
Interleukin-18 Binding Protein in Immune Regulation and Autoimmune Diseases


Abstract
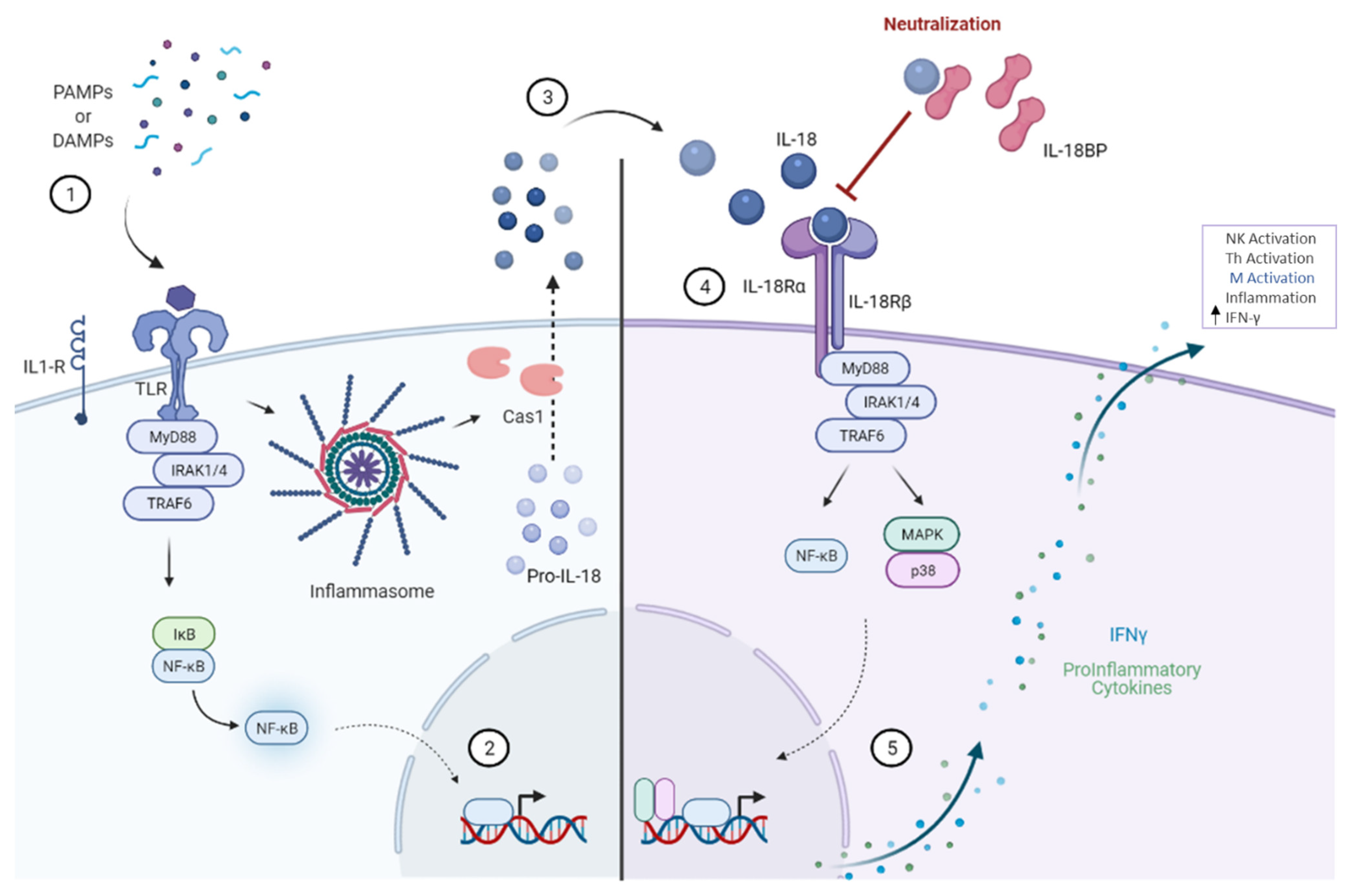
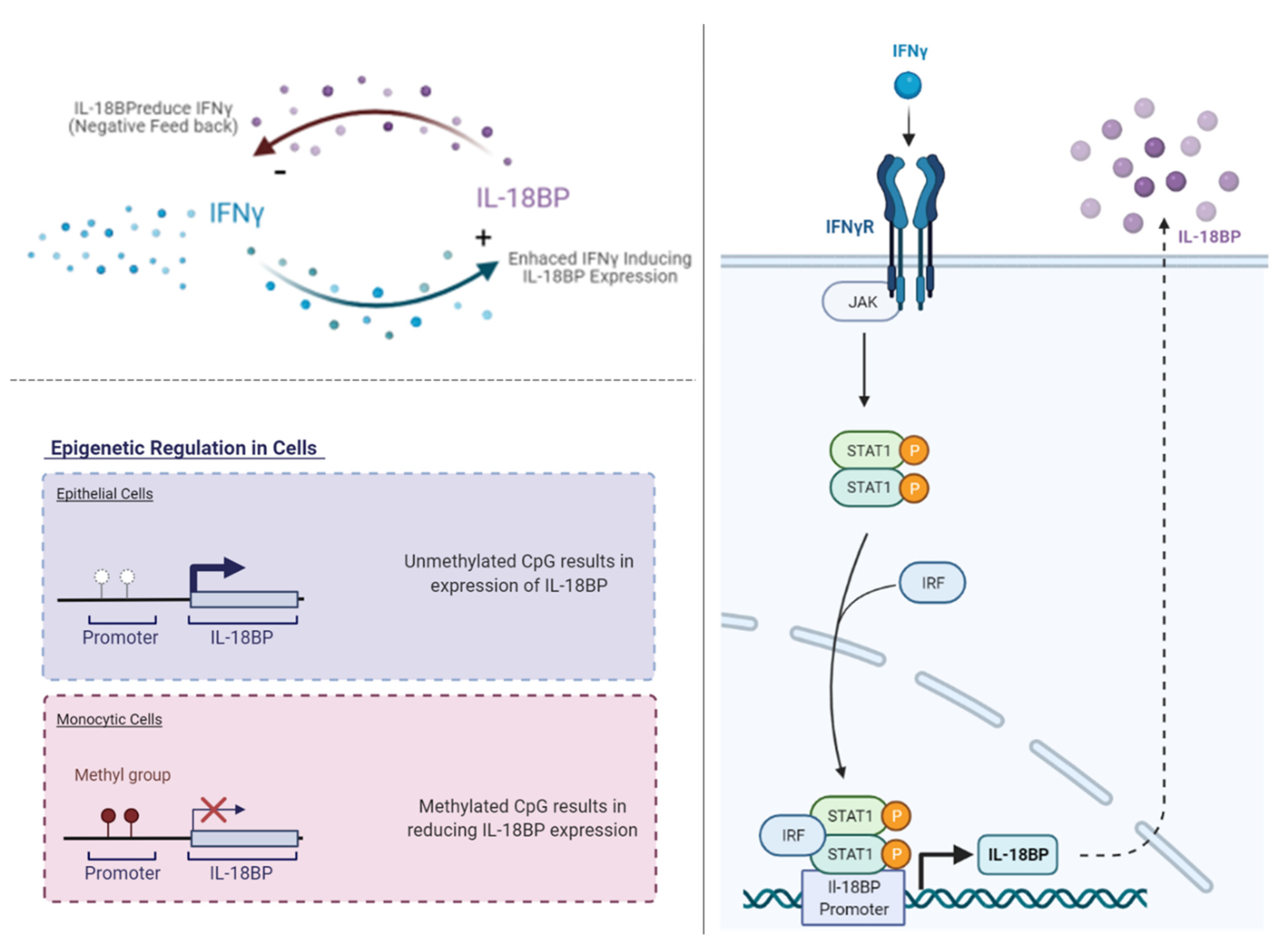
1. Regulation of IL-18BP
2. Difference in the Biological Activity of IL-18BP Isoforms
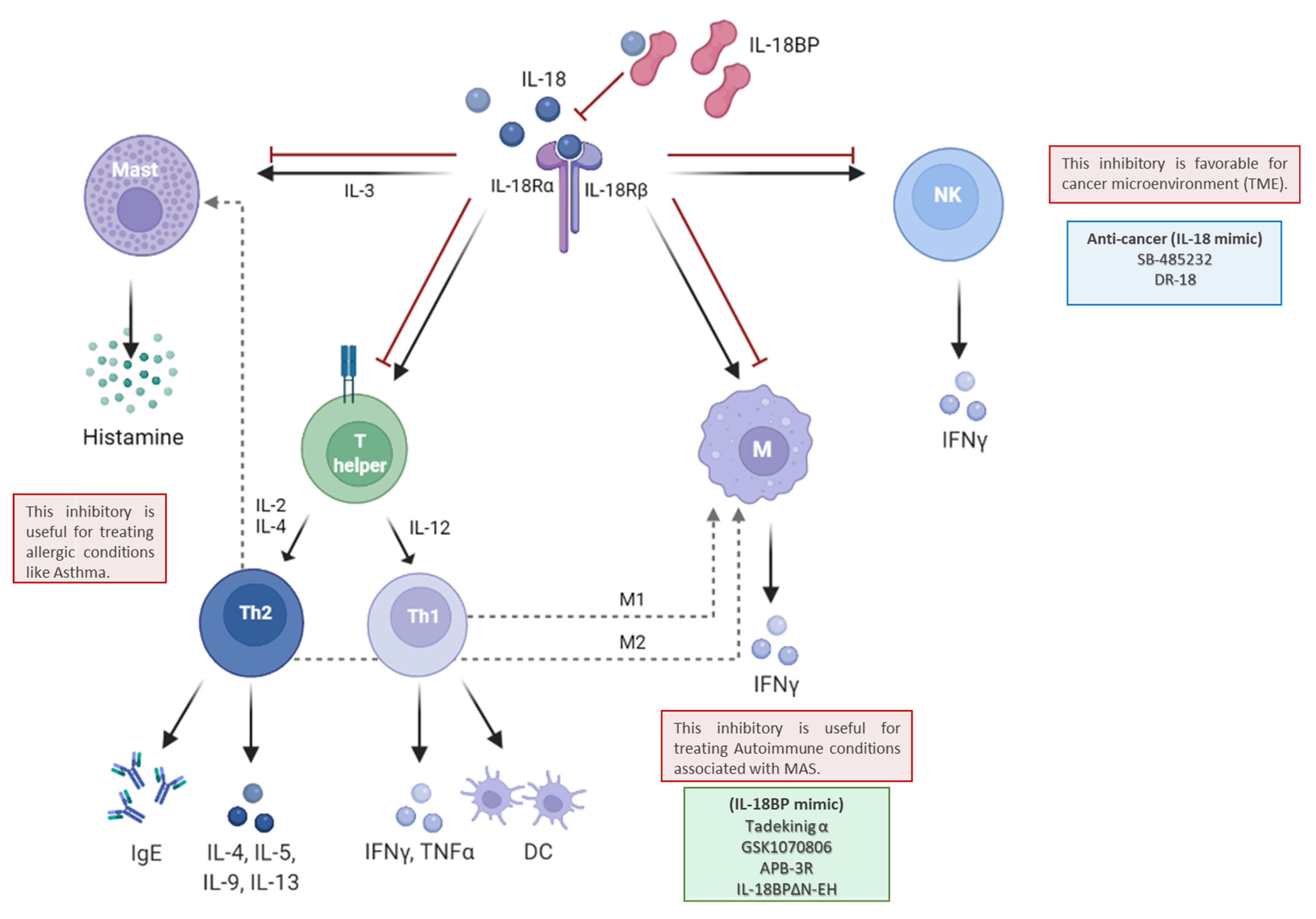
3. IL-18BP in Immunity Balancing between Th1 and Th2 Immune Response
4. IL-18BP in Immunity Balancing between Th17 and Treg
5. IL-18BP in Autoimmune Diseases
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Atopic dermatitis |
| AOSD | Adult-onset Still’s disease |
| APCs | Antigen-presenting cells |
| CAP | Community-acquired pneumonia |
| CD | Crohn’s disease |
| CEBP-β | CCAAT-enhancer binding protein beta |
| COVID-19 | Coronavirus disease 2019 |
| CRP | C-reactive protein |
| DAMPs | damage-associated molecular patterns |
| DCs | Dendritic cells |
| GAS | Gamma-activated sequence |
| GvHD | Graft versus host disease |
| HLH | hemophagocytic lymphohistiocytosis |
| Ig | Immunoglobulin |
| IBD | Inflammatory bowel disease |
| IFN | Interferon |
| IFNγR1 | IFNγ receptor 1 |
| IFNγR2 | IFNγ receptor 2 |
| IRAK | interleukin-1 receptor-associated kinase |
| IRFs | Interferon regulatory factors |
| IRF-E | IFN regulatory factor 1 response element |
| IL | Interleukin |
| IL-1R | IL-1 receptor |
| IL-18BP | IL-18 binding protein |
| IL-18Rα | IL-18 receptor alpha chain |
| IL-18Rβ | IL-18 receptor beta chain |
| JAK | Janus kinase |
| MAS | Macrophage activated syndrome |
| miRNAs | MicroRNAs |
| MTB | Mycobacterium tuberculosis |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK | Natural killer |
| NLRC4 | NLR family CARD domain containing 4 |
| PAMPs | Pathogen-associated molecular patterns |
| PAPA | Pyoderma gangrenosum and acne |
| Pro-IL-18 | pro-interleukin IL-18 |
| PRRs | Pattern recognition receptors |
| PTM | Post translational modifications |
| RA | Rheumatoid arthritis |
| SARS CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| STAT1 | signal transducer and activator of transcription 1 |
| TB | Tuberculosis |
| TLR | Toll-like receptors |
| TRAF | TNF receptor associated factor |
| vIL-18BP | viral IL-18BP |
References
- Okamura, H.; Nagata, K.; Komatsu, T.; Tanimoto, T.; Nukata, Y.; Tanabe, F.; Akita, K.; Torigoe, K.; Okura, T.; Fukuda, S. A novel costimulatory factor for gamma interferon induction found in the livers of mice causes endotoxic shock. Infect. Immun. 1995, 63, 3966–3972. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 Binding Protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef] [PubMed]
- Halim, A.; Nilsson, J.; Rüetschi, U.; Hesse, C.; Larson, G. Human urinary glycoproteomics; attachment site specific analysis of N-and O-linked glycosylations by CID and ECD. Mol. Cell. Proteom. 2012, 11, M111.013649. [Google Scholar] [CrossRef]
- Kim, S.-H.; Eisenstein, M.; Reznikov, L.; Fantuzzi, G.; Novick, D.; Rubinstein, M.; Dinarello, C.A. Structural requirements of six naturally occurring isoforms of the IL-18 binding protein to inhibit IL-18. Proc. Natl. Acad. Sci. USA 2000, 97, 1190–1195. [Google Scholar] [CrossRef]
- Kaplanski, G. Interleukin-18: Biological properties and role in disease pathogenesis. Immunol. Rev. 2018, 281, 138–153. [Google Scholar] [CrossRef]
- Novick, D.; Kim, S.-H.; Fantuzzi, G.; Reznikov, L.L.; Dinarello, C.A.; Rubinstein, M. Interleukin-18 Binding Protein: A Novel Modulator of the Th1 Cytokine Response. Immunity 1999, 10, 127–136. [Google Scholar] [CrossRef]
- Aizawa, Y.; Akita, K.; Taniai, M.; Torigoe, K.; Mori, T.; Nishida, Y.; Ushio, S.; Nukada, Y.; Tanimoto, T.; Ikegami, H.; et al. Cloning and expression of interleukin-18 binding protein. FEBS Lett. 1999, 445, 338–342. [Google Scholar] [CrossRef]
- Im, S.-H.; Kim, S.-H.; Azam, T.; Venkatesh, N.; Dinarello, C.A.; Fuchs, S.; Souroujon, M.C. Rat Interleukin-18 Binding Protein: Cloning, Expression, and Characterization. J. Interf. Cytokine Res. 2002, 22, 321–328. [Google Scholar] [CrossRef]
- Paulukat, J.; Bosmann, M.; Nold, M.; Garkisch, S.; Kämpfer, H.; Frank, S.; Raedle, J.; Zeuzem, S.; Pfeilschifter, J.; Mühl, H. Expression and release of IL-18 binding protein in response to IFN-γ. J. Immunol. 2001, 167, 7038–7043. [Google Scholar] [CrossRef]
- Veenstra, K.G.; Jonak, Z.L.; Trulli, S.; Gollob, J.A. IL-12 induces monocyte IL-18 binding protein expression via IFN-γ. J. Immunol. 2002, 168, 2282–2287. [Google Scholar] [CrossRef] [PubMed]
- Hurgin, V.; Novick, D.; Rubinstein, M. The promoter of IL-18 binding protein: Activation by an IFN-γ-induced complex of IFN regulatory factor 1 and CCAAT/enhancer binding protein β. Proc. Natl. Acad. Sci. USA 2002, 99, 16957–16962. [Google Scholar] [CrossRef]
- Möller, B.; Paulukat, J.; Nold, M.; Behrens, M.; Kukoc-Zivojnov, N.; Kaltwasser, J.P.; Pfeilschifter, J.; Mühl, H. Interferon-gamma induces expression of interleukin-18 binding protein in fibroblast-like synoviocytes. Rheumatology 2003, 42, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Okamura, H.; Wada, M.; Nagata, K.; Tamura, T. Endotoxin-induced serum factor that stimulates gamma interferon production. Infect. Immun. 1989, 57, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Esmailbeig, M.; Ghaderi, A. Interleukin-18: A regulator of cancer and autoimmune diseases. Eur. Cytokine Netw. 2017, 28, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Chen, J.; Lu, J.; Yi, L.; Tong, X.; Kang, L.; Pei, S.; Ouyang, Y.; Jiang, L.; Ding, Y.; et al. Roles of inflammation factors in melanogenesis. Mol. Med. Rep. 2020, 21, 1421–1430. [Google Scholar] [CrossRef]
- Vecchié, A.; Bonaventura, A.; Toldo, S.; Dagna, L.; Dinarello, C.A.; Abbate, A. IL-18 and infections: Is there a role for targeted therapies? J. Cell. Physiol. 2021, 236, 1638–1657. [Google Scholar] [CrossRef]
- Novick, D.; Schwartsburd, B.; Pinkus, R.; Suissa, D.; Belzer, I.; Sthoeger, Z.; Keane, W.F.; Chvatchko, Y.; Kim, S.-H.; Fantuzzi, G.; et al. A novel IL-18BP ELISA shows elevated serum IL-18BP in sepsis and extensive decrease of free IL-18. Cytokine 2001, 14, 334–342. [Google Scholar] [CrossRef]
- Principe, S.; Jones, E.E.; Kim, Y.; Sinha, A.; Nyalwidhe, J.O.; Brooks, J.; Semmes, O.J.; Troyer, D.A.; Lance, R.S.; Kislinger, T.; et al. In-depth proteomic analyses of exosomes isolated from expressed prostatic secretions in urine. Proteomics 2013, 13, 1667–1671. [Google Scholar] [CrossRef]
- Sonek, J.; Blumenfeld, M.; Foley, M.; Johnson, F.; Iams, J. Cervical length may change during ultrasonographic examination. Am. J. Obstet. Gynecol. 1990, 62, 1355–1357. [Google Scholar] [CrossRef]
- Taylor, T.D.; Noguchi, H.; Totoki, Y.; Toyoda, A.; Kuroki, Y.; Dewar, K.; Lloyd, C.; Itoh, T.; Takeda, T.; Kim, D.-W.; et al. Human chromosome 11 DNA sequence and analysis including novel gene identification. Nature 2006, 440, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.R.; Mummidi, S.; Valente, A.J.; Yoshida, T.; Somanna, N.K.; Delafontaine, P.; Dinarello, C.A.; Chandrasekar, B. β2 adrenergic activation induces the expression of IL-18 binding protein, a potent inhibitor of isoproterenol induced cardiomyocyte hypertrophy in vitro and myocardial hypertrophy in vivo. J. Mol. Cell. Cardiol. 2012, 52, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, M.; Doble, R.; Bachmann, M.; Pfeilschifter, J.; Werfel, T.; Mühl, H. IL-27 Regulates IL-18 Binding Protein in Skin Resident Cells. PLoS ONE 2012, 7, e38751. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.; Paulukat, J.; Pfeilschifter, J.; Mühl, H. Molecular mechanisms of IL-18BP regulation in DLD-1 cells: Pivotal direct action of the STAT1/GAS axis on the promoter level. J. Cell. Mol. Med. 2009, 13, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, H.; Liu, Y.; Jiao, Y.; Yang, L.; Wang, X.; Yu, W.; Su, D.; Tian, J. Remifentanil upregulates hepatic IL-18 binding protein (IL-18BP) expression through transcriptional control. Lab. Investig. 2018, 98, 1588–1599. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.; Pfeilschifter, J.; Muhl, H. Epigenetic regulation by CpG methylation splits strong from retarded IFNgamma-induced IL-18BP in epithelial versus monocytic cells. Biochim. Biophys. Acta (BBA)—Gene Regul. Mech. 2018, 1861, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, Z.-X.; Lian, D.; Haig, A.; Bhattacharjee, R.N.; Jevnikar, A.M. IL-37 inhibits IL-18-induced tubular epithelial cell expression of pro-inflammatory cytokines and renal ischemia-reperfusion injury. Kidney Int. 2015, 87, 396–408. [Google Scholar] [CrossRef]
- Leng, J.; Yao, H.; Shen, J.; Wang, K.; Zhuo, G.; Wang, Z. Co-expression of IL-18 binding protein and IL-4 regulates Th1/Th2 cytokine response in murine collagen-induced arthritis. Acta Biochim. Biophys. Sin. 2008, 40, 116–124. [Google Scholar] [CrossRef]
- Chirathaworn, C.; Rianthavorn, P.; Wuttirattanakowit, N.; Poovorawan, Y. Serum IL-18 and IL-18BP Levels in Patients with Chikungunya Virus Infection. Viral Immunol. 2010, 23, 113–117. [Google Scholar] [CrossRef]
- Iannello, A.; Boulassel, M.; Samarani, S.; Tremblay, C.; Toma, E.; Routy, J.; Ahmad, A. HIV-1 Causes an Imbalance in the Production of Interleukin-18 and Its Natural Antagonist in HIV-Infected Individuals: Implications for Enhanced Viral Replication. J. Infect. Dis. 2010, 201, 608–617. [Google Scholar] [CrossRef]
- Fujita, K.; Ewing, C.M.; Isaacs, W.B.; Pavlovich, C.P. Immunomodulatory IL-18 binding protein is produced by prostate cancer cells and its levels in urine and serum correlate with tumor status. Int. J. Cancer 2011, 129, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Mazodier, K.; Marin, V.; Novick, D.; Farnarier, C.; Robitail, S.; Schleinitz, N.; Veit, V.; Paul, P.; Rubinstein, M.; Dinarello, C.A.; et al. Severe imbalance of IL-18/IL-18BP in patients with secondary hemophagocytic syndrome. Blood 2005, 106, 3483–3489. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Vizio, B.; Novarino, A.; Mauri, F.A.; Geuna, M.; Robino, C.; Brondino, G.; Prati, A.; Giacobino, A.; Campra, D.; et al. IL-18 Paradox in Pancreatic Carcinoma: Elevated Serum Levels of Free IL-18 are Correlated With Poor Survival. J. Immunother. 2009, 32, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Carbotti, G.; Barisione, G.; Orengo, A.M.; Brizzolara, A.; Airoldi, I.; Bagnoli, M.; Pinciroli, P.; Mezzanzanica, D.; Centurioni, M.G.; Fabbi, M.; et al. The IL-18 antagonist IL-18-binding protein is produced in the human ovarian cancer microenvironment. Clin. Cancer Res. 2013, 19, 4611–4620. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Liu, N.; Li, G.-F.; Meng, Q.-C.; Yao, J.-H.; Wang, N. IL-18 expression in clinical human pituitary adenoma. Technol. Health Care 2022, 30, 11–16. [Google Scholar] [CrossRef]
- Kaur, D.; Chachi, L.; Gomez, E.; Sylvius, N.; Brightling, C.E. Interleukin-18, IL-18 binding protein and IL-18 receptor expression in asthma: A hypothesis showing IL-18 promotes epithelial cell differentiation. Clin. Transl. Immunol. 2021, 10, e1301. [Google Scholar] [CrossRef]
- Hirooka, Y.; Nozaki, Y. Interleukin-18 in Inflammatory Kidney Disease. Front. Med. 2021, 8, 639103. [Google Scholar] [CrossRef]
- Liang, R.; Zheng, L.; Ji, T.; Zheng, J.; Liu, J.; Yuan, C.; Huang, Q.; Yang, M. Elevated serum free IL-18 in neuropsychiatric systemic lupus erythematosus patients with seizure disorders. Lupus 2022, 31, 187–193. [Google Scholar] [CrossRef]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 Regulates Both Th1 and Th2 Responses. Annu. Rev. Immunol. 2001, 19, 423–474. [Google Scholar] [CrossRef]
- Abbas, A.K.; Murphy, K.M.; Sher, A. Functional diversity of helper T lymphocytes. Nature 1996, 383, 787–793. [Google Scholar] [CrossRef]
- Kamradt, T.; Burmester, G.R. Cytokines and arthritis: Is the Th1/Th2 paradigm useful for understanding pathogenesis? J. Rheumatol. 1998, 25, 6–8. [Google Scholar]
- Shan, N.N.; Ji, X.B.; Wang, X.; Li, Y.; Liu, X.; Zhu, X.J.; Hou, M. In vitro recovery of Th1/Th2 balance in PBMCs from patients with immune thrombocytopenia through the actions of IL-18BPa/Fc. Thromb. Res. 2011, 128, e119–e124. [Google Scholar] [CrossRef]
- Novick, D.; Kim, S.; Kaplanski, G.; Dinarello, C.A. Interleukin-18, more than a Th1 cytokine. Semin. Immunol. 2013, 25, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Min, H.K.; Kim, S.; Lee, J.-Y.; Kim, K.-W.; Lee, S.-H.; Kim, H.-R. IL-18 binding protein suppresses IL-17-induced osteoclastogenesis and rectifies type 17 helper T cell / regulatory T cell imbalance in rheumatoid arthritis. J. Transl. Med. 2021, 19, 392. [Google Scholar] [CrossRef] [PubMed]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 is a unique cytokine that stimulates both Th1 and Th2 responses depending on its cytokine milieu. Cytokine Growth Factor Rev. 2001, 12, 53–72. [Google Scholar] [CrossRef]
- Nakamura, K.; Okamura, H.; Nagata, K.; Komatsu, T.; Tamura, T. Purification of a factor which provides a costimulatory signal for gamma interferon production. Infect. Immun. 1993, 61, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Fehniger, T.A.; Shah, M.H.; Turner, M.J.; VanDeusen, J.B.; Whitman, S.P.; Cooper, M.A.; Suzuki, K.; Wechser, M.; Goodsaid, F.; Caligiuri, M.A. Differential cytokine and chemokine gene expression by human NK cells following activation with IL-18 or IL-15 in combination with IL-12: Implications for the innate immune response. J. Immunol. 1999, 162, 4511–4520. [Google Scholar]
- Munder, M.; Mallo, M.; Eichmann, K.; Modolell, M. Murine macrophages secrete interferon γ upon combined stimulation with interleukin (IL)-12 and IL-18: A novel pathway of autocrine macrophage activation. J. Exp. Med. 1998, 187, 2103–2108. [Google Scholar] [CrossRef]
- Tsutsui, H.; Nakanishi, K.; Matsui, K.; Higashino, K.; Okamura, H.; Miyazawa, Y.; Kaneda, K. IFN-gamma-inducing factor up-regulates Fas ligand-mediated cytotoxic activity of murine natural killer cell clones. J. Immunol. 1996, 157, 3967–3973. [Google Scholar]
- Dinarello, C.A. IL-18: A TH1 -inducing, proinflammatory cytokine and new member of the IL-1 family. J. Allergy Clin. Immunol. 1999, 103, 11–24. [Google Scholar] [CrossRef]
- Swain, S.L. Interleukin 18: Tipping the balance towards a T helper cell 1 response. J. Exp. Med. 2001, 194, F11–F14. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Yoshimoto, T.; Maruyama, H.; Tegoshi, T.; Ohta, N.; Arizono, N.; Nakanishi, K. IL-18 with IL-2 protects against Strongyloides venezuelensis infection by activating mucosal mast cell–dependent type 2 innate immunity. J. Exp. Med. 2005, 202, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Miyazaki, N.; Oashi, K.; Teramoto, S.; Shiratori, M.; Hashimoto, M.; Ohmichi, M.; Abe, S. IL-18 might reflect disease activity in mild and moderate asthma exacerbation. J. Allergy Clin. Immunol. 2001, 107, 331–336. [Google Scholar] [CrossRef]
- Hoshino, T.; Wiltrout, R.H.; A Young, H. IL-18 is a potent coinducer of IL-13 in NK and T cells: A new potential role for IL-18 in modulating the immune response. J. Immunol. 1999, 162, 5070–5077. [Google Scholar]
- Yoshimoto, T.; Mizutani, H.; Tsutsui, H.; Noben-Trauth, N.; Yamanaka, K.-I.; Tanaka, M.; Izumi, S.; Okamura, H.; Paul, W.E.; Nakanishi, K. IL-18 induction of IgE: Dependence on CD4+ T cells, IL-4 and STAT6. Nat. Immunol. 2000, 1, 132–137. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Tsutsui, H.; Tominaga, K.; Hoshino, K.; Okamura, H.; Akira, S.; Akira, S.; Paul, W.E.; Nakanishi, K. IL-18, although antiallergic when administered with IL-12, stimulates IL-4 and histamine release by basophils. Proc. Natl. Acad. Sci. USA 1999, 96, 13962–13966. [Google Scholar] [CrossRef] [PubMed]
- Sawada, M.; Kawayama, T.; Imaoka, H.; Sakazaki, Y.; Oda, H.; Takenaka, S.-I.; Kaku, Y.; Azuma, K.; Tajiri, M.; Edakuni, N.; et al. IL-18 Induces Airway Hyperresponsiveness and Pulmonary Inflammation via CD4+ T Cell and IL-13. PLoS ONE 2013, 8, e54623. [Google Scholar] [CrossRef] [PubMed]
- Oda, H.; Kawayama, T.; Imaoka, H.; Sakazaki, Y.; Kaku, Y.; Okamoto, M.; Kitasato, Y.; Edakuni, N.; Takenaka, S.-I.; Yoshida, M.; et al. Interleukin-18 expression, CD8+ T cells, and eosinophils in lungs of nonsmokers with fatal asthma. Ann. Allergy Asthma Immunol. 2014, 112, 23–28.e1. [Google Scholar] [CrossRef] [PubMed]
- Conforti-Andreoni, C.; Spreafico, R.; Qian, H.L.; Riteau, N.; Ryffel, B.; Ricciardi-Castagnoli, P.; Mortellaro, A. Uric Acid-Driven Th17 Differentiation Requires Inflammasome-Derived IL-1 and IL-18. J. Immunol. 2011, 187, 5842–5850. [Google Scholar] [CrossRef]
- Mills, K.H.G.; Dungan, L.S.; Jones, S.A.; Harris, J. The role of inflammasome-derived IL-1 in driving IL-17 responses. J. Leukoc. Biol. 2013, 93, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.M.; Li, Q.W.; Jiang, G.X.; Liu, J.S.; Cheng, Q. IL-18 induced IL-23/IL-17 expression impairs Abeta clearance in cultured THP-1 and BV2 cells. Cytokine 2019, 119, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, M.N.; Shukla, P.; Kakaji, M.; Tyagi, A.M.; Srivastava, K.; Shukla, M.; Dixit, M.; Kureel, J.; Gupta, S.; Singh, D. IL-18BP is decreased in osteoporotic women: Prevents Inflammasome mediated IL-18 activation and reduces Th17 differentiation. Sci. Rep. 2016, 6, 33680. [Google Scholar] [CrossRef] [PubMed]
- Born, T.L.; Morrison, L.A.; Esteban, D.J.; Vandenbos, T.; Thebeau, L.G.; Chen, N.; Spriggs, M.K.; Sims, J.; Buller, R.M.L. A Poxvirus Protein That Binds to and Inactivates IL-18, and Inhibits NK Cell Response. J. Immunol. 2000, 164, 3246–3254. [Google Scholar] [CrossRef] [PubMed]
- Reading, P.C.; Smith, G.L. Vaccinia Virus Interleukin-18-Binding Protein Promotes Virulence by Reducing Gamma Interferon Production and Natural Killer and T-Cell Activity. J. Virol. 2003, 77, 9960–9968. [Google Scholar] [CrossRef]
- Smith, V.P.; Bryant, N.A.; Alcamı, A. Ectromelia, vaccinia and cowpox viruses encode secreted interleukin-18-binding proteins. J. Gen. Virol. 2000, 81, 1223–1230. [Google Scholar] [CrossRef]
- Meng, X.; Leman, M.; Xiang, Y. Variola virus IL-18 binding protein interacts with three human IL-18 residues that are part of a binding site for human IL-18 receptor alpha subunit. Virology 2007, 358, 211–220. [Google Scholar] [CrossRef]
- Krumm, B.; Meng, X.; Wang, Z.; Xiang, Y.; Deng, J. A Unique Bivalent Binding and Inhibition Mechanism by the Yatapoxvirus Interleukin 18 Binding Protein. PLoS Pathog. 2012, 8, e1002876. [Google Scholar] [CrossRef]
- Nazarian, S.H.; Rahman, M.M.; Werden, S.J.; Villeneuve, D.; Meng, X.; Brunetti, C.; Valeriano, C.; Wong, C.; Singh, R.; Barrett, J.W.; et al. Yaba Monkey Tumor Virus Encodes a Functional Inhibitor of Interleukin-18. J. Virol. 2008, 82, 522–528. [Google Scholar] [CrossRef][Green Version]
- Xiang, Y.; Moss, B. IL-18 binding and inhibition of interferon γ induction by human poxvirus-encoded proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11537–11542. [Google Scholar] [CrossRef]
- Xiang, Y.; Moss, B. Molluscum Contagiosum Virus Interleukin-18 (IL-18) Binding Protein Is Secreted as a Full-Length Form That Binds Cell Surface Glycosaminoglycans through the C-Terminal Tail and a Furin-Cleaved Form with Only the IL-18 Binding Domain. J. Virol. 2003, 77, 2623–2630. [Google Scholar] [CrossRef]
- Falivene, J.; Zajac, M.P.D.M.; Pascutti, M.F.; Rodriguez, A.M.; Maeto, C.; Perdiguero, B.; Gomez, C.E.; Esteban, M.; Calamante, G.; Gherardi, M.M. Improving the MVA Vaccine Potential by Deleting the Viral Gene Coding for the IL-18 Binding Protein. PLoS ONE 2012, 7, e32220. [Google Scholar] [CrossRef]
- Wawrocki, S.; Seweryn, M.; Kielnierowski, G.; Rudnicka, W.; Wlodarczyk, M.; Druszczynska, M. IL-18/IL-37/IP-10 signalling complex as a potential biomarker for discriminating active and latent TB. PLoS ONE 2019, 14, e0225556. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.C.; Jung, J.Y.; Nau, G.J. Interferon-gamma, tumor necrosis factor, and interleukin-18 cooperate to control growth of Mycobacterium tuberculosis in human macrophages. Cytokine 2012, 60, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Wawrocki, S.; Seweryn, M.; Kielnierowski, G.; Rudnicka, W.; Druszczynska, M. IL-18 and related function proteins associated with tuberculosis severity and screening for active TB among patients with non-mycobacterial community-acquired pneumonia (CAP). Saudi J. Biol. Sci. 2020, 27, 3035–3045. [Google Scholar] [CrossRef]
- Wawrocki, S.; Kielnierowski, G.; Rudnicka, W.; Seweryn, M.; Druszczynska, M. Interleukin-18, functional IL-18 receptor and IL-18 binding protein expression in active and latent tuberculosis. Pathogens 2020, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Fabbi, M.; Carbotti, G.; Ferrini, S. Context-dependent role of IL-18 in cancer biology and counter-regulation by IL-18BP. J. Leukoc. Biol. 2015, 97, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Dixon, K.O.; Kuchroo, V.K. IL-18: Throwing off the shackles to boost anti-tumor immunity. Cell Res. 2020, 30, 831–832. [Google Scholar] [CrossRef]
- Nakamura, K.; Bald, T.; Smyth, M.J. Cancer-killing, decoy-resistant interleukin-18. Immunol. Cell Biol. 2020, 98, 434–436. [Google Scholar] [CrossRef]
- Abrahamsson, A.; Rzepecka, A.; Dabrosin, C. Equal Pro-inflammatory Profiles of CCLs, CXCLs, and Matrix Metalloproteinases in the Extracellular Microenvironment In Vivo in Human Dense Breast Tissue and Breast Cancer. Front. Immunol. 2017, 8, 1994. [Google Scholar] [CrossRef]
- Muhl, H.; Bachmann, M. IL-18/IL-18BP and IL-22/IL-22BP: Two interrelated couples with therapeutic potential. Cell. Signal. 2019, 63, 109388. [Google Scholar] [CrossRef] [PubMed]
- Tarhini, A.A.; Millward, M.; Mainwaring, P.; Kefford, R.; Logan, T.; Pavlick, A.; Kathman, S.J.; Laubscher, K.H.; Dar, M.M.; Kirkwood, J.M. A phase 2, randomized study of SB-485232, rhIL-18, in patients with previously untreated metastatic melanoma. Cancer 2009, 115, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Simpkins, F.; Flores, A.; Chu, C.; Berek, J.S.; Lucci, J.; Murray, S.; Bauman, J.; Struemper, H.; Germaschewski, F.; Jonak, Z.; et al. Chemoimmunotherapy Using Pegylated Liposomal Doxorubicin and Interleukin-18 in Recurrent Ovarian Cancer: A Phase I Dose-Escalation StudyIL-18 Plus PLD in Ovarian Cancer. Cancer Immunol. Res. 2013, 1, 168–178. [Google Scholar] [CrossRef]
- Zhou, T.; Damsky, W.; Weizman, O.-E.; McGeary, M.K.; Hartmann, K.P.; Rosen, C.E.; Fischer, S.; Jackson, R.; Flavell, R.A.; Wang, J.; et al. IL-18BP is a secreted immune checkpoint and barrier to IL-18 immunotherapy. Nature 2020, 583, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Li, J.; Lin, W.; Cen, W.; Zhang, W.; Zhu, Z.; Lu, B.; Yu, J. Inhibition of heat shock protein 90 alleviates cholestatic liver injury by decreasing IL-1beta and IL-18 expression. Exp. Ther. Med. 2021, 21, 241. [Google Scholar] [CrossRef]
- Dokumacioğlu, E.; Iskender, H.; Kapakin KA, T.; Yenice, G.; Mokthare, B.; Bolat, I.; Hayirli, A. Effect of betulinic acid administration on TLR-9/NF-ΚB/IL-18 levels in experimentalliver injury. Turk. J. Med. Sci. 2021, 51, 1544–1553. [Google Scholar] [CrossRef]
- Endre, Z.H.; Westhuyzen, J. Early detection of acute kidney injury: Emerging new biomarkers (Review Article). Nephrology 2008, 13, 91–98. [Google Scholar] [CrossRef]
- Washburn, K.K.; Zappitelli, M.; Arikan, A.A.; Loftis, L.; Yalavarthy, R.; Parikh, C.R.; Edelstein, C.L.; Goldstein, S.L. Urinary interleukin-18 is an acute kidney injury biomarker in critically ill children. Nephrol. Dial. Transplant. 2008, 23, 566–572. [Google Scholar] [CrossRef]
- Belkaya, S.; Michailidis, E.; Korol, C.B.; Kabbani, M.; Cobat, A.; Bastard, P.; Lee, Y.S.; Hernandez, N.; Drutman, S.; De Jong, Y.P.; et al. Inherited IL-18BP deficiency in human fulminant viral hepatitis. J. Exp. Med. 2019, 216, 1777–1790. [Google Scholar] [CrossRef]
- Novick, D.; Elbirt, D.; Dinarello, C.A.; Rubinstein, M.; Sthoeger, Z.M. Interleukin-18 Binding Protein in the Sera of Patients with Wegener’s Granulomatosis. J. Clin. Immunol. 2009, 29, 38–45. [Google Scholar] [CrossRef]
- Novick, D.; Elbirt, D.; Miller, G.; Dinarello, C.A.; Rubinstein, M.; Sthoeger, Z.M. High circulating levels of free interleukin-18 in patients with active SLE in the presence of elevated levels of interleukin-18 binding protein. J. Autoimmun. 2010, 34, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Banda, N.K.; Vondracek, A.; Kraus, D.; Dinarello, C.A.; Kim, S.-H.; Bendele, A.; Senaldi, G.; Arend, W.P. Mechanisms of Inhibition of Collagen-Induced Arthritis by Murine IL-18 Binding Protein. J. Immunol. 2003, 170, 2100–2105. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Muraoka, M.; Yokoyama, T.; Toma, T.; Kanegane, H.; Yachie, A. Cytokine profiles in children with primary Epstein-Barr virus infection. Pediatr. Blood Cancer 2013, 60, E46–E48. [Google Scholar] [CrossRef]
- Shimizu, M. Macrophage activation syndrome in systemic juvenile idiopathic arthritis. Immunol. Med. 2021, 44, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Maeno, N.; Takei, S.; Imanaka, H.; Yamamoto, K.; Kuriwaki, K.; Kawano, Y.; Oda, H. Increased interleukin-18 expression in bone marrow of a patient with systemic juvenile idiopathic arthritis and unrecognized macrophage-activation syndrome. Arthritis Care Res. 2004, 50, 1935–1938. [Google Scholar] [CrossRef] [PubMed]
- Emmenegger, U.; Reimers, A.; Frey, U.; Fux, C.H.; Bihl, F.; Semela, D.; Cottagnoud, P.; Cerny, A.; Spaethe, P.; Neftel, K.A. Reactive macrophage activation syndrome: A simple screening strategy and its potential in early treatment initiation. Swiss Med. Wkly. 2002, 132, 230–236. [Google Scholar] [PubMed]
- Nold-Petry, C.A.; Lehrnbecher, T.; Jarisch, A.; Schwabe, D.; Pfeilschifter, J.M.; Muhl, H.; Nold, M.F. Failure of interferon gamma to induce the anti-inflammatory interleukin 18 binding protein in familial hemophagocytosis. PLoS ONE 2010, 5, e8663. [Google Scholar] [CrossRef]
- Honda, K.; Ohga, S.; Takada, H.; Nomura, A.; Ohshima, K.; Kinukawa, N.; Mizuno, Y.; Hara, T. Neuron-specific enolase in hemophagocytic lymphohistiocytosis: A potential indicator for macrophage activation? Int. J. Hematol. 2000, 72, 55–60. [Google Scholar]
- Fitzgerald, A.A.; LeClercq, S.A.; Yan, A.; Homik, J.E.; Dinarello, C.A. Rapid responses to anakinra in patients with refractory adult-onset Still’s disease. Arthritis Care Res. 2005, 52, 1794–1803. [Google Scholar] [CrossRef]
- Larroche, C.; Mouthon, L. Pathogenesis of hemophagocytic syndrome (HPS). Autoimmun. Rev. 2004, 3, 69–75. [Google Scholar] [CrossRef]
- Carter, S.J.; Tattersall, R.S.; Ramanan, A.V. Macrophage activation syndrome in adults: Recent advances in pathophysiology, diagnosis and treatment. Rheumatology 2019, 58, 5–17. [Google Scholar] [CrossRef]
- Giacomelli, R.; Ruscitti, P.; Shoenfeld, Y. A comprehensive review on adult onset Still’s disease. J. Autoimmun. 2018, 93, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Yuan, Y.; Wang, Y.; Jiang, J.; Ye, Z.; Liu, T.; Jiang, Z.; Zhao, L. Clinical characteristics of macrophage activation syndrome in adult-onset Still’s disease. Clin. Exp. Rheumatol. 2021, 39 (Suppl. 132), 59–66. [Google Scholar] [CrossRef] [PubMed]
- Yasin, S.; Solomon, K.; Canna, S.W.; Girard-Guyonvarc’H, C.; Gabay, C.; Schiffrin, E.; Sleight, A.; Grom, A.A.; Schulert, G.S. IL-18 as therapeutic target in a patient with resistant systemic juvenile idiopathic arthritis and recurrent macrophage activation syndrome. Rheumatology 2020, 59, 442–445. [Google Scholar] [CrossRef]
- Behrens, E.M.; Beukelman, T.; Paessler, M.; Cron, R.Q. Occult macrophage activation syndrome in patients with systemic juvenile idiopathic arthritis. J. Rheumatol. 2007, 34, 1133–1138. [Google Scholar] [PubMed]
- Crayne, C.B.; Albeituni, S.; Nichols, K.E.; Cron, R.Q. The Immunology of Macrophage Activation Syndrome. Front. Immunol. 2019, 10, 119. [Google Scholar] [CrossRef]
- Grom, A.A.; Horne, A.; De Benedetti, F. Macrophage activation syndrome in the era of biologic therapy. Nat. Rev. Rheumatol. 2016, 12, 259–268. [Google Scholar] [CrossRef]
- Schulert, G.S.; Grom, A.A. Pathogenesis of Macrophage Activation Syndrome and Potential for Cytokine- Directed Therapies. Annu. Rev. Med. 2015, 66, 145–159. [Google Scholar] [CrossRef]
- Kawashima, M.; Yamamura, M.; Taniai, M.; Yamauchi, H.; Tanimoto, T.; Kurimoto, M.; Miyawaki, S.; Amano, T.; Takeuchi, T.; Makino, H. Levels of interleukin-18 and its binding inhibitors in the blood circulation of patients with adult-onset Still’s disease. Arthritis Rheum. 2001, 44, 550–560. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, H.-S.; Lee, J.S.; Kim, J.J.; Jung, K.-H.; Park, Y.-W.; Yoo, D.-H. Natural Killer Cell Cytolytic Function in Korean Patients with Adult-onset Still’s Disease. J. Rheumatol. 2012, 39, 2000–2007. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Terajima, H.; Harigai, M.; Hara, M.; Kamatani, N. Interleukin-18 as a novel diagnostic marker and indicator of disease severity in adult-onset Still’s disease. Arthritis Care Res. 2001, 44, 1716–1717. [Google Scholar] [CrossRef]
- Colafrancesco, S.; Priori, R.; Alessandri, C.; Perricone, C.; Pendolino, M.; Picarelli, G.; Valesini, G. IL-18 Serum Level in Adult Onset Still’s Disease: A Marker of Disease Activity. Int. J. Inflamm. 2012, 2012, 156890. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Rech, J.; Brown, M.; Allali, D.; Roux-Lombard, P.; Spertini, F.; Schiffrin, E.J.; Schett, G.; Manger, B.; Bas, S.; et al. Elevated serum levels of free interleukin-18 in adult-onset Still’s disease. Rheumatology 2016, 55, 2237–2247. [Google Scholar] [CrossRef] [PubMed]
- Grom, A.A. Macrophage activation syndrome and reactive hemophagocytic lymphohistiocytosis: The same entities? Curr. Opin. Rheumatol. 2003, 15, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Grom, A.A.; Mellins, E.D. Macrophage activation syndrome: Advances towards understanding pathogenesis. Curr. Opin. Rheumatol. 2010, 22, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Grom, A.A.; Villanueva, J.; Lee, S.; Goldmuntz, E.A.; Passo, M.H.; Filipovich, A. Natural killer cell dysfunction in patients with systemic-onset juvenile rheumatoid arthritis and macrophage activation syndrome. J. Pediatr. 2003, 142, 292–296. [Google Scholar] [CrossRef]
- Villanueva, J.; Lee, S.; Giannini, E.H.; Graham, T.B.; Passo, M.H.; Filipovich, A.; Grom, A.A. Natural killer cell dysfunction is a distinguishing feature of systemic onset juvenile rheumatoid arthritis and macrophage activation syndrome. Arthritis Res. Ther. 2005, 7, R30–R37. [Google Scholar] [CrossRef] [PubMed]
- Janka, G. Familial and Acquired Hemophagocytic Lymphohistiocytosis. Annu. Rev. Med. 2012, 63, 233–246. [Google Scholar] [CrossRef]
- Chen, O.; Shan, N.; Zhu, X.; Wang, Y.; Ren, P.; Wei, D.E.; Sun, R. The imbalance of IL-18/IL-18BP in patients with systemic juvenile idiopathic arthritis. Acta Biochim. Biophys. Sin. 2013, 45, 339–341. [Google Scholar] [CrossRef][Green Version]
- Weiss, E.S.; Girard-Guyonvarc’h, C.; Holzinger, D.; de Jesus, A.A.; Tariq, Z.; Picarsic, J.; Schiffrin, E.J.; Foell, D.; Grom, A.A.; Ammann, S.; et al. Interleukin-18 diagnostically distinguishes and pathogenically promotes human and murine macrophage activation syndrome. Blood 2018, 131, 1442–1455. [Google Scholar] [CrossRef]
- Jelušić, M.; Lukić, I.K.; Tambić-Bukovac, L.; Dubravčić, K.; Malčić, I.; Rudan, I.; Batinić, D. Interleukin-18 as a mediator of systemic juvenile idiopathic arthritis. Clin. Rheumatol. 2007, 26, 1332–1334. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lotito AP, N.; Campa, A.; Silva, C.A.; Kiss, M.H.; Mello, S.B. Interleukin 18 as a marker of disease activity and severity in patients with juvenile idiopathic arthritis. J. Rheumatol. 2007, 34, 823–830. [Google Scholar] [PubMed]
- Kim, H.A.; An, J.M.; Nam, J.Y.; Jeon, J.Y.; Suh, C.H. Serum S100A8/A9, but not follistatin-like protein 1 and interleukin 18, may be a useful biomarker of disease activity in adult-onset Still’s disease. J. Rheumatol. 2012, 39, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Nakagishi, Y.; Inoue, N.; Mizuta, M.; Ko, G.; Saikawa, Y.; Kubota, T.; Yamasaki, Y.; Takei, S.; Yachie, A. Interleukin-18 for predicting the development of macrophage activation syndrome in systemic juvenile idiopathic arthritis. Clin. Immunol. 2015, 160, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Put, K.; Avau, A.; Brisse, E.; Mitera, T.; Put, S.; Proost, P.; Bader-Meunier, B.; Westhovens, R.; van den Eynde, B.J.; Orabona, C.; et al. Cytokines in systemic juvenile idiopathic arthritis and haemophagocytic lymphohistiocytosis: Tipping the balance between interleukin-18 and interferon-gamma. Rheumatology 2015, 54, 1507–1517. [Google Scholar]
- Inoue, N.; Shimizu, M.; Tsunoda, S.; Kawano, M.; Matsumura, M.; Yachie, A. Cytokine profile in adult-onset Still’s disease: Comparison with systemic juvenile idiopathic arthritis. Clin. Immunol. 2016, 169, 8–13. [Google Scholar] [CrossRef]
- Tomaras, S.; Goetzke, C.; Kallinich, T.; Feist, E. Adult-Onset Still’s Disease: Clinical Aspects and Therapeutic Approach. J. Clin. Med. 2021, 10, 733. [Google Scholar] [CrossRef]
- Tang, S.; Li, S.; Zheng, S.; Ding, Y.; Zhu, D.; Sun, C.; Hu, Y.; Qiao, J.; Fang, H. Understanding of cytokines and targeted therapy in macrophage activation syndrome. Semin. Arthritis Rheum. 2021, 51, 198–210. [Google Scholar] [CrossRef]
- Gabay, C.; Fautrel, B.; Rech, J.; Spertini, F.; Feist, E.; Kötter, I.; Hachulla, E.; Morel, J.; Schaeverbeke, T.; Hamidou, M.A.; et al. Open-label, multicentre, dose-escalating phase II clinical trial on the safety and efficacy of tadekinig alfa (IL-18BP) in adult-onset Still’s disease. Ann. Rheum. Dis. 2018, 77, 840–847. [Google Scholar] [CrossRef]
- Kiltz, U.; Kiefer, D.; Braun, J.; Schiffrin, E.J.; Girard-Guyonvarc’H, C.; Gabay, C. Prolonged treatment with Tadekinig alfa in adult-onset Still’s disease. Ann. Rheum. Dis. 2020, 79, e10. [Google Scholar] [CrossRef]
- Canna, S.W.; Girard, C.; Malle, L.; de Jesus, A.; Romberg, N.; Kelsen, J.; Surrey, L.F.; Russo, P.; Sleight, A.; Schiffrin, E.; et al. Life-threatening NLRC4-associated hyperinflammation successfully treated with IL-18 inhibition. J. Allergy Clin. Immunol. 2017, 139, 1698–1701. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.L.; Chen, Y.M.; Hsieh, C.W.; Chen, H.H.; Lee, H.C.; Hung, W.T.; Tang, K.-Y.; Chen, D.Y. Upregulation of circulating microRNA-134 in adult-onset Still’s disease and its use as potential biomarker. Sci. Rep. 2017, 7, 4214. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Canna, S.W.; De Jesus, A.A.; Gouni, S.; Brooks, S.R.; Marrero, B.; Liu, Y.; DiMattia, M.A.; Zaal, K.J.; Sanchez, G.A.M.; Kim, H.; et al. An activating NLRC4 inflammasome mutation causes autoinflammation with recurrent macrophage activation syndrome. Nat. Genet. 2014, 46, 1140–1146. [Google Scholar] [CrossRef]
- Romberg, N.; Al Moussawi, K.; Nelson-Williams, C.; Stiegler, A.L.; Loring, E.; Choi, M.; Overton, J.; Meffre, E.; Khokha, M.K.; Huttner, A.J.; et al. Mutation of NLRC4 causes a syndrome of enterocolitis and autoinflammation. Nat. Genet. 2014, 46, 1135–1139. [Google Scholar] [CrossRef]
- Dinarello, C.A. Targeting interleukin 18 with interleukin 18 binding protein. Ann. Rheum. Dis. 2000, 59 (Suppl. 1), i17–i20. [Google Scholar] [CrossRef]
- Vasilev, G.; Manolova, I.; Ivanova, M.; Stanilov, I.; Miteva, L.; Stanilova, S. The role of IL-18 in addition to Th17 cytokines in rheumatoid arthritis development and treatment in women. Sci. Rep. 2021, 11, 15391. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.-H.; Yuan, F.-L.; Wang, S.-J.; Xu, H.-Y.; Li, C.-W.; Tong, X. Association of interleukin-18 and asthma. Inflammation 2017, 40, 324–327. [Google Scholar] [CrossRef]
- Xiang, M.; Feng, Y.; Wang, Y.; Wang, J.; Zhang, Z.; Liang, J.; Xu, J. Correlation between circulating interleukin-18 level and systemic lupus erythematosus: A meta-analysis. Sci. Rep. 2021, 11, 4707. [Google Scholar] [CrossRef]
- Forouzandeh, M.; Besen, J.; Keane, R.W.; Vaccari, J.P.D.R. The Inflammasome Signaling Proteins ASC and IL-18 as Biomarkers of Psoriasis. Front. Pharmacol. 2020, 11, 1238. [Google Scholar] [CrossRef]
- Fukaura, H.; Kikuchi, S. [IL-18 in multiple sclerosis]. Nihon Rinsho 2003, 61, 1416–1421. [Google Scholar]
- Franke, E.I.; Vanderbrink, B.A.; Hile, K.L.; Zhang, H.; Cain, A.; Matsui, F.; Meldrum, K.K. Renal IL-18 Production Is Macrophage Independent During Obstructive Injury. PLoS ONE 2012, 7, e47417. [Google Scholar] [CrossRef]
- Williams, M.; O’Callaghan, A.; Corr, S.C. IL-33 and IL-18 in Inflammatory Bowel Disease Etiology and Microbial Interactions. Front. Immunol. 2019, 10, 1091. [Google Scholar] [CrossRef]
- Su, Y.; Zhao, H. Predisposition of Inflammatory Bowel Disease Is Influenced by IL-8, IL-10, and IL-18 Polymorphisms: A Meta-Analysis. Int. Arch. Allergy Immunol. 2020, 181, 799–806. [Google Scholar] [CrossRef]
- Karaba, A.H.; Figueroa, A.; Werbel, W.A.; Dioverti, M.V.; Steinke, S.M.; Ray, S.C.; Cox, A.L.; Avery, R.K. Interleukin-18 and tumor necrosis factor-α are elevated in solid organ transplant recipients with possible cytomegalovirus end-organ disease. Transpl. Infect. Dis. 2021, 23, e13682. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, J.; Liu, B.; Yuan, S.; Shou, D.; Wen, L.; Wu, X.; Gong, W. Role of IL-18 in transplant biology. Eur. Cytokine Netw. 2018, 29, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, J.E.; Lee, J.Y.; Cho, B.K.; Lee, W.J.; Kim, T.; Yoon, D.; Cho, D. Increased expression of IL-18 in cutaneous graft-versus-host disease. Immunol. Lett. 2004, 95, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Stone, D.L.; Ombrello, A.; Arostegui, J.I.; Schneider, C.; Dang, V.; de Jesus, A.; Girard-Guyonvarc’H, C.; Gabay, C.; Lee, W.; Chae, J.J.; et al. Excess Serum Interleukin-18 Distinguishes Patients With Pathogenic Mutations in PSTPIP1. Arthritis Rheumatol. 2022, 74, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Wise, C.A.; Gillum, J.D.; Seidman, C.E.; Lindor, N.M.; Veile, R.; Bashiardes, S.; Lovett, M. Mutations in CD2BP1 disrupt binding to PTP PEST and are responsible for PAPA syndrome, an autoinflammatory disorder. Hum. Mol. Genet. 2002, 11, 961–969. [Google Scholar] [CrossRef]
- Shoham, N.G.; Centola, M.; Mansfield, E.; Hull, K.M.; Wood, G.; Wise, C.A.; Kastner, D.L. Pyrin binds the PSTPIP1/CD2BP1 protein, defining familial Mediterranean fever and PAPA syndrome as disorders in the same pathway. Proc. Natl. Acad. Sci. USA 2003, 100, 13501–13506. [Google Scholar] [CrossRef]
- Wlodek, E.; Kirkpatrick, R.B.; Andrews, S.; Noble, R.; Schroyer, R.; Scott, J.; Watson, C.J.E.; Clatworthy, M.; Harrison, E.M.; Wigmore, S.J.; et al. A pilot study evaluating GSK1070806 inhibition of interleukin-18 in renal transplant delayed graft function. PLoS ONE 2021, 16, e0247972. [Google Scholar] [CrossRef] [PubMed]
- Galozzi, P.; Bindoli, S.; Doria, A.; Sfriso, P. Progress in Biological Therapies for Adult-Onset Still’s Disease. Biol. Targets Ther. 2022, 16, 21–34. [Google Scholar] [CrossRef]
- Detry, S.; Andries, J.; Bloch, Y.; Gabay, C.; Clancy, D.M.; Savvides, S.N. Structural basis of human IL-18 sequestration by the decoy receptor IL-18 binding protein (IL-18BP) in inflammation and tumor immunity. J. Biol. Chem. 2022, 298, 101808. [Google Scholar] [CrossRef] [PubMed]
- Flament, H.; Rouland, M.; Beaudoin, L.; Toubal, A.; Bertrand, L.; Lebourgeois, S.; Rousseau, C.; Soulard, P.; Gouda, Z.; Cagninacci, L.; et al. Outcome of SARS-CoV-2 infection is linked to MAIT cell activation and cytotoxicity. Nat. Immunol. 2021, 22, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef]
- Rodriguez-Smith, J.J.; Verweyen, E.L.; Clay, G.M.; Esteban, Y.M.; de Loizaga, S.R.; Baker, E.J.; Do, T.; Dhakal, S.; Lang, S.M.; Grom, A.A.; et al. Inflammatory biomarkers in COVID-19-associated multisystem inflammatory syndrome in children, Kawasaki disease, and macrophage activation syndrome: A cohort study. Lancet Rheumatol. 2021, 3, e574–e584. [Google Scholar] [CrossRef]
- Huang, W.; Li, M.; Luo, G.; Wu, X.; Su, B.; Zhao, L.; Zhang, S.; Chen, X.; Jia, M.; Zhu, J.; et al. The inflammatory factors associated with disease severity to predict COVID-19 progression. J. Immunol. 2021, 206, 1597–1608. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-alpha and IFN-gamma Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef]
- Wilson, J.G.; Simpson, L.J.; Ferreira, A.-M.; Rustagi, A.; Roque, J.; Asuni, A.; Ranganath, T.; Grant, P.M.; Subramanian, A.; Rosenberg-Hasson, Y.; et al. Cytokine profile in plasma of severe COVID-19 does not differ from ARDS and sepsis. JCI Insight 2020, 5, 140289. [Google Scholar] [CrossRef]
- Gerasimenko, V.J.; Petersen, O.H.; Gerasimenko, O.V. SARS-CoV-2 S Protein Subunit 1 Elicits Ca2+ Influx - Dependent Ca2+ Signals in Pancreatic Stellate Cells and Macrophages In Situ. Function 2022, 3, zqac002. [Google Scholar] [CrossRef]
- Jamilloux, Y.; Henry, T.; Belot, A.; Viel, S.; Fauter, M.; El Jammal, T.; Walzer, T.; François, B.; Sève, P. Should we stimulate or suppress immune responses in COVID-19? Cytokine and anti-cytokine interventions. Autoimmun. Rev. 2020, 19, 102567. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition/Disease | Clinical Phase | Clinical Trial Status | No. of Participants | Type of Intervention or Treatment | NCT Number |
|---|---|---|---|---|---|
| Still’s Disease, Adult-Onset | Phase II | Completed | 23 participants | Biological: Tadekinig alfa (recombinant human IL-18 binding protein) | NCT02398435 |
| XIAP Deficiency, NLRC4-MAS | Phase III | Recruiting | 10 participants | Drug: Tadekinig alfa | NCT03512314 |
| XIAP Deficiency, NLRC4-MAS | Phase III | Recruiting | 10 participants | Drug: Tadekinig alfa | NCT03113760 |
| Inflammatory Bowel Diseases | Phase I | Completed | 78 participants | Drug: GSK1070806 (block IL-18) | NCT01035645 |
| Kidney Transplantation (Status Post) | Phase II | Terminated * | 7 participants | Drug: GSK1070806 + others | NCT02723786 |
| Melanoma | Phase II | Completed | 64 participants | Drug: SB-485232 (rhIL-18) | NCT00107718 |
| Dermatitis, Atopic | Phase I | Recruiting | 48 participants | Drug: GSK1070806 | NCT04975438 |
| Crohn Disease | Phase I Phase II | Completed | 5 participants | Drug: GSK1070806 | NCT03681067 |
| Behcet’s Disease | Phase II | Unknown # | 12 participants | Drug: GSK1070806 | NCT03522662 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.Y.; Hisham, Y.; Shin, H.M.; Yeom, S.C.; Kim, S. Interleukin-18 Binding Protein in Immune Regulation and Autoimmune Diseases. Biomedicines 2022, 10, 1750. https://doi.org/10.3390/biomedicines10071750
Park SY, Hisham Y, Shin HM, Yeom SC, Kim S. Interleukin-18 Binding Protein in Immune Regulation and Autoimmune Diseases. Biomedicines. 2022; 10(7):1750. https://doi.org/10.3390/biomedicines10071750
Chicago/Turabian StylePark, Seung Yong, Yasmin Hisham, Hyun Mu Shin, Su Cheong Yeom, and Soohyun Kim. 2022. "Interleukin-18 Binding Protein in Immune Regulation and Autoimmune Diseases" Biomedicines 10, no. 7: 1750. https://doi.org/10.3390/biomedicines10071750
APA StylePark, S. Y., Hisham, Y., Shin, H. M., Yeom, S. C., & Kim, S. (2022). Interleukin-18 Binding Protein in Immune Regulation and Autoimmune Diseases. Biomedicines, 10(7), 1750. https://doi.org/10.3390/biomedicines10071750