
Comparing Intraperitoneal and Intravenous Personalized ErbB2CAR-T for the Treatment of Epithelial Ovarian Cancer
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Cell Lines and Culture
2.3. Construction of the CAR Retroviral Vector
2.4. Preparation of Packaging Cells
2.5. T-Cell Transduction
2.6. Flow Cytometry Analysis
2.7. In Vitro CAR-T Cell Mediated Killing
2.8. In Vitro IFNγ Secretion Assay
2.9. Luciferase Gene Transduction of NAR Cells
2.10. In Vivo Experiments
2.11. RT-PCR
2.12. Single-Cell Isolation from Mouse Tissues and Human Ovarian Tissues
2.13. Ex-Vivo Organoid Culture (EVOC) System
2.14. Scoring of Ex Vivo Organ Culture
2.15. Statistical Analysis
3. Results
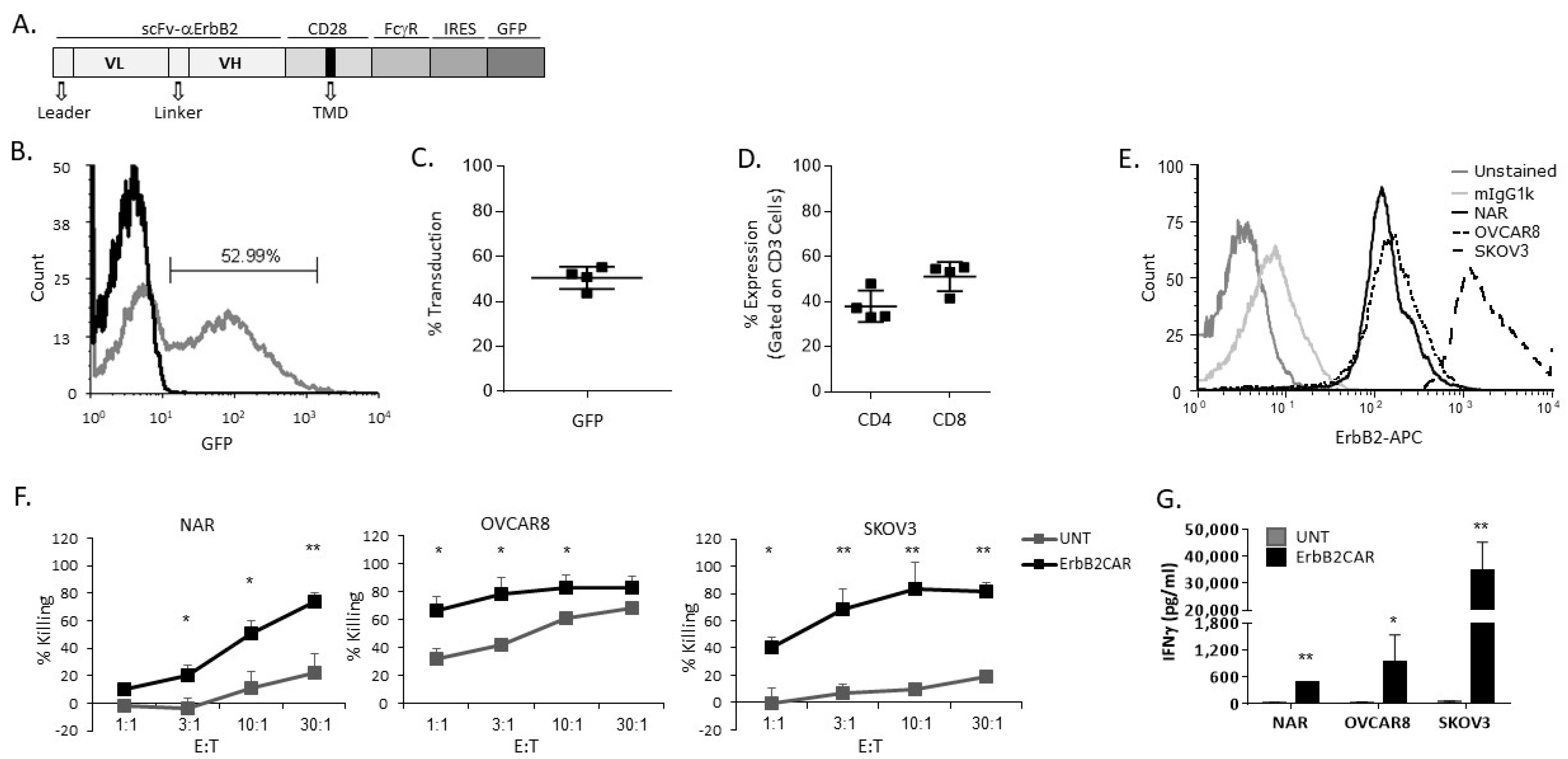
3.1. Characterization and In Vitro Activity of ErbB2CAR-T Cells
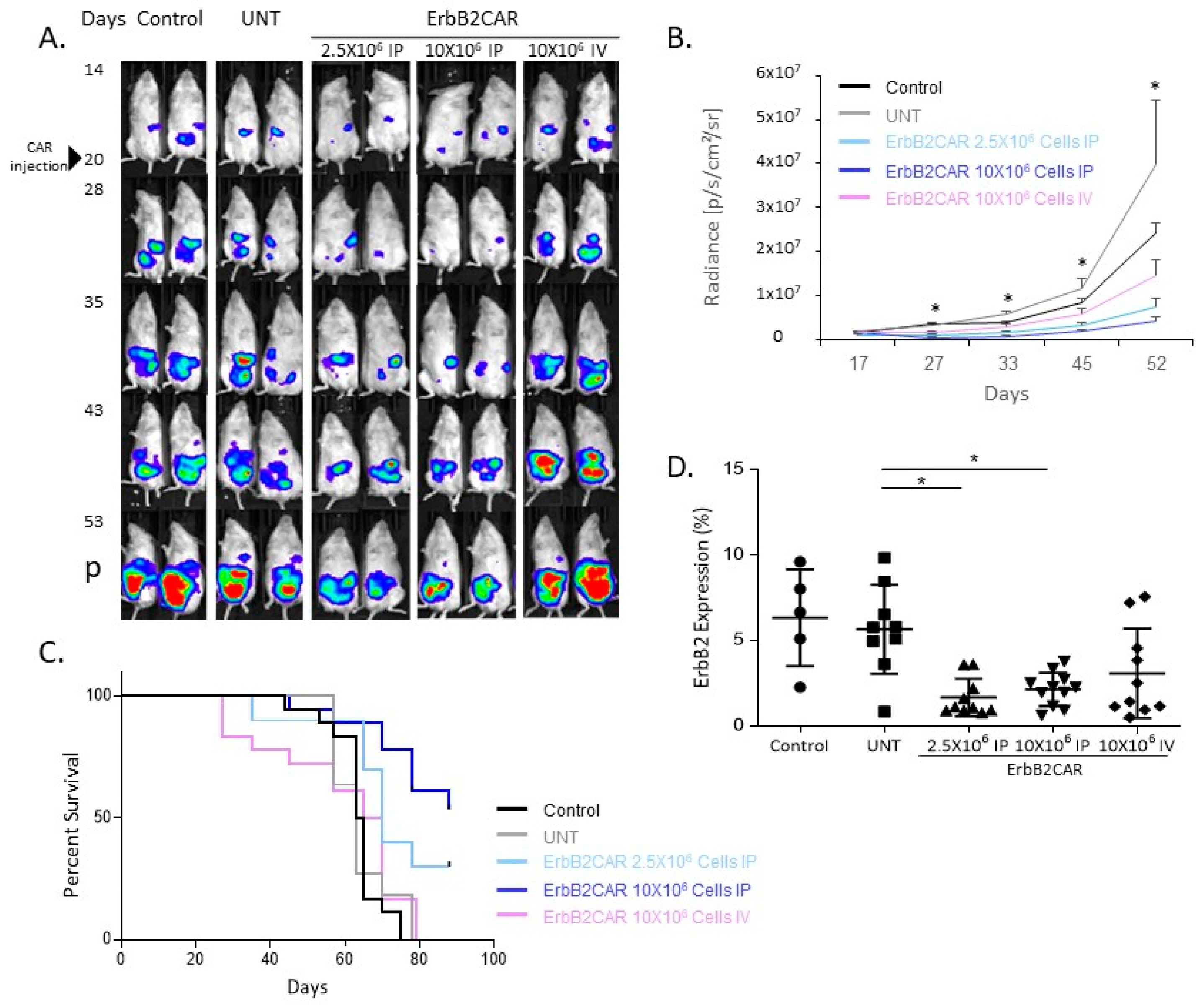
3.2. Antitumor Effect of Local and Systemic ErbB2CAR Treatment in an OC Mouse Model
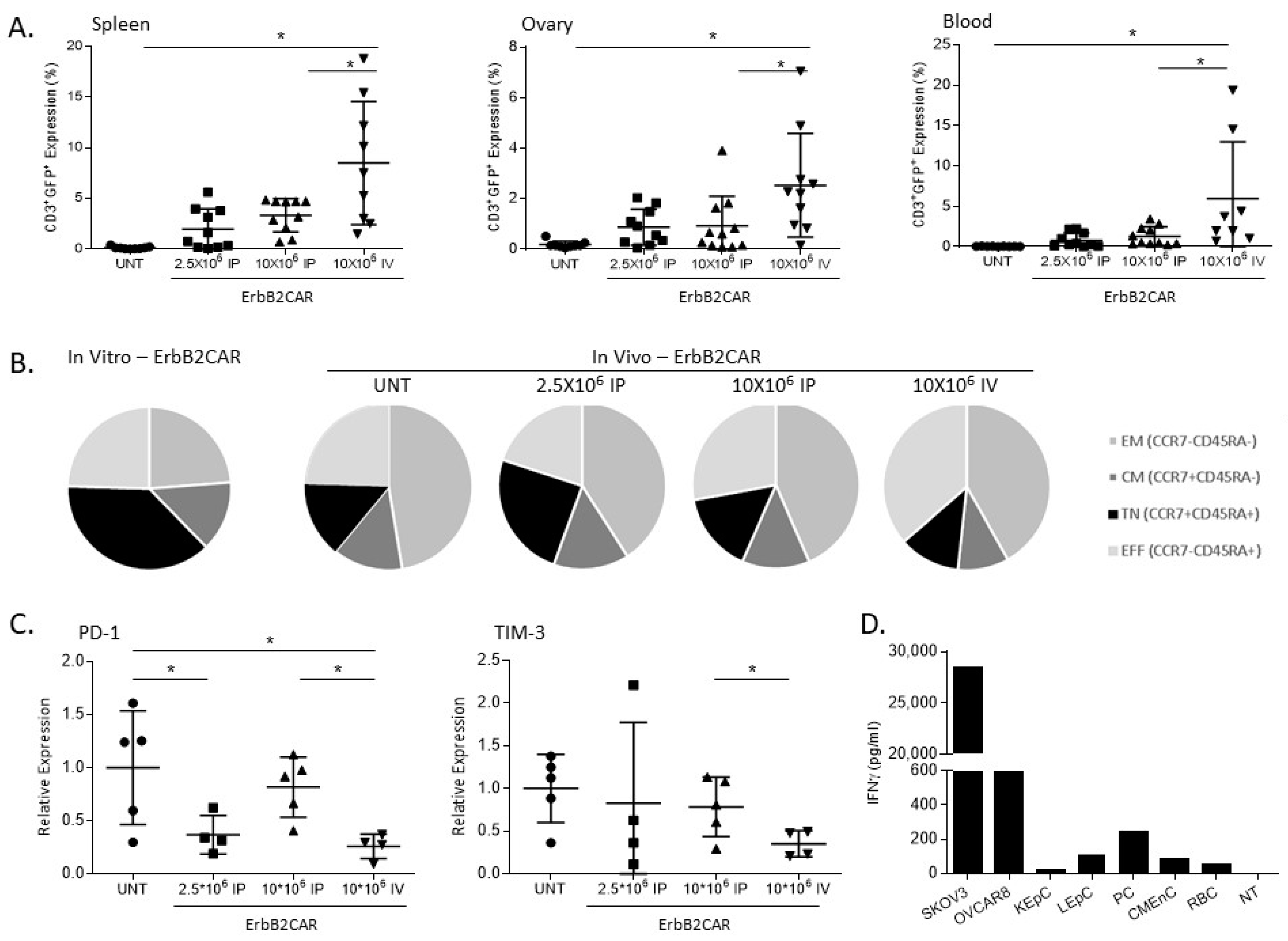
3.3. In Vivo Persistence and Safety Analysis of ErbB2CAR-T Cells
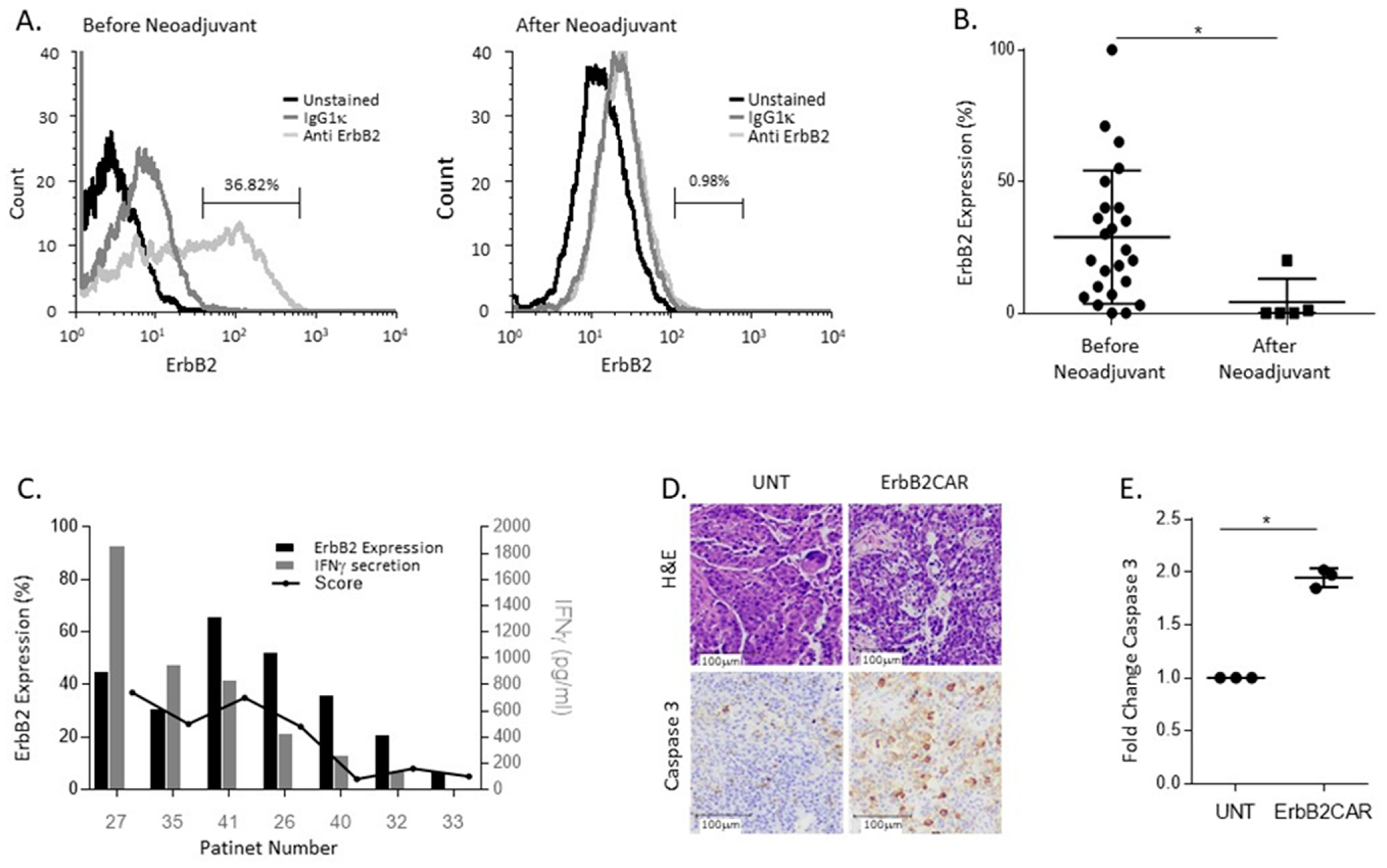
3.4. ErbB2CAR Promotes Killing of Primary Ovarian Tumors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Lheureux, S.; Braunstein, M.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [PubMed]
- Tebbutt, N.; Pedersen, M.W.; Johns, T.G. Targeting the ERBB family in cancer: Couples therapy. Nat. Rev. Cancer 2013, 13, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Teplinsky, E.; Muggia, F. Targeting HER2 in ovarian and uterine cancers: Challenges and future directions. Gynecol. Oncol. 2014, 135, 364–370. [Google Scholar] [CrossRef]
- Reibenwein, J.; Krainer, M. Targeting signaling pathways in ovarian cancer. Expert Opin. Ther. Targets 2008, 12, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Schuster, S.J.; Bishop, M.R.; Tam, C.S.; Waller, E.K.; Borchmann, P.; McGuirk, J.P.; Jager, U.; Jaglowski, S.; Andreadis, C.; Westin, J.R.; et al. Tisagenlecleucel in Adult Relapsed or Refractory Diffuse Large B-Cell Lymphoma. N. Engl. J. Med. 2019, 380, 45–56. [Google Scholar] [CrossRef]
- Chardin, L.; Leary, A. Immunotherapy in Ovarian Cancer: Thinking Beyond PD-1/PD-L1. Front. Oncol. 2021, 11, 795547. [Google Scholar] [CrossRef] [PubMed]
- Leary, A.; Tan, D.; Ledermann, J. Immune checkpoint inhibitors in ovarian cancer: Where do we stand? Ther. Adv. Med. Oncol. 2021, 13, 17588359211039899. [Google Scholar] [CrossRef]
- Maiorano, B.A.; Maiorano, M.F.P.; Lorusso, D.; Maiello, E. Ovarian Cancer in the Era of Immune Checkpoint Inhibitors: State of the Art and Future Perspectives. Cancers 2021, 13, 4438. [Google Scholar] [CrossRef]
- Gross, G.; Eshhar, Z. Endowing T cells with antibody specificity using chimeric T cell receptors. FASEB J. 1992, 6, 3370–3378. [Google Scholar] [CrossRef] [PubMed]
- Eshhar, Z.; Waks, T.; Gross, G. The emergence of T-bodies/CAR T cells. Cancer J. 2014, 20, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Globerson Levin, A.; Riviere, I.; Eshhar, Z.; Sadelain, M. CAR T cells: Building on the CD19 paradigm. Eur. J. Immunol. 2021, 51, 2151–2163. [Google Scholar] [CrossRef]
- Yan, W.; Hu, H.; Tang, B. Advances Of Chimeric Antigen Receptor T Cell Therapy In Ovarian Cancer. Oncol. Targets Ther. 2019, 12, 8015–8022. [Google Scholar] [CrossRef]
- Papa, S.; van Schalkwyk, M.; Maher, J. Clinical Evaluation of ErbB-Targeted CAR T-Cells, Following Intracavity Delivery in Patients with ErbB-Expressing Solid Tumors. Methods Mol. Biol. 2015, 1317, 365–382. [Google Scholar] [CrossRef]
- Wright, A.A.; Cronin, A.; Milne, D.E.; Bookman, M.A.; Burger, R.A.; Cohn, D.E.; Cristea, M.C.; Griggs, J.J.; Keating, N.L.; Levenback, C.F.; et al. Use and Effectiveness of Intraperitoneal Chemotherapy for Treatment of Ovarian Cancer. J. Clin. Oncol. 2015, 33, 2841–2847. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.K.; Walker, J.L. Role of Intraperitoneal Therapy in the Initial Management of Ovarian Cancer. J. Clin. Oncol. 2019, 37, 2416–2419. [Google Scholar] [CrossRef]
- Luo, H.; Xu, X.; Ye, M.; Sheng, B.; Zhu, X. The prognostic value of HER2 in ovarian cancer: A meta-analysis of observational studies. PLoS ONE 2018, 13, e0191972. [Google Scholar] [CrossRef] [PubMed]
- Stancovski, I.; Schindler, D.G.; Waks, T.; Yarden, Y.; Sela, M.; Eshhar, Z. Targeting of T lymphocytes to Neu/HER2-expressing cells using chimeric single chain Fv receptors. J. Immunol. 1993, 151, 6577–6582. [Google Scholar]
- Naviaux, R.K.; Costanzi, E.; Haas, M.; Verma, I.M. The pCL vector system: Rapid production of helper-free, high-titer, recombinant retroviruses. J. Virol. 1996, 70, 5701–5705. [Google Scholar] [CrossRef] [PubMed]
- Globerson Levin, A.; Rawet Slobodkin, M.; Waks, T.; Horn, G.; Ninio-Many, L.; Deshet Unger, N.; Ohayon, Y.; Suliman, S.; Cohen, Y.; Tartakovsky, B.; et al. Treatment of Multiple Myeloma Using Chimeric Antigen Receptor T Cells with Dual Specificity. Cancer Immunol. Res. 2020, 8, 1485–1495. [Google Scholar] [CrossRef] [PubMed]
- Aharon, A.; Horn, G.; Bar-Lev, T.H.; Zagagi Yohay, E.; Waks, T.; Levin, M.; Deshet Unger, N.; Avivi, I.; Globerson Levin, A. Extracellular Vesicles Derived from Chimeric Antigen Receptor-T Cells: A Potential Therapy for Cancer. Hum. Gene Ther. 2021, 32, 1224–1241. [Google Scholar] [CrossRef]
- Gavert, N.; Zwang, Y.; Weiser, R.; Greenberg, O.; Halperin, S.; Jacobi, O.; Mallel, G.; Sandler, O.; Berger, A.J.; Stossel, E.; et al. Ex vivo organotypic cultures for synergistic therapy prioritization identify patient-specific responses to combined MEK and Src inhibition in colorectal cancer. Nat. Cancer 2022, 3, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.W.Y.; Dand, S.; Doig, L.; Papenfuss, A.T.; Scott, C.L.; Ho, G.; Ooi, J.D. T-Cell Receptor Therapy in the Treatment of Ovarian Cancer: A Mini Review. Front. Immunol. 2021, 12, 672502. [Google Scholar] [CrossRef]
- Akter, S.; Rahman, M.A.; Hasan, M.N.; Akhter, H.; Noor, P.; Islam, R.; Shin, Y.; Rahman, M.D.H.; Gazi, M.S.; Huda, M.N.; et al. Recent Advances in Ovarian Cancer: Therapeutic Strategies, Potential Biomarkers, and Technological Improvements. Cells 2022, 11, 650. [Google Scholar] [CrossRef]
- Moore, K.; Colombo, N.; Scambia, G.; Kim, B.G.; Oaknin, A.; Friedlander, M.; Lisyanskaya, A.; Floquet, A.; Leary, A.; Sonke, G.S.; et al. Maintenance Olaparib in Patients with Newly Diagnosed Advanced Ovarian Cancer. N. Engl. J. Med. 2018, 379, 2495–2505. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F.; Qari, H.A.; Upadhyay, T.K.; Alkhateeb, A.F.; Oves, M. Revolutionization in Cancer Therapeutics via Targeting Major Immune Checkpoints PD-1, PD-L1 and CTLA-4. Pharmaceuticals 2022, 15, 335. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, N.; Shi, H. Driving better and safer HER2-specific CARs for cancer therapy. Oncotarget 2017, 8, 62730–62741. [Google Scholar] [CrossRef]
- Szoor, A.; Toth, G.; Zsebik, B.; Szabo, V.; Eshhar, Z.; Abken, H.; Vereb, G. Trastuzumab derived HER2-specific CARs for the treatment of trastuzumab-resistant breast cancer: CAR T cells penetrate and eradicate tumors that are not accessible to antibodies. Cancer Lett. 2020, 484, 1–8. [Google Scholar] [CrossRef] [PubMed]
- de Bree, E.; Michelakis, D. An overview and update of hyperthermic intraperitoneal chemotherapy in ovarian cancer. Expert Opin. Pharmacother. 2020, 21, 1479–1492. [Google Scholar] [CrossRef] [PubMed]
- Koneru, M.; O’Cearbhaill, R.; Pendharkar, S.; Spriggs, D.R.; Brentjens, R.J. A phase I clinical trial of adoptive T cell therapy using IL-12 secreting MUC-16(ecto) directed chimeric antigen receptors for recurrent ovarian cancer. J. Transl. Med. 2015, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Chekmasova, A.A.; Rao, T.D.; Nikhamin, Y.; Park, K.J.; Levine, D.A.; Spriggs, D.R.; Brentjens, R.J. Successful eradication of established peritoneal ovarian tumors in SCID-Beige mice following adoptive transfer of T cells genetically targeted to the MUC16 antigen. Clin. Cancer Res. 2010, 16, 3594–3606. [Google Scholar] [CrossRef] [Green Version]
- Morgan, R.A.; Yang, J.C.; Kitano, M.; Dudley, M.E.; Laurencot, C.M.; Rosenberg, S.A. Case report of a serious adverse event following the administration of T cells transduced with a chimeric antigen receptor recognizing ERBB2. Mol. Ther. 2010, 18, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Vitanza, N.A.; Johnson, A.J.; Wilson, A.L.; Brown, C.; Yokoyama, J.K.; Kunkele, A.; Chang, C.A.; Rawlings-Rhea, S.; Huang, W.; Seidel, K.; et al. Locoregional infusion of HER2-specific CAR T cells in children and young adults with recurrent or refractory CNS tumors: An interim analysis. Nat. Med. 2021, 27, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jiang, S.; Fang, C.; Yang, S.; Olalere, D.; Pequignot, E.C.; Cogdill, A.P.; Li, N.; Ramones, M.; Granda, B.; et al. Affinity-Tuned ErbB2 or EGFR Chimeric Antigen Receptor T Cells Exhibit an Increased Therapeutic Index against Tumors in Mice. Cancer Res. 2015, 75, 3596–3607. [Google Scholar] [CrossRef]
- Kloss, C.C.; Condomines, M.; Cartellieri, M.; Bachmann, M.; Sadelain, M. Combinatorial antigen recognition with balanced signaling promotes selective tumor eradication by engineered T cells. Nat. Biotechnol. 2013, 31, 71–75. [Google Scholar] [CrossRef]
- Wilkie, S.; van Schalkwyk, M.C.; Hobbs, S.; Davies, D.M.; van der Stegen, S.J.; Pereira, A.C.; Burbridge, S.E.; Box, C.; Eccles, S.A.; Maher, J. Dual targeting of ErbB2 and MUC1 in breast cancer using chimeric antigen receptors engineered to provide complementary signaling. J. Clin. Immunol. 2012, 32, 1059–1070. [Google Scholar] [CrossRef]
- Shi, H.; Liu, L.; Wang, Z. Improving the efficacy and safety of engineered T cell therapy for cancer. Cancer Lett. 2013, 328, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Brawley, V.S.; Hegde, M.; Robertson, C.; Ghazi, A.; Gerken, C.; Liu, E.; Dakhova, O.; Ashoori, A.; Corder, A.; et al. Human Epidermal Growth Factor Receptor 2 (HER2) -Specific Chimeric Antigen Receptor-Modified T Cells for the Immunotherapy of HER2-Positive Sarcoma. J. Clin. Oncol. 2015, 33, 1688–1696. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer 5′ to 3′ | Reverse Primer 5′ to 3′ |
|---|---|---|
| PD-1 | CCAGGATGGTTCTTAGACTCCC | TTTAGCACGAAGCTCTCCGAT |
| TIM-3 | AGACAGTGGGATCTACTGCTG | CCTGGTGGTAAGCATCCTTGG |
| GAPDH | AGGGCCCTGACAACTCTTTT | TTACTCCTTGGAGGCCATGT |
| Patient Number | Age | Grade | Tumor Histology | Surgery | Chemotherapy |
|---|---|---|---|---|---|
| 1 | 67 | 3 | HGSOC | Primary Debulking | Carbo Taxol |
| 2 | 65 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 3 | 59 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 4 | 28 | 3 | HGSOC | Primary Debulking | Carbo Taxol + Olaparib |
| 5 | 71 | 3 | HGSOC | Primary Debulking | Carbo Taxol + Avastin |
| 6 | 48 | 3 | HGSOC | Primary Debulking | Carbo Taxol + Avastin |
| 7 | 61 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 8 | 68 | 3 | HGSOC | Primary Debulking | Carbo Taxol + Avastin |
| 9 | 71 | 3 | HGSOC | Interval Debulking + Hipec | Carbo Taxol |
| 10 | 47 | 3 | HGSOC | Primary Debulking | Carbo Taxol |
| 11 | 63 | 3 | HGSOC | Interval Debulking + Hipec | Carbo Taxol |
| 12 | 47 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 13 | 56 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
| 14 | 69 | 3 | HGSOC | Primary Debulking | Carbo Taxol |
| 15 | 64 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
| 16 | 60 | 3 | HGSOC | Primary Debulking | Carbo Taxol |
| 17 | 66 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 18 | 67 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 19 | 83 | 3 | HGSOC | Primary Debulking | No Chemotherapy |
| 20 | 73 | 3 | HGSOC | Primary Debulking | Carbo Taxol |
| 21 | 67 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
| 22 | 56 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
| 23 | 53 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 24 | 46 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
| 25 | 62 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
| 26 | 59 | 3 | HGSOC | Secondary Debulking | |
| 27 | 76 | 3 | HGSOC | Interval Debulking | Carbo Taxol |
| 28 | 70 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
| 29 | 75 | 3 | HGSOC | Interval Debulking | Carbo Taxol + Avastin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deshet-Unger, N.; Horn, G.; Rawet-Slobodkin, M.; Waks, T.; Laskov, I.; Michaan, N.; Raz, Y.; Bar, V.; Zundelevich, A.; Aharon, S.; et al. Comparing Intraperitoneal and Intravenous Personalized ErbB2CAR-T for the Treatment of Epithelial Ovarian Cancer. Biomedicines 2022, 10, 2216. https://doi.org/10.3390/biomedicines10092216
Deshet-Unger N, Horn G, Rawet-Slobodkin M, Waks T, Laskov I, Michaan N, Raz Y, Bar V, Zundelevich A, Aharon S, et al. Comparing Intraperitoneal and Intravenous Personalized ErbB2CAR-T for the Treatment of Epithelial Ovarian Cancer. Biomedicines. 2022; 10(9):2216. https://doi.org/10.3390/biomedicines10092216
Chicago/Turabian StyleDeshet-Unger, Naamit, Galit Horn, Moran Rawet-Slobodkin, Tova Waks, Ido Laskov, Nadav Michaan, Yael Raz, Vered Bar, Adi Zundelevich, Sara Aharon, and et al. 2022. "Comparing Intraperitoneal and Intravenous Personalized ErbB2CAR-T for the Treatment of Epithelial Ovarian Cancer" Biomedicines 10, no. 9: 2216. https://doi.org/10.3390/biomedicines10092216
APA StyleDeshet-Unger, N., Horn, G., Rawet-Slobodkin, M., Waks, T., Laskov, I., Michaan, N., Raz, Y., Bar, V., Zundelevich, A., Aharon, S., Turovsky, L., Mallel, G., Salpeter, S., Neev, G., Hollander, K. S., Katz, B.-Z., Grisaru, D., & Globerson Levin, A. (2022). Comparing Intraperitoneal and Intravenous Personalized ErbB2CAR-T for the Treatment of Epithelial Ovarian Cancer. Biomedicines, 10(9), 2216. https://doi.org/10.3390/biomedicines10092216