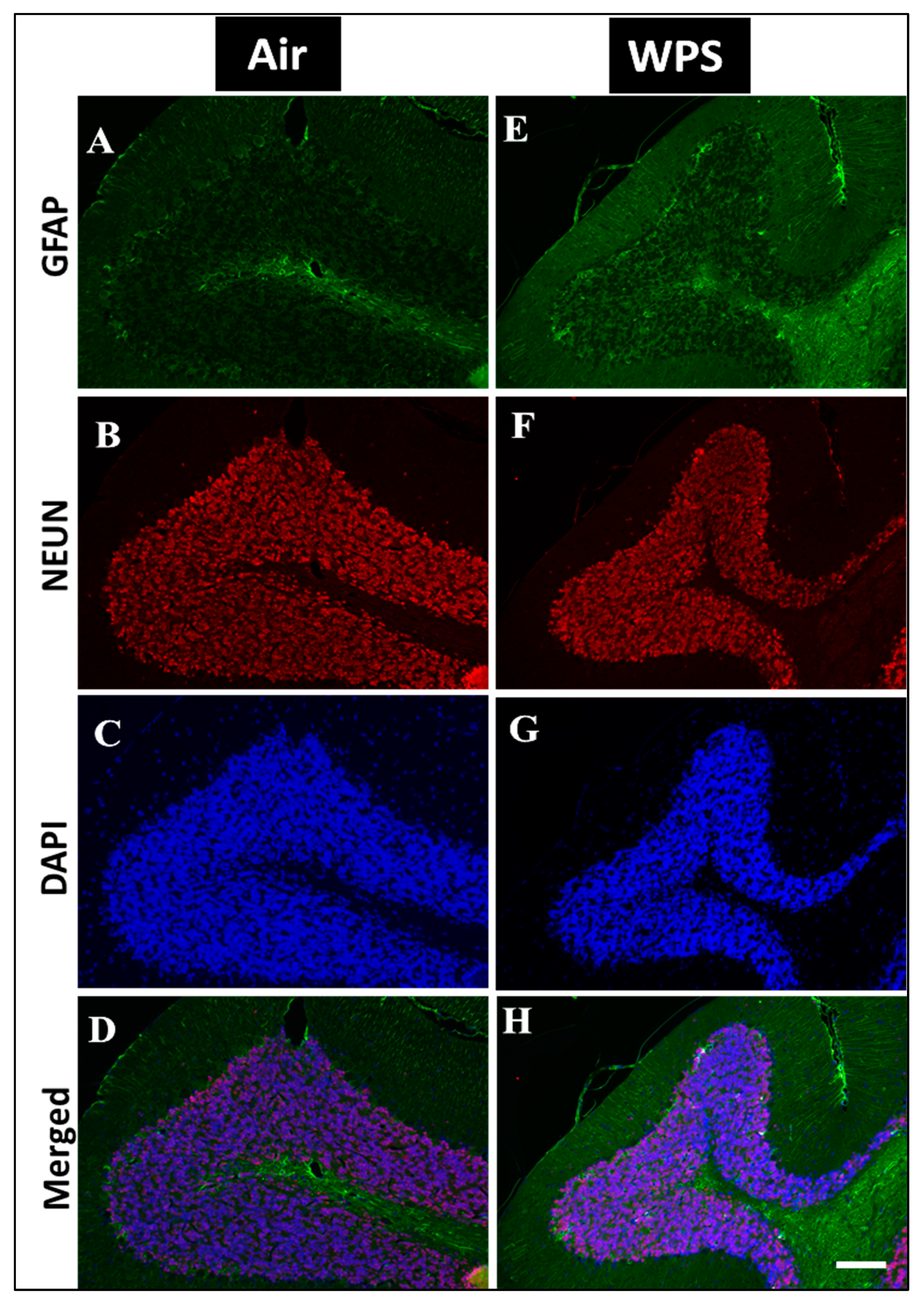
3.7. Effect of Chronic Exposure to WPS on the GFAP Labeling and Quantification of GFAP-Positive Astroglia in the Cerebellum
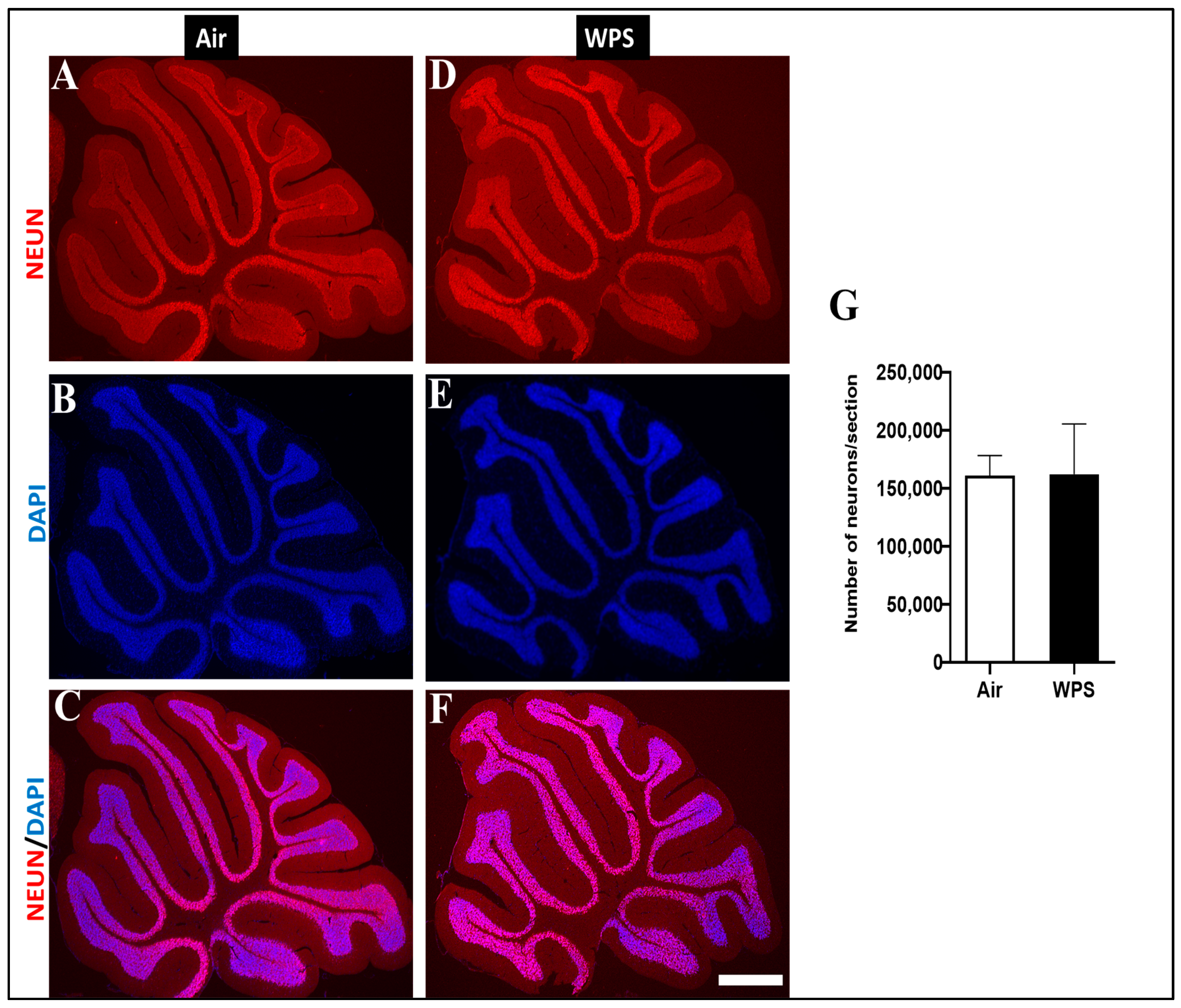
3.8. Effect of Chronic Exposure to WPS on the NEUN/DAPI Labeling and Quantification of Neurons in the Cerebellum
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bauer, U.E.; Briss, P.A.; Goodman, R.A.; Bowman, B.A. Prevention of chronic disease in the 21st century: Elimination of the leading preventable causes of premature death and disability in the USA. Lancet 2014, 384, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Maziak, W. The waterpipe: An emerging global risk for cancer. Cancer Epidemiol. 2013, 37, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Jamal, A.; Phillips, E.; Gentzke, A.S.; Homa, D.M.; Babb, S.D.; King, B.A.; Neff, L.J. Current Cigarette Smoking Among Adults-United States, 2016. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Salloum, R.G.; Thrasher, J.F.; Kates, F.R.; Maziak, W. Water pipe tobacco smoking in the United States: Findings from the National Adult Tobacco Survey. Prev. Med. 2015, 71, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Ward, K.D. The waterpipe: An emerging global epidemic in need of action. Tob. Control 2015, 24 (Suppl. S1), i1–i2. [Google Scholar] [CrossRef]
- Javed, F.; Alharthi, S.S.; BinShabaib, M.S.; Gajendra, S.; Romanos, G.E.; Rahman, I. Toxicological impact of waterpipe smoking and flavorings in the oral cavity and respiratory system. Inhal. Toxicol. 2017, 29, 389–396. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Yuvaraju, P.; Beegam, S.; Yasin, J.; Ali, B.H. Chronic exposure to water-pipe smoke induces cardiovascular dysfunction in mice. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H329–H339. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Zaaba, N.E.; Yasin, J.; Ali, B.H. Waterpipe Tobacco Smoke Inhalation Triggers Thrombogenicity, Cardiac Inflammation and Oxidative Stress in Mice: Effects of Flavouring. Int. J. Mol. Sci. 2020, 21, 1291. [Google Scholar] [CrossRef] [PubMed]
- Sibai, A.M.; Tohme, R.A.; Almedawar, M.M.; Itani, T.; Yassine, S.I.; Nohra, E.A.; Isma’eel, H.A. Lifetime cumulative exposure to waterpipe smoking is associated with coronary artery disease. Atherosclerosis 2014, 234, 454–460. [Google Scholar] [CrossRef]
- Ali, B.H.; Al Balushi, K.A.; Ashique, M.; Shalaby, A.; Al Kindi, M.A.; Adham, S.A.; Karaca, T.; Beegam, S.; Yuvaraju, P.; Nemmar, A. Chronic Water-Pipe Smoke Exposure Induces Injurious Effects to Reproductive System in Male Mice. Front. Physiol. 2017, 8, 158. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Ali, B.H. Comparative Study on Pulmonary Toxicity in Mice Induced by Exposure to Unflavoured and Apple- and Strawberry-Flavoured Tobacco Waterpipe Smoke. Oxidative Med. Cell. Longev. 2020, 2020, 6450450. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Al Hemeiri, A.; Al Hammadi, N.; Yuvaraju, P.; Beegam, S.; Yasin, J.; Elwasila, M.; Ali, B.H.; Adeghate, E. Early pulmonary events of nose-only water pipe (shisha) smoking exposure in mice. Physiol. Rep. 2015, 3, e12258. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, F.K.; Er, M.; Hasanoglu, H.C.; Kilic, H.; Senturk, A.; Karalezli, A.; Ergin, M.; Erel, O. Deteriorations of pulmonary function, elevated carbon monoxide levels and increased oxidative stress amongst water-pipe smokers. Int. J. Occup. Med. Environ. Health 2017, 30, 731–742. [Google Scholar] [CrossRef]
- Shah, R.S.; Cole, J.W. Smoking and stroke: The more you smoke the more you stroke. Expert Rev. Cardiovasc. Ther. 2010, 8, 917–932. [Google Scholar] [CrossRef]
- Cataldo, J.K.; Prochaska, J.J.; Glantz, S.A. Cigarette smoking is a risk factor for Alzheimer’s Disease: An analysis controlling for tobacco industry affiliation. J. Alzheimer’s Dis. JAD 2010, 19, 465–480. [Google Scholar] [CrossRef]
- Sundström, P.; Nyström, L.; Hallmans, G. Smoke exposure increases the risk for multiple sclerosis. Eur. J. Neurol. 2008, 15, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, H.; Wang, J.; Xue, Q.; Yang, X.; Kang, Y.; Li, M.; Xu, J.; Li, G.; Li, C.; et al. Association of Cigarette Smoking With Cerebrospinal Fluid Biomarkers of Neurodegeneration, Neuroinflammation, and Oxidation. JAMA Netw. Open 2020, 3, e2018777. [Google Scholar] [CrossRef]
- Luchese, C.; Pinton, S.; Nogueira, C.W. Brain and lungs of rats are differently affected by cigarette smoke exposure: Antioxidant effect of an organoselenium compound. Pharmacol. Res. 2009, 59, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Guo, M.; Mehra, M.; Royal, W., 3rd. Inflammation and oxidative stress induced by cigarette smoke in Lewis rat brains. J. Neuroimmunol. 2013, 254, 69–75. [Google Scholar] [CrossRef]
- Ramesh, T.; Sureka, C.; Bhuvana, S.; Begum, V.H. Oxidative stress in the brain of cigarette smoke-induced noxiousness: Neuroprotective role of Sesbania grandiflora. Metab. Brain Dis. 2015, 30, 573–582. [Google Scholar] [CrossRef]
- Anbarasi, K.; Vani, G.; Balakrishna, K.; Devi, C.S. Effect of bacoside A on brain antioxidant status in cigarette smoke exposed rats. Life Sci. 2006, 78, 1378–1384. [Google Scholar] [CrossRef]
- Sivandzade, F.; Alqahtani, F.; Sifat, A.; Cucullo, L. The cerebrovascular and neurological impact of chronic smoking on post-traumatic brain injury outcome and recovery: An in vivo study. J. Neuroinflamm. 2020, 17, 133. [Google Scholar] [CrossRef] [PubMed]
- Vani, G.; Anbarasi, K.; Shyamaladevi, C.S. Bacoside A: Role in Cigarette Smoking Induced Changes in Brain. Evid.-Based Complement. Altern. Med. Ecam 2015, 2015, 286137. [Google Scholar] [CrossRef] [PubMed]
- Vayssier, M.; Banzet, N.; François, D.; Bellmann, K.; Polla, B.S. Tobacco smoke induces both apoptosis and necrosis in mammalian cells: Differential effects of HSP70. Am. J. Physiol. 1998, 275, L771-9. [Google Scholar] [CrossRef]
- Alomari, M.A.; Al-Sheyab, N.A.; Khabour, O.F.; Alzoubi, K.H. Brain-derived neutrophic factor in adolescents smoking waterpipe: The Irbid TRY. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2018, 67, 14–18. [Google Scholar] [CrossRef]
- Karakayalı, O.; Utku, U.; Yılmaz, S. Evaluation of Cerebral Blood Flow Alterations and Acute Neuronal Damage due to Water-Pipe Smoking. Balk. Med. J. 2019, 36, 106–112. [Google Scholar] [CrossRef]
- Toukan, Y.; Hakim, F.; Bentur, Y.; Aharon-Peretz, J.; Elemy, A.; Gur, M.; Hanna, M.; Fisher, T.; Scherb, I.; Bentur, L. The Effect of a 30-Min Water-Pipe Smoking Session on Cognitive Measures and Cardio-Pulmonary Parameters. Nicotine Tob. Res. Off. J. Soc. Res. Nicotine Tob. 2020, 22, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.M.; Swiss, G.M.S.; Hall, F.S.; Hikmat, S.; Sari, Y.; Al-Qirim, T.M.; Amawi, H.A. Ceftriaxone Reduces Waterpipe Tobacco Smoke Withdrawal-induced Anxiety in rats via Modulating the Expression of TNF-α/NFĸB, Nrf2, and GLT-1. Neuroscience 2021, 463, 128–142. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Khabour, O.F.; Alharahshah, E.A.; Alhashimi, F.H.; Shihadeh, A.; Eissenberg, T. The Effect of Waterpipe Tobacco Smoke Exposure on Learning and Memory Functions in the Rat Model. J. Mol. Neurosci. MN 2015, 57, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Halboup, A.M.; Alomari, M.A.; Khabour, O.F. The neuroprotective effect of vitamin E on waterpipe tobacco smoking-induced memory impairment: The antioxidative role. Life Sci. 2019, 222, 46–52. [Google Scholar] [CrossRef]
- Klein, A.P.; Ulmer, J.L.; Quinet, S.A.; Mathews, V.; Mark, L.P. Nonmotor Functions of the Cerebellum: An Introduction. AJNR. Am. J. Neuroradiol. 2016, 37, 1005–1009. [Google Scholar] [CrossRef]
- Popa, L.S.; Hewitt, A.L.; Ebner, T.J. The cerebellum for jocks and nerds alike. Front. Syst. Neurosci. 2014, 8, 113. [Google Scholar] [CrossRef]
- Schmahmann, J.D. The cerebellum and cognition. Neurosci. Lett. 2019, 688, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Beuriat, P.A.; Cohen-Zimerman, S.; Smith, G.N.L.; Krueger, F.; Gordon, B.; Grafman, J. A New Insight on the Role of the Cerebellum for Executive Functions and Emotion Processing in Adults. Front. Neurol. 2020, 11, 593490. [Google Scholar] [CrossRef] [PubMed]
- Hoche, F.; Guell, X.; Sherman, J.C.; Vangel, M.G.; Schmahmann, J.D. Cerebellar Contribution to Social Cognition. Cerebellum 2016, 15, 732–743. [Google Scholar] [CrossRef]
- Kühn, S.; Romanowski, A.; Schilling, C.; Mobascher, A.; Warbrick, T.; Winterer, G.; Gallinat, J. Brain grey matter deficits in smokers: Focus on the cerebellum. Brain Struct. Funct. 2012, 217, 517–522. [Google Scholar] [CrossRef]
- Shen, Z.; Huang, P.; Wang, C.; Qian, W.; Yang, Y.; Zhang, M. Cerebellar Gray Matter Reductions Associate With Decreased Functional Connectivity in Nicotine-Dependent Individuals. Nicotine Tob. Res. Off. J. Soc. Res. Nicotine Tob. 2018, 20, 440–447. [Google Scholar] [CrossRef]
- Nemmar, A.; Yuvaraju, P.; Beegam, S.; John, A.; Raza, H.; Ali, B.H. Cardiovascular effects of nose-only water-pipe smoking exposure in mice. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H740–H746. [Google Scholar] [CrossRef]
- Nemmar, A.; Raza, H.; Yuvaraju, P.; Beegam, S.; John, A.; Yasin, J.; Hameed, R.S.; Adeghate, E.; Ali, B.H. Nose-only water-pipe smoking effects on airway resistance, inflammation, and oxidative stress in mice. J. Appl. Physiol. 2013, 115, 1316–1323. [Google Scholar] [CrossRef]
- Nemmar, A.; Yuvaraju, P.; Beegam, S.; Ali, B.H. Short-term nose-only water-pipe (shisha) smoking exposure accelerates coagulation and causes cardiac inflammation and oxidative stress in mice. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 35, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Al-Salam, S.; Yuvaraju, P.; Beegam, S.; Yasin, J.; Ali, B.H. Chronic Exposure to Water-Pipe Smoke Induces Alveolar Enlargement, DNA Damage and Impairment of Lung Function. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 38, 982–992. [Google Scholar] [CrossRef]
- Hamadi, N.; Sheikh, A.; Madjid, N.; Lubbad, L.; Amir, N.; Shehab, S.A.; Khelifi-Touhami, F.; Adem, A. Increased pro-inflammatory cytokines, glial activation and oxidative stress in the hippocampus after short-term bilateral adrenalectomy. BMC Neurosci. 2016, 17, 61. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Hamadi, N.; Ali, B.H. In Vivo Protective Effects of Nootkatone against Particles-Induced Lung Injury Caused by Diesel Exhaust Is Mediated via the NF-κB Pathway. Nutrients 2018, 10, 263. [Google Scholar] [CrossRef]
- Nemmar, A.; Beegam, S.; Yuvaraju, P.; Yasin, J.; Ali, B.H.; Adeghate, E. Nose-Only Water-Pipe Smoke Exposure in Mice Elicits Renal Histopathological Alterations, Inflammation, Oxidative Stress, DNA Damage, and Apoptosis. Front. Physiol. 2020, 11, 46. [Google Scholar] [CrossRef]
- Arunachalam, S.; Nagoor Meeran, M.F.; Azimullah, S.; Sharma, C.; Goyal, S.N.; Ojha, S. Nerolidol Attenuates Oxidative Stress, Inflammation, and Apoptosis by Modulating Nrf2/MAPK Signaling Pathways in Doxorubicin-Induced Acute Cardiotoxicity in Rats. Antioxidants 2021, 10, 984. [Google Scholar] [CrossRef]
- Juan, D.; Zhou, D.H.; Li, J.; Wang, J.Y.; Gao, C.; Chen, M. A 2-year follow-up study of cigarette smoking and risk of dementia. Eur. J. Neurol. 2004, 11, 277–282. [Google Scholar] [CrossRef]
- Ho, Y.S.; Yang, X.; Yeung, S.C.; Chiu, K.; Lau, C.F.; Tsang, A.W.; Mak, J.C.; Chang, R.C. Cigarette smoking accelerated brain aging and induced pre-Alzheimer-like neuropathology in rats. PLoS ONE 2012, 7, e36752. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Kim, C.; Kim, H.J.; Ye, B.S.; Kim, Y.J.; Jung, N.Y.; Son, T.O.; Cho, E.B.; Jang, H.; Lee, J.; et al. Impact of smoking on neurodegeneration and cerebrovascular disease markers in cognitively normal men. Eur. J. Neurol. 2016, 23, 110–119. [Google Scholar] [CrossRef]
- Rauf, A.; Badoni, H.; Abu-Izneid, T.; Olatunde, A.; Rahman, M.M.; Painuli, S.; Semwal, P.; Wilairatana, P.; Mubarak, M.S. Neuroinflammatory Markers: Key Indicators in the Pathology of Neurodegenerative Diseases. Molecules 2022, 27, 3194. [Google Scholar] [CrossRef] [PubMed]
- Aljadani, R.H.; Algabbani, A.M.; Alamir, J.A.; Alqahtani, A.S.; BinDhim, N.F. Waterpipe Tobacco Chemical Content, Microbial Contamination, and Genotoxic Effects: A Systematic Review. Int. J. Toxicol. 2020, 39, 256–262. [Google Scholar] [CrossRef]
- Weng, M.W.; Hsiao, Y.M.; Chen, C.J.; Wang, J.P.; Chen, W.C.; Ko, J.L. Benzo[a]pyrene diol epoxide up-regulates COX-2 expression through NF-kappaB in rat astrocytes. Toxicol. Lett. 2004, 151, 345–355. [Google Scholar] [CrossRef]
- Ghosh, D.; Mishra, M.K.; Das, S.; Kaushik, D.K.; Basu, A. Tobacco carcinogen induces microglial activation and subsequent neuronal damage. J. Neurochem. 2009, 110, 1070–1081. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, L.; Fernandez, F.; Johnson, J.B.; Naiker, M.; Owoola, A.G.; Broszczak, D.A. Oxidative stress in alzheimer’s disease: A review on emergent natural polyphenolic therapeutics. Complement. Ther. Med. 2020, 49, 102294. [Google Scholar] [CrossRef]
- Jenner, P. Oxidative stress in Parkinson’s disease. Ann. Neurol. 2003, 53 (Suppl. S3), S26–S36; discussion S36–S38. [Google Scholar] [CrossRef]
- Ohl, K.; Tenbrock, K.; Kipp, M. Oxidative stress in multiple sclerosis: Central and peripheral mode of action. Exp. Neurol. 2016, 277, 58–67. [Google Scholar] [CrossRef]
- Coppedè, F.; Migliore, L. DNA damage in neurodegenerative diseases. Mutat. Res. 2015, 776, 84–97. [Google Scholar] [CrossRef]
- Martinet, W.; Knaapen, M.W.; De Meyer, G.R.; Herman, A.G.; Kockx, M.M. Elevated levels of oxidative DNA damage and DNA repair enzymes in human atherosclerotic plaques. Circulation 2002, 106, 927–932. [Google Scholar] [CrossRef]
- Neofytou, E.; Tzortzaki, E.G.; Chatziantoniou, A.; Siafakas, N.M. DNA damage due to oxidative stress in Chronic Obstructive Pulmonary Disease (COPD). Int. J. Mol. Sci. 2012, 13, 16853–16864. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2’ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health. Part C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef]
- Mattson, M.P. Apoptosis in neurodegenerative disorders. Nat. Rev. Mol. Cell Biol. 2000, 1, 120–129. [Google Scholar] [CrossRef]
- Wallauer, M.M.; Huf, F.; Tortorelli, L.S.; Rahmeier, F.L.; Carvalho, F.B.; Meurer, R.T.; da Cruz Fernandes, M. Morphological changes in the cerebellum as a result of ethanol treatment and cigarette smoke exposure: A study on astrogliosis, apoptosis and Purkinje cells. Neurosci. Lett. 2018, 672, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Yang, J.; Jones, D.P. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Et Biophys. Acta 1998, 1366, 139–149. [Google Scholar] [CrossRef]
- Hosseini, M.-J.; Naserzadeh, P.; Salimi, A.; Pourahmad, J. Toxicity of cigarette smoke on isolated lung, heart, and brain mitochondria: Induction of oxidative stress and cytochrome c release. Toxicol. Environ. Chem. 2013, 95, 1624–1637. [Google Scholar] [CrossRef]
- Ali, B.H.; Al-Salam, S.; Adham, S.A.; Al Balushi, K.; Al Za’abi, M.; Beegam, S.; Yuvaraju, P.; Manoj, P.; Nemmar, A. Testicular Toxicity of Water Pipe Smoke Exposure in Mice and the Effect of Treatment with Nootkatone Thereon. Oxidative Med. Cell. Longev. 2019, 2019, 2416935. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Helweg, L.P.; Greiner, J.F.W.; Kaltschmidt, C. NF-κB in neurodegenerative diseases: Recent evidence from human genetics. Front. Mol. Neurosci. 2022, 15, 954541. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Zaaba, N.E.; Elzaki, O.; Yasin, J.; Ali, B.H. Waterpipe smoke-induced hypercoagulability and cardiac injury in mice: Influence of cessation of exposure. Biomed. Pharmacother. Biomed. Pharmacother. 2022, 146, 112493. [Google Scholar] [CrossRef]
- Waisman, A.; Ginhoux, F.; Greter, M.; Bruttger, J. Homeostasis of Microglia in the Adult Brain: Review of Novel Microglia Depletion Systems. Trends Immunol. 2015, 36, 625–636. [Google Scholar] [CrossRef]
- Song, N.; Wang, J.; Jiang, H.; Xie, J. Astroglial and microglial contributions to iron metabolism disturbance in Parkinson’s disease. Biochim. Et Biophys. Acta. Mol. Basis Dis. 2018, 1864, 967–973. [Google Scholar] [CrossRef]
- Ransohoff, R.M. Microgliosis: The questions shape the answers. Nat. Neurosci. 2007, 10, 1507–1509. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Nissen, J.C.; Ji, K.; Tsirka, S.E. The experimental autoimmune encephalomyelitis disease course is modulated by nicotine and other cigarette smoke components. PLoS ONE 2014, 9, e107979. [Google Scholar] [CrossRef]
- Liu, Z.; Chopp, M. Astrocytes, therapeutic targets for neuroprotection and neurorestoration in ischemic stroke. Prog. Neurobiol. 2016, 144, 103–120. [Google Scholar] [CrossRef]
- Ransom, B.R.; Ransom, C.B. Astrocytes: Multitalented stars of the central nervous system. Methods Mol. Biol. (Clifton N.J.) 2012, 814, 3–7. [Google Scholar]
- Katz, I.R.; Iacovitti, L.M.; Reis, D.J. In vitro studies on the characterization of cellular proliferation following neuronal injury in the adult rat brain. J. Neuroimmunol. 1990, 29, 33–48. [Google Scholar] [CrossRef]
- Abdel-Rahman, A.; Dechkovskaia, A.M.; Sutton, J.M.; Chen, W.C.; Guan, X.; Khan, W.A.; Abou-Donia, M.B. Maternal exposure of rats to nicotine via infusion during gestation produces neurobehavioral deficits and elevated expression of glial fibrillary acidic protein in the cerebellum and CA1 subfield in the offspring at puberty. Toxicology 2005, 209, 245–261. [Google Scholar] [CrossRef]
- Fuller, B.F.; Gold, M.S.; Wang, K.K.; Ottens, A.K. Effects of environmental tobacco smoke on adult rat brain biochemistry. J. Mol. Neurosci. MN 2010, 41, 165–171. [Google Scholar] [CrossRef]
- Rouach, N.; Avignone, E.; Même, W.; Koulakoff, A.; Venance, L.; Blomstrand, F.; Giaume, C. Gap junctions and connexin expression in the normal and pathological central nervous system. Biol. Cell 2002, 94, 457–475. [Google Scholar] [CrossRef]
- Ransohoff, R.M.; Brown, M.A. Innate immunity in the central nervous system. J. Clin. Investig. 2012, 122, 1164–1171. [Google Scholar] [CrossRef]
- Sprung, C.N.; Ivashkevich, A.; Forrester, H.B.; Redon, C.E.; Georgakilas, A.; Martin, O.A. Oxidative DNA damage caused by inflammation may link to stress-induced non-targeted effects. Cancer Lett. 2015, 356, 72–81. [Google Scholar] [CrossRef]
- Salucci, S.; Bartoletti Stella, A.; Battistelli, M.; Burattini, S.; Bavelloni, A.; Cocco, L.I.; Gobbi, P.; Faenza, I. How Inflammation Pathways Contribute to Cell Death in Neuro-Muscular Disorders. Biomolecules 2021, 11, 1109. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamadi, N.; Beegam, S.; Zaaba, N.E.; Elzaki, O.; Altamimi, M.A.; Nemmar, A. Neuroinflammation, Oxidative Stress, Apoptosis, Microgliosis and Astrogliosis in the Cerebellum of Mice Chronically Exposed to Waterpipe Smoke. Biomedicines 2023, 11, 1104. https://doi.org/10.3390/biomedicines11041104
Hamadi N, Beegam S, Zaaba NE, Elzaki O, Altamimi MA, Nemmar A. Neuroinflammation, Oxidative Stress, Apoptosis, Microgliosis and Astrogliosis in the Cerebellum of Mice Chronically Exposed to Waterpipe Smoke. Biomedicines. 2023; 11(4):1104. https://doi.org/10.3390/biomedicines11041104
Chicago/Turabian StyleHamadi, Naserddine, Sumaya Beegam, Nur Elena Zaaba, Ozaz Elzaki, Mariam Abdulla Altamimi, and Abderrahim Nemmar. 2023. "Neuroinflammation, Oxidative Stress, Apoptosis, Microgliosis and Astrogliosis in the Cerebellum of Mice Chronically Exposed to Waterpipe Smoke" Biomedicines 11, no. 4: 1104. https://doi.org/10.3390/biomedicines11041104
APA StyleHamadi, N., Beegam, S., Zaaba, N. E., Elzaki, O., Altamimi, M. A., & Nemmar, A. (2023). Neuroinflammation, Oxidative Stress, Apoptosis, Microgliosis and Astrogliosis in the Cerebellum of Mice Chronically Exposed to Waterpipe Smoke. Biomedicines, 11(4), 1104. https://doi.org/10.3390/biomedicines11041104