
Regeneration of Panniculus Carnosus Muscle in Fetal Mice Is Characterized by the Presence of Actin Cables

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Wound Model
2.3. Preparation of Tissue Sections
2.4. Staining Tissue Sections
2.5. Imaging
2.6. Statistical Analysis
3. Results
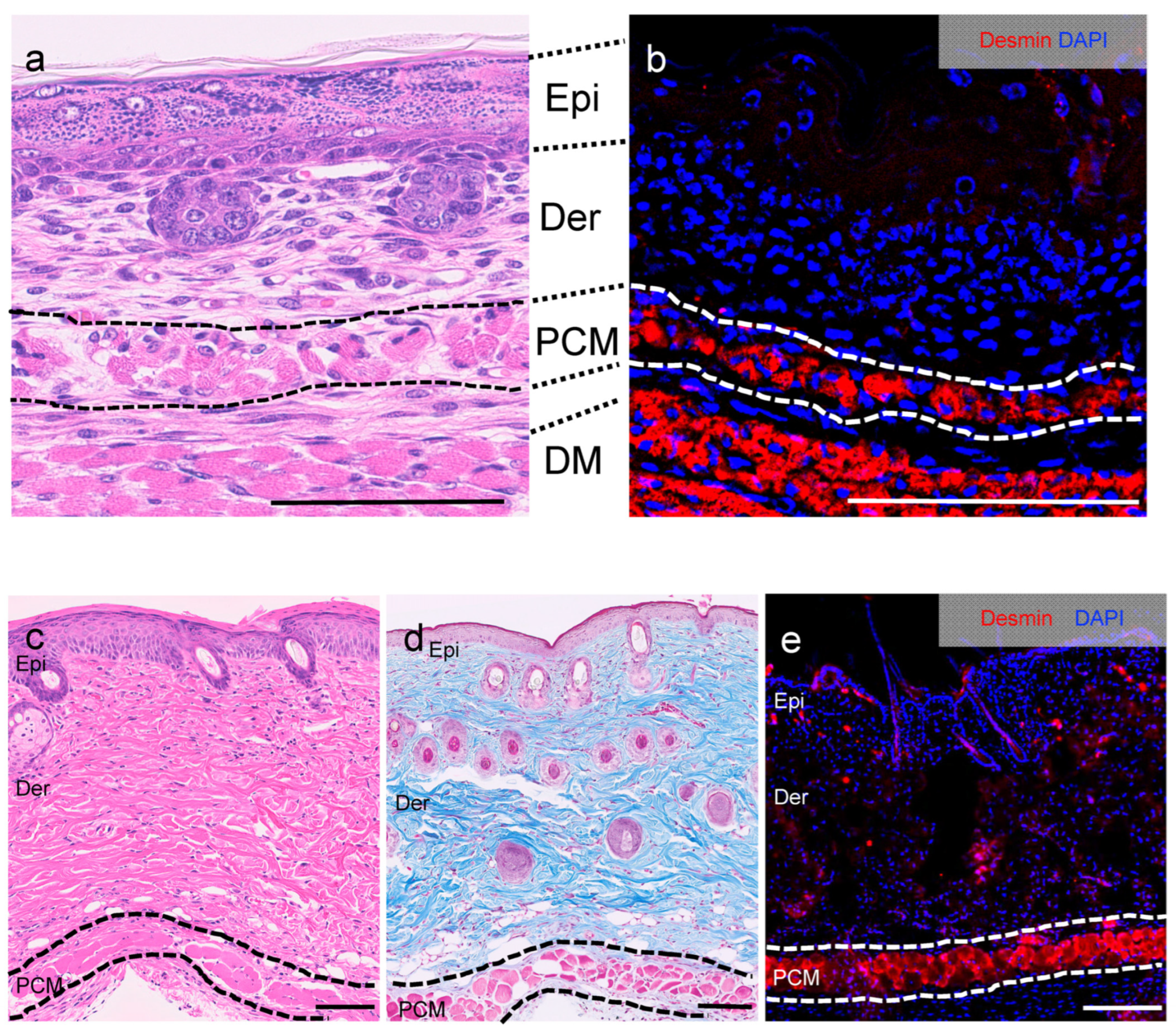
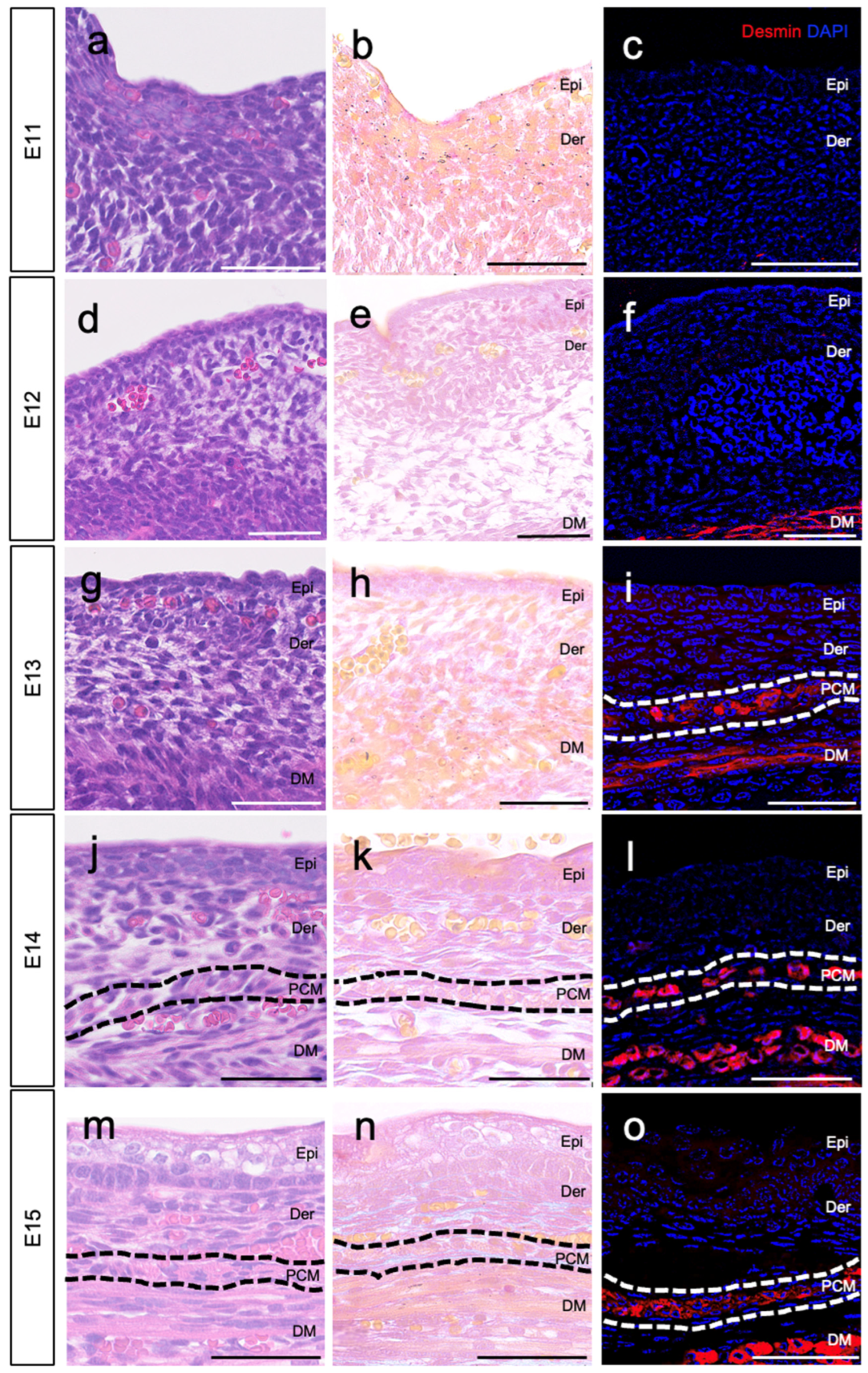
3.1. Location of PCM on E13
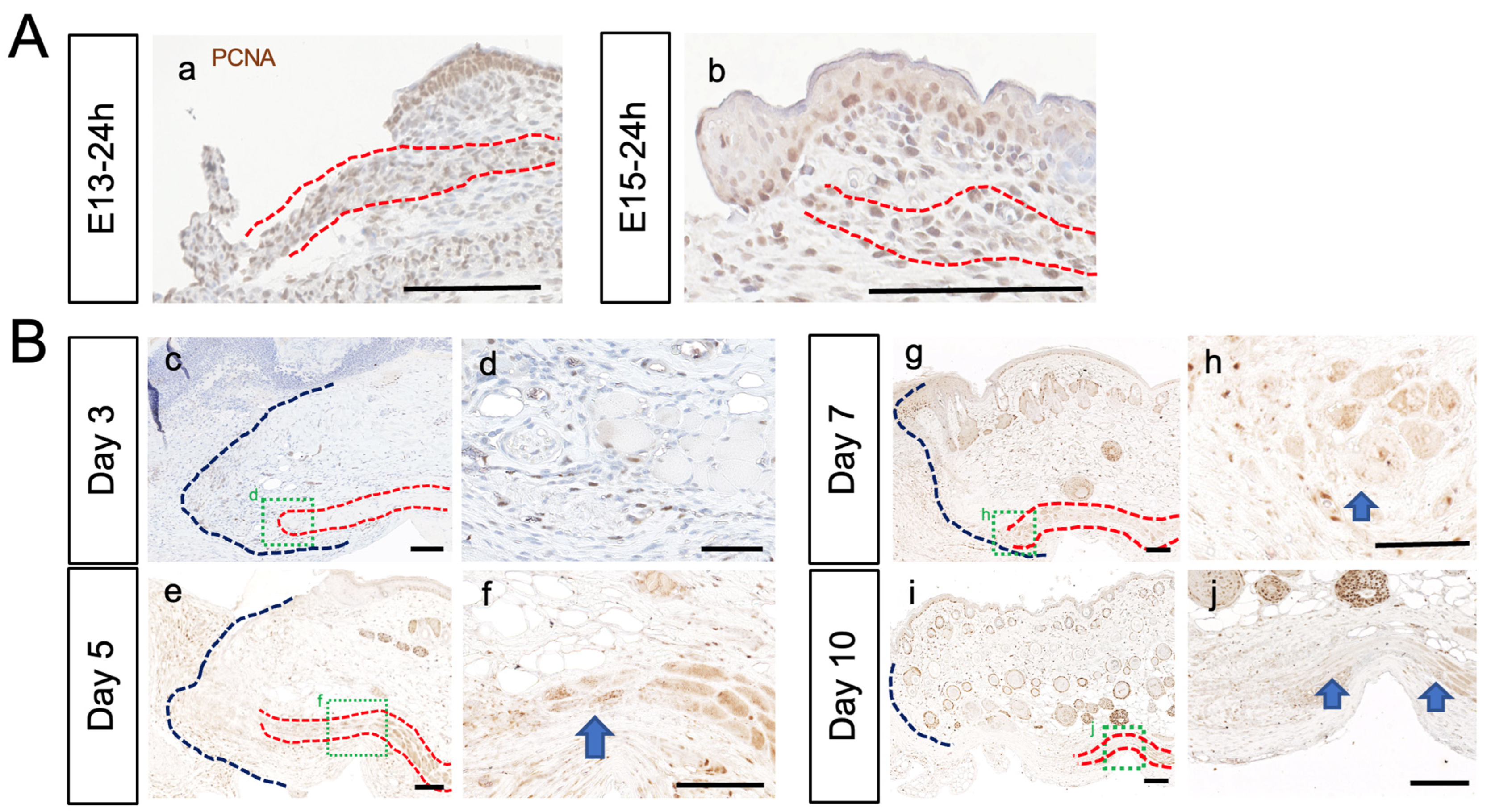
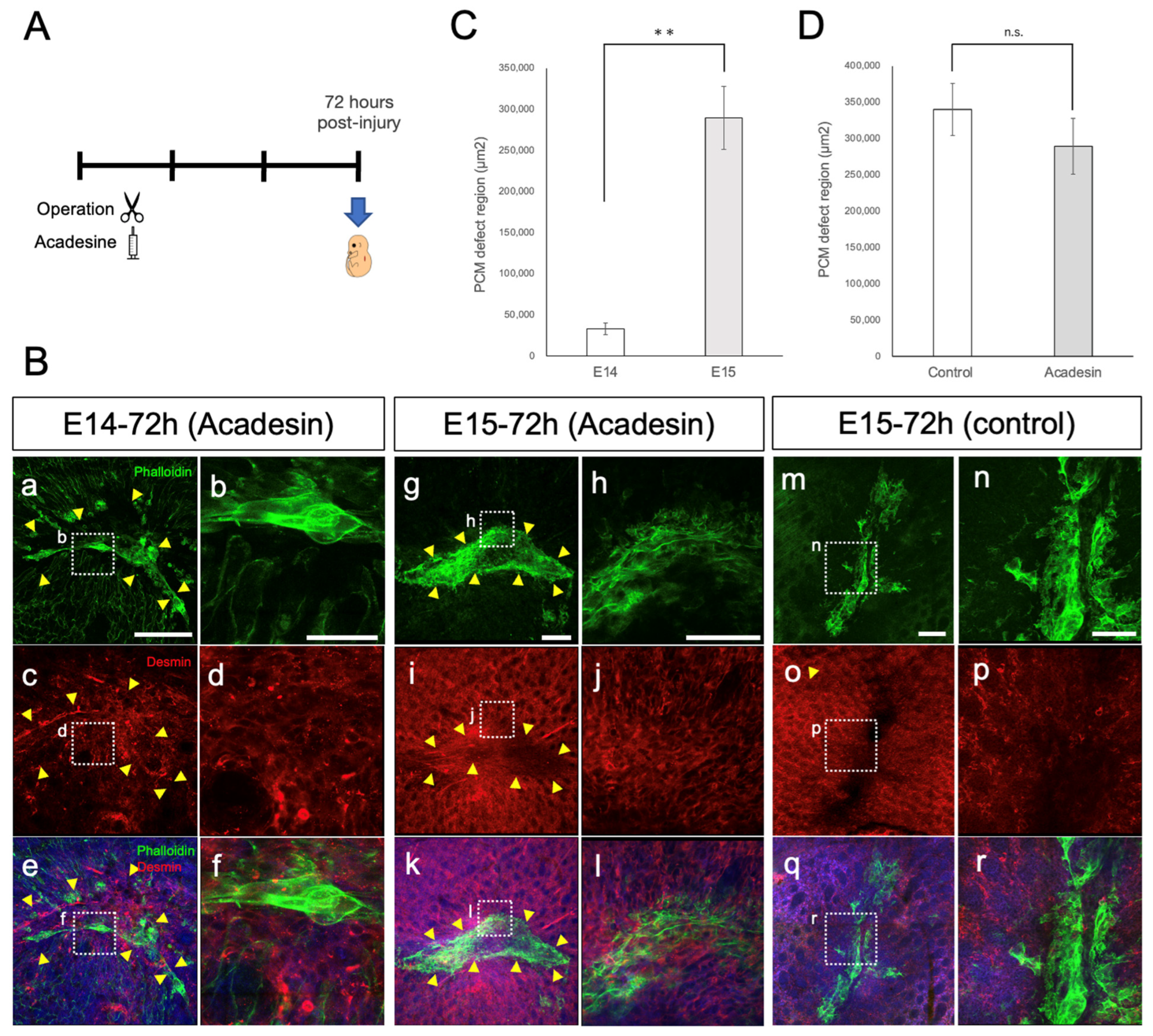
3.2. PCM Regenerates at E13, but Not after E14
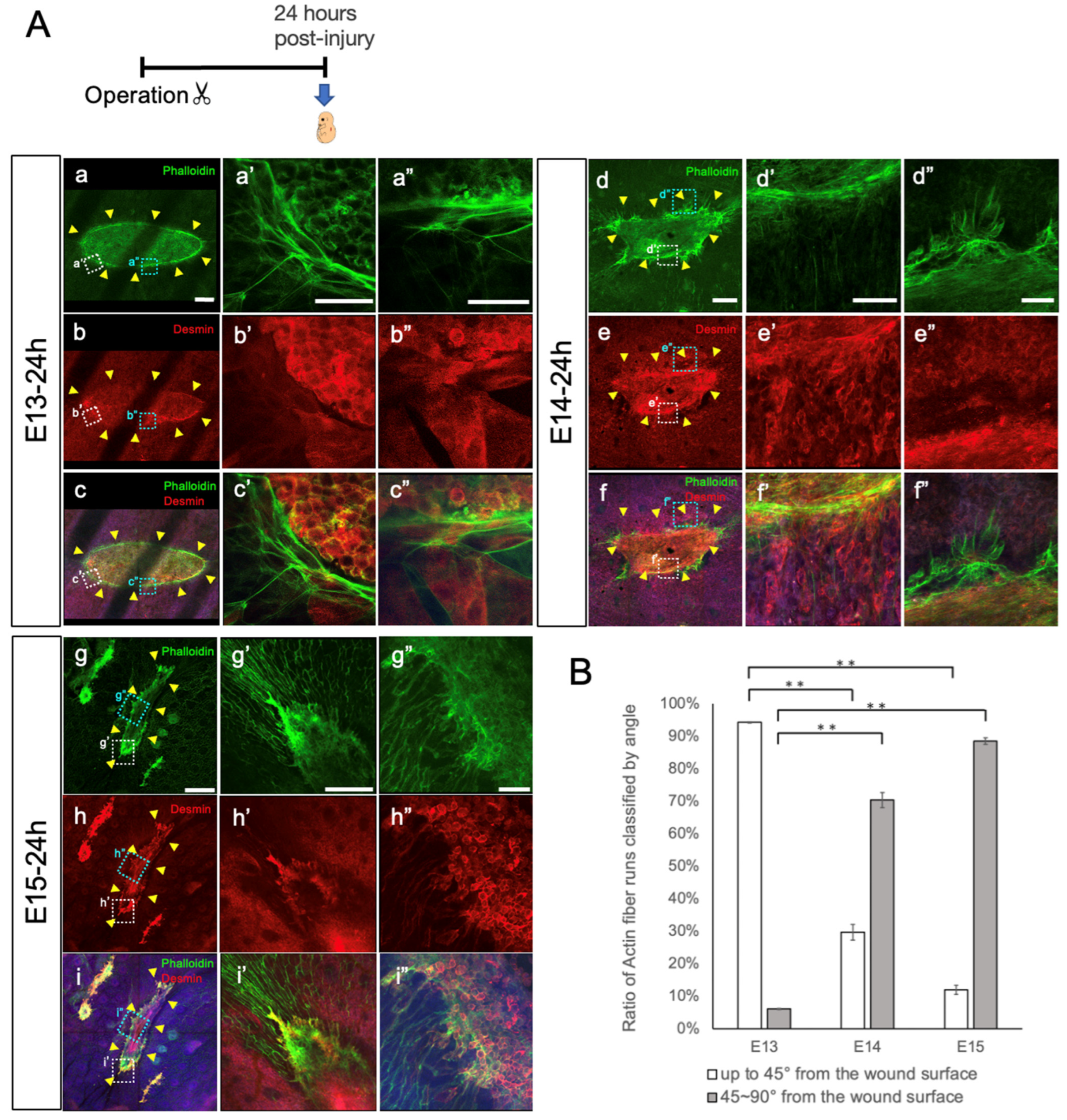
3.3. PCM Closure via Actin Cables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yokoyama, H.; Maruoka, T.; Aruga, A.; Amano, T.; Ohgo, S.; Shiroishi, T.; Tamura, K. Prx-1 expression in Xenopus laevis scarless skin-wound healing and its resemblance to epimorphic regeneration. J. Investig. Dermatol. 2011, 131, 2477–2485. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Takashimizu, I.; Casco-Robles, M.M.; Taya, Y.; Yuzuriha, S.; Toyama, F.; Maruo, F.; Kishi, K.; Chiba, C. Skin wound healing of the adult newt, Cynops pyrrhogaster: A unique re-epithelialization and scarless model. Biomedicines 2021, 9, 1892. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Sacri, R.F.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct fibroblast lineages determine dermal architecture in skin development and repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Takaya, K.; Sunohara, A.; Aramaki-Hattori, N.; Sakai, S.; Okabe, K.; Kanazawa, H.; Asou, T.; Kishi, K. Role of Wnt signaling in mouse fetal skin wound healing. Biomedicines 2022, 10, 1536. [Google Scholar] [CrossRef] [PubMed]
- Hess, A. Reactions of mammalian fetal tissues to injury. II. Skin. Anat. Rec. 1954, 119, 435–447. [Google Scholar] [CrossRef]
- Whitby, D.J.; Longaker, M.T.; Harrison, M.R.; Adzick, N.S.; Ferguson, M.W. Rapid epithelialisation of fetal wounds is associated with the early deposition of tenascin. J. Cell Sci. 1991, 99, 583–586. [Google Scholar] [CrossRef]
- Longaker, M.T.; Adzick, N.S. The biology of fetal wound healing: A review. Plast. Reconstr. Surg. 1991, 87, 788–798. [Google Scholar] [CrossRef]
- Cass, D.L.; Bullard, K.M.; Sylvester, K.G.; Yang, E.Y.; Longaker, M.T.; Adzick, N.S. Wound size and gestational age modulate scar formation in fetal wound repair. J. Pediatr. Surg. 1997, 32, 411–415. [Google Scholar] [CrossRef]
- Takaya, K.; Okabe, K.; Ishigami, A.; Imbe, Y.; Kanazawa, H.; Sakai, S.; Aramaki-Hattori, N.; Kishi, K. Actin cable formation and epidermis–dermis positional relationship during complete skin regeneration. Sci. Rep. 2022, 12, 15913. [Google Scholar] [CrossRef]
- Takaya, K.; Aramaki-Hattori, N.; Sakai, S.; Okabe, K.; Asou, T.; Kishi, K. Effect of all-trans retinoic acid on panniculus carnosus muscle regeneration in fetal mouse wound healing. Plast. Reconstr. Surg. Glob. Open. 2022, 10, e4533. [Google Scholar] [CrossRef]
- Takaya, K.; Aramaki-Hattori, N.; Sakai, S.; Okabe, K.; Asou, T.; Kishi, K. Fibroblast growth factor 7 suppresses fibrosis and promotes epithelialization during wound healing in mouse fetuses. Int. J. Mol. Sci. 2022, 23, 7087. [Google Scholar] [CrossRef] [PubMed]
- Spyrou, G.E.; Watt, D.A.; Naylor, I.L. The origin and mode of fibroblast migration and proliferation in granulation tissue. Br. J. Plast. Surg. 1998, 51, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowicz, K.; Gledhill, K.; Ambler, C.A.; Manning, C.B.; Jahoda, C.A. Development of the mouse dermal adipose layer occurs independently of subcutaneous adipose tissue and is marked by restricted early expression of FABP4. PLoS ONE 2013, 8, e59811. [Google Scholar] [CrossRef]
- Jiang, D.; Rinkevich, Y. Furnishing wound repair by the subcutaneous fascia. Int. J. Mol. Sci. 2021, 22, 9006. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Imanishi, N.; Minabe, T.; Kishi, K.; Aiso, S. Anatomical study of subcutaneous adipofascial tissue: A concept of the protective adipofascial system (PAFS) and lubricant adipofascial system (LAFS). Scand. J. Plast. Reconstr. Surg. Hand. Surg. 2004, 38, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Naldaiz-Gastesi, N.; Bahri, O.A.; López de Munain, A.; McCullagh, K.J.A.; Izeta, A. The panniculus carnosus muscle: An evolutionary enigma at the intersection of distinct research fields. J. Anat. 2018, 233, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Zammit, P.S. Satellite cells are essential for skeletal muscle regeneration: The cell on the edge returns centre stage. Development 2012, 139, 2845–2856. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Rudnicki, M.A. A new look at the origin, function, and ‘stem-cell’ status of muscle satellite cells. Dev. Biol. 2000, 218, 115–124. [Google Scholar] [CrossRef]
- Hawke, T.J.; Garry, D.J. Myogenic satellite cells: Physiology to molecular biology. J. Appl. Physiol. (1985) 2001, 91, 534–551. [Google Scholar] [CrossRef]
- Schultz, E.; McCormick, K.M. Skeletal muscle satellite cells. Rev. Physiol. Biochem. Pharmacol. 1994, 123, 213–257. [Google Scholar] [CrossRef]
- Hernández-Hernández, J.M.; García-González, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zammit, P.S.; Golding, J.P.; Nagata, Y.; Hudon, V.; Partridge, T.A.; Beauchamp, J.R. Muscle satellite cells adopt divergent fates: A mechanism for self-renewal? J. Cell Biol. 2004, 166, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Kokabu, S.; Nakatomi, C.; Matsubara, T.; Ono, Y.; Addison, W.N.; Lowery, J.W.; Urata, M.; Hudnall, A.M.; Hitomi, S.; Nakatomi, M.; et al. The transcriptional co-repressor TLE3 regulates myogenic differentiation by repressing the activity of the MyoD transcription factor. J. Biol. Chem. 2017, 292, 12885–12894. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Schüler, S.C.; Hüttner, S.S.; von Eyss, B.; von Maltzahn, J. Adult stem cells at work: Regenerating skeletal muscle. Cell. Mol. Life Sci. 2019, 76, 2559–2570. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, C.; Iantomasi, T.; Brandi, M.L. Available in vitro models for human satellite cells from skeletal muscle. Int. J. Mol. Sci. 2021, 22, 13221. [Google Scholar] [CrossRef]
- Soleimani, V.D.; Punch, V.G.; Kawabe, Y.; Jones, A.E.; Palidwor, G.A.; Porter, C.J.; Cross, J.W.; Carvajal, J.J.; Kockx, C.E.; van Ijcken, W.F.; et al. Transcriptional dominance of Pax7 in adult myogenesis is due to high-affinity recognition of homeodomain motifs. Dev. Cell 2012, 22, 1208–1220. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Dilworth, F.J. Differential modulation of cell cycle progression distinguishes members of the myogenic regulatory factor family of transcription factors. FEBS J. 2013, 280, 3991–4003. [Google Scholar] [CrossRef]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Guillamat-Prats, R. The role of MSC in wound healing, scarring and regeneration. Cells 2021, 10, 1729. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Skin fibrosis. Identification and isolation of a dermal lineage with intrinsic fibrogenic potential: Science 2015, 348, aaa2151. [Google Scholar] [CrossRef]
- Jiang, D.; Christ, S.; Correa-Gallegos, D.; Ramesh, P.; Kalgudde Gopal, S.; Wannemacher, J.; Mayr, C.H.; Lupperger, V.; Yu, Q.; Ye, H.; et al. Injury triggers fascia fibroblast collective cell migration to drive scar formation through N-cadherin. Nat. Commun. 2020, 11, 5653. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Lewis, J. Actin cables and epidermal movement in embryonic wound healing. Nature 1992, 360, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Danjo, Y.; Gipson, I.K. Actin ‘purse string’ filaments are anchored by E-cadherin-mediated adherens junctions at the leading edge of the epithelial wound, providing coordinated cell movement. J. Cell Sci. 1998, 111, 3323–3332. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y.; Coulson-Gilmer, C.; Millard, T.H. Endocytosis-dependent coordination of multiple actin regulators is required for wound healing. J. Cell Biol. 2015, 210, 419–433. [Google Scholar] [CrossRef]
- Abreu-Blanco, M.T.; Watts, J.J.; Verboon, J.M.; Parkhurst, S.M. Cytoskeleton responses in wound repair. Cell. Mol. Life Sci. 2012, 69, 2469–2483. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamada, M.; Takaya, K.; Wang, Q.; Otaki, M.; Imbe, Y.; Nakajima, Y.; Sakai, S.; Okabe, K.; Aramaki-Hattori, N.; Kishi, K. Regeneration of Panniculus Carnosus Muscle in Fetal Mice Is Characterized by the Presence of Actin Cables. Biomedicines 2023, 11, 3350. https://doi.org/10.3390/biomedicines11123350
Hamada M, Takaya K, Wang Q, Otaki M, Imbe Y, Nakajima Y, Sakai S, Okabe K, Aramaki-Hattori N, Kishi K. Regeneration of Panniculus Carnosus Muscle in Fetal Mice Is Characterized by the Presence of Actin Cables. Biomedicines. 2023; 11(12):3350. https://doi.org/10.3390/biomedicines11123350
Chicago/Turabian StyleHamada, Mariko, Kento Takaya, Qi Wang, Marika Otaki, Yuka Imbe, Yukari Nakajima, Shigeki Sakai, Keisuke Okabe, Noriko Aramaki-Hattori, and Kazuo Kishi. 2023. "Regeneration of Panniculus Carnosus Muscle in Fetal Mice Is Characterized by the Presence of Actin Cables" Biomedicines 11, no. 12: 3350. https://doi.org/10.3390/biomedicines11123350
APA StyleHamada, M., Takaya, K., Wang, Q., Otaki, M., Imbe, Y., Nakajima, Y., Sakai, S., Okabe, K., Aramaki-Hattori, N., & Kishi, K. (2023). Regeneration of Panniculus Carnosus Muscle in Fetal Mice Is Characterized by the Presence of Actin Cables. Biomedicines, 11(12), 3350. https://doi.org/10.3390/biomedicines11123350