
Far-Infrared Therapy Decreases Orthotopic Allograft Transplantation Vasculopathy

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Equipment
2.2. Animal Study
2.2.1. Authorization of Animal Study
2.2.2. Orthotopic Aortic Transplantation
2.2.3. Animal Grouping
2.2.4. Biochemical Measurements and Enzyme-Linked Immunosorbent Assays
2.2.5. Morphological Analysis
2.2.6. Flow Cytometry
2.3. In Vitro Study
2.3.1. Cultivation of Human EPCs and FIR Therapy
2.3.2. Tubing Formation Assay
2.3.3. Cellular Senescence Assay
2.3.4. Migration Assay (Wound-Healing Assay)
2.3.5. Real-Time Quantitative Polymerase Chain Reactions
2.3.6. Western Blotting Analysis
2.4. Statistical Analyses
3. Results
3.1. FIR Therapy Affected the Inflammation-Related Proteins Expression, Not the Biochemical Characteristics, in OAT ACI/NKyo Rats
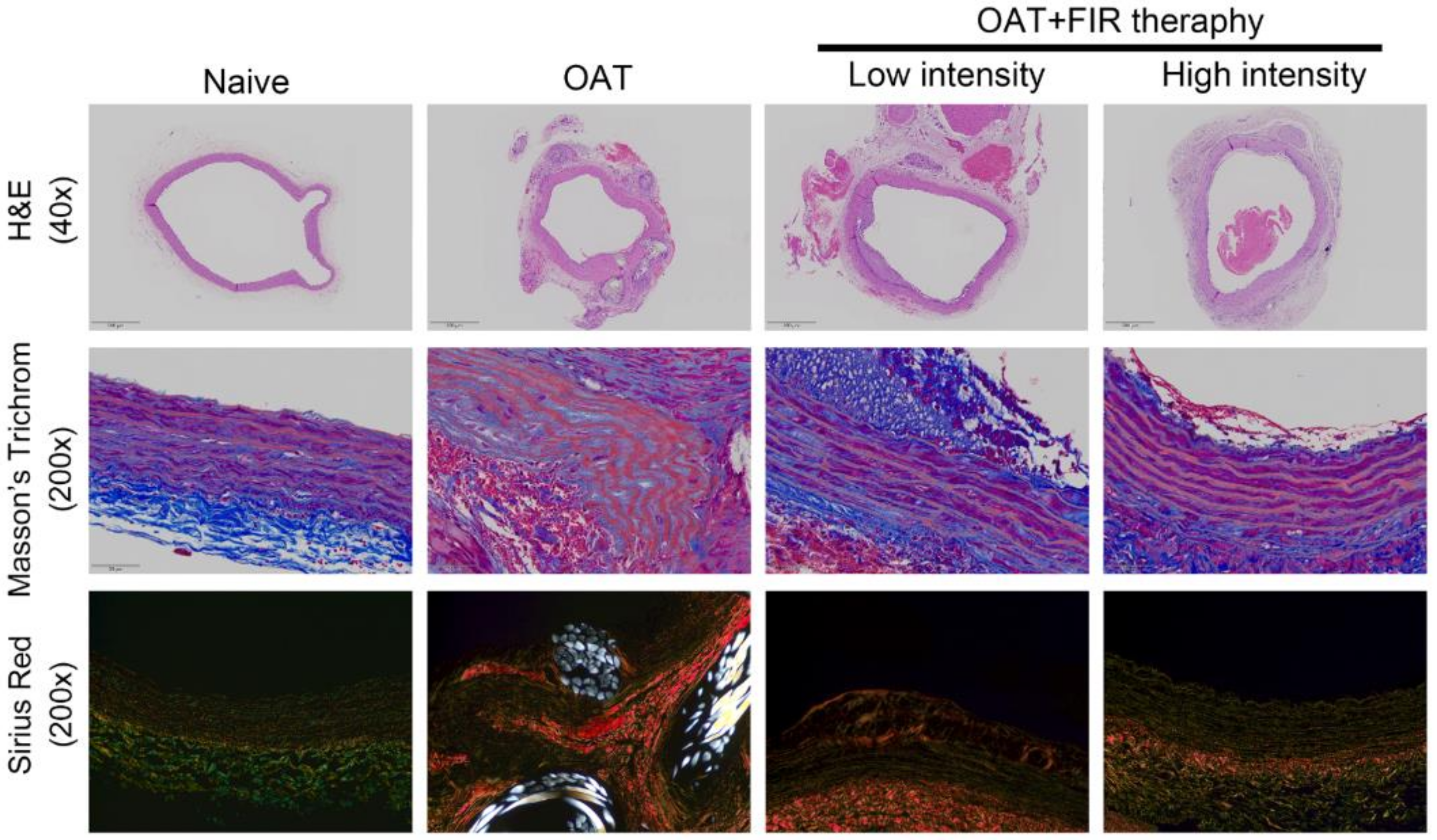
3.2. FIR Therapy Decreases Vascular Damage and Accumulation of Collagen in OAT ACI/NKyo Rats
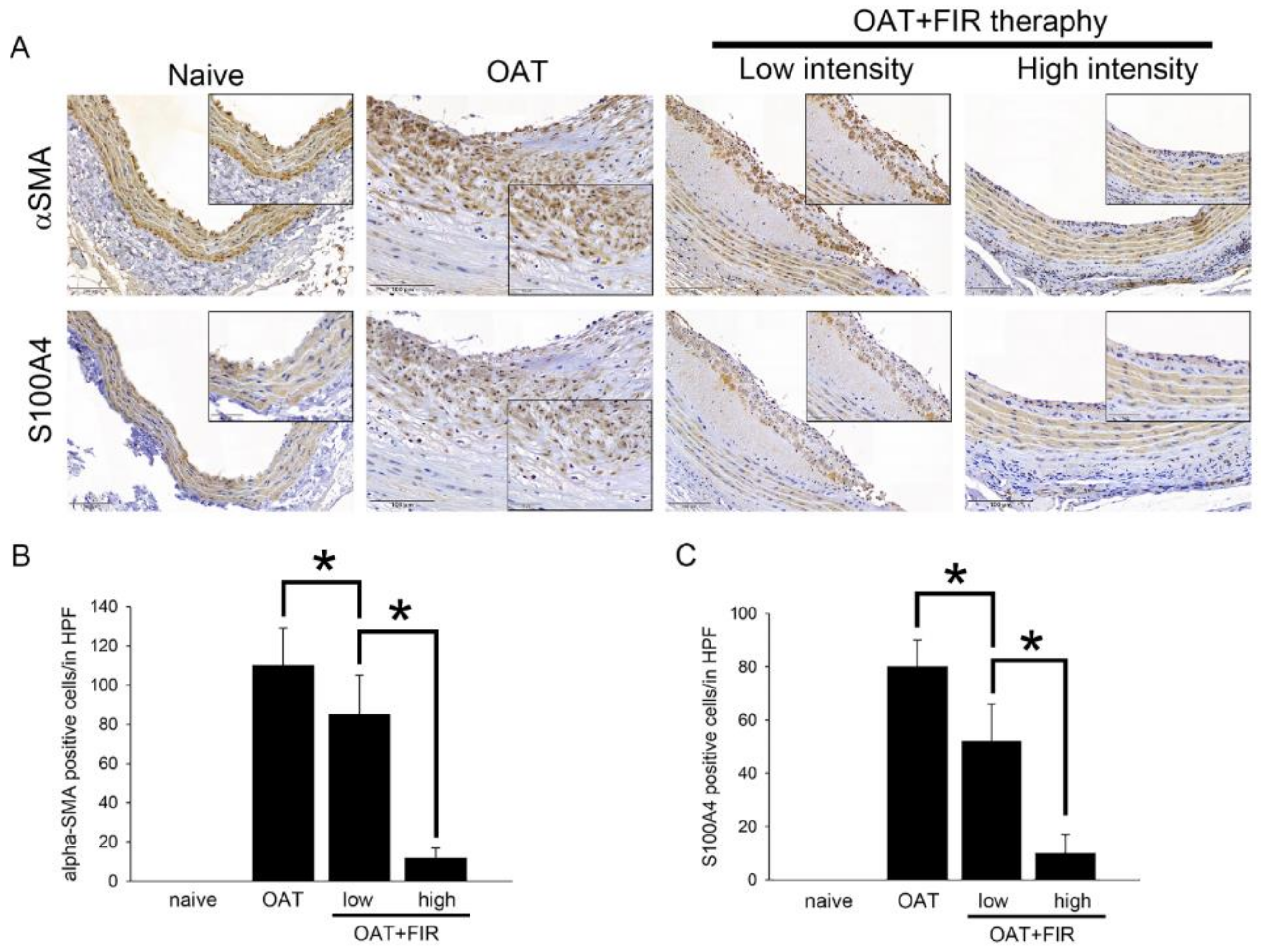
3.3. Reduced Proliferation of SMCs and Fibroblasts in the Aortic Wall of FIR Therapy-Administered OAT ACI/Nkyo Rats
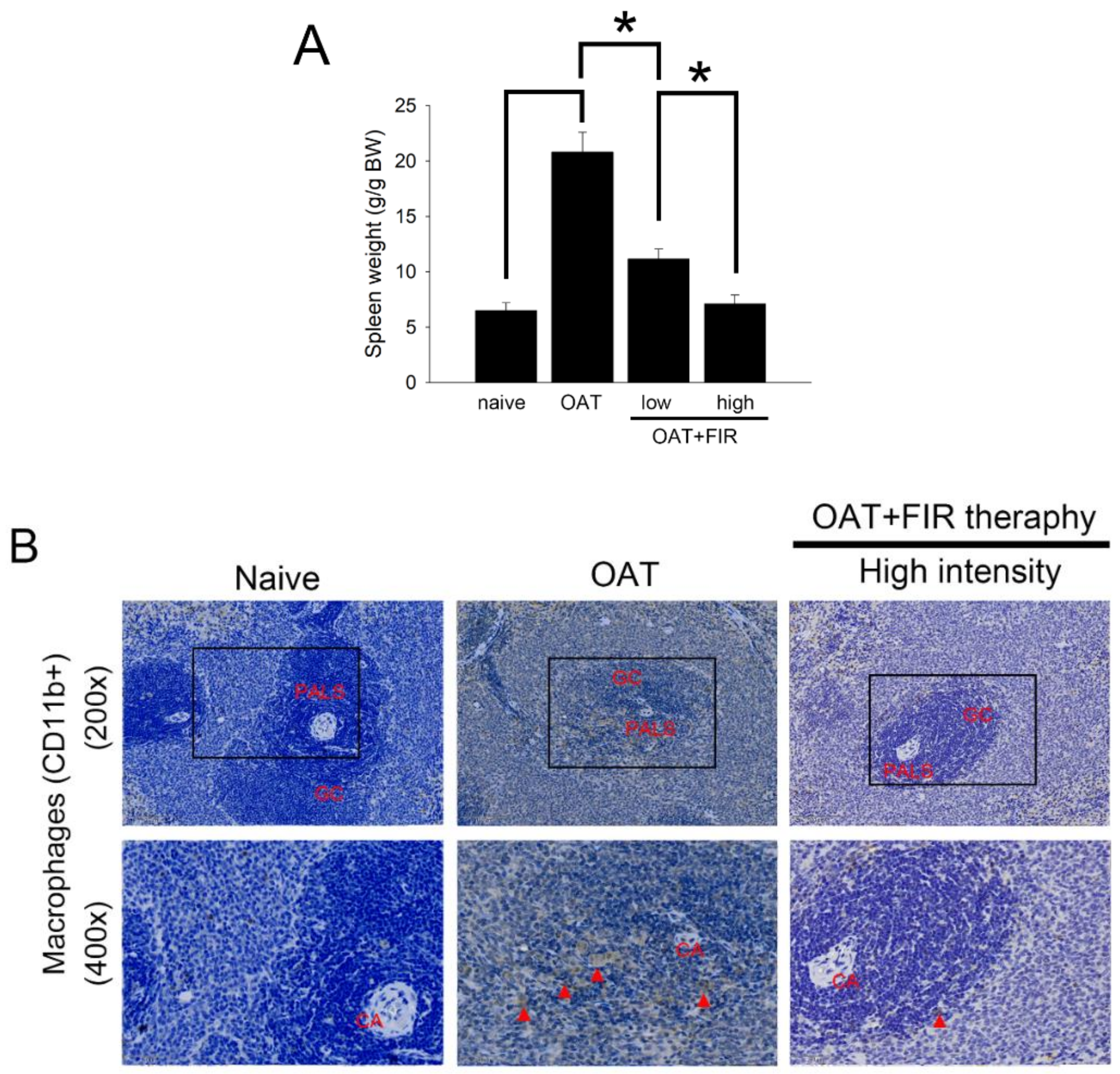
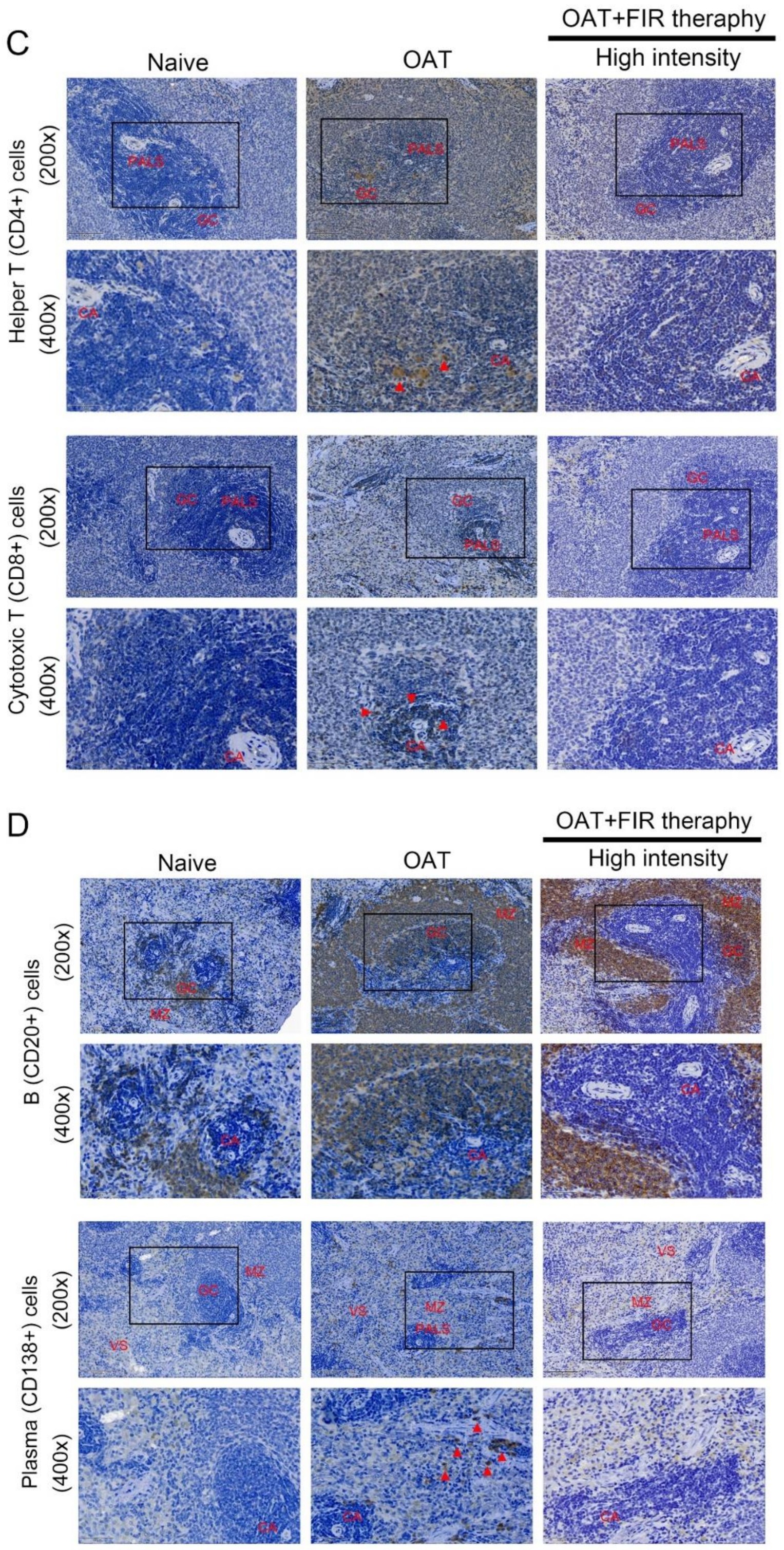
3.4. FIR Therapy Reduced Immune Responses in OAT ACI/NKyo Rats
3.5. FIR Therapy Lower Cytokines and Inflammation-Related Proteins Production in OAT ACI/NKyo Rats
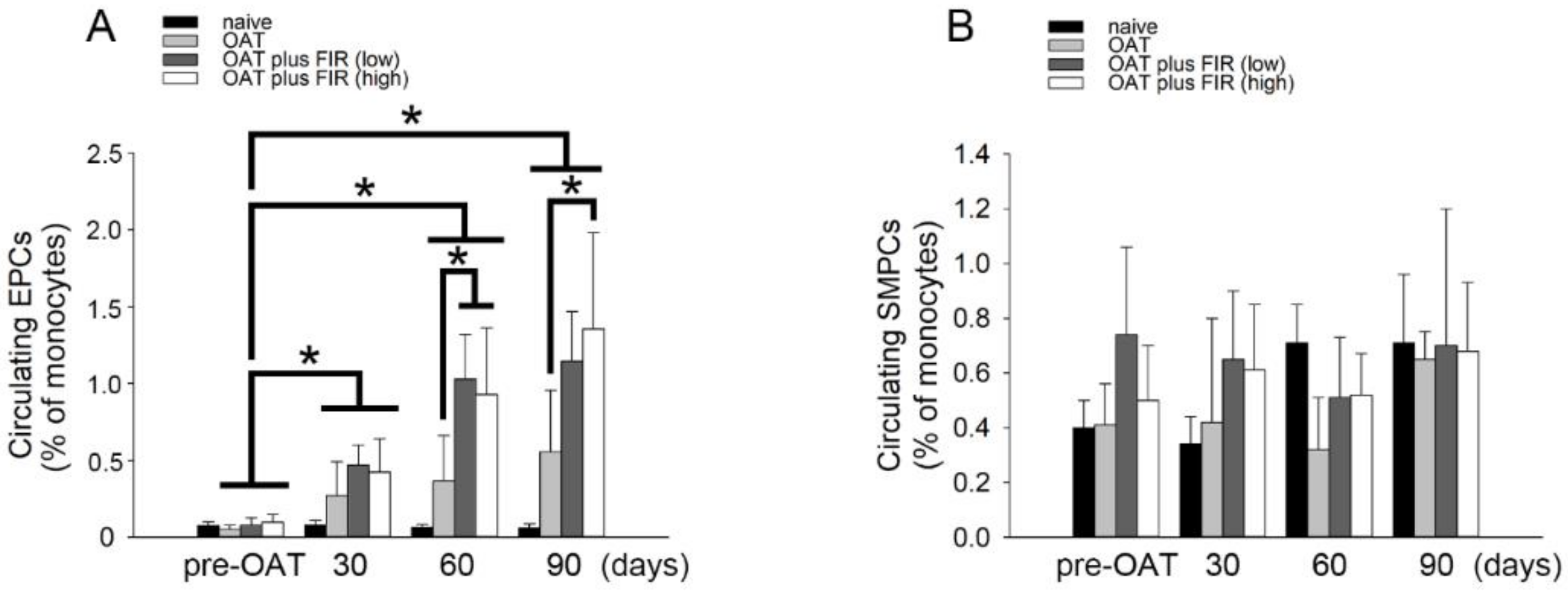
3.6. FIR Therapy Mobilized Circulating EPCs, Not SMPCs, in OAT ACI/NKyo Rats
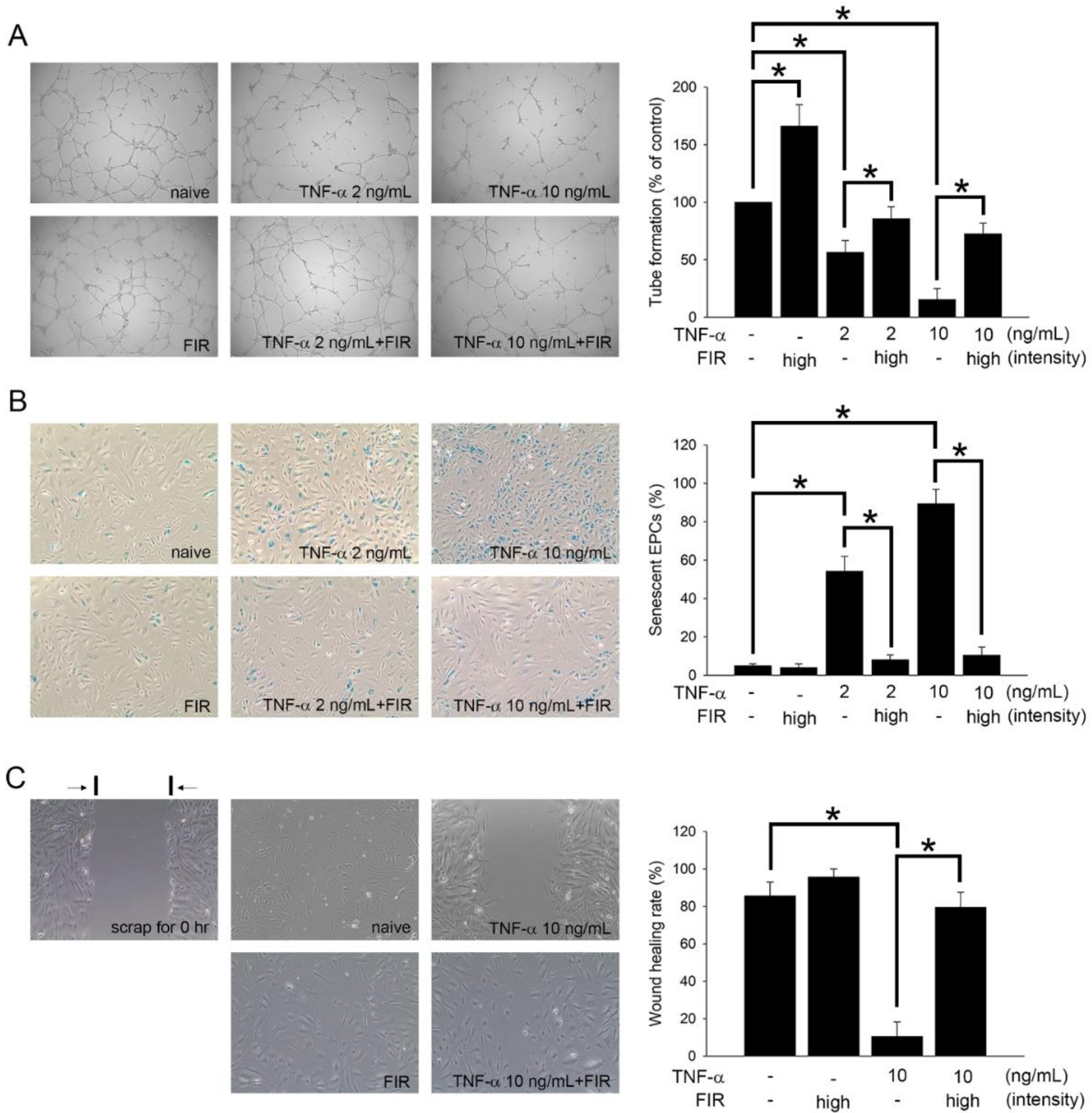
3.7. FIR Treatment That Regulates the Functions of EPCs May Mediate OAT Vasculopathy
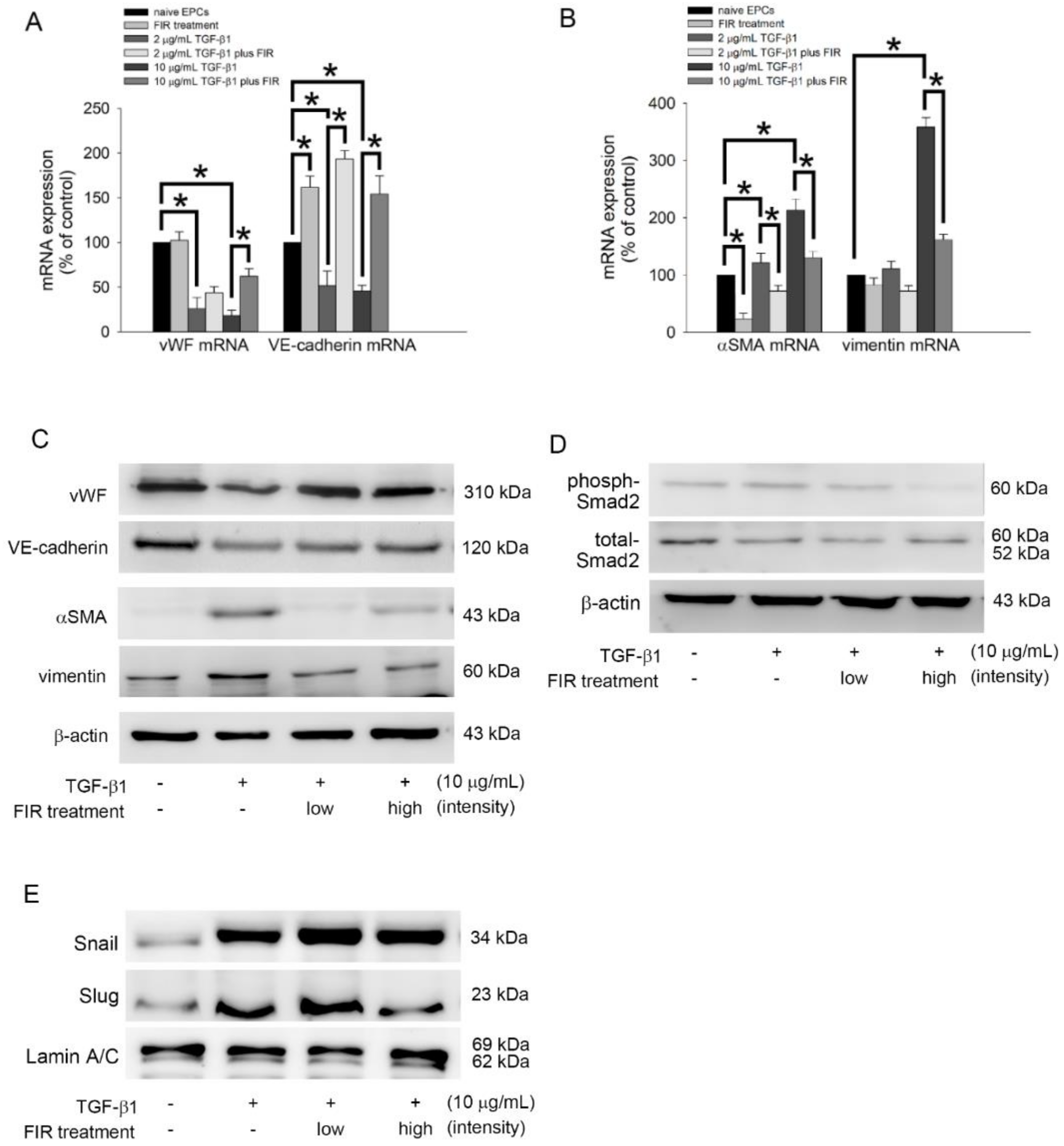
3.8. FIR Treatment Regulates the EndoMT of EPCs and May Mediate OAT Vasculopathy
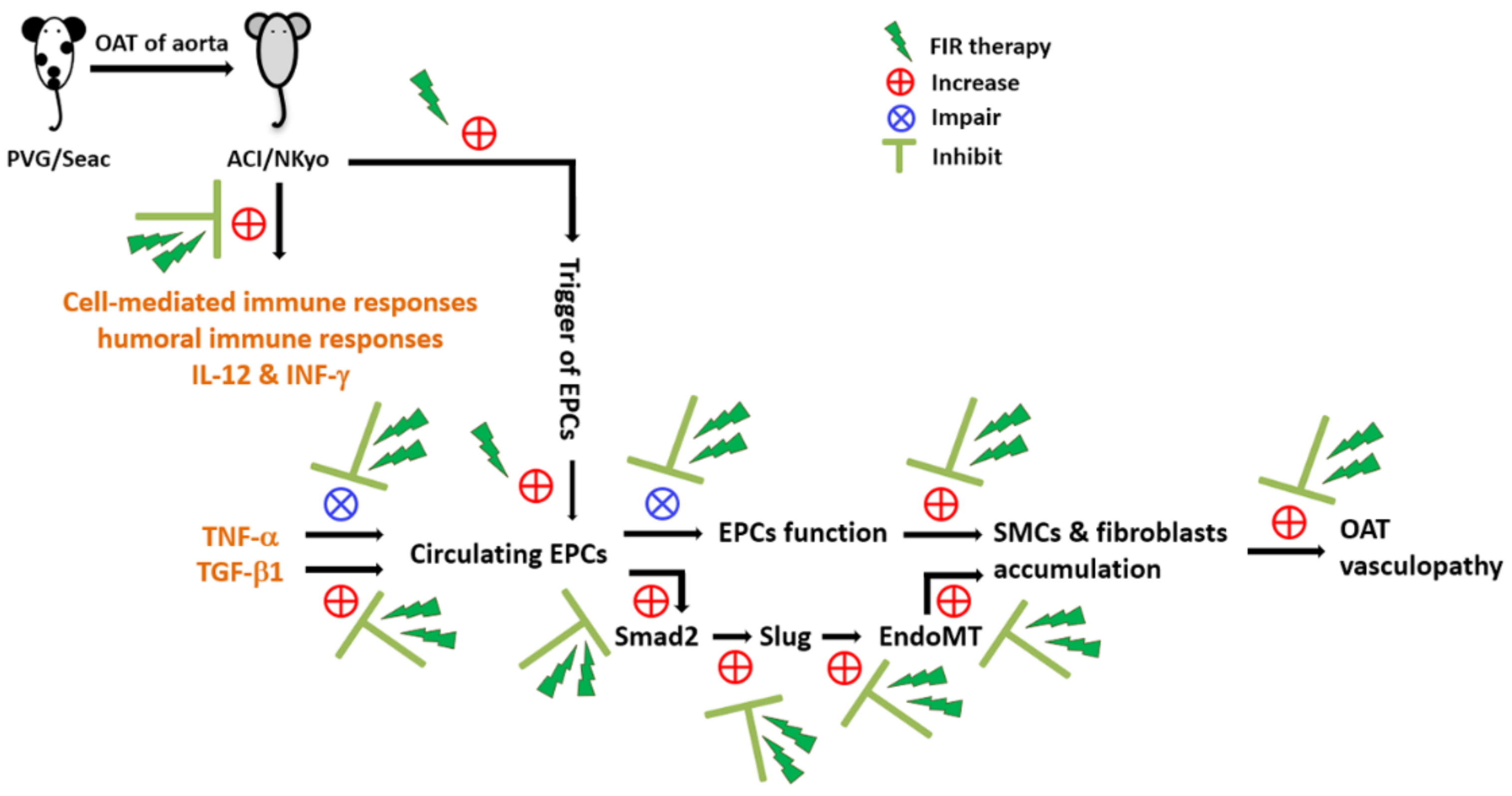
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sobajima, M.; Nozawa, T.; Ihori, H.; Shida, T.; Ohori, T.; Suzuki, T.; Matsuki, A.; Yasumura, S.; Inoue, H. Repeated sauna therapy improves myocardial perfusion in patients with chronically occluded coronary artery-related ische-mia. Int. J. Cardiol. 2013, 167, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Ohori, T.; Nozawa, T.; Ihori, H.; Shida, T.; Sobajima, M.; Matsuki, A.; Yasumura, S.; Inoue, H. Effect of repeated sauna treatment on exercise tolerance and endothelial function in patients with chronic heart failure. Am. J. Cardiol. 2012, 109, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chang, C.F.; Lai, M.Y.; Chen, T.W.; Lee, P.C.; Yang, W.C. Far-infrared therapy: A novel treatment to im-prove access blood flow and unassisted patency of arteriovenous fistula in hemodialysis patients. J. Am. Soc. Nephrol. 2007, 18, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.C.; Fang, H.C.; Mar, G.Y.; Liou, J.C.; Tseng, C.J.; Liu, C.P. Post-angioplasty far infrared radiation therapy improves 1-year angioplasty-free hemodialysis access patency of recurrent obstructive lesions. Eur. J. Vasc. Endovasc. Surg. 2013, 46, 726–732. [Google Scholar] [CrossRef]
- Tu, Y.P.; Chen, S.C.; Liu, Y.H.; Chen, C.F.; Hour, T.C. Postconditioning with far-infrared irradiation increases heme oxygenase-1 expression and protects against ischemia/reperfusion injury in rat testis. Life Sci. 2013, 92, 35–41. [Google Scholar] [CrossRef]
- Yu, S.Y.; Chiu, J.H.; Yang, S.D.; Hsu, Y.C.; Lui, W.Y.; Wu, C.W. Biological effect of far-infrared therapy on increas-ing skin microcirculation in rats. Photodermatol. Photoimmunol. Photomed. 2006, 22, 78–86. [Google Scholar] [CrossRef]
- Chen, T.Y.; Yang, Y.C.; Sha, Y.N.; Chou, J.R.; Liu, B.S. Far-Infrared Therapy Promotes Nerve Repair following End-to-End Neurorrhaphy in Rat Models of Sciatic Nerve Injury. Evid. Based Complement. Altern. Med. 2015, 2015, 207245. [Google Scholar] [CrossRef]
- Huang, P.H.; Chen, J.W.; Lin, C.P.; Chen, Y.H.; Wang, C.H.; Leu, H.B.; Lin, S.J. Far infra-red therapy promotes is-chemia-induced angiogenesis in diabetic mice and restores high glucose-suppressed endothelial progenitor cell functions. Cardiovasc. Diabetol. 2012, 11, 99. [Google Scholar] [CrossRef]
- Shui, S.; Wang, X.; Chiang, J.Y.; Zheng, L. Far-infrared therapy for cardiovascular, autoimmune, and other chronic health problems: A systematic review. Exp. Biol. Med. 2015, 240, 1257–1265. [Google Scholar] [CrossRef]
- Lund, L.H.; Edwards, L.B.; Kucheryavaya, A.Y.; Dipchand, A.I.; Benden, C.; Christie, J.D.; Dobbels, F.; Kirk, R.; Rahmel, A.O.; Yusen, R.D.; et al. The Registry of the International Society for Heart and Lung Transplantation: Thir-tieth Official Adult Heart Transplant Report--2013; focus theme: Age. J. Heart Lung Transplant. 2013, 32, 951–964. [Google Scholar] [CrossRef]
- Costello, J.P.; Mohanakumar, T.; Nath, D.S. Mechanisms of chronic cardiac allograft rejection. Tex. Heart Inst. J. 2013, 40, 395–399. [Google Scholar] [PubMed]
- Nath, D.S.; Basha, H.I.; Mohanakumar, T. Antihuman leukocyte antigen antibody-induced autoimmunity: Role in chronic rejection. Curr. Opin. Organ Transplant. 2010, 15, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.D. Overview of Immunosuppressive Therapy in Solid Organ Transplantation. Anesthesiol. Clin. 2017, 35, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Jasiak, N.M.; Park, J.M. Immunosuppression in Solid-Organ Transplantation: Essentials and Practical Tips. Crit. Care Nurs. Q. 2016, 39, 227–240. [Google Scholar] [CrossRef]
- Poston, R.S.; Billingham, M.; Hoyt, E.G.; Pollard, J.; Shorthouse, R.; Morris, R.E.; Robbins, R.C. Rapamycin reverses chronic graft vascular disease in a novel cardiac allograft model. Circulation 1999, 100, 67–74. [Google Scholar] [CrossRef]
- Bedi, D.S.; Riella, L.V.; Tullius, S.G.; Chandraker, A. Animal models of chronic allograft injury: Contributions and limitations to understanding the mechanism of long-term graft dysfunction. Transplantation 2010, 90, 935–944. [Google Scholar] [CrossRef]
- Chen, Y.H.; Lin, S.J.; Lin, F.Y.; Wu, T.C.; Tsao, C.R.; Huang, P.H.; Liu, P.L.; Chen, Y.L.; Chen, J.W. High glucose impairs early and late endothelial progenitor cells by modifying nitric oxide-related but not oxidative stress-mediated mechanisms. Diabetes 2007, 56, 1559–1568. [Google Scholar] [CrossRef]
- Chen, J.Z.; Zhu, J.H.; Wang, X.X.; Xie, X.D.; Sun, J.; Shang, Y.P.; Guo, X.G.; Dai, H.M.; Hu, S.J. Effects of homocys-teine on number and activity of endothelial progenitor cells from peripheral blood. J. Mol. Cell. Cardiol. 2004, 36, 233–239. [Google Scholar] [CrossRef]
- Goldstein, S. Replicative senescence: The human fibroblast comes of age. Science 1990, 249, 1129–1133. [Google Scholar] [CrossRef]
- Lin, F.Y.; Shih, C.M.; Huang, C.Y.; Tsai, Y.T.; Loh, S.H.; Li, C.Y.; Lin, C.Y.; Lin, Y.W.; Tsai, C.S. Dipeptidyl Pepti-dase-4 Inhibitor Decreases Allograft Vasculopathy Via Regulating the Functions of Endothelial Progenitor Cells in Normoglycemic Rats. Cardiovasc. Drugs Ther. 2021, 35, 1111–1127. [Google Scholar]
- Zou, H.; Yang, Y.; Gao, M.; Zhang, B.; Ming, B.; Sun, Y.; Chen, H.; Tang, X.; Chen, Z.; Xiong, P.; et al. HMGB1 is involved in chronic rejection of cardiac allograft via promoting inflammatory-like mDCs. Am. J. Transplant. 2014, 14, 1765–1777. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.T.; Mirza, A.B.; Zahid, R.; Haleem, A.; Al Hussaini, H.; Al Sulaiman, M.; Mousa, D. Antibody-mediated rejection: Importance of lactate dehydrogenase and neutrophilia in early diagnosis. Saudi J. Kidney Dis. Transplant. 2011, 22, 525–530. [Google Scholar]
- Kato, G.J.; Hebbel, R.P.; Steinberg, M.H.; Gladwin, M.T. Vasculopathy in sickle cell disease: Biology, pathophysi-ology, genetics, translational medicine, and new research directions. Am. J. Hematol. 2009, 84, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Rua, R.; McGavern, D.B. Elucidation of monocyte/macrophage dynamics and function by intravital imaging. J. Leukoc. Biol. 2015, 98, 319–332. [Google Scholar] [CrossRef]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef]
- Fadini, G.P.; Sartore, S.; Albiero, M.; Baesso, I.; Murphy, E.; Menegolo, M.; Grego, F.; Vigili de Kreutzenberg, S.; Tiengo, A.; Agostini, C.; et al. Number and function of endothelial progenitor cells as a marker of severity for dia-betic vasculopathy. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2140–2146. [Google Scholar] [CrossRef]
- Pintavorn, P.; Ballermann, B.J. TGF-beta and the endothelium during immune injury. Kidney Int. 1997, 51, 1401–1412. [Google Scholar] [CrossRef]
- Weis, M.; Wildhirt, S.M.; Schulze, C.; Pehlivanli, S.; Fraunberger, P.; Meiser, B.M.; von Scheidt, W. Modulation of coronary vasomotor tone by cytokines in cardiac transplant recipients. Transplantation 1999, 68, 1263–1267. [Google Scholar] [CrossRef]
- Del Papa, N.; Pignataro, F. The Role of Endothelial Progenitors in the Repair of Vascular Damage in Systemic Sclerosis. Front. Immunol. 2018, 9, 1383. [Google Scholar] [CrossRef]
- Tilling, L.; Chowienczyk, P.; Clapp, B. Progenitors in motion: Mechanisms of mobilization of endothelial progen-itor cells. Br. J. Clin. Pharmacol. 2009, 68, 484–492. [Google Scholar] [CrossRef]
- Oh, S.A.; Li, M.O. TGF-beta: Guardian of T cell function. J. Immunol. 2013, 191, 3973–3979. [Google Scholar] [CrossRef] [PubMed]
- Knight, R.J.; Liu, H.; Fishman, E.; Reis, E.D. Cold ischemic injury, aortic allograft vasculopathy, and pro-inflammatory cytokine expression. J. Surg. Res. 2003, 113, 201–207. [Google Scholar] [CrossRef]
- Kovacic, J.C.; Dimmeler, S.; Harvey, R.P.; Finkel, T.; Aikawa, E.; Krenning, G.; Baker, A.H. Endothelial to Mesen-chymal Transition in Cardiovascular Disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2019, 73, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.M.; Serbanovic-Canic, J.; Feng, S.; Souilhol, C.; Xing, R.; Hsiao, S.; Mammoto, A.; Chen, J.; Ariaans, M.; Francis, S.E.; et al. Shear stress induces endothelial-to-mesenchymal transition via the transcription factor Snail. Sci. Rep. 2017, 7, 3375. [Google Scholar] [CrossRef] [PubMed]
- Platel, V.; Faure, S.; Corre, I.; Clere, N. Endothelial-to-Mesenchymal Transition (EndoMT): Roles in Tumorigene-sis, Metastatic Extravasation and Therapy Resistance. J. Oncol. 2019, 2019, 8361945. [Google Scholar] [CrossRef] [PubMed]
- Tei, C. Waon therapy: Soothing warmth therapy. J. Cardiol. 2007, 49, 301–304. [Google Scholar] [PubMed]
- Tei, C.; Horikiri, Y.; Park, J.C.; Jeong, J.W.; Chang, K.S.; Toyama, Y.; Tanaka, N. Acute hemodynamic improve-ment by thermal vasodilation in congestive heart failure. Circulation 1995, 91, 2582–2590. [Google Scholar] [CrossRef]
- Akasaki, Y.; Miyata, M.; Eto, H.; Shirasawa, T.; Hamada, N.; Ikeda, Y.; Biro, S.; Otsuji, Y.; Tei, C. Repeated thermal therapy up-regulates endothelial nitric oxide synthase and augments angiogenesis in a mouse model of hindlimb ischemia. Circ. J. 2006, 70, 463–470. [Google Scholar] [CrossRef]
- Ise, N.; Katsuura, T.; Kikuchi, Y.; Miwa, E. Effect of far-infrared radiation on forearm skin blood flow. Ann. Physiol. Anthropol. 1987, 6, 31–32. [Google Scholar] [CrossRef]
- Kihara, T.; Biro, S.; Imamura, M.; Yoshifuku, S.; Takasaki, K.; Ikeda, Y.; Otuji, Y.; Minagoe, S.; Toyama, Y.; Tei, C. Repeated sauna treatment improves vascular endothelial and cardiac function in patients with chronic heart failure. J. Am. Coll. Cardiol. 2002, 39, 754–759. [Google Scholar] [CrossRef]
- Ryotokuji, K.; Ishimaru, K.; Kihara, K.; Namiki, Y.; Hozumi, N. Effect of pinpoint plantar long-wavelength infra-red light irradiation on subcutaneous temperature and stress markers. Laser Ther. 2013, 22, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Su, L.H.; Wu, K.D.; Lee, L.S.; Wang, H.; Liu, C.F. Effects of far infrared acupoint stimulation on autonomic activ-ity and quality of life in hemodialysis patients. Am. J. Chin. Med. 2009, 37, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Oosterveld, F.G.; Rasker, J.J.; Floors, M.; Landkroon, R.; van Rennes, B.; Zwijnenberg, J.; van de Laar, M.A.; Koel, G.J. Infrared sauna in patients with rheumatoid arthritis and ankylosing spondylitis. A pilot study showing good tolerance, short-term improvement of pain and stiffness, and a trend towards long-term beneficial effects. Clin. Rheumatol. 2009, 28, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Biro, S.; Kamogawa, Y.; Yoshifuku, S.; Eto, H.; Orihara, K.; Yu, B.; Kihara, T.; Miyata, M.; Hamasaki, S.; et al. Repeated sauna therapy increases arterial endothelial nitric oxide synthase expression and nitric oxide pro-duction in cardiomyopathic hamsters. Circ. J. 2005, 69, 722–729. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, S.; Cho, D.H.; Park, Y.M.; Kang, D.H.; Jo, I. Far-infrared radiation acutely increases nitric oxide production by increasing Ca(2+) mobilization and Ca(2+)/calmodulin-dependent protein kinase II-mediated phos-phorylation of endothelial nitric oxide synthase at serine 1179. Biochem. Biophys. Res. Commun. 2013, 436, 601–606. [Google Scholar] [CrossRef]
- Anggard, E. Nitric oxide: Mediator, murderer, and medicine. Lancet 1994, 343, 1199–1206. [Google Scholar] [CrossRef]
- Patrono, C.; FitzGerald, G.A. Isoprostanes: Potential markers of oxidant stress in atherothrombotic disease. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 2309–2315. [Google Scholar] [CrossRef]
- Malek, A.M.; Izumo, S.; Alper, S.L. Modulation by pathophysiological stimuli of the shear stress-induced up-regulation of endothelial nitric oxide synthase expression in endothelial cells. Neurosurgery 1999, 45, 334–344, discussion 344–335. [Google Scholar] [CrossRef]
- Aicher, A.; Heeschen, C.; Mildner-Rihm, C.; Urbich, C.; Ihling, C.; Technau-Ihling, K.; Zeiher, A.M.; Dimmeler, S. Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nat. Med. 2003, 9, 1370–1376. [Google Scholar] [CrossRef]
- Kuehbacher, A.; Urbich, C.; Zeiher, A.M.; Dimmeler, S. Role of Dicer and Drosha for endothelial microRNA expression and angiogenesis. Circ. Res. 2007, 101, 59–68. [Google Scholar] [CrossRef]
- Weber, M.; Baker, M.B.; Moore, J.P.; Searles, C.D. MiR-21 is induced in endothelial cells by shear stress and mod-ulates apoptosis and eNOS activity. Biochem. Biophys. Res. Commun. 2010, 393, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Ni, C.W.; Qiu, H.; Jo, H. MicroRNA-663 upregulated by oscillatory shear stress plays a role in inflammatory re-sponse of endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1762–H1769. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, V.; Zaccagnini, G.; Capogrossi, M.C.; Martelli, F. microRNAs as peripheral blood biomarkers of cardiovascular disease. Vascul. Pharmacol. 2011, 55, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Pei, F.; Zhu, X.; Duan, D.D.; Zeng, C. Circulating microRNAs as novel and sensitive biomarkers of acute myocardial Infarction. Clin. Biochem. 2012, 45, 727–732. [Google Scholar] [CrossRef]
- Qu, K.; Wang, Z.; Lin, X.L.; Zhang, K.; He, X.L.; Zhang, H. MicroRNAs: Key regulators of endothelial progenitor cell functions. Clin. Chim. Acta 2015, 448, 65–73. [Google Scholar] [CrossRef]
- Kaczmarek, I.; Deutsch, M.A.; Kauke, T.; Beiras-Fernandez, A.; Schmoeckel, M.; Vicol, C.; Sodian, R.; Reichart, B.; Spannagl, M.; Ueberfuhr, P. Donor-specific HLA alloantibodies: Long-term impact on cardiac allograft vasculopathy and mortality after heart transplant. Exp. Clin. Transplant. 2008, 6, 229–235. [Google Scholar]
- Weiss, M.J.; Madsen, J.C.; Rosengard, B.R.; Allan, J.S. Mechanisms of chronic rejection in cardiothoracic trans-plantation. Front. Biosci. 2008, 13, 2980–2988. [Google Scholar] [CrossRef]
- Benatti, R.D.; Taylor, D.O. Evolving concepts and treatment strategies for cardiac allograft vasculopathy. Curr. Treat. Options Cardiovasc. Med. 2014, 16, 278. [Google Scholar] [CrossRef]
- Skoric, B.; Cikes, M.; Ljubas Macek, J.; Baricevic, Z.; Skorak, I.; Gasparovic, H.; Biocina, B.; Milicic, D. Cardiac al-lograft vasculopathy: Diagnosis, therapy, and prognosis. Croat. Med. J. 2014, 55, 562–576. [Google Scholar] [CrossRef]
- Borthwick, L.A.; Parker, S.M.; Brougham, K.A.; Johnson, G.E.; Gorowiec, M.R.; Ward, C.; Lordan, J.L.; Corris, P.A.; Kirby, J.A.; Fisher, A.J. Epithelial to mesenchymal transition (EMT) and airway remodelling after human lung transplantation. Thorax 2009, 64, 770–777. [Google Scholar] [CrossRef]
- Chen, P.Y.; Qin, L.; Barnes, C.; Charisse, K.; Yi, T.; Zhang, X.; Ali, R.; Medina, P.P.; Yu, J.; Slack, F.J.; et al. FGF reg-ulates TGF-beta signaling and endothelial-to-mesenchymal transition via control of let-7 miRNA expression. Cell Rep. 2012, 2, 1684–1696. [Google Scholar] [CrossRef] [PubMed]
- Piera-Velazquez, S.; Jimenez, S.A. Molecular mechanisms of endothelial to mesenchymal cell transition (En-doMT) in experimentally induced fibrotic diseases. Fibrogenes. Tissue Repair 2012, 5, S7. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, D.A.; Kajstura, J.; Hosoda, T.; Gatti, A.; Bello, R.; Mosna, F.; Bardelli, S.; Zheng, H.; D’Amario, D.; Padin-Iruegas, M.E.; et al. Progenitor cells from the explanted heart generate immunocompatible myocardium within the transplanted donor heart. Circ. Res. 2009, 105, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Hillebrands, J.L.; Klatter, F.A.; Rozing, J. Origin of vascular smooth muscle cells and the role of circulating stem cells in transplant arteriosclerosis. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 380–387. [Google Scholar] [CrossRef][Green Version]
- Sathya, C.J.; Sheshgiri, R.; Prodger, J.; Tumiati, L.; Delgado, D.; Ross, H.J.; Rao, V. Correlation between circulating endothelial progenitor cell function and allograft rejection in heart transplant patients. Transpl. Int. 2010, 23, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Simper, D.; Wang, S.; Deb, A.; Holmes, D.; McGregor, C.; Frantz, R.; Kushwaha, S.S.; Caplice, N.M. Endothelial progenitor cells are decreased in blood of cardiac allograft patients with vasculopathy and endothelial cells of non-cardiac origin are enriched in transplant atherosclerosis. Circulation 2003, 108, 143–149. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| vWF | 5′-GGC TGC AGT ATG TCA AGG TGG-3′ | 5′-AGA GCC ATT GGT GCA GTG CAG-3′ |
| VE-cadherin | 5′-AGA CAA TGG GAT GCC AAG TCB-3′ | 5′-AAG ATG AGC AGG GTG ATC ACT G-3′ |
| αSMA | 5′-CTA TCA GGG GGC ACC ACT ATG-3′ | 5′-CCG ATC CAG ACA GAG TAT TTG CG-3′ |
| vimentin | 5′-AGG CAA AGC AGG AGT CCA CTG A-3′ | 5′-ATC TGG CGT TCC AGG GAC TCAT -3′ |
| GAPDH | 5′-TGC CCC CTC TGC TGA TGC C-3′ | 5′-CCT CCG ACG CCT GCT TCA CCA C-3′ |
| Sham Control | OAT | OAT+FIR Low Intensity | OAT+FIR High Intensity | |||||
|---|---|---|---|---|---|---|---|---|
| baseline | 90 days | baseline | 90 days | baseline | 90 days | baseline | 90 days | |
| Body weight (g) | 256.8 ± 12.8 | 340.2 ± 9.5 | 270.8 ± 10.8 | 341.6 ± 10.7 | 258.4 ± 7.6 | 345.8 ± 4.4 | 266.2 ± 16.4 | 346.0 ± 13.5 |
| BUN (mg/dL) | 26.5 ± 1.4 | 29.6 ± 2.4 | 30.4 ± 2.4 | 29.6 ± 1.7 | 28.7 ± 3.1 | 29.7 ± 2.3 | 30.5 ± 2.9 | 33.7 ± 2.3 |
| Creatinine (mg/dL) | 0.4 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.5 ± 0.2 | 0.5 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.2 | 0.4 ± 0.2 |
| ALT (IU/L) | 26.3 ± 1.6 | 26.0 ± 1.8 | 27.7 ± 1.8 | 28.8 ± 1.2 | 26.7 ± 1.1 | 28.0 ± 1.6 | 26.2 ± 0.9 | 26.4 ± 1.8 |
| AST (IU/L) | 34.4 ± 1.3 | 33.8 ± 1.3 | 34.4 ± 1.8 | 34.6 ± 1.4 | 33.9 ± 1.2 | 32.6 ± 1.7 | 35.2 ± 2.5 | 34.5 ± 1.7 |
| LDH (IU/L) | 786.6 ± 37.8 | 732.7 ± 20.3 | 750.8 ± 37.3 | 1166.2 ± 111.5 ab | 737.6 ± 26.6 | 1001.0 ± 142.0 ab | 757.1 ± 30.5 | 823.7 ± 57.4 c |
| HMGB1 (ng/mL) | 3.3 ± 0.7 | 3.4 ± 0.5 | 3.2 ± 0.5 | 127.5 ± 89.5 ab | 2.4 ± 0.9 | 36.5 ± 19.8 abc | 2.5 ± 0.5 | 35.0 ± 10.9 abc |
| Sham Control | OAT | OAT+FIR Low Intensity | OAT+FIR High Intensity | |||||
|---|---|---|---|---|---|---|---|---|
| Proteins | baseline | 90 days | baseline | 90 days | baseline | 90 days | baseline | 90 days |
| CRP (mg/dL) | 30.9 ± 6.1 | 33.5 ± 14.6 | 33.6 ± 12.5 | 39.4 ± 8.5 | 32.6 ± 6.7 | 32.0 ± 11.7 | 33.9 ± 13.1 | 27.9 ± 10.5 |
| SDF-1α (pg/mL) | 180.5 ± 42.4 | 192.2 ± 41.1 | 200.7 ± 50.8 | 376.8 ± 103.2 ab | 169.1 ± 63.9 | 367.2 ± 81.5 ab | 259.3 ± 95.5 | 444.2 ± 106.9 ab |
| TNF-α (pg/mL) | 68.2 ± 11.4 | 61.6 ± 9.8 | 59.0 ± 9.3 | 243.8 ± 65.2 ab | 62.0 ± 7.0 | 110.9 ± 44.2 abc | 70.1 ± 10.8 | 113.2 ± 47.7 c |
| TGF-β1 (ng/mL) | 50.6 ± 11.2 | 52.5 ± 10.8 | 46.6 ± 13.3 | 418.4 ± 102.6 ab | 41.6 ± 16.9 | 155.1 ± 54.9 abc | 39.0 ± 7.3 | 136.3 ± 54.6 abc |
| INF-γ (pg/mL) | 2.9 ± 0.8 | 2.9 ± 0.7 | 3.5 ± 1.0 | 23.2 ± 7.5 ab | 3.5 ± 1.3 | 15.5 ± 3.8 abc | 3.3 ± 1.5 | 4.7 ± 1.4 bc |
| IL-12 (pg/mL) | 195.5 ± 38.9 | 181.6 ± 32.1 | 202.5 ± 34.7 | 404.6 ± 88.7 ab | 176.7 ± 58.8 | 327.6 ± 60.3 ab | 191.3 ± 45.4 | 187.7 ± 49.9 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-W.; Tsai, C.-S.; Huang, C.-Y.; Tsai, Y.-T.; Shih, C.-M.; Lin, S.-J.; Li, C.-Y.; Lin, C.-Y.; Sung, S.-Y.; Lin, F.-Y. Far-Infrared Therapy Decreases Orthotopic Allograft Transplantation Vasculopathy. Biomedicines 2022, 10, 1089. https://doi.org/10.3390/biomedicines10051089
Lin Y-W, Tsai C-S, Huang C-Y, Tsai Y-T, Shih C-M, Lin S-J, Li C-Y, Lin C-Y, Sung S-Y, Lin F-Y. Far-Infrared Therapy Decreases Orthotopic Allograft Transplantation Vasculopathy. Biomedicines. 2022; 10(5):1089. https://doi.org/10.3390/biomedicines10051089
Chicago/Turabian StyleLin, Yi-Wen, Chien-Sung Tsai, Chun-Yao Huang, Yi-Ting Tsai, Chun-Ming Shih, Shing-Jong Lin, Chi-Yuan Li, Cheng-Yen Lin, Shih-Ying Sung, and Feng-Yen Lin. 2022. "Far-Infrared Therapy Decreases Orthotopic Allograft Transplantation Vasculopathy" Biomedicines 10, no. 5: 1089. https://doi.org/10.3390/biomedicines10051089
APA StyleLin, Y.-W., Tsai, C.-S., Huang, C.-Y., Tsai, Y.-T., Shih, C.-M., Lin, S.-J., Li, C.-Y., Lin, C.-Y., Sung, S.-Y., & Lin, F.-Y. (2022). Far-Infrared Therapy Decreases Orthotopic Allograft Transplantation Vasculopathy. Biomedicines, 10(5), 1089. https://doi.org/10.3390/biomedicines10051089