
Autism Spectrum Disorders: A Recent Update on Targeting Inflammatory Pathways with Natural Anti-Inflammatory Agents


 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Pathophysiology of ASD
2.1. Genetics
2.2. Epigenetic
2.3. Environmental Factors
2.3.1. Heavy Metal
2.3.2. Organophosphate
2.4. Congenital Viral Infection
2.5. Other Factors
3. Current Therapies for ASD
3.1. Medications
3.2. Behavior and Communication Therapy
4. Inflammatory Pathways and Immunoinflammatory Link of ASD
4.1. NF-κB Pathway
4.2. JAK/STAT Pathway
4.3. MAPK Pathway
4.4. TGF-β Signalling Pathway
5. Significance of Natural Anti-Inflammatory Agents in ASD
6. Preclinical Studies Targeting Inflammatory Pathways
7. Clinical Studies Aimed at Targeting Inflammatory Pathways
7.1. BDNF-TrkB Pathway
7.2. JAK-STAT Pathway
7.3. mTOR Pathway
7.4. NF-κB Pathway
7.5. Toll-Like Receptor Pathway
7.6. Mitogen-Activated Protein Kinase Pathway
8. Translational Studies Targeting Inflammatory Pathways (Patents)

{kind=link}
{kind=link}
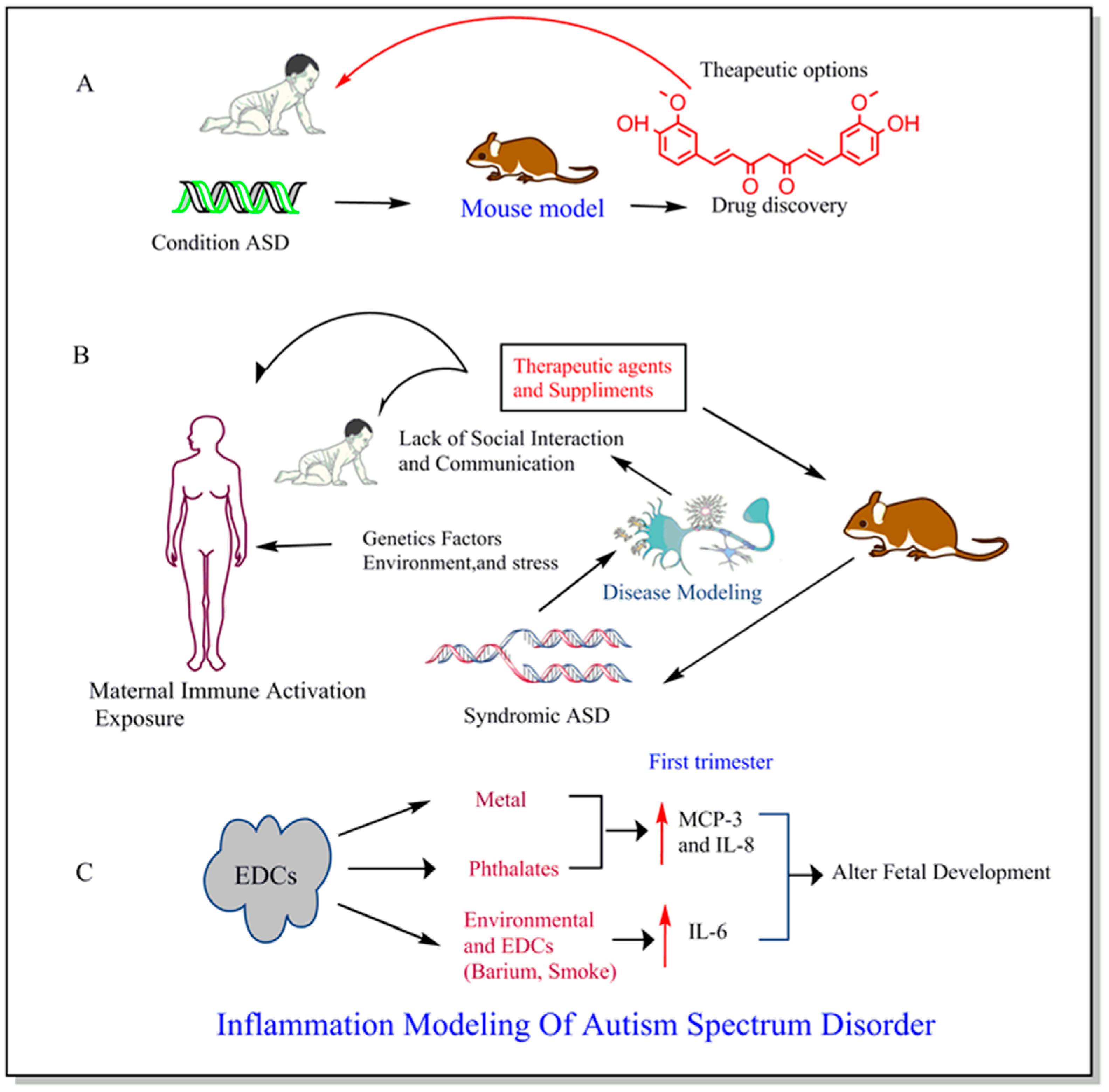{kind=link}
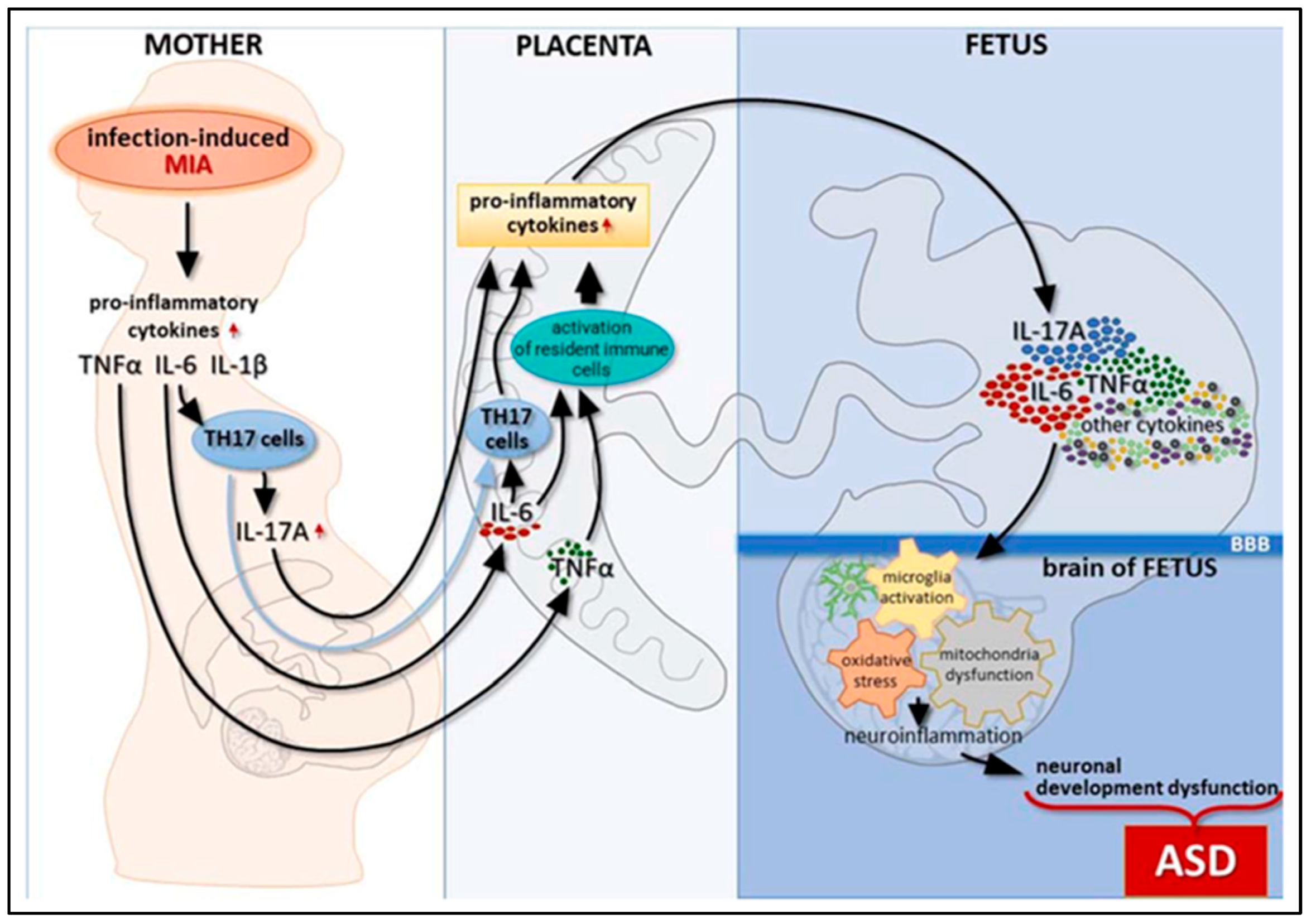{kind=link}
| S. No. | Patent No. | Natural Anti-Inflammatory Agent | Target | Method of Treatment | Reference |
|---|---|---|---|---|---|
| 1. | US8937050B2 |
| Cellular stress response | Improvements in behavioral symptoms comprise one or more of the following: a decrease in
| [101] |
| 2. | US20110104100A1 |
| IL-10, IL-4, or TGF family members | Method of treating a pervasive developmental disorder comprising:
| [102] |
9. Future Perspectives
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pizzarelli, R.; Cherubini, E. Alterations of GABAergic Signaling in Autism Spectrum Disorders. Neural Plast. 2011, 2011, 297153. [Google Scholar] [CrossRef]
- Frye, R.E. Social Skills Deficits in Autism Spectrum Disorder: Potential Biological Origins and Progress in Developing Therapeutic Agents. CNS Drugs 2018, 32, 713–734. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, U.; Quinn, T.; LaRosa, D.A.; Dickinson, H.; Walker, D.W. Prenatal Exposure to the Viral Mimetic Poly I:C Alters Fetal Brain Cytokine Expression and Postnatal Behaviour. Dev. Neurosci. 2014, 36, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Pardo, C.A.; Vargas, D.L.; Zimmerman, A.W. Immunity, Neuroglia and Neuroinflammation in Autism. Int. Rev. Psychiatry 2005, 17, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Rylaarsdam, L.; Guemez-Gamboa, A. Genetic Causes and Modifiers of Autism Spectrum Disorder. Front. Cell. Neurosci. 2019, 13, 385. [Google Scholar] [CrossRef]
- Chauhan, A.; Sahu, J.K.; Jaiswal, N.; Kumar, K.; Agarwal, A.; Kaur, J.; Singh, S.; Singh, M. Prevalence of Autism Spectrum Disorder in Indian Children: A Systematic Review and Meta-Analysis. Neurol. India 2019, 67, 100–104. [Google Scholar] [CrossRef]
- Freitas, B.C.; Mei, A.; Mendes, A.P.D.; Beltrão-Braga, P.C.B.; Marchetto, M.C. Modeling Inflammation in Autism Spectrum Disorders Using Stem Cells. Front. Pediatr. 2018, 6, 394. [Google Scholar] [CrossRef]
- Maenner, M.J. Prevalence of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2016. MMWR Surveill. Summ. 2020, 69, 1–12. [Google Scholar] [CrossRef]
- Noriega, D.B.; Savelkoul, H.F.J. Immune Dysregulation in Autism Spectrum Disorder. Eur. J. Pediatr. 2014, 173, 33–43. [Google Scholar] [CrossRef]
- Sociodemographic and Clinical Profile of Children with Autism Spectrum Disorders–An Observational Study from a Tertiary Care Hospital Bhat BA, Hussain A, Qadir W, Dar SA–BLDE Univ J Health Sci. Available online: https://www.bldeujournalhs.in/article.asp?issn=2468-838X;year=2019;volume=4;issue=2;spage=72;epage=77;aulast=Bhat (accessed on 4 August 2022).
- Marchezan, J.; Winkler Dos Santos, E.G.A.; Deckmann, I.; Riesgo, R.D.S. Immunological Dysfunction in Autism Spectrum Disorder: A Potential Target for Therapy. Neuroimmunomodulation 2018, 25, 300–319. [Google Scholar] [CrossRef]
- Warren, R.P.; Cole, P.; Odell, J.D.; Pingree, C.B.; Warren, W.L.; White, E.; Yonk, J.; Singh, V.K. Detection of Maternal Antibodies in Infantile Autism. J. Am. Acad. Child Adolesc. Psychiatry 1990, 29, 873–877. [Google Scholar] [CrossRef]
- Kurtys, E.A.; Dierckx, R.A.J.O.; Eisel, U.L.M. The Impact of Nutrition on Neuroinflammation in Vitro and in Vivo. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2017. [Google Scholar]
- Elias, R.; Sullivan, J.B.; Lee, Y.W.; White, S.W. Exploring the Potential Role of Inflammation as an Etiological Process in ASD. Rev. J. Autism. Dev. Disord. 2015, 2, 273–286. [Google Scholar] [CrossRef][Green Version]
- Samsam, M.; Ahangari, R.; Naser, S.A. Pathophysiology of Autism Spectrum Disorders: Revisiting Gastrointestinal Involvement and Immune Imbalance. World J. Gastroenterol. 2014, 20, 9942–9951. [Google Scholar] [CrossRef]
- Yalcin, O.; Sener, S.; Gokce Saripinar, E.; Soysal Acar, A.; Guney, E.; Akin Sari, B.; İşerï, E. Comparison between Cognitive Functions of Children and Adolescents with Obsessive-Compulsive Disorder and Healthy Controls: A Neuropsychological Study of Large Sample. Noropsikiyatri Ars. Arch. Neuropsychiatr. 2012, 49, 119–129. [Google Scholar] [CrossRef]
- Chaste, P.; Leboyer, M. Autism Risk Factors: Genes, Environment, and Gene-Environment Interactions. Dialogues Clin. Neurosci. 2012, 14, 281–292. [Google Scholar] [CrossRef]
- Shuid, A.N.; Jayusman, P.A.; Shuid, N.; Ismail, J.; Kamal Nor, N.; Mohamed, I.N. Association between Viral Infections and Risk of Autistic Disorder: An Overview. Int. J. Environ. Res. Public Health 2021, 18, 2817. [Google Scholar] [CrossRef]
- Robinson, E.B.; Lichtenstein, P.; Anckarsäter, H.; Happé, F.; Ronald, A. Examining and Interpreting the Female Protective Effect against Autistic Behavior. Proc. Natl. Acad. Sci. USA 2013, 110, 5258–5262. [Google Scholar] [CrossRef]
- Wiśniowiecka-Kowalnik, B.; Nowakowska, B.A. Genetics and Epigenetics of Autism Spectrum Disorder-Current Evidence in the Field. J. Appl. Genet. 2019, 60, 37–47. [Google Scholar] [CrossRef] [PubMed]
- LaSalle, J.M. Epigenomic Strategies at the Interface of Genetic and Environmental Risk Factors for Autism. J. Hum. Genet. 2013, 58, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Tognon, M.; Martini, F. Epigenetic Dysregulations in Merkel Cell Polyomavirus-Driven Merkel Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 11464. [Google Scholar] [CrossRef] [PubMed]
- Loke, Y.J.; Hannan, A.J.; Craig, J.M. The Role of Epigenetic Change in Autism Spectrum Disorders. Front. Neurol. 2015, 6, 107. [Google Scholar] [CrossRef] [PubMed]
- Schepici, G.; Cavalli, E.; Bramanti, P.; Mazzon, E. Autism Spectrum Disorder and MiRNA: An Overview of Experimental Models. Brain Sci. 2019, 9, 265. [Google Scholar] [CrossRef] [PubMed]
- Grabrucker, A.M. Environmental Factors in Autism. Front. Psychiatry 2012, 3, 118. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, V.; Singh, A.V.; Maharjan, R.S.; Dakua, S.P.; Balakrishnan, S.; Dash, S.; Laux, P.; Luch, A.; Singh, S.; Pradhan, M. Perspectives on the Technological Aspects and Biomedical Applications of Virus-Like Particles/Nanoparticles in Reproductive Biology: Insights on the Medicinal and Toxicological Outlook. Adv. NanoBiomed. Res. 2022, 2, 2200010. [Google Scholar] [CrossRef]
- Persico, A.M.; Merelli, S. Environmental Factors and Autism Spectrum Disorder. Autism Spectr. Disord. 2015, 180, 113–134. [Google Scholar] [CrossRef]
- Calviello, G.; Su, H.-M.; Weylandt, K.H.; Fasano, E.; Serini, S.; Cittadini, A. Experimental Evidence of ω-3 Polyunsaturated Fatty Acid Modulation of Inflammatory Cytokines and Bioactive Lipid Mediators: Their Potential Role in Inflammatory, Neurodegenerative, and Neoplastic Diseases. Biomed. Res. Int. 2013, 2013, 743171. [Google Scholar] [CrossRef]
- Katsinelos, T.; Tuck, B.J.; Mukadam, A.S.; McEwan, W.A. The Role of Antibodies and Their Receptors in Protection Against Ordered Protein Assembly in Neurodegeneration. Front. Immunol. 2019, 10, 1139. [Google Scholar] [CrossRef]
- Frazier, T.W.; Thompson, L.; Youngstrom, E.A.; Law, P.; Hardan, A.Y.; Eng, C.; Morris, N. A Twin Study of Heritable and Shared Environmental Contributions to Autism. J. Autism. Dev. Disord. 2014, 44, 2013–2025. [Google Scholar] [CrossRef]
- Psychiatric Rehabilitation–3rd Edition. Available online: https://www.elsevier.com/books/psychiatric-rehabilitation/barrett/978-0-12-387002-5 (accessed on 15 December 2022).
- Down Syndrome and Autistic Spectrum Disorder: A Look at What We Know. Available online: https://www.kennedykrieger.org/stories/down-syndrome-and-autistic-spectrum-disorder-look-what-we-know (accessed on 15 October 2022).
- Salcedo-Arellano, M.J.; Cabal-Herrera, A.M.; Punatar, R.H.; Clark, C.J.; Romney, C.A.; Hagerman, R.J. Overlapping Molecular Pathways Leading to Autism Spectrum Disorders, Fragile X Syndrome, and Targeted Treatments. Neurotherapeutics 2021, 18, 265–283. [Google Scholar] [CrossRef]
- Laelago, T. Herbal Medicine Use during Pregnancy: Benefits and Untoward Effects; IntechOpen: London, UK, 2018; ISBN 978-1-78984-783-3. [Google Scholar]
- Hyman, S.L.; Levy, S.E.; Myers, S.M. Council on children with disabilities, section on developmental and behavioral pediatrics Identification, Evaluation, and Management of Children With Autism Spectrum Disorder. Pediatrics 2020, 145, e20193447. [Google Scholar] [CrossRef]
- Hafizi, S.; Tabatabaei, D.; Lai, M.-C. Review of Clinical Studies Targeting Inflammatory Pathways for Individuals With Autism. Front. Psychiatry 2019, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Zawadzka, A.; Cieślik, M.; Adamczyk, A. The Role of Maternal Immune Activation in the Pathogenesis of Autism: A Review of the Evidence, Proposed Mechanisms and Implications for Treatment. Int. J. Mol. Sci. 2021, 22, 11516. [Google Scholar] [CrossRef]
- Morrison, D.K. MAP Kinase Pathways. Cold. Spring Harb. Perspect. Biol. 2012, 4, a011254. [Google Scholar] [CrossRef]
- Liao, X.; Li, Y. Nuclear Factor Kappa B in Autism Spectrum Disorder: A Systematic Review. Pharmacol. Res. 2020, 159, 104918. [Google Scholar] [CrossRef]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The Role of JAK/STAT Signaling Pathway and Its Inhibitors in Diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Nadeem, A.; Ansari, M.A.; Bakheet, S.A.; Al-Ayadhi, L.Y.; Attia, S.M. Upregulation of IL-9 and JAK-STAT Signaling Pathway in Children with Autism. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The Maternal Interleukin-17a Pathway in Mice Promotes Autism-like Phenotypes in Offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Sometani, A.; Kataoka, H.; Nitta, A.; Fukumitsu, H.; Nomoto, H.; Furukawa, S. Transforming Growth Factor-Beta1 Enhances Expression of Brain-Derived Neurotrophic Factor and Its Receptor, TrkB, in Neurons Cultured from Rat Cerebral Cortex. J. Neurosci. Res. 2001, 66, 369–376. [Google Scholar] [CrossRef]
- Ohja, K.; Gozal, E.; Fahnestock, M.; Cai, L.; Cai, J.; Freedman, J.H.; Switala, A.; El-Baz, A.; Barnes, G.N. Neuroimmunologic and Neurotrophic Interactions in Autism Spectrum Disorders: Relationship to Neuroinflammation. Neuromol. Med. 2018, 20, 161–173. [Google Scholar] [CrossRef]
- Kutuk, M.O.; Tufan, E.; Gokcen, C.; Kilicaslan, F.; Karadag, M.; Mutluer, T.; Yektas, C.; Coban, N.; Kandemir, H.; Buber, A.; et al. Cytokine Expression Profiles in Autism Spectrum Disorder: A Multi-Center Study from Turkey. Cytokine 2020, 133, 155152. [Google Scholar] [CrossRef]
- Skaper, S.D. The Brain as a Target for Inflammatory Processes and Neuroprotective Strategies. Ann. N. Y. Acad. Sci. 2007, 1122, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, P.; Mehdi, I.; Kaith, R.; Ahmad, F.; Anwar, M.S. Potential Natural Products for the Management of Autism Spectrum Disorder. Ibrain 2022, 8, 365–376. [Google Scholar] [CrossRef]
- Tsilioni, I.; Taliou, A.; Francis, K.; Theoharides, T.C. Children with Autism Spectrum Disorders, Who Improved with a Luteolin-Containing Dietary Formulation, Show Reduced Serum Levels of TNF and IL-6. Transl. Psychiatry 2015, 5, e647. [Google Scholar] [CrossRef] [PubMed]
- Taliou, A.; Zintzaras, E.; Lykouras, L.; Francis, K. An Open-Label Pilot Study of a Formulation Containing the Anti-Inflammatory Flavonoid Luteolin and Its Effects on Behavior in Children with Autism Spectrum Disorders. Clin. Ther. 2013, 35, 592–602. [Google Scholar] [CrossRef]
- Kelley, A.S.; Banker, M.; Goodrich, J.M.; Dolinoy, D.C.; Burant, C.; Domino, S.E.; Smith, Y.R.; Song, P.X.K.; Padmanabhan, V. Early Pregnancy Exposure to Endocrine Disrupting Chemical Mixtures Are Associated with Inflammatory Changes in Maternal and Neonatal Circulation. Sci. Rep. 2019, 9, 5422. [Google Scholar] [CrossRef]
- Mabunga, D.F.N.; Gonzales, E.L.T.; Kim, J.-W.; Kim, K.C.; Shin, C.Y. Exploring the Validity of Valproic Acid Animal Model of Autism. Exp. Neurobiol. 2015, 24, 285–300. [Google Scholar] [CrossRef]
- Bertolino, B.; Crupi, R.; Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Beneficial Effects of Co-Ultramicronized Palmitoylethanolamide/Luteolin in a Mouse Model of Autism and in a Case Report of Autism. CNS Neurosci. Ther. 2017, 23, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Parker-Athill, E.; Luo, D.; Bailey, A.; Giunta, B.; Tian, J.; Shytle, R.D.; Murphy, T.; Legradi, G.; Tan, J. Flavonoids, a Prenatal Prophylaxis via Targeting JAK2/STAT3 Signaling to Oppose IL-6/MIA Associated Autism. J. Neuroimmunol. 2009, 217, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Kopschina Feltes, P.; Doorduin, J.; Klein, H.C.; Juárez-Orozco, L.E.; Dierckx, R.A.; Moriguchi-Jeckel, C.M.; de Vries, E.F. Anti-Inflammatory Treatment for Major Depressive Disorder: Implications for Patients with an Elevated Immune Profile and Non-Responders to Standard Antidepressant Therapy. J. Psychopharmacol. 2017, 31, 1149–1165. [Google Scholar] [CrossRef]
- Depino, A.M. Peripheral and Central Inflammation in Autism Spectrum Disorders. Mol. Cell. Neurosci. 2013, 53, 69–76. [Google Scholar] [CrossRef]
- Lyall, K.; Schmidt, R.J.; Hertz-Picciotto, I. Maternal Lifestyle and Environmental Risk Factors for Autism Spectrum Disorders. Int. J. Epidemiol. 2014, 43, 443–464. [Google Scholar] [CrossRef]
- Kumar, S.; Reynolds, K.; Ji, Y.; Gu, R.; Rai, S.; Zhou, C.J. Impaired Neurodevelopmental Pathways in Autism Spectrum Disorder: A Review of Signaling Mechanisms and Crosstalk. J. Neurodev. Disord. 2019, 11, 10. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Immune Mediators in the Brain and Peripheral Tissues in Autism Spectrum Disorder. Nat. Rev. Neurosci. 2015, 16, 469–486. [Google Scholar] [CrossRef]
- Cristiano, C.; Lama, A.; Lembo, F.; Mollica, M.P.; Calignano, A.; Mattace Raso, G. Interplay Between Peripheral and Central Inflammation in Autism Spectrum Disorders: Possible Nutritional and Therapeutic Strategies. Front. Physiol. 2018, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Stigler, K.A. Psychopharmacologic Management of Serious Behavioral Disturbance in ASD. Child. Adolesc. Psychiatr. Clin. N. Am. 2014, 23, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Minakova, E.; Warner, B.B. Maternal Immune Activation, Central Nervous System Development and Behavioral Phenotypes. Birth Defects Res. 2018, 110, 1539–1550. [Google Scholar] [CrossRef] [PubMed]
- Karl, T.; Pabst, R.; von Hörsten, S. Behavioral Phenotyping of Mice in Pharmacological and Toxicological Research. Exp. Toxicol. Pathol. 2003, 55, 69–83. [Google Scholar] [CrossRef]
- Iyer, S.S.; Cheng, G. Role of Interleukin 10 Transcriptional Regulation in Inflammation and Autoimmune Disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef]
- Patterson, P.H. Maternal Infection and Immune Involvement in Autism. Trends Mol. Med. 2011, 17, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Urakubo, A.; Jarskog, L.F.; Lieberman, J.A.; Gilmore, J.H. Prenatal Exposure to Maternal Infection Alters Cytokine Expression in the Placenta, Amniotic Fluid, and Fetal Brain. Schizophr. Res. 2001, 47, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Mosser, C.-A.; Baptista, S.; Arnoux, I.; Audinat, E. Microglia in CNS Development: Shaping the Brain for the Future. Prog. Neurobiol. 2017, 149–150, 1–20. [Google Scholar] [CrossRef]
- Dheen, S.T.; Kaur, C.; Ling, E.-A. Microglial Activation and Its Implications in the Brain Diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Valles, A.; Rodrigue, B.; Matta-Camacho, E. Maternal Immune Activation and the Development of Dopaminergic Neurotransmission of the Offspring: Relevance for Schizophrenia and Other Psychoses. Front. Psychiatry 2020, 11, 852. [Google Scholar] [CrossRef]
- Boulanger-Bertolus, J.; Pancaro, C.; Mashour, G.A. Increasing Role of Maternal Immune Activation in Neurodevelopmental Disorders. Front. Behav. Neurosci. 2018, 12, 230. [Google Scholar] [CrossRef]
- Ross, H.E.; Guo, Y.; Coleman, K.; Ousley, O.; Miller, A.H. Association of IL-12p70 and IL-6:IL-10 Ratio with Autism-Related Behaviors in 22q11.2 Deletion Syndrome: A Preliminary Report. Brain. Behav. Immun. 2013, 31, 76–81. [Google Scholar] [CrossRef]
- Pendyala, G.; Chou, S.; Jung, Y.; Coiro, P.; Spartz, E.; Padmashri, R.; Li, M.; Dunaevsky, A. Maternal Immune Activation Causes Behavioral Impairments and Altered Cerebellar Cytokine and Synaptic Protein Expression. Neuropsychopharmacology 2017, 42, 1435–1446. [Google Scholar] [CrossRef]
- Mostafa, G.A.; Al-Ayadhi, L.Y. The Relationship between the Increased Frequency of Serum Antineuronal Antibodies and the Severity of Autism in Children. Eur. J. Paediatr. Neurol. 2012, 16, 464–468. [Google Scholar] [CrossRef]
- Olmos, G.; Lladó, J. Tumor Necrosis Factor Alpha: A Link between Neuroinflammation and Excitotoxicity. Mediators Inflamm. 2014, 2014, 861231. [Google Scholar] [CrossRef]
- Blaylock, R.L.; Strunecka, A. Immune-Glutamatergic Dysfunction as a Central Mechanism of the Autism Spectrum Disorders. Curr. Med. Chem. 2009, 16, 157–170. [Google Scholar] [CrossRef]
- Conway, F.; Brown, A.S. Maternal Immune Activation and Related Factors in the Risk of Offspring Psychiatric Disorders. Front. Psychiatry 2019, 10, 430. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Lima, C.N.; Doifode, T.; Colodel, A.; Sayana, P.; Giridharan, V.V.; Macedo, D.S.; Pinjari, O.F.; Barichello, T. Maternal Immune Activation as a Risk Factor for Schizophrenia: Evidence From Preclinical and Clinical Studies. In Perinatal Inflammation and Adult Psychopathology: From Preclinical Models to Humans; Teixeira, A.L., Macedo, D., Baune, B.T., Eds.; Progress in Inflammation Research; Springer International Publishing: Cham, The Netherlands, 2020; pp. 129–154. ISBN 978-3-030-39335-9. [Google Scholar]
- Bölte, S.; Girdler, S.; Marschik, P.B. The Contribution of Environmental Exposure to the Etiology of Autism Spectrum Disorder. Cell. Mol. Life Sci. 2019, 76, 1275–1297. [Google Scholar] [CrossRef] [PubMed]
- Buka, S.L.; Cannon, T.D.; Torrey, E.F.; Yolken, R.H.; Collaborative Study Group on the Perinatal Origins of Severe Psychiatric Disorders. Maternal Exposure to Herpes Simplex Virus and Risk of Psychosis among Adult Offspring. Biol. Psychiatry 2008, 63, 809–815. [Google Scholar] [CrossRef]
- Wang, L.; Wang, F.-S.; Gershwin, M.E. Human Autoimmune Diseases: A Comprehensive Update. J. Intern. Med. 2015, 278, 369–395. [Google Scholar] [CrossRef]
- Chen, S.-W.; Zhong, X.-S.; Jiang, L.-N.; Zheng, X.-Y.; Xiong, Y.-Q.; Ma, S.-J.; Qiu, M.; Huo, S.-T.; Ge, J.; Chen, Q. Maternal Autoimmune Diseases and the Risk of Autism Spectrum Disorders in Offspring: A Systematic Review and Meta-Analysis. Behav. Brain Res. 2016, 296, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.G.; Sullivan, G.E. Glutathione, Vitamin C and Cysteine Use in Children with Autism and Severe Behavior Concerns: A Double Blind, Placebo-Controlled Crossover Study; University of Louisville, Louisville, KY, USA, 2016. Available online: clinicaltrials.gov (accessed on 30 November 2022).
- Jankowsky, J.L.; Patterson, P.H. Cytokine and Growth Factor Involvement in Long-Term Potentiation. Mol. Cell. Neurosci. 1999, 14, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Neuropeptides and Neurotrophins in Neonatal Blood of Children with Autism or Mental Retardation–Nelson–2001–Annals of Neurology–Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/ana.1024 (accessed on 19 December 2022).
- Miyazaki, K.; Narita, N.; Sakuta, R.; Miyahara, T.; Naruse, H.; Okado, N.; Narita, M. Serum Neurotrophin Concentrations in Autism and Mental Retardation: A Pilot Study. Brain Dev. 2004, 26, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Katoh-Semba, R.; Wakako, R.; Komori, T.; Shigemi, H.; Miyazaki, N.; Ito, H.; Kumagai, T.; Tsuzuki, M.; Shigemi, K.; Yoshida, F.; et al. Age-Related Changes in BDNF Protein Levels in Human Serum: Differences between Autism Cases and Normal Controls. Int. J. Dev. Neurosci. 2007, 25, 367–372. [Google Scholar] [CrossRef]
- Xu, N.; Li, X.; Zhong, Y. Inflammatory Cytokines: Potential Biomarkers of Immunologic Dysfunction in Autism Spectrum Disorders. Mediat. Inflamm. 2015, 2015, 531518. [Google Scholar] [CrossRef]
- Zhang, J.-C.; Yao, W.; Hashimoto, K. Brain-Derived Neurotrophic Factor (BDNF)-TrkB Signaling in Inflammation-Related Depression and Potential Therapeutic Targets. Curr. Neuropharmacol. 2016, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Baranova, J.; Dragunas, G.; Botellho, M.C.S.; Ayub, A.L.P.; Bueno-Alves, R.; Alencar, R.R.; Papaiz, D.D.; Sogayar, M.C.; Ulrich, H.; Correa, R.G. Autism Spectrum Disorder: Signaling Pathways and Prospective Therapeutic Targets. Cell. Mol. Neurobiol. 2021, 41, 619–649. [Google Scholar] [CrossRef] [PubMed]
- Sato, A. MTOR, a Potential Target to Treat Autism Spectrum Disorder. CNS Neurol. Disord. Drug Targets 2016, 15, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.; Ahmad, S.F.; Bakheet, S.A.; Al-Harbi, N.O.; Al-Ayadhi, L.Y.; Attia, S.M.; Zoheir, K.M.A. Toll-like Receptor 4 Signaling Is Associated with Upregulated NADPH Oxidase Expression in Peripheral T Cells of Children with Autism. Brain Behav. Immun. 2017, 61, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Manicassamy, S.; Pulendran, B. Modulation of Adaptive Immunity with Toll-like Receptors. Semin. Immunol. 2009, 21, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Fekonja, O.; Avbelj, M.; Jerala, R. Suppression of TLR Signaling by Targeting TIR Domain-Containing Proteins. Curr. Protein Pept. Sci. 2012, 13, 776–788. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Taniguchi, T. IRFs: Master Regulators of Signalling by Toll-like Receptors and Cytosolic Pattern-Recognition Receptors. Nat. Rev. Immunol. 2006, 6, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Al-Harbi, N.O.; Nadeem, A.; Ahmad, S.F.; Al-Ayadhi, L.Y.; Al-Harbi, M.M.; As Sobeai, H.M.; Ibrahim, K.E.; Bakheet, S.A. Elevated Expression of Toll-like Receptor 4 Is Associated with NADPH Oxidase-Induced Oxidative Stress in B Cells of Children with Autism. Int. Immunopharmacol. 2020, 84, 106555. [Google Scholar] [CrossRef]
- Sanz-Moreno, V.; Casar, B.; Crespo, P. P38alpha Isoform Mxi2 Binds to Extracellular Signal-Regulated Kinase 1 and 2 Mitogen-Activated Protein Kinase and Regulates Its Nuclear Activity by Sustaining Its Phosphorylation Levels. Mol. Cell. Biol. 2003, 23, 3079–3090. [Google Scholar] [CrossRef]
- Vithayathil, J.; Pucilowska, J.; Landreth, G.E. ERK/MAPK Signaling and Autism Spectrum Disorders. Prog. Brain Res. 2018, 241, 63–112. [Google Scholar] [CrossRef]
- Sulakhiya, K.; Keshavlal, G.P.; Bezbaruah, B.B.; Dwivedi, S.; Gurjar, S.S.; Munde, N.; Jangra, A.; Lahkar, M.; Gogoi, R. Lipopolysaccharide Induced Anxiety- and Depressive-like Behaviour in Mice Are Prevented by Chronic Pre-Treatment of Esculetin. Neurosci. Lett. 2016, 611, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Pangrazzi, L.; Balasco, L.; Bozzi, Y. Natural Antioxidants: A Novel Therapeutic Approach to Autism Spectrum Disorders? Antioxidants 2020, 9, 1186. [Google Scholar] [CrossRef] [PubMed]
- Talalay, P.; Zimmerman, A.W.; Smith, K.D. Methods and Compositions for Treatment of Autism. U.S. Patent No. 8,937,050, 20 January 2015. [Google Scholar]
- Riordan, N.H.; Ichim, T.E. Compositions and Methods of Stem Cell Therapy for Autism. Google Patents WO2009046377A2, 9 April 2009. Available online: https://patents.google.com/patent/WO2009046377A2/en (accessed on 4 August 2022).




| S. No. | Pathway | Anti-Inflammatory Agent | Target | Remark | References |
|---|---|---|---|---|---|
| 1. | NF-κB | Resveratrol | IL-1β, TNF-α IL-17A |
| [40] |
| Palmitoylethanolamide & luteoiln | IL-1b, IL-6, TNF-α, NO, NF-κB |
| [54] | ||
| 2. | JAK/STAT | Luteolin Diosmine | IL-6, IL-12, IFN-γ |
| [41,55] |
| Quercetine | IL-12 |
| [55] | ||
| 3. | ERK/MAPK | IL-17A antibody | IL-17A |
| [38,43] |
| S. No. | Molecule | Targets | Patient Detail | Type of Study | Remarks | Reference |
|---|---|---|---|---|---|---|
| 1. | Dietary Supplement: Luteolin, Quercetin & Rutin combined in a capsule (Neuroprotek) | IL-6 & 8 TNF-α | N = 50 (42M/8F) 4–10 years | Open-label trial |
| [51] |
| 2. | GSH, Vit. C, NAC | - | N = 24 | Interventional |
| [83] |
| 3. | Luteolin formulation | IL-6 & 8 TNF-α | N = 40 (34M/6F) 4–10 years | Open-label trial |
| [50] |
| 4. | Palmitoylethanolamide & luteolin | IL-6 | A 10-year-old child | ATEC test |
| [54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.; Kisku, A.; Kungumaraj, H.; Nagaraj, V.; Pal, A.; Kumar, S.; Sulakhiya, K. Autism Spectrum Disorders: A Recent Update on Targeting Inflammatory Pathways with Natural Anti-Inflammatory Agents. Biomedicines 2023, 11, 115. https://doi.org/10.3390/biomedicines11010115
Singh R, Kisku A, Kungumaraj H, Nagaraj V, Pal A, Kumar S, Sulakhiya K. Autism Spectrum Disorders: A Recent Update on Targeting Inflammatory Pathways with Natural Anti-Inflammatory Agents. Biomedicines. 2023; 11(1):115. https://doi.org/10.3390/biomedicines11010115
Chicago/Turabian StyleSingh, Ramu, Anglina Kisku, Haripriya Kungumaraj, Vini Nagaraj, Ajay Pal, Suneel Kumar, and Kunjbihari Sulakhiya. 2023. "Autism Spectrum Disorders: A Recent Update on Targeting Inflammatory Pathways with Natural Anti-Inflammatory Agents" Biomedicines 11, no. 1: 115. https://doi.org/10.3390/biomedicines11010115
APA StyleSingh, R., Kisku, A., Kungumaraj, H., Nagaraj, V., Pal, A., Kumar, S., & Sulakhiya, K. (2023). Autism Spectrum Disorders: A Recent Update on Targeting Inflammatory Pathways with Natural Anti-Inflammatory Agents. Biomedicines, 11(1), 115. https://doi.org/10.3390/biomedicines11010115