
Paneth Cells Regulate Lymphangiogenesis under Control of Microbial Signals during Experimental Portal Hypertension

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Genotyping of Mice
2.3. RNA Isolation
2.4. mRNA-Sequencing in Small Intestinal Tissue
2.5. RT2 Profiler qPCR Array and Gene Expression Quantification by Real-Time PCR
2.6. Immunohistochemistry
2.7. Isolation of Crypts
2.8. Development and Passaging of Intestinal Organoids
2.9. Stimulation of Intestinal Organoids with PAMPs
2.10. Whole Mount Lysozyme Immunofluorescence of Organoids and Imaging
2.11. Lymphatic Endothelial Cell Tube Formation Assay
2.12. Wound Healing Assay
2.13. Proteomic Analysis of Conditioned Media
2.14. Statistical Analysis
3. Results
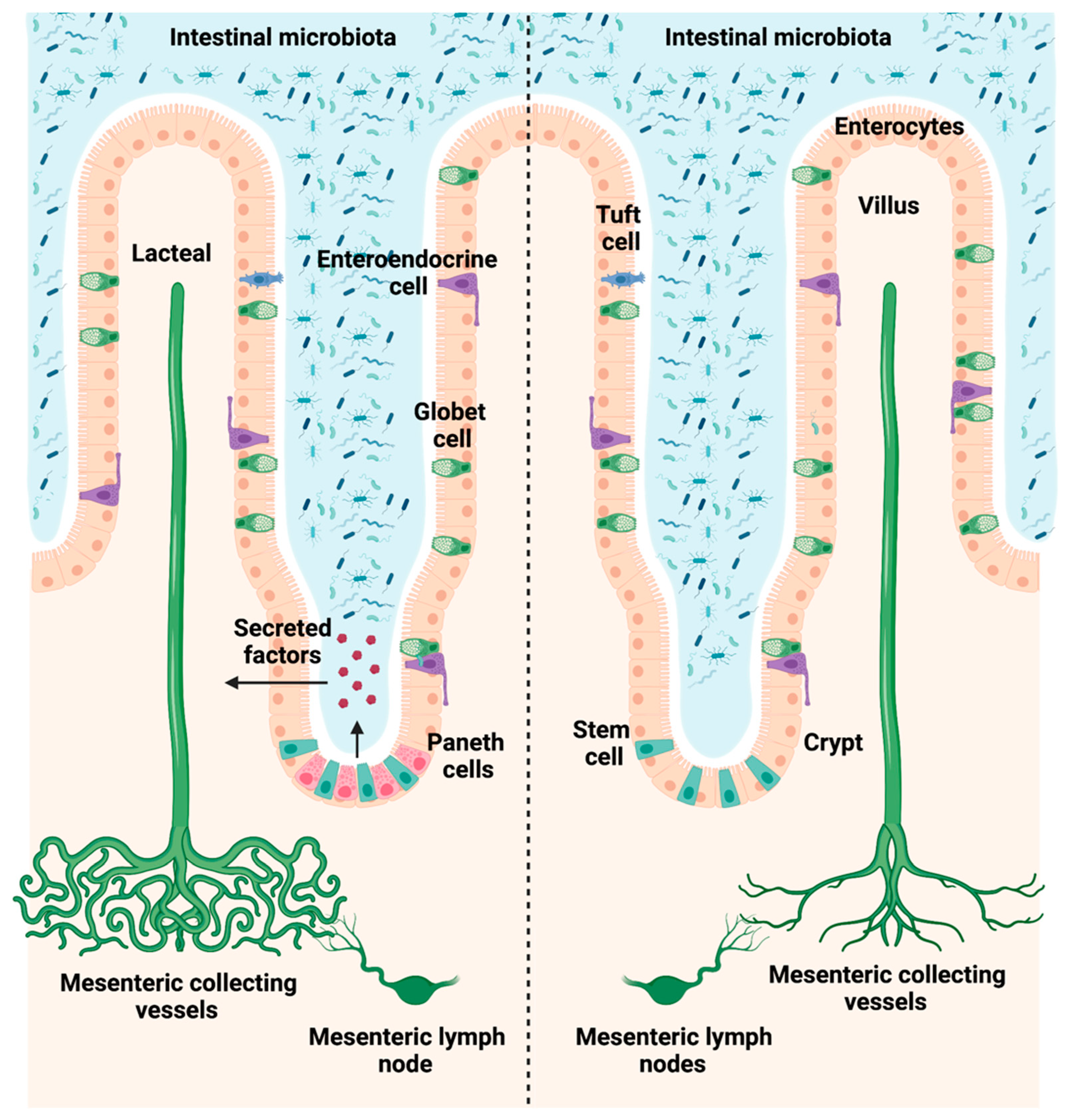
3.1. Paneth Cells Are Depleted in the Intestine of Math1Lox/LoxVilcreERT2 Mice after Tamoxifen Injection
3.2. Small Intestinal Gene Expression Is Altered in the Absence of Paneth Cells after Experimental Portal Hypertension
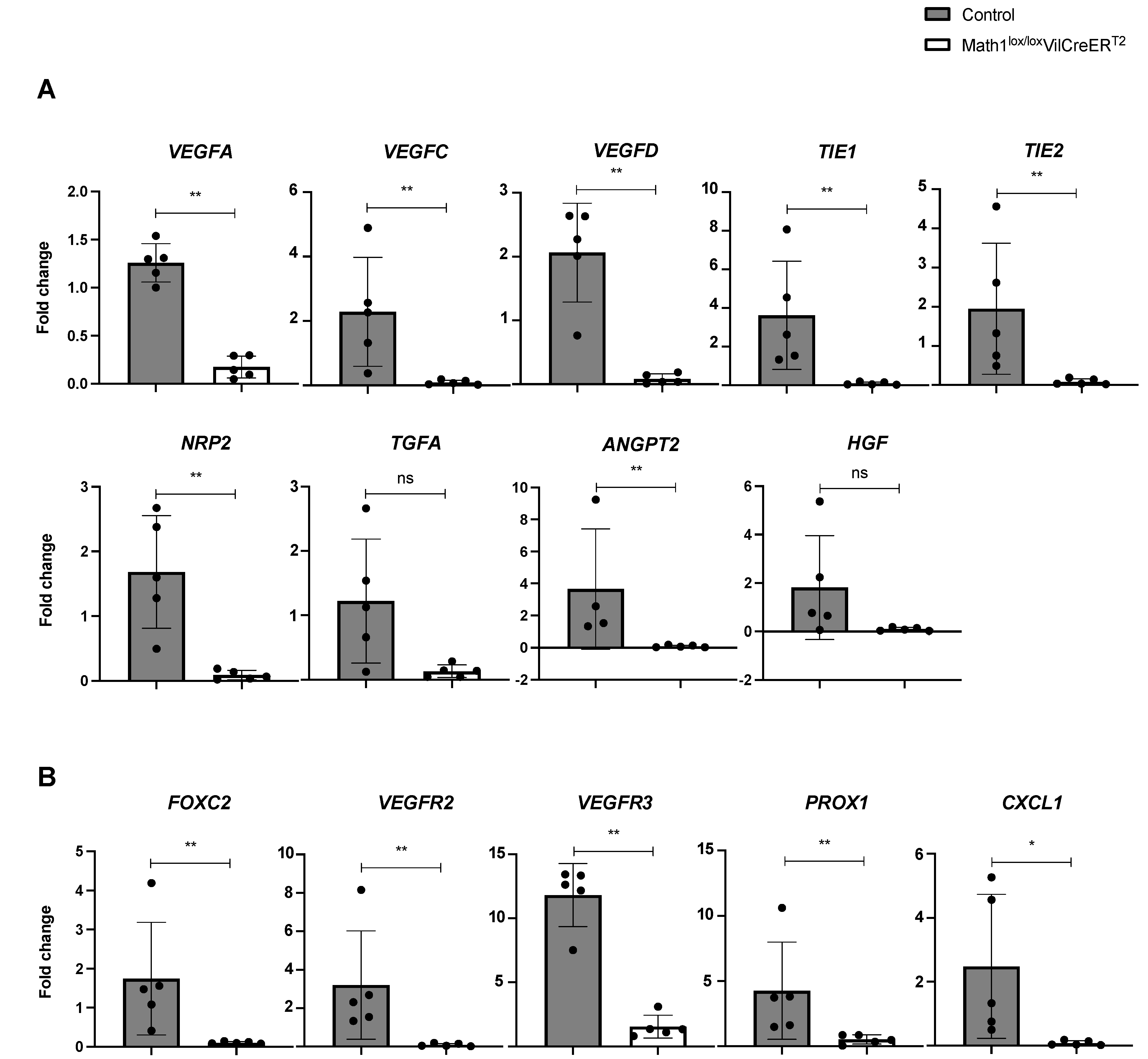
3.3. The Absence of Paneth Cells Is Associated with Reduced Expression of Lymphangiogenic Genes in Small Intestine of Portal Hypertensive Mice
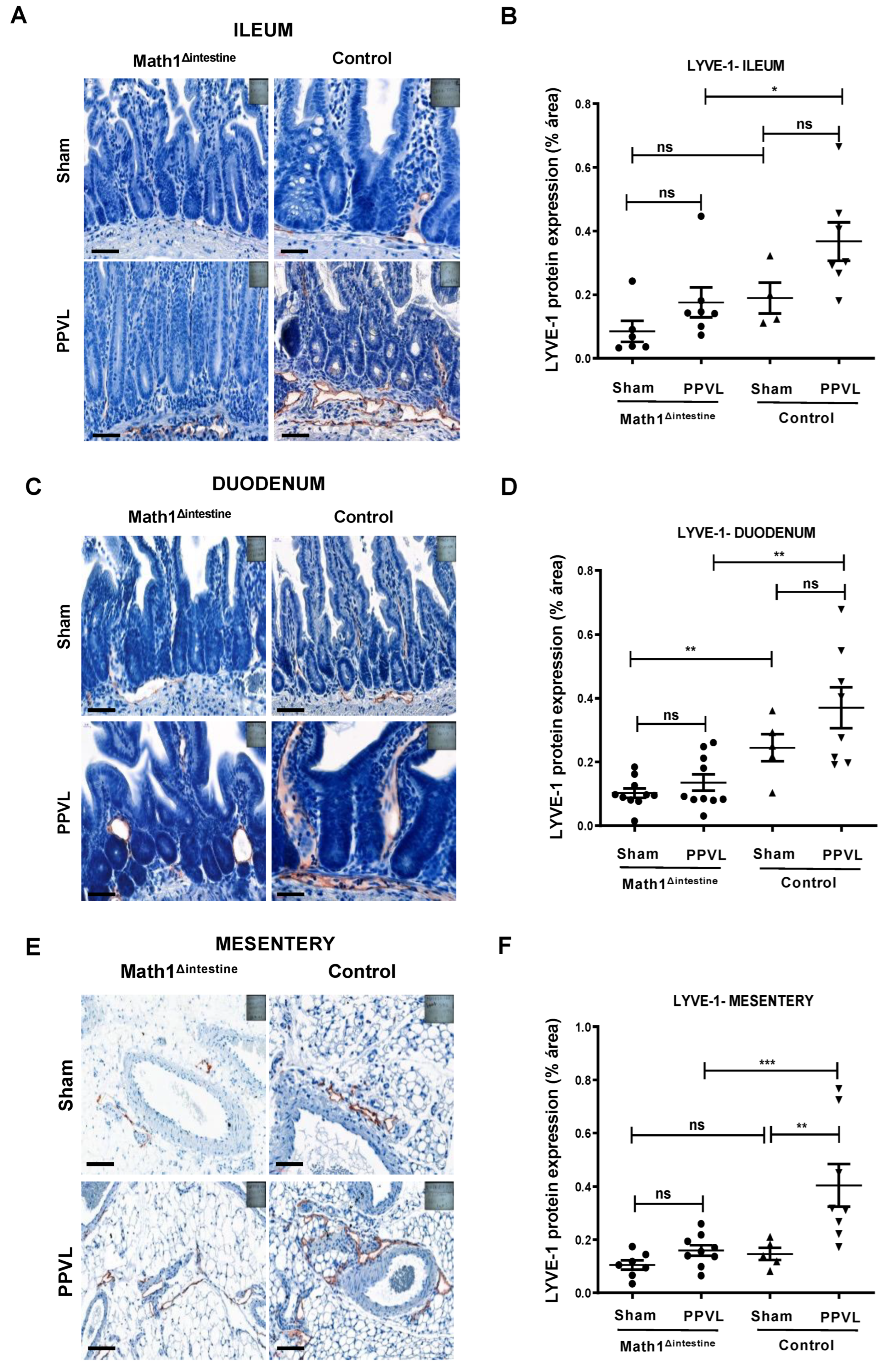
3.4. Loss of Paneth Cells Is Associated with Reduced Abundance of Intestinal and Mesenteric Lymphatic Vessels after PPVL
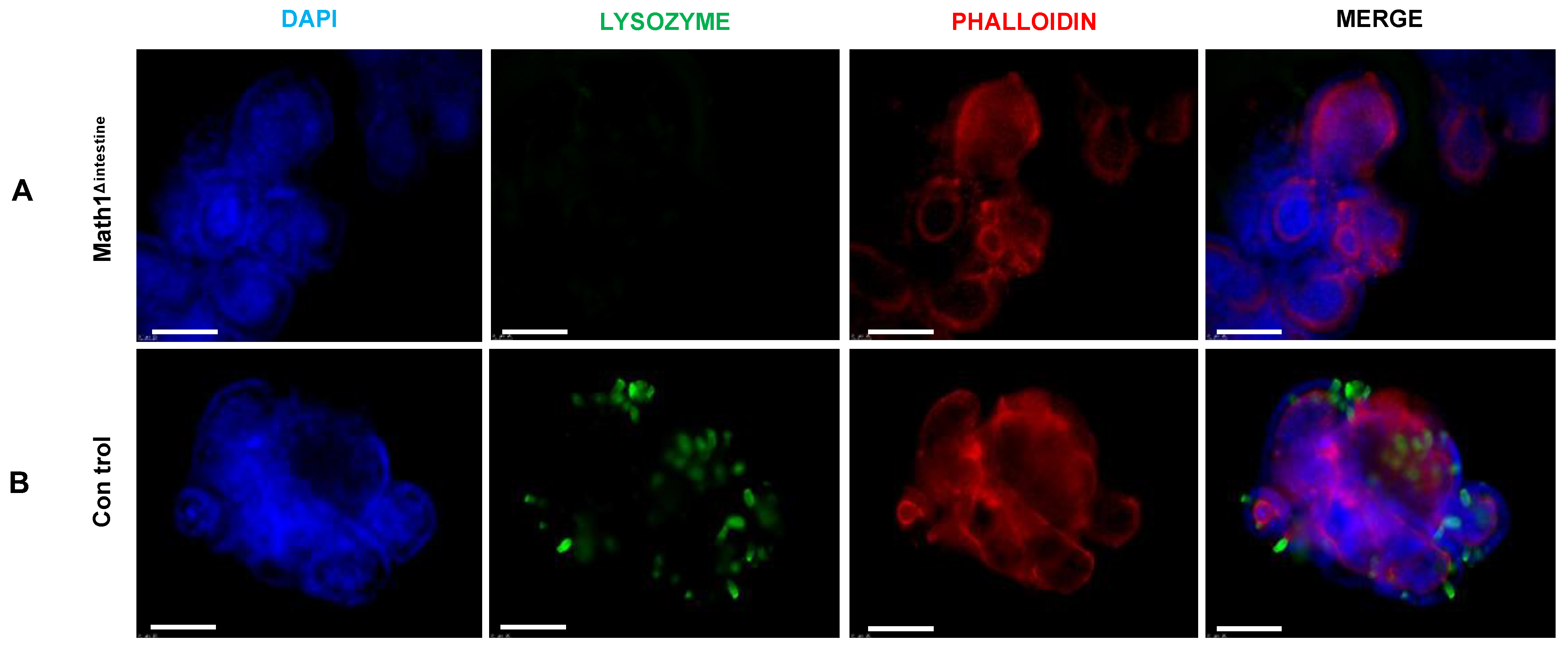
3.5. The Growth of Intestinal Organoids Is Not Affected by the Absence of Paneth Cells
3.6. PCs Secretory Products Stimulate Lymphangiogenic Responses In Vitro
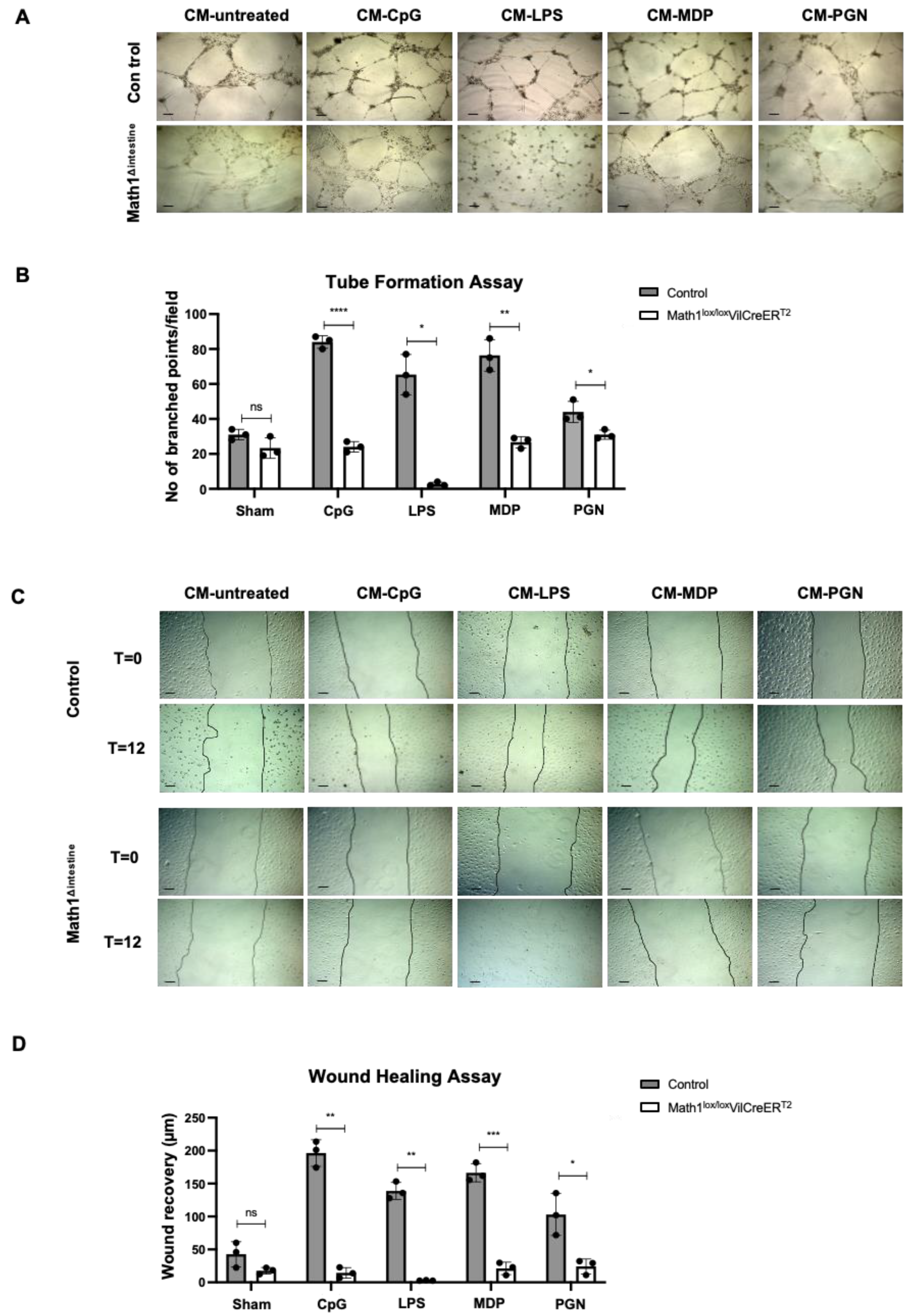
3.7. Proteins Secreted by Paneth Cells Are Associated with Metabolic Processes, Cell Proliferation and Cell Growth
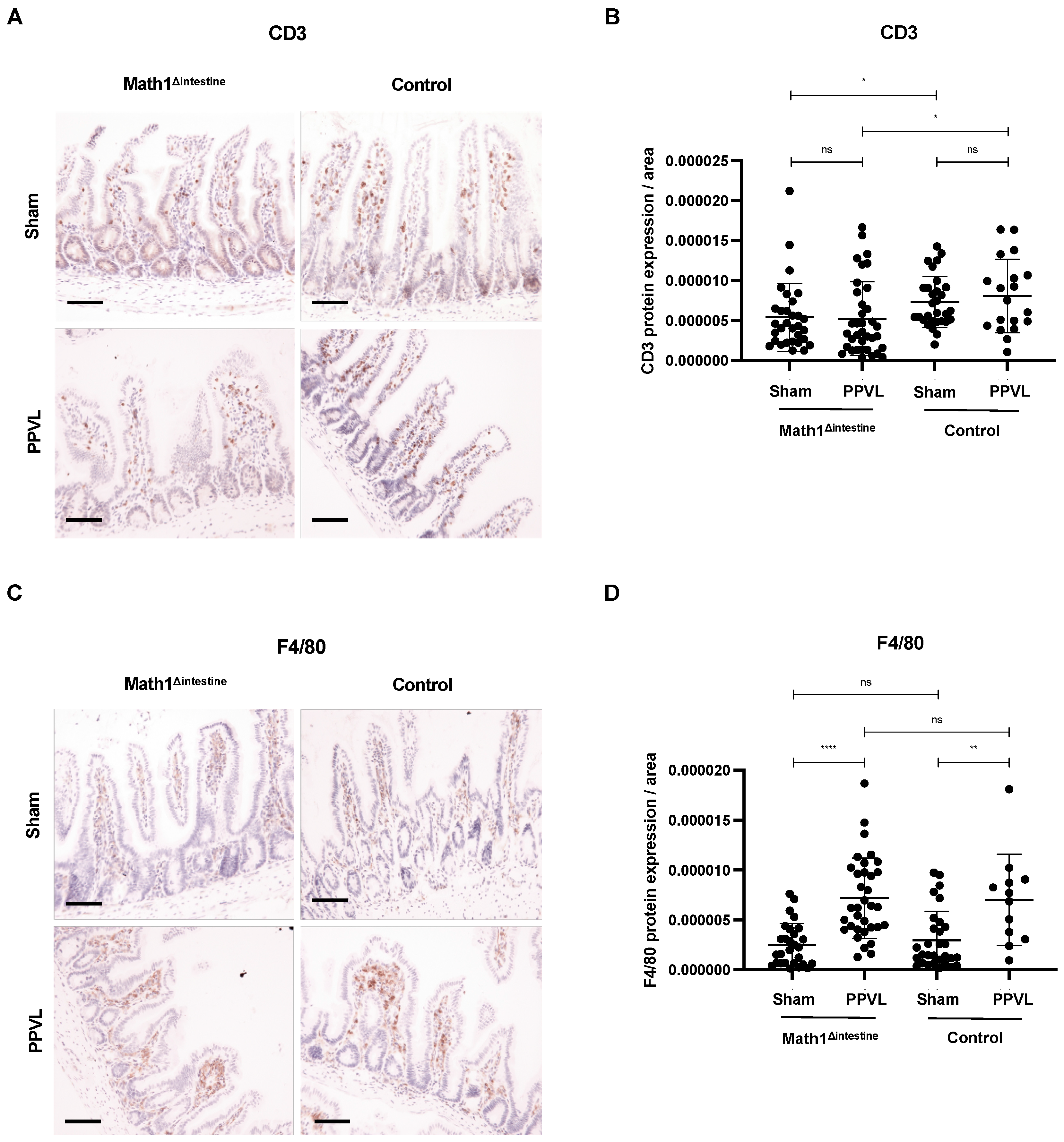
3.8. Intestinal Immune Cell Infiltration Is Slightly Affected by the Presence or Absence of Paneth Cells

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21st Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [Google Scholar] [CrossRef] [PubMed]
- Baluk, P.; Fuxe, J.; Hashizume, H.; Romano, T.; Lashnits, E.; Butz, S.; Vestweber, D.; Corada, M.; Molendini, C.; Dejana, E.; et al. Functionally specialized junctions between endothelial cells of lymphatic vessels. J. Exp. Med. 2007, 204, 2349–2362. [Google Scholar] [CrossRef] [PubMed]
- Petrova, T.V.; Koh, G.Y. Biological functions of lymphatic vessels. Science 2020, 369, eaax4063. [Google Scholar] [CrossRef] [PubMed]
- Bernier-Latmani, J.; Petrova, T.V. Intestinal lymphatic vasculature: Structure, mechanisms and functions. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 510–526. [Google Scholar] [CrossRef]
- Van Kruiningen, H.J.; Colombel, J.F. The forgotten role of lymphangitis in Crohn’s disease. Gut 2008, 57, 1–4. [Google Scholar] [CrossRef]
- Colombel, J.F.; Watson, A.J.; Neurath, M.F. The 10 remaining mysteries of inflammatory bowel disease. Gut 2008, 57, 429–433. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Iwakiri, Y. The Hepatic Lymphatic Vascular System: Structure, Function, Markers, and Lymphangiogenesis. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 733–749. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.; Iwakiri, Y. The lymphatic vascular system in liver diseases: Its role in ascites formation. Clin. Mol. Hepatol. 2013, 19, 99–104. [Google Scholar] [CrossRef]
- Witte, M.H.; Dumont, A.E.; Cole, W.R.; Witte, C.L.; Kintner, K. Lymph circulation in hepatic cirrhosis: Effect of portacaval shunt. Ann. Intern. Med. 1969, 70, 303–310. [Google Scholar] [CrossRef]
- Ribera, J.; Córdoba-Jover, B.; Portolés, I.; Morales-Ruiz, M. The Role of Hepatic and Splanchnic Lymphatic System in Portal Hypertension and Ascites. Curr. Hepatol. Rep. 2019, 18, 157–163. [Google Scholar] [CrossRef]
- Cardenas, A.; Bataller, R.; Arroyo, V. Mechanisms of ascites formation. Clin. Liver. Dis. 2000, 4, 447–465. [Google Scholar] [CrossRef]
- Henriksen, J.H. Estimation of lymphatic conductance: A model based on protein-kinetic studies and haemodynamic measurements in patients with cirrhosis of the liver and in pigs. Scand. J. Clin. Lab. Investig. 1985, 45, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Michitaka, K.; Onji, M. Morphometric analysis of lymphatic and blood vessels in human chronic viral liver diseases. Am. J. Pathol. 1998, 153, 1131–1137. [Google Scholar] [CrossRef] [Green Version]
- Yokomori, H.; Oda, M.; Kaneko, F.; Kawachi, S.; Tanabe, M.; Yoshimura, K.; Kitagawa, Y.; Hibi, T. Lymphatic marker podoplanin/D2-40 in human advanced cirrhotic liver-Re-evaluations of microlymphatic abnormalities. BMC Gastroenterol. 2010, 10, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.; Tanaka, M.; Iwakiri, Y. Hepatic lymphatic vascular system in health and disease. J. Hepatol. 2022, 77, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Moghadamrad, S.; McCoy, K.D.; Geuking, M.B.; Sägesser, H.; Kirundi, J.; Macpherson, A.J.; De Gottardi, A. Attenuated portal hypertension in germ-free mice: Function of bacterial flora on the development of mesenteric lymphatic and blood vessels. Hepatology 2015, 61, 1685–1695. [Google Scholar] [CrossRef]
- Salzman, N.H.; Hung, K.; Haribhai, D.; Chu, H.; Karlsson-Sjöberg, J.; Amir, E.; Teggatz, P.; Barman, M.; Hayward, M.; Eastwood, D.; et al. Enteric defensins are essential regulators of intestinal microbial ecology. Nat. Immunol. 2010, 11, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.I.; Regnault, B.; Mulet, C.; Tanguy, M.; Jay, P.; Sansonetti, P.J.; Pédron, T. Maturation of paneth cells induces the refractory state of newborn mice to Shigella infection. J. Immunol. 2008, 180, 4924–4930. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.; Moghadamrad, S.; Sorribas, M.; Muntet, S.G.; Kellmann, P.; Trentesaux, C.; Fraudeau, M.; Nanni, P.; Wolski, W.; Keller, I.; et al. Paneth cells promote angiogenesis and regulate portal hypertension in response to microbial signals. J. Hepatol. 2020, 73, 628–639. [Google Scholar] [CrossRef]
- Hooper, L.V.; Stappenbeck, T.S.; Hong, C.V.; Gordon, J.I. Angiogenins: A new class of microbicidal proteins involved in innate immunity. Nat. Immunol. 2003, 4, 269–273. [Google Scholar] [CrossRef]
- Stappenbeck, T.S.; Hooper, L.V.; Gordon, J.I. Developmental regulation of intestinal angiogenesis by indigenous microbes via Paneth cells. Proc. Natl. Acad. Sci. USA 2002, 99, 15451–15455. [Google Scholar] [CrossRef] [Green Version]
- Durand, A.; Donahue, B.; Peignon, G.; Letourneur, F.; Cagnard, N.; Slomianny, C.; Perret, C.; Shroyer, N.F.; Romagnolo, B. Functional intestinal stem cells after Paneth cell ablation induced by the loss of transcription factor Math1 (Atoh1). Proc. Natl. Acad. Sci. USA 2012, 109, 8965–8970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Moolenbeek, C.; Ruitenberg, E.J. The “Swiss roll”: A simple technique for histological studies of the rodent intestine. Lab. Anim. 1981, 15, 57–59. [Google Scholar] [CrossRef]
- Mahe, M.M.; Aihara, E.; Schumacher, M.A.; Zavros, Y.; Montrose, M.H.; Helmrath, M.A.; Sato, T.; Shroyer, N.F. Establishment of Gastrointestinal Epithelial Organoids. Curr. Protoc. Mouse Biol. 2013, 3, 217–240. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Trachsel, C.; Panse, C.; Kockmann, T.; Wolski, W.E.; Grossmann, J.; Schlapbach, R. rawDiag: An R Package Supporting Rational LC-MS Method Optimization for Bottom-up Proteomics. J. Proteome Res. 2018, 17, 2908–2914. [Google Scholar] [CrossRef] [Green Version]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Vasaikar, S.; Shi, Z.; Greer, M.; Zhang, B. WebGestalt 2017: A more comprehensive, powerful, flexible and interactive gene set enrichment analysis toolkit. Nucleic Acids Res. 2017, 45, W130–W137. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Aspelund, A.; Alitalo, K. Lymphangiogenic factors, mechanisms, and applications. J. Clin. Investig. 2014, 124, 878–887. [Google Scholar] [CrossRef]
- Barrowman, J.A.; Granger, D.N. Effects of experimental cirrhosis on splanchnic microvascular fluid and solute exchange in the rat. Gastroenterology 1984, 87, 165–172. [Google Scholar] [CrossRef]
- Oikawa, H.; Masuda, T.; Sato, S.; Yashima, A.; Suzuki, K.; Satodate, R. Changes in lymph vessels and portal veins in the portal tract of patients with idiopathic portal hypertension: A morphometric study. Hepatology 1998, 27, 1607–1610. [Google Scholar] [CrossRef]
- Ayabe, T.; Satchell, D.P.; Wilson, C.L.; Parks, W.C.; Selsted, M.E.; Ouellette, A.J. Secretion of microbicidal alpha-defensins by intestinal Paneth cells in response to bacteria. Nat. Immunol. 2000, 1, 113–118. [Google Scholar] [CrossRef]
- Wehkamp, J.; Salzman, N.H.; Porter, E.; Nuding, S.; Weichenthal, M.; Petras, R.E.; Shen, B.; Schaeffeler, E.; Schwab, M.; Linzmeier, R.; et al. Reduced Paneth cell alpha-defensins in ileal Crohn’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 18129–18134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teltschik, Z.; Wiest, R.; Beisner, J.; Nuding, S.; Hofmann, C.; Schoelmerich, J.; Bevins, C.L.; Stange, E.F.; Wehkamp, J. Intestinal bacterial translocation in rats with cirrhosis is related to compromised Paneth cell antimicrobial host defense. Hepatology 2012, 55, 1154–1163. [Google Scholar] [CrossRef]
- Adams, R.H.; Alitalo, K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 2007, 8, 464–478. [Google Scholar] [CrossRef]
- Partanen, T.A.; Arola, J.; Saaristo, A.; Jussila, L.; Ora, A.; Miettinen, M.; Stacker, S.A.; Achen, M.G.; Alitalo, K. VEGF-C and VEGF-D expression in neuroendocrine cells and their receptor, VEGFR-3, in fenestrated blood vessels in human tissues. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2000, 14, 2087–2096. [Google Scholar] [CrossRef]
- Ohtani, O.; Ohtani, Y. Lymph circulation in the liver. Anat. Rec. (Hoboken) 2008, 291, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Petrova, T.V.; Mäkinen, T.; Mäkelä, T.P.; Saarela, J.; Virtanen, I.; Ferrell, R.E.; Finegold, D.N.; Kerjaschki, D.; Ylä-Herttuala, S.; Alitalo, K. Lymphatic endothelial reprogramming of vascular endothelial cells by the Prox-1 homeobox transcription factor. EMBO J. 2002, 21, 4593–4599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäkinen, T.; Veikkola, T.; Mustjoki, S.; Karpanen, T.; Catimel, B.; Nice, E.C.; Wise, L.; Mercer, A.; Kowalski, H.; Kerjaschki, D.; et al. Isolated lymphatic endothelial cells transduce growth, survival and migratory signals via the VEGF-C/D receptor VEGFR-3. EMBO J. 2001, 20, 4762–4773. [Google Scholar] [CrossRef] [Green Version]
- Ribera, J.; Pauta, M.; Melgar-Lesmes, P.; Tugues, S.; Fernández-Varo, G.; Held, K.F.; Soria, G.; Tudela, R.; Planas, A.M.; Fernández-Hernando, C.; et al. Increased nitric oxide production in lymphatic endothelial cells causes impairment of lymphatic drainage in cirrhotic rats. Gut 2013, 62, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.D.; Goldblum, S.E. Mechanisms of bacterial lipopolysaccharide-induced endothelial apoptosis. Am. J. Physiol. Lung. Cell Mol. Physiol. 2003, 284, L899–L914. [Google Scholar] [CrossRef] [Green Version]
- Pulido, D.; Nogués, M.V.; Boix, E.; Torrent, M. Lipopolysaccharide neutralization by antimicrobial peptides: A gambit in the innate host defense strategy. J. Innate Immun. 2012, 4, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Kerjaschki, D. The crucial role of macrophages in lymphangiogenesis. J. Clin. Investig. 2005, 115, 2316–2319. [Google Scholar] [CrossRef] [Green Version]
- Watari, K.; Nakao, S.; Fotovati, A.; Basaki, Y.; Hosoi, F.; Bereczky, B.; Higuchi, R.; Miyamoto, T.; Kuwano, M.; Ono, M. Role of macrophages in inflammatory lymphangiogenesis: Enhanced production of vascular endothelial growth factor C and D through NF-kappaB activation. Biochem. Biophys. Res. Commun. 2008, 377, 826–831. [Google Scholar] [CrossRef]
- Ogata, F.; Fujiu, K.; Matsumoto, S.; Nakayama, Y.; Shibata, M.; Oike, Y.; Koshima, I.; Watabe, T.; Nagai, R.; Manabe, I. Excess Lymphangiogenesis Cooperatively Induced by Macrophages and CD4(+) T Cells Drives the Pathogenesis of Lymphedema. J. Investig. Dermatol. 2016, 136, 706–714. [Google Scholar] [CrossRef] [Green Version]
- Tsuru, S.; Ito, Y.; Matsuda, H.; Hosono, K.; Inoue, T.; Nakamoto, S.; Kurashige, C.; Mishima, T.; Tsujikawa, K.; Okamoto, H.; et al. RAMP1 signaling in immune cells regulates inflammation-associated lymphangiogenesis. Lab. Investig. 2020, 100, 738–750. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham Control vs. Sham Math1Δint | ||||
|---|---|---|---|---|
| S.No. | Gene_ID | Mgi_Symbol | log2 Fold Change | p-adj |
| 1 | ENSMUSG00000096499 | Ighv1-5 | 1.813 | 0.002 |
| PPVL Control vs. PPVL Math1Δint | ||||
|---|---|---|---|---|
| S.No. | Gene_ID | Mgi_Symbol | log2 Fold Change | p-adj |
| 1 | ENSMUSG00000020407 | Upp1 | 1.356 | 0.007 |
| 2 | ENSMUSG00000003355 | Fkbp11 | 1.241 | 0.007 |
| 3 | ENSMUSG00000030659 | Nucb2 | 1.222 | 0.007 |
| 4 | ENSMUSG00000053695 | Defb37 | −1.598 | 0.007 |
| 5 | ENSMUSG00000025196 | Cpn1 | −1.669 | 0.009 |
| 6 | ENSMUSG00000070427 | Il18bp | 1.480 | 0.010 |
| 7 | ENSMUSG00000093394 | Gm20621 | −1.598 | 0.011 |
| 8 | ENSMUSG00000000440 | Pparg | −1.242 | 0.013 |
| 9 | ENSMUSG00000040543 | Pitpnm3 | −1.317 | 0.015 |
| 10 | ENSMUSG00000029417 | Cxcl9 | 1.482 | 0.016 |
| 11 | ENSMUSG00000061825 | Ces2c | −1.147 | 0.018 |
| 12 | ENSMUSG00000095981 | Ighv10-1 | 1.518 | 0.020 |
| 13 | ENSMUSG00000096727 | Psmb9 | 1.195 | 0.023 |
| 14 | ENSMUSG00000074934 | Grem1 | −0.802 | 0.026 |
| 15 | ENSMUSG00000079116 | Gm15293 | 1.321 | 0.026 |
| 16 | ENSMUSG00000081769 | Gm12216 | 1.080 | 0.026 |
| 17 | ENSMUSG00000004933 | Matk | −1.476 | 0.026 |
| 18 | ENSMUSG00000027514 | Zbp1 | 1.467 | 0.026 |
| 19 | ENSMUSG00000079339 | Ifit1bl1 | 1.476 | 0.026 |
| 20 | ENSMUSG00000055748 | Gsdmc4 | −1.433 | 0.026 |
| 21 | ENSMUSG00000047844 | Bex4 | −1.457 | 0.026 |
| 22 | ENSMUSG00000110639 | Gm6831 | −1.460 | 0.026 |
| 23 | ENSMUSG00000067599 | Klra7 | 1.428 | 0.026 |
| 24 | ENSMUSG00000032418 | Me1 | −1.458 | 0.026 |
| 25 | ENSMUSG00000094662 | Defa36 | 1.334 | 0.026 |
| 26 | ENSMUSG00000022216 | Psme1 | 0.700 | 0.026 |
| 27 | ENSMUSG00000076633 | Ighv5-2 | −1.433 | 0.027 |
| 28 | ENSMUSG00000035692 | Isg15 | 1.421 | 0.027 |
| 29 | ENSMUSG00000056293 | Gsdmc2 | −1.374 | 0.027 |
| 30 | ENSMUSG00000040808 | S100g | −1.354 | 0.027 |
| 31 | ENSMUSG00000046718 | Bst2 | 0.892 | 0.027 |
| 32 | ENSMUSG00000068614 | Actc1 | −1.414 | 0.030 |
| 33 | ENSMUSG00000022964 | Tmem50b | 0.447 | 0.030 |
| 34 | ENSMUSG00000095328 | Defa-ps6 | 1.422 | 0.031 |
| 35 | ENSMUSG00000035105 | Egln3 | −1.316 | 0.031 |
| 36 | ENSMUSG00000094797 | Igkv6-15 | 1.381 | 0.031 |
| 37 | ENSMUSG00000021730 | Hcn1 | −1.415 | 0.031 |
| 38 | ENSMUSG00000078612 | Fyb2 | −1.370 | 0.034 |
| 39 | ENSMUSG00000055827 | Gsdmc3 | −1.336 | 0.034 |
| 40 | ENSMUSG00000076512 | Igkv9-123 | 1.225 | 0.034 |
| 41 | ENSMUSG00000039323 | Igfbp2 | −1.388 | 0.036 |
| 42 | ENSMUSG00000022587 | Ly6e | 0.953 | 0.036 |
| 43 | ENSMUSG00000020469 | Myl7 | 0.872 | 0.036 |
| 44 | ENSMUSG00000025041 | Nt5c2 | −0.953 | 0.036 |
| 45 | ENSMUSG00000026104 | Stat1 | 1.133 | 0.036 |
| 46 | ENSMUSG00000030107 | Usp18 | 1.304 | 0.036 |
| 47 | ENSMUSG00000098557 | Kctd12 | −1.194 | 0.037 |
| 48 | ENSMUSG00000094687 | Defa25 | 1.231 | 0.037 |
| 49 | ENSMUSG00000078853 | Igtp | 1.372 | 0.037 |
| 50 | ENSMUSG00000028965 | Tnfrsf9 | 1.239 | 0.037 |
| 51 | ENSMUSG00000052373 | Mpp3 | −1.332 | 0.038 |
| 52 | ENSMUSG00000095565 | Ighv2-9-1 | 1.294 | 0.041 |
| 53 | ENSMUSG00000027397 | Slc20a1 | −1.194 | 0.043 |
| 54 | ENSMUSG00000030173 | Klra5 | 1.349 | 0.043 |
| 55 | ENSMUSG00000018211 | Wfdc15b | −1.296 | 0.043 |
| 56 | ENSMUSG00000026981 | Il1rn | −1.278 | 0.046 |
| 57 | ENSMUSG00000050097 | Ces2b | −1.344 | 0.046 |
| 58 | ENSMUSG00000020319 | Wdpcp | 1.168 | 0.046 |
| 59 | ENSMUSG00000024600 | Slc27a6 | −1.306 | 0.048 |
| 60 | ENSMUSG00000054169 | Ceacam10 | 1.288 | 0.048 |
| 61 | ENSMUSG00000046460 | Sh2d7 | −1.333 | 0.049 |
| 62 | ENSMUSG00000029307 | Dmp1 | −1.330 | 0.049 |
| 63 | ENSMUSG00000020571 | Pdia6 | 0.609 | 0.049 |
| GO.ID | Term | Annotated | Significant | Weight01.Fisher p-Value | Genes |
|---|---|---|---|---|---|
| GO:0071222 | Cellular response to lipopolysaccharide | 177 | 5 | 0.00 | Cxcl9, Defa25, Gm15293, Il1rn, Stat1 |
| GO:0061844 | Antimicrobial humoral immune response mediated by antimicrobial peptide | 56 | 3 | 0.00 | Cxcl9, Defa25, Gm15293 |
| GO:0042742 | Defense response to bacterium | 185 | 5 | 0.00 | Defa25, Defb37, Gm15293, Isg15, Wfdc15b |
| GO:0046321 | Positive regulation of fatty acid oxidation | 17 | 2 | 0.00 | Nucb2, Pparg |
| GO:0045648 | Positive regulation of erythrocyte differentiation | 23 | 2 | 0.00 | Isg15, Stat1 |
| GO:0051673 | Membrane disruption in other organism | 25 | 2 | 0.00 | Defa25, Gm15293 |
| GO:0002227 | Innate immune response in mucosa | 31 | 2 | 0.00 | Defa25, Gm15293 |
| GO:0072608 | Interleukin-10 secretion | 12 | 2 | 0.01 | Isg15, Tnfrsf9 |
| GO:0061326 | Renal tubule development | 80 | 2 | 0.01 | Grem1, Stat1 |
| GO:0042127 | Regulation of cell proliferation | 1393 | 8 | 0.01 | Bex4, Egln3, Grem1, Igfbp2, Matk, Pparg, Stat1, Tnfrsf9 |
| GO:0019731 | Antibacterial humoral response | 38 | 2 | 0.01 | Defa25, Gm15293 |
| GO:0035458 | Cellular response to interferon-beta | 45 | 2 | 0.01 | Igtp, Stat1 |
| GO:0010742 | Macrophage derived foam cell differentiation | 23 | 2 | 0.01 | Pparg, Stat1 |
| GO:1900077 | Negative regulation of cellular response to insulin stimulus | 40 | 2 | 0.01 | Nucb2, Pparg |
| GO:0090278 | Negative regulation of peptide hormone secretion | 56 | 2 | 0.02 | Bst2, Isg15 |
| GO:0051893 | Regulation of focal adhesion assembly | 56 | 2 | 0.02 | Grem1, Wdpcp |
| GO:0034340 | Response to type I interferon | 37 | 3 | 0.02 | Isg15, Stat1, Zbp1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.; Juanola, O.; Keller, I.; Nanni, P.; Wolski, W.; Martínez-López, S.; Caparrós, E.; Francés, R.; Moghadamrad, S. Paneth Cells Regulate Lymphangiogenesis under Control of Microbial Signals during Experimental Portal Hypertension. Biomedicines 2022, 10, 1503. https://doi.org/10.3390/biomedicines10071503
Hassan M, Juanola O, Keller I, Nanni P, Wolski W, Martínez-López S, Caparrós E, Francés R, Moghadamrad S. Paneth Cells Regulate Lymphangiogenesis under Control of Microbial Signals during Experimental Portal Hypertension. Biomedicines. 2022; 10(7):1503. https://doi.org/10.3390/biomedicines10071503
Chicago/Turabian StyleHassan, Mohsin, Oriol Juanola, Irene Keller, Paolo Nanni, Witold Wolski, Sebastián Martínez-López, Esther Caparrós, Rubén Francés, and Sheida Moghadamrad. 2022. "Paneth Cells Regulate Lymphangiogenesis under Control of Microbial Signals during Experimental Portal Hypertension" Biomedicines 10, no. 7: 1503. https://doi.org/10.3390/biomedicines10071503
APA StyleHassan, M., Juanola, O., Keller, I., Nanni, P., Wolski, W., Martínez-López, S., Caparrós, E., Francés, R., & Moghadamrad, S. (2022). Paneth Cells Regulate Lymphangiogenesis under Control of Microbial Signals during Experimental Portal Hypertension. Biomedicines, 10(7), 1503. https://doi.org/10.3390/biomedicines10071503