
Bladder Tissue Microbiome Composition in Patients of Bladder Cancer or Benign Prostatic Hyperplasia and Related Human Beta Defensin Levels


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Isolation, 16S rRNA Gene Library Preparation and MiSeq Sequencing
2.3. Defensin Expression Assays
2.4. ELISA (Enzyme Linked Immunosorbent Assay)
2.5. Statistical Analysis
2.6. Ethical Considerations
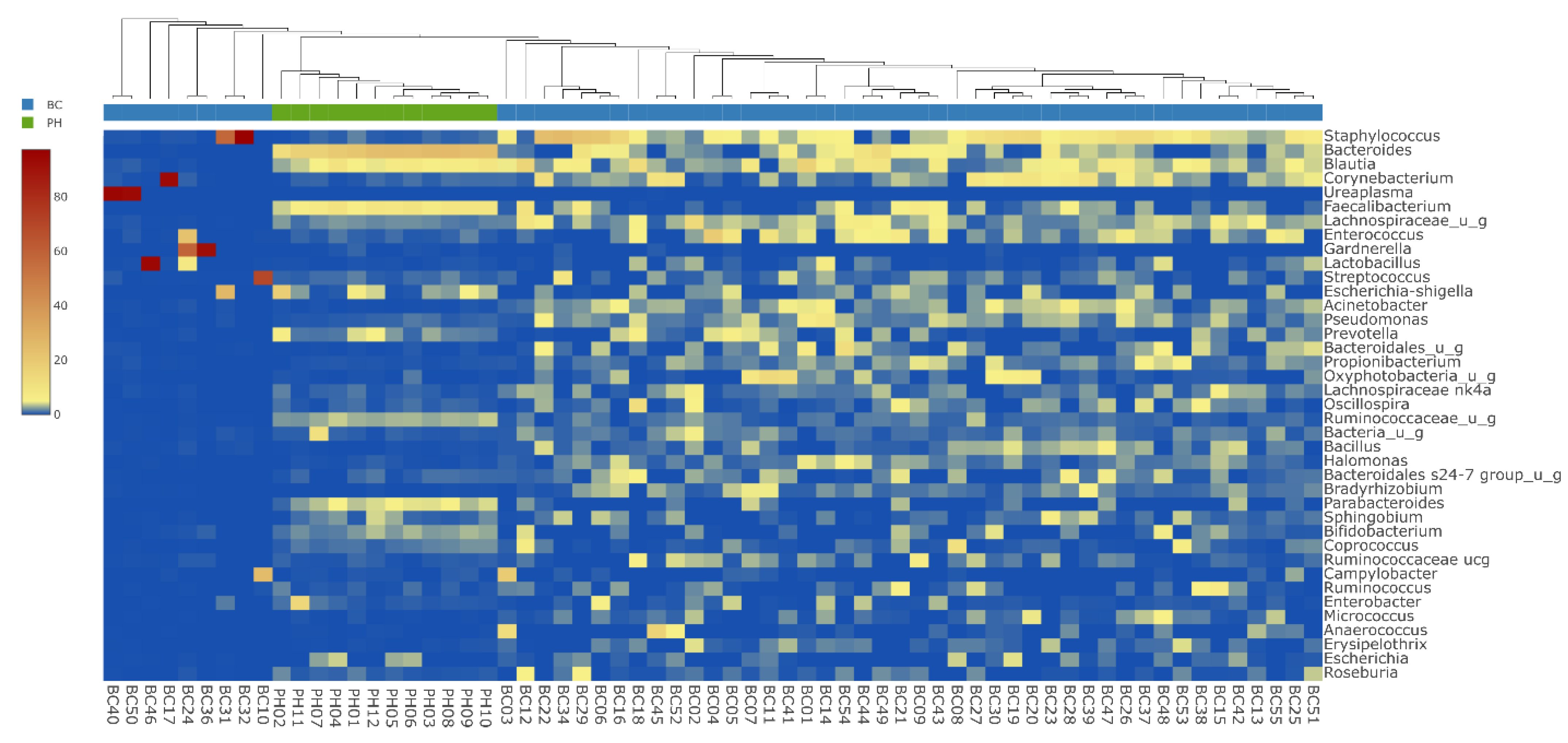
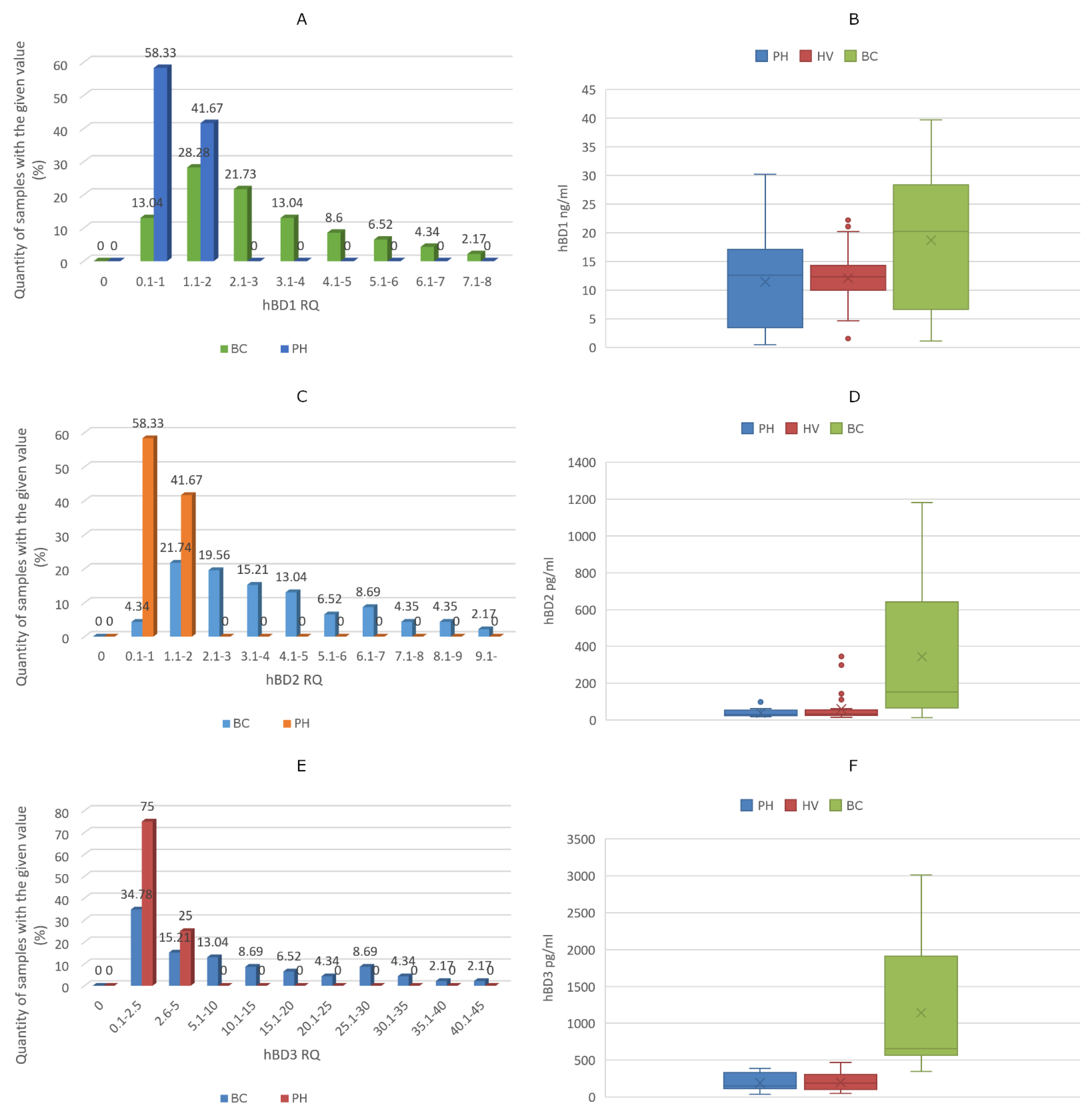
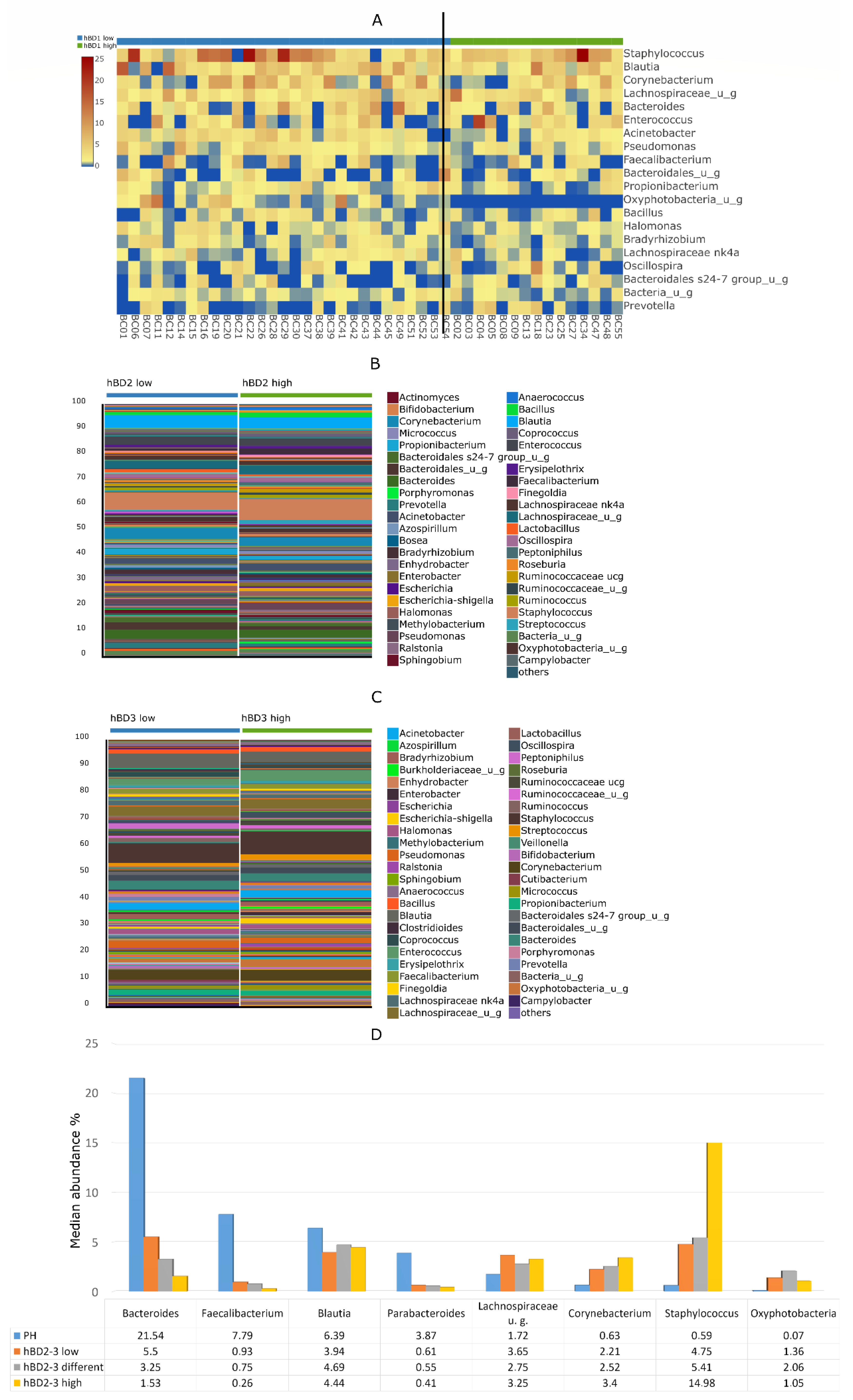
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richters, A.; Aben, K.K.H.; Kiemeney, L.A.L.M. The global burden of urinary bladder cancer: An update. World J. Urol. 2020, 38, 1895–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powles, T.; Bellmunt, J.; Comperat, E.; De Santis, M.; Huddart, R.; Loriot, Y.; Necchi, A.; Valderrama, B.P.; Ravaud, A.; Shariat, S.F.; et al. Electronic address: Clinicalguidelines@esmo.org. Bladder cancer: ESMO Clinical Practice Guideline for diagnosis, treatment and follow-up. Ann. Oncol. 2022, 33, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Cumberbatch, M.G.; Rota, M.; Catto, J.W.; La Vecchia, C. The Role of Tobacco Smoke in Bladder and Kidney Carcinogenesis: A Comparison of Exposures and Meta-analysis of Incidence and Mortality Risks. Eur. Urol. 2016, 70, 458–466. [Google Scholar] [CrossRef] [Green Version]
- Cogliano, V.J.; Baan, R.; Straif, K.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; et al. Preventable exposures associated with human cancers. J. Natl. Cancer Inst. 2011, 103, 1827–1839. [Google Scholar] [CrossRef] [PubMed]
- Steinmaus, C.M.; Ferreccio, C.; Romo, J.A.; Yuan, Y.; Cortes, S.; Marshall, G.; Moore, L.E.; Balmes, J.R.; Liaw, J.; Golden, T.; et al. Drinking water arsenic in northern chile: High cancer risks 40 years after exposure cessation. Cancer Epidemiol. Biomark. Prev. 2013, 22, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duell, B.L.; Carey, A.J.; Tan, C.K.; Cui, X.; Webb, R.I.; Totsika, M.; Schembri, M.A.; Derrington, P.; Irving-Rodgers, H.; Brooks, A.J.; et al. Innate transcriptional networks activated in bladder in response to uropathogenic Escherichia coli drive diverse biological pathways and rapid synthesis of IL-10 for defense against bacterial urinary tract infection. J. Immunol. 2012, 188, 781–792. [Google Scholar] [CrossRef] [Green Version]
- Bajic, P.; Wolfe, A.J.; Gupta, G.N. The Urinary Microbiome: Implications in Bladder Cancer Pathogenesis and Therapeutics. Urology 2019, 126, 10–15. [Google Scholar] [CrossRef]
- Bucevic Popovic, V.; Situm, M.; Chow, C.T.; Chan, L.S.; Roje, B.; Terzic, J. The urinary microbiome associated with bladder cancer. Sci. Rep. 2018, 8, 12157. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Yang, L.; Lee, P.; Huang, W.C.; Nossa, C.; Ma, Y.; Deng, F.M.; Zhou, M.; Melamed, J.; Pei, Z. Mini-review: Perspective of the microbiome in the pathogenesis of urothelial carcinoma. Am. J. Clin. Exp. Urol. 2014, 2, 57–61. [Google Scholar]
- Wu, P.; Zhang, G.; Zhao, J.; Chen, J.; Chen, Y.; Huang, W.; Zhong, J.; Zeng, J. Profiling the Urinary Microbiota in Male Patients With Bladder Cancer in China. Front. Cell Infect. Microbiol. 2018, 8, 167. [Google Scholar] [CrossRef]
- Han, Q.; Wang, R.; Sun, C.; Jin, X.; Liu, D.; Zhao, X.; Wang, L.; Ji, N.; Li, J.; Zhou, Y.; et al. Human beta-defensin-1 suppresses tumor migration and invasion and is an independent predictor for survival of oral squamous cell carcinoma patients. PLoS ONE 2014, 9, e91867. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, Á.H.; Martínez Velázquez, M.; Prado Montes de Oca, E. Human β-defensin 1 update: Potential clinical applications of the restless warrior. Int. J. Biochem. Cell Biol. 2018, 104, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.A.; Kim, K.H.; Lee, T.J.; Park, E.S.; Kim, M.K.; Myung, S.C. A role of human beta defensin-1 in predicting prostatic adenocarcinoma in cases of false-negative biopsy. APMIS 2017, 125, 1063–1069. [Google Scholar] [CrossRef]
- Sun, C.Q.; Arnold, R.; Fernandez-Golarz, C.; Parrish, A.B.; Almekinder, T.; He, J.; Ho, S.M.; Svoboda, P.; Pohl, J.; Marshall, F.F.; et al. Human beta-defensin-1, a potential chromosome 8p tumor suppressor: Control of transcription and induction of apoptosis in renal cell carcinoma. Cancer Res. 2006, 66, 8542–8549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.Q.; Arnold, R.S.; Hsieh, C.L.; Dorin, J.R.; Lian, F.; Li, Z.; Petros, J.A. Discovery and mechanisms of host defense to oncogenesis: Targeting the beta-defensin-1 peptide as a natural tumor inhibitor. Cancer Biol. Ther. 2019, 20, 774–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Kim, S.J.; Lee, K.M.; Chang, I.H. Human beta-defensin 2 may inhibit internalisation of bacillus Calmette-Guerin (BCG) in bladder cancer cells. BJU Int. 2013, 112, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Whang, Y.M.; Jin, S.B.; Park, S.I.; Chang, I.H. MEK inhibition enhances efficacy of bacillus Calmette-Guerin on bladder cancer cells by reducing release of Toll-like receptor 2-activated antimicrobial peptides. Oncotarget 2017, 8, 53168–53179. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Huang, Z.; Huang, P.; Li, K.; Zeng, J.; Wen, Y.; Li, B.; Zhao, J.; Wu, P. Urogenital Microbiota:Potentially Important Determinant of PD-L1 Expression in Male Patients with Non-muscle Invasive Bladder Cancer. BMC Microbiol. 2022, 22, 7. [Google Scholar] [CrossRef]
- Mansour, B.; Monyok, A.; Makra, N.; Gajdacs, M.; Vadnay, I.; Ligeti, B.; Juhasz, J.; Szabo, D.; Ostorhazi, E. Bladder cancer-related microbiota: Examining differences in urine and tissue samples. Sci. Rep. 2020, 10, 11042. [Google Scholar] [CrossRef]
- Yan, Q.; Wi, Y.M.; Thoendel, M.J.; Raval, Y.S.; Greenwood-Quaintance, K.E.; Abdel, M.P.; Jeraldo, P.R.; Chia, N.; Patel, R. Evaluation of the CosmosID Bioinformatics Platform for Prosthetic Joint-Associated Sonicate Fluid Shotgun Metagenomic Data Analysis. J. Clin. Microbiol. 2019, 57, e01182-18. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Eidinger, D.; Bruce, A.W. Intracavitary Bacillus Calmette-Guerin in the treatment of superficial bladder tumors. J. Urol. 1976, 116, 180–183. [Google Scholar] [CrossRef]
- Yu, D.S.; Wu, C.L.; Ping, S.Y.; Keng, C.; Shen, K.H. Bacille Calmette-Guerin can induce cellular apoptosis of urothelial cancer directly through toll-like receptor 7 activation. Kaohsiung J. Med. Sci. 2015, 31, 391–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- See, W.A.; Zhang, G.; Chen, F.; Cao, Y.; Langenstroer, P.; Sandlow, J. Bacille-Calmette Guerin induces caspase-independent cell death in urothelial carcinoma cells together with release of the necrosis-associated chemokine high molecular group box protein 1. BJU Int. 2009, 103, 1714–1720. [Google Scholar] [CrossRef]
- Shah, G.; Zielonka, J.; Chen, F.; Zhang, G.; Cao, Y.; Kalyanaraman, B.; See, W. H2O2 generation by bacillus Calmette-Guerin induces the cellular oxidative stress response required for bacillus Calmette-Guerin direct effects on urothelial carcinoma biology. J. Urol. 2014, 192, 1238–1248. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A.; Brown, R.; Williams, J.; White, P.; Jacobson, S.K.; Marchesi, J.R.; Drake, M.J. The human urinary microbiome; bacterial DNA in voided urine of asymptomatic adults. Front. Cell Infect. Microbiol. 2013, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Pan, T.; Yan, L.; Jin, T.; Zhang, R.; Chen, B.; Feng, J.; Duan, T.; Xiang, Y.; Zhang, M.; et al. The inflammatory microenvironment and the urinary microbiome in the initiation and progression of bladder cancer. Genes Dis. 2021, 8, 781–797. [Google Scholar] [CrossRef]
- Oresta, B.; Braga, D.; Lazzeri, M.; Frego, N.; Saita, A.; Faccani, C.; Fasulo, V.; Colombo, P.; Guazzoni, G.; Hurle, R.; et al. The Microbiome of Catheter Collected Urine in Males with Bladder Cancer According to Disease Stage. J. Urol. 2021, 205, 86–93. [Google Scholar] [CrossRef]
- Bersanelli, M.; Santoni, M.; Ticinesi, A.; Buti, S. The Urinary Microbiome and Anticancer Immunotherapy: The Potentially Hidden Role of Unculturable Microbes. Target Oncol. 2019, 14, 247–252. [Google Scholar] [CrossRef]
- Li, W.T.; Iyangar, A.S.; Reddy, R.; Chakladar, J.; Bhargava, V.; Sakamoto, K.; Ongkeko, W.M.; Rajasekaran, M. The Bladder Microbiome Is Associated with Epithelial-Mesenchymal Transition in Muscle Invasive Urothelial Bladder Carcinoma. Cancers 2021, 13, 3649. [Google Scholar] [CrossRef]
- Cho, M.J.; Kim, M.J.; Kim, K.; Choi, Y.W.; Lee, S.J.; Whang, Y.M.; Chang, I.H. The immunotherapeutic effects of recombinant Bacillus Calmette-Guerin resistant to antimicrobial peptides on bladder cancer cells. Biochem. Biophys. Res. Commun. 2019, 509, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Pederzoli, F.; Bandini, M.; Raggi, D.; Marandino, L.; Basile, G.; Alfano, M.; Colombo, R.; Salonia, A.; Briganti, A.; Gallina, A.; et al. Is There a Detrimental Effect of Antibiotic Therapy in Patients with Muscle-invasive Bladder Cancer Treated with Neoadjuvant Pembrolizumab? Eur. Urol. 2021, 80, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Ishiyama, Y.; Kondo, T.; Nemoto, Y.; Kobari, Y.; Ishihara, H.; Tachibana, H.; Yoshida, K.; Hashimoto, Y.; Takagi, T.; Iizuka, J.; et al. Antibiotic use and survival of patients receiving pembrolizumab for chemotherapy-resistant metastatic urothelial carcinoma. Urol. Oncol. 2021, 39, 834. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Halder, C.V.; Faria, A.V.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Sha, S.; Ni, L.; Stefil, M.; Dixon, M.; Mouraviev, V. The human gastrointestinal microbiota and prostate cancer development and treatment. Investig. Clin. Urol. 2020, 61, S43–S50. [Google Scholar] [CrossRef]
- Ma, J.; Sun, L.; Liu, Y.; Ren, H.; Shen, Y.; Bi, F.; Zhang, T.; Wang, X. Alter between gut bacteria and blood metabolites and the anti-tumor effects of Faecalibacterium prausnitzii in breast cancer. BMC Microbiol. 2020, 20, 82. [Google Scholar] [CrossRef]
- Ma, W.; Zhang, W.; Shen, L.; Liu, J.; Yang, F.; Maskey, N.; Wang, H.; Zhang, J.; Yan, Y.; Yao, X. Can Smoking Cause Differences in Urine Microbiome in Male Patients with Bladder Cancer? A Retrospective Study. Front. Oncol. 2021, 11, 677605. [Google Scholar] [CrossRef]
- Pederzoli, F.; Ferrarese, R.; Amato, V.; Locatelli, I.; Alchera, E.; Luciano, R.; Nebuloni, M.; Briganti, A.; Gallina, A.; Colombo, R.; et al. Sex-specific Alterations in the Urinary and Tissue Microbiome in Therapy-naive Urothelial Bladder Cancer Patients. Eur. Urol. Oncol. 2020, 3, 784–788. [Google Scholar] [CrossRef]
- Nienhouse, V.; Gao, X.; Dong, Q.; Nelson, D.E.; Toh, E.; McKinley, K.; Schreckenberger, P.; Shibata, N.; Fok, C.S.; Mueller, E.R.; et al. Interplay between bladder microbiota and urinary antimicrobial peptides: Mechanisms for human urinary tract infection risk and symptom severity. PLoS ONE 2014, 9, e114185. [Google Scholar]
- Miao, C.; Ren, Y.; Chen, M.; Wang, Z.; Wang, T. Microcystin-LR promotes migration and invasion of colorectal cancer through matrix metalloproteinase-13 up-regulation. Mol. Carcinog. 2016, 55, 514–524. [Google Scholar] [CrossRef]
- Brunner, A.; Medvecz, M.; Makra, N.; Sardy, M.; Komka, K.; Gugolya, M.; Szabo, D.; Gajdacs, M.; Ostorhazi, E. Human beta defensin levels and vaginal microbiome composition in post-menopausal women diagnosed with lichen sclerosus. Sci. Rep. 2021, 11, 15999. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.J.; Unholzer, A.; Schaller, M.; Schäfer-Korting, M.; Korting, H.C. Human defensins. J. Mol. Med. 2005, 83, 587–595. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansour, B.; Monyók, Á.; Gajdács, M.; Stercz, B.; Makra, N.; Pénzes, K.; Vadnay, I.; Szabó, D.; Ostorházi, E. Bladder Tissue Microbiome Composition in Patients of Bladder Cancer or Benign Prostatic Hyperplasia and Related Human Beta Defensin Levels. Biomedicines 2022, 10, 1758. https://doi.org/10.3390/biomedicines10071758
Mansour B, Monyók Á, Gajdács M, Stercz B, Makra N, Pénzes K, Vadnay I, Szabó D, Ostorházi E. Bladder Tissue Microbiome Composition in Patients of Bladder Cancer or Benign Prostatic Hyperplasia and Related Human Beta Defensin Levels. Biomedicines. 2022; 10(7):1758. https://doi.org/10.3390/biomedicines10071758
Chicago/Turabian StyleMansour, Bassel, Ádám Monyók, Márió Gajdács, Balázs Stercz, Nóra Makra, Kinga Pénzes, István Vadnay, Dóra Szabó, and Eszter Ostorházi. 2022. "Bladder Tissue Microbiome Composition in Patients of Bladder Cancer or Benign Prostatic Hyperplasia and Related Human Beta Defensin Levels" Biomedicines 10, no. 7: 1758. https://doi.org/10.3390/biomedicines10071758
APA StyleMansour, B., Monyók, Á., Gajdács, M., Stercz, B., Makra, N., Pénzes, K., Vadnay, I., Szabó, D., & Ostorházi, E. (2022). Bladder Tissue Microbiome Composition in Patients of Bladder Cancer or Benign Prostatic Hyperplasia and Related Human Beta Defensin Levels. Biomedicines, 10(7), 1758. https://doi.org/10.3390/biomedicines10071758