
Analysis of Human and Microbial Salivary Proteomes in Children Offers Insights on the Molecular Pathogenesis of Molar-Incisor Hypomineralization

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Standardized Sample Collection
2.3. Sample Processing
2.4. LC-MS/MS Analysis
2.5. Data Analysis
2.5.1. Host Proteome Analysis
2.5.2. Metaproteomic Analysis
3. Results
3.1. Clinical Characteristics of the Study Population
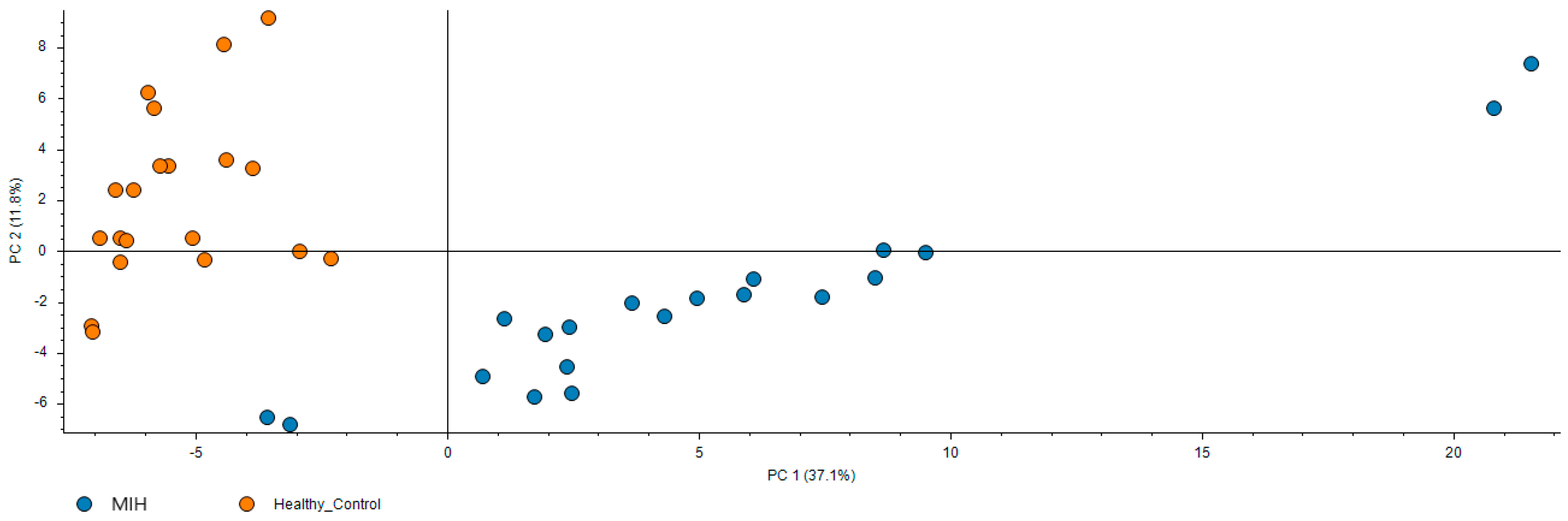
3.2. Proteomic Analysis: Molecular Pathways and Differential Expression of Proteins Involved in Molar-Incisor Hypomineralization
3.3. Microbiome–Microbial Analysis in Molar-Incisor Hypomineralization
4. Discussion
4.1. Differential Expression of Proteins and Immune Response Deregulation in Patients with MIH
4.2. The Oral Microbiome in Patients with MIH
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weerheijm, K.L.; Jalevik, B.; Alaluusua, S. Molar-incisor hypomineralisation. Caries Res. 2001, 35, 390–391. [Google Scholar] [CrossRef] [PubMed]
- Garot, E.; Denis, A.; Delbos, Y.; Manton, D.; Silva, M.; Rouas, P. Are hypomineralised lesions on second primary molars (HSPM) a predictive sign of molar incisor hypomineralisation (MIH)? A systematic review and a meta-analysis. J. Dent. 2018, 72, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Lygidakis, N.A.; Garot, E.; Somani, C.; Taylor, G.D.; Rouas, P.; Wong, F.S.L. Best clinical practice guidance for clinicians dealing with children presenting with molar-incisor-hypomineralisation (MIH): An updated European Academy of Paediatric Dentistry policy document. Eur. Arch. Paediatr. Dent. 2022, 23, 3–21. [Google Scholar] [CrossRef]
- Zhao, D.; Dong, B.; Yu, D.; Ren, Q.; Sun, Y. The prevalence of molar incisor hypomineralization: Evidence from 70 studies. Int. J. Paediatr. Dent. 2018, 28, 170–179. [Google Scholar] [CrossRef]
- Dave, M.; Taylor, G. Global prevalence of molar incisor hypomineralisation. Evid. Based Dent. 2018, 19, 78–79. [Google Scholar] [CrossRef]
- Schwendicke, F.; Elhennawy, K.; Reda, S.; Bekes, K.; Manton, D.J.; Krois, J. Global burden of molar incisor hypomineralization. J. Dent. 2018, 68, 10–18. [Google Scholar] [CrossRef]
- Lygidakis, N.A.; Wong, F.; Jalevik, B.; Vierrou, A.M.; Alaluusua, S.; Espelid, I. Best Clinical Practice Guidance for clinicians dealing with children presenting with Molar-Incisor-Hypomineralisation (MIH): An EAPD Policy Document. Eur. Arch. Paediatr. Dent. 2010, 11, 75–81. [Google Scholar] [CrossRef]
- Garot, E.; Rouas, P.; Somani, C.; Taylor, G.D.; Wong, F.; Lygidakis, N.A. An update of the aetiological factors involved in molar incisor hypomineralisation (MIH): A systematic review and meta-analysis. Eur. Arch. Paediatr. Dent. 2022, 23, 23–38. [Google Scholar] [CrossRef]
- Vieira, A.R.; Kup, E. On the Etiology of Molar-Incisor Hypomineralization. Caries Res. 2016, 50, 166–169. [Google Scholar] [CrossRef]
- Perez, V.A.; Mangum, J.E.; Hubbard, M.J. Pathogenesis of Molar Hypomineralisation: Aged Albumin Demarcates Chalky Regions of Hypomineralised Enamel. Front. Physiol. 2020, 11, 579015. [Google Scholar] [CrossRef]
- Hubbard, M.J.; Mangum, J.E.; Perez, V.A.; Williams, R. A Breakthrough in Understanding the Pathogenesis of Molar Hypomineralisation: The Mineralisation-Poisoning Model. Front. Physiol. 2021, 12, 802833. [Google Scholar] [CrossRef] [PubMed]
- Javaid, M.A.; Ahmed, A.S.; Durand, R.; Tran, S.D. Saliva as a diagnostic tool for oral and systemic diseases. J. Oral Biol. Craniofac. Res. 2016, 6, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Pappa, E.; Kousvelari, E.; Vastardis, H. Saliva in the “Omics” era: A promising tool in paediatrics. Oral Dis. 2019, 25, 16–25. [Google Scholar] [CrossRef]
- Pappa, E.; Vastardis, H.; Mermelekas, G.; Gerasimidi-Vazeou, A.; Zoidakis, J.; Vougas, K. Saliva Proteomics Analysis Offers Insights on Type 1 Diabetes Pathology in a Pediatric Population. Front. Physiol. 2018, 9, 444. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, Q.; Lin, Q.; Duan, Y. Emerging salivary biomarkers by mass spectrometry. Clin. Chim. Acta Int. J. Clin. Chem. 2015, 438, 214–221. [Google Scholar] [CrossRef]
- Pappa, E.; Vougas, K.; Zoidakis, J.; Vastardis, H. Proteomic advances in salivary diagnostics. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140494. [Google Scholar] [CrossRef]
- Bekes, K.; Mitulovic, G.; Meissner, N.; Resch, U.; Gruber, R. Saliva proteomic patterns in patients with molar incisor hypomineralization. Sci. Rep. 2020, 10, 7560. [Google Scholar] [CrossRef]
- Weerheijm, K.L.; Duggal, M.; Mejare, I.; Papagiannoulis, L.; Koch, G.; Martens, L.C.; Hallonsten, A.L. Judgement criteria for molar incisor hypomineralisation (MIH) in epidemiologic studies: A summary of the European meeting on MIH held in Athens, 2003. Eur. J. Paediatr. Dent. 2003, 4, 110–113. [Google Scholar]
- Castagnola, M.; Picciotti, P.M.; Messana, I.; Fanali, C.; Fiorita, A.; Cabras, T.; Calo, L.; Pisano, E.; Passali, G.C.; Iavarone, F.; et al. Potential applications of human saliva as diagnostic fluid. Acta Otorhinolaryngol. Ital. 2011, 31, 347–357. [Google Scholar]
- Castagnola, M.; Cabras, T.; Iavarone, F.; Fanali, C.; Nemolato, S.; Peluso, G.; Bosello, S.L.; Faa, G.; Ferraccioli, G.; Messana, I. The human salivary proteome: A critical overview of the results obtained by different proteomic platforms. Expert Rev. Proteom. 2012, 9, 33–46. [Google Scholar] [CrossRef]
- Pastorino, B.; Touret, F.; Gilles, M.; de Lamballerie, X.; Charrel, R.N. Heat Inactivation of Different Types of SARS-CoV-2 Samples: What Protocols for Biosafety, Molecular Detection and Serological Diagnostics? Viruses 2020, 12, 735. [Google Scholar] [CrossRef]
- Chen, Z.; Sikorski, T.W. Safety considerations in the bioanalytical laboratories handling specimens from coronavirus disease 2019 patients. Bioanalysis 2020, 12, 1219–1222. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Stroggilos, R.; Mokou, M.; Latosinska, A.; Makridakis, M.; Lygirou, V.; Mavrogeorgis, E.; Drekolias, D.; Frantzi, M.; Mullen, W.; Fragkoulis, C.; et al. Proteome-based classification of Nonmuscle Invasive Bladder Cancer. Int. J. Cancer 2020, 146, 281–294. [Google Scholar] [CrossRef]
- Hughes, C.S.; Moggridge, S.; Muller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; Garcia-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Messana, I.; Inzitari, R.; Fanali, C.; Cabras, T.; Castagnola, M. Facts and artifacts in proteomics of body fluids. What proteomics of saliva is telling us? J. Sep. Sci. 2008, 31, 1948–1963. [Google Scholar] [CrossRef]
- Ai, J.Y.; Smith, B.; Wong, D.T. Bioinformatics advances in saliva diagnostics. Int. J. Oral Sci. 2012, 4, 85–87. [Google Scholar] [CrossRef][Green Version]
- Dawes, C.; Wong, D.T.W. Role of Saliva and Salivary Diagnostics in the Advancement of Oral Health. J. Dent. Res. 2019, 98, 133–141. [Google Scholar] [CrossRef]
- Bussaneli, D.G.; Restrepo, M.; Fragelli, C.M.B.; Santos-Pinto, L.; Jeremias, F.; Cordeiro, R.C.L.; Bezamat, M.; Vieira, A.R.; Scarel-Caminaga, R.M. Genes Regulating Immune Response and Amelogenesis Interact in Increasing the Susceptibility to Molar-Incisor Hypomineralization. Caries Res. 2019, 53, 217–227. [Google Scholar] [CrossRef]
- Americano, G.C.; Jacobsen, P.E.; Soviero, V.M.; Haubek, D. A systematic review on the association between molar incisor hypomineralization and dental caries. Int. J. Paediatr. Dent. 2017, 27, 11–21. [Google Scholar] [CrossRef]
- Farah, R.; Drummond, B.; Swain, M.; Williams, S. Linking the clinical presentation of molar-incisor hypomineralisation to its mineral density. Int. J. Paediatr. Dent. 2010, 20, 353–360. [Google Scholar] [CrossRef]
- Elhennawy, K.; Schwendicke, F. Managing molar-incisor hypomineralization: A systematic review. J. Dent. 2016, 55, 16–24. [Google Scholar] [CrossRef]
- Mangum, J.E.; Crombie, F.A.; Kilpatrick, N.; Manton, D.J.; Hubbard, M.J. Surface integrity governs the proteome of hypomineralized enamel. J. Dent. Res. 2010, 89, 1160–1165. [Google Scholar] [CrossRef]
- Yuan, Z.A.; McAndrew, K.S.; Collier, P.M.; Koyama, E.; Chen, E.; Sandgren, E.P.; Gibson, C.W. Albumin gene expression during mouse odontogenesis. Adv. Dent. Res. 1996, 10, 119–124. [Google Scholar] [CrossRef]
- Palomo, J.; Dietrich, D.; Martin, P.; Palmer, G.; Gabay, C. The interleukin (IL)-1 cytokine family--Balance between agonists and antagonists in inflammatory diseases. Cytokine 2015, 76, 25–37. [Google Scholar] [CrossRef]
- Suarez, L.J.; Garzon, H.; Arboleda, S.; Rodriguez, A. Oral Dysbiosis and Autoimmunity: From Local Periodontal Responses to an Imbalanced Systemic Immunity. A Review. Front Immunol. 2020, 11, 591255. [Google Scholar] [CrossRef]
- Gao, L.; Cheng, Z.; Zhu, F.; Bi, C.; Shi, Q.; Chen, X. The Oral Microbiome and Its Role in Systemic Autoimmune Diseases: A Systematic Review of Big Data Analysis. Front. Big Data 2022, 5, 927520. [Google Scholar] [CrossRef]
- Belibasakis, G.N.; Bostanci, N.; Marsh, P.D.; Zaura, E. Applications of the oral microbiome in personalized dentistry. Arch. Oral Biol. 2019, 104, 7–12. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| MIH | Controls | |
|---|---|---|
| Number of Subjects | 10 | 10 |
| Age (yrs), mean (SD) | 14.8 ± 1.7 | 15.2 ± 1.8 |
| Gender, n (M/F) | 4/6 | 4/6 |
| DMFT | 2.3 ± 0.6 | 2.5 ± 0.4 |
| Plaque Index (PI) | 0.8 ± 0.05 | 0.9 ± 0.03 |
| Gingival Index (GI) | 0.7 ± 0.2 | 0.8 ± 0.3 |
| Protein Name | Ratio MIH/Control | p Value | Function |
|---|---|---|---|
| Immunoglobulin lambda variable 2-11 | 2.44 | 7.12 × 10−6 | Immune response |
| Complement C5 | 2.32 | 2.42 × 10−5 | Complement cascade |
| Plasma kallikrein | 1.67 | 1.40 × 10−2 | Inflammatory protease |
| Bactericidal permeability-increasing protein | 1.53 | 4.57 × 10−2 | Antibacterial activity |
| Antileukoproteinase | 0.61 | 4.25 × 10−3 | Anti-inflammatory protease inhibitor |
| Glutaredoxin-3 | Only in Control | 1.00 × 10−17 | Antioxidant defence |
| Glutathione S-transferase Mu 4 | Only in Control | 1.00 × 10−17 | Antioxidant defence |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pappa, E.; Vastardis, H.; Makridakis, M.; Zoidakis, J.; Vougas, K.; Stamatakis, G.; Samiotaki, M.; Rahiotis, C. Analysis of Human and Microbial Salivary Proteomes in Children Offers Insights on the Molecular Pathogenesis of Molar-Incisor Hypomineralization. Biomedicines 2022, 10, 2061. https://doi.org/10.3390/biomedicines10092061
Pappa E, Vastardis H, Makridakis M, Zoidakis J, Vougas K, Stamatakis G, Samiotaki M, Rahiotis C. Analysis of Human and Microbial Salivary Proteomes in Children Offers Insights on the Molecular Pathogenesis of Molar-Incisor Hypomineralization. Biomedicines. 2022; 10(9):2061. https://doi.org/10.3390/biomedicines10092061
Chicago/Turabian StylePappa, Eftychia, Heleni Vastardis, Manousos Makridakis, Jerome Zoidakis, Konstantinos Vougas, George Stamatakis, Martina Samiotaki, and Christos Rahiotis. 2022. "Analysis of Human and Microbial Salivary Proteomes in Children Offers Insights on the Molecular Pathogenesis of Molar-Incisor Hypomineralization" Biomedicines 10, no. 9: 2061. https://doi.org/10.3390/biomedicines10092061
APA StylePappa, E., Vastardis, H., Makridakis, M., Zoidakis, J., Vougas, K., Stamatakis, G., Samiotaki, M., & Rahiotis, C. (2022). Analysis of Human and Microbial Salivary Proteomes in Children Offers Insights on the Molecular Pathogenesis of Molar-Incisor Hypomineralization. Biomedicines, 10(9), 2061. https://doi.org/10.3390/biomedicines10092061