
Sodium Accumulation and Blood Capillary Rarefaction in the Skin Predispose Spontaneously Hypertensive Rats to Salt Sensitive Hypertension

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Protocol
2.3. Tissue Electrolyte Determination
2.4. Gene Expression Profiling
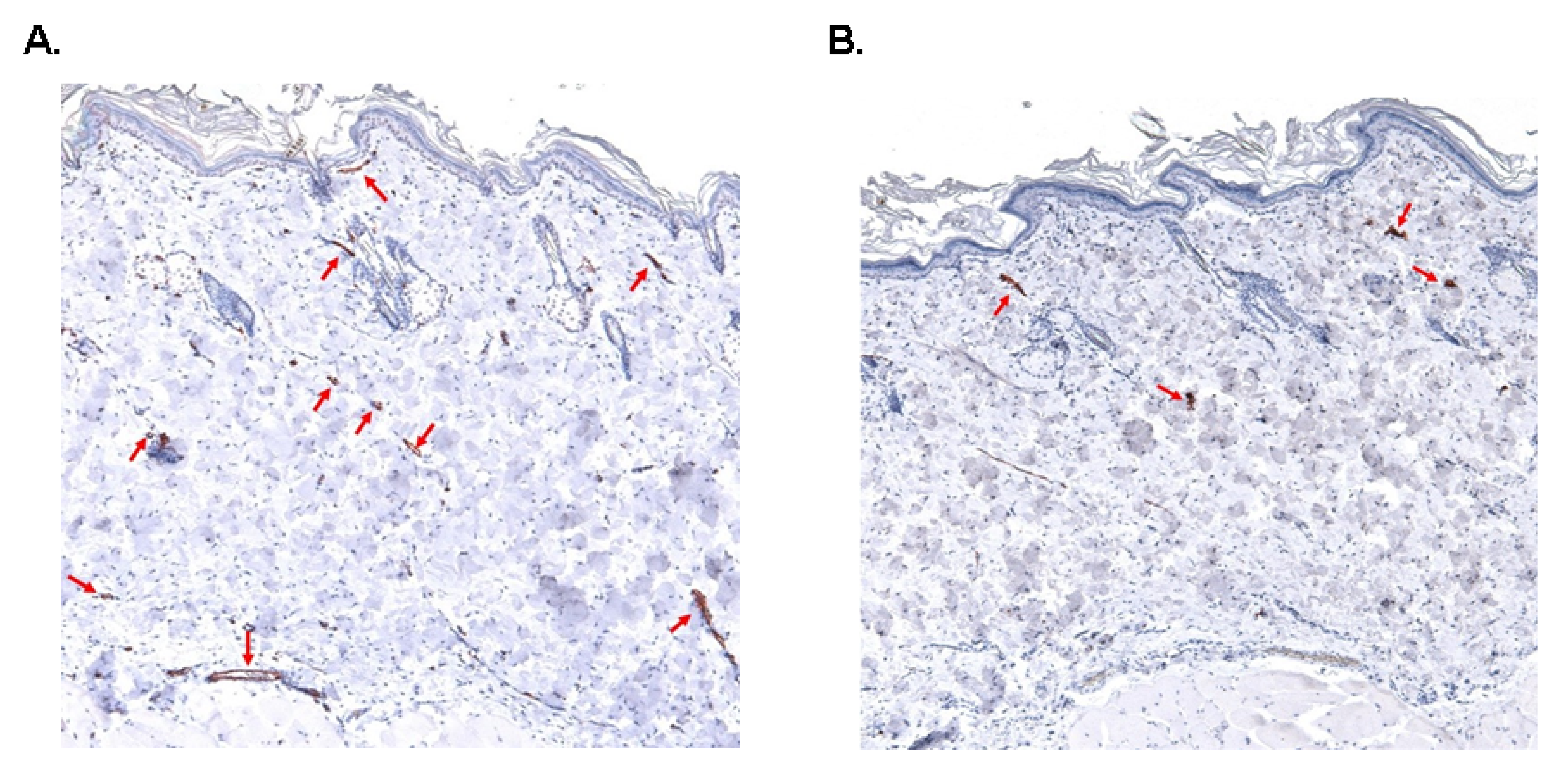
2.5. Number of Blood and Lymph Capillaries
2.6. Statistical Analysis
3. Results
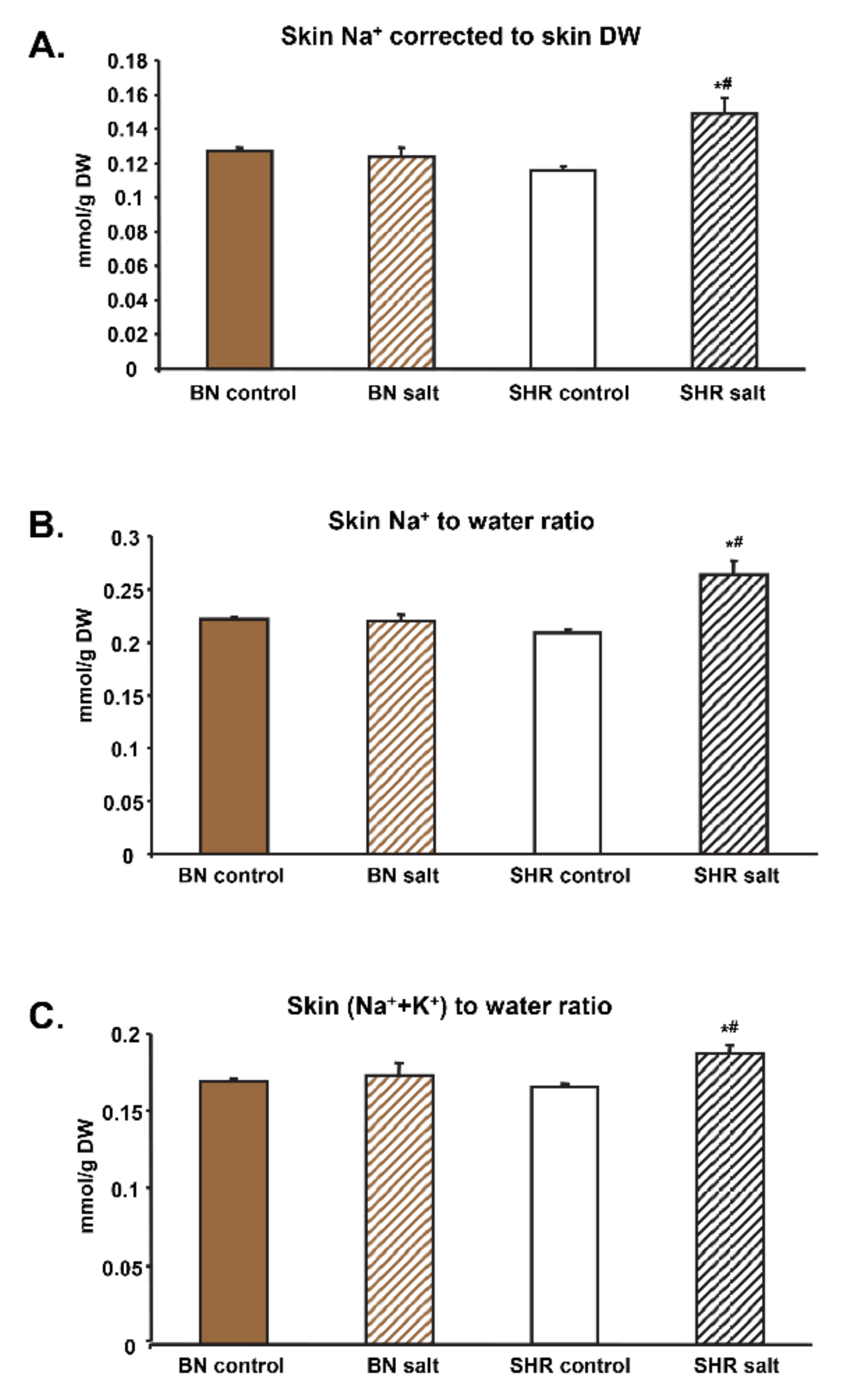
3.1. Tissue Electrolyte Concentrations and Water Content
3.2. Na+-to-Water, K+-to-Water and (Na++K+)-to-Water Ratios
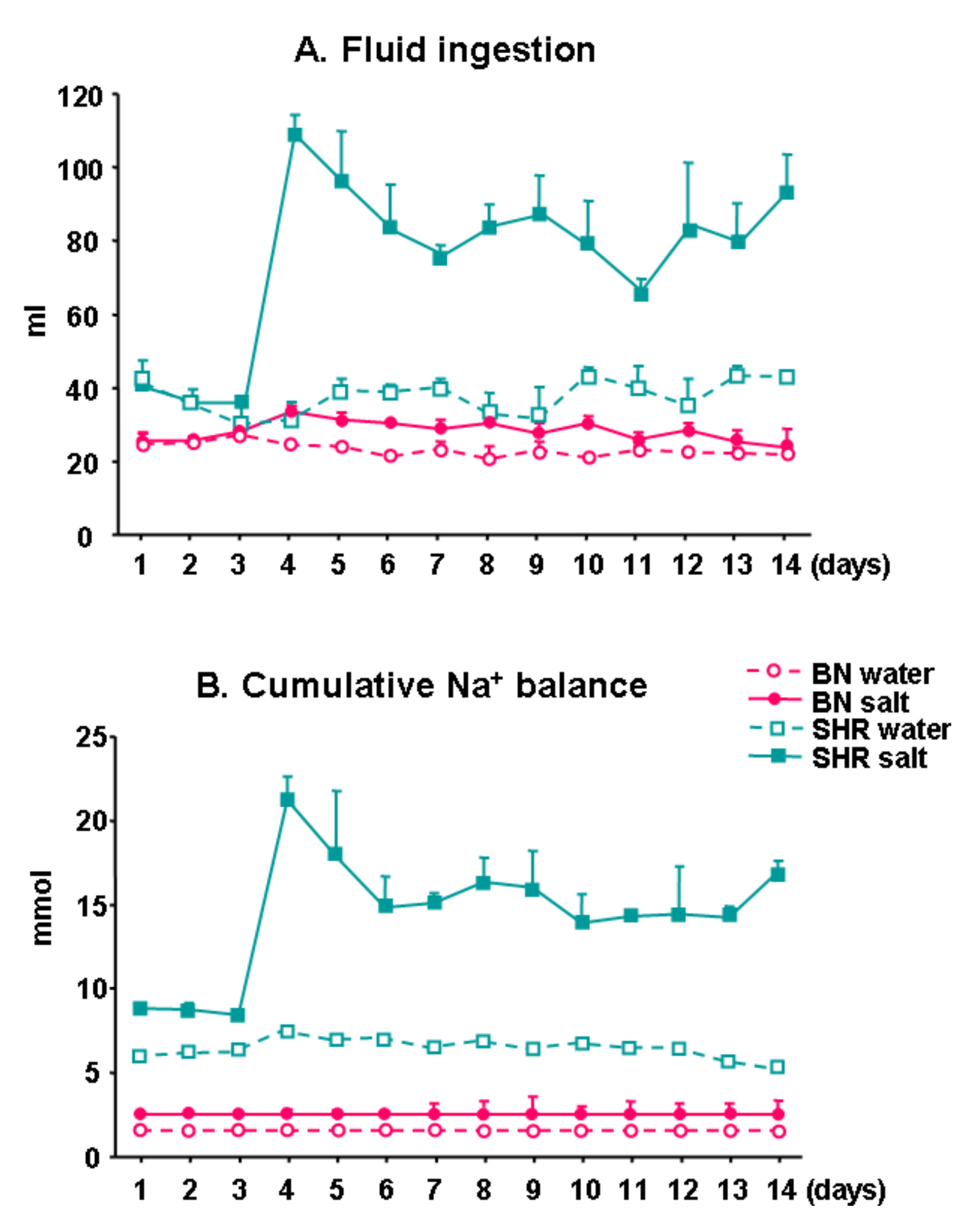
3.3. Fluid Ingestion and Cumulative Na+ Balance
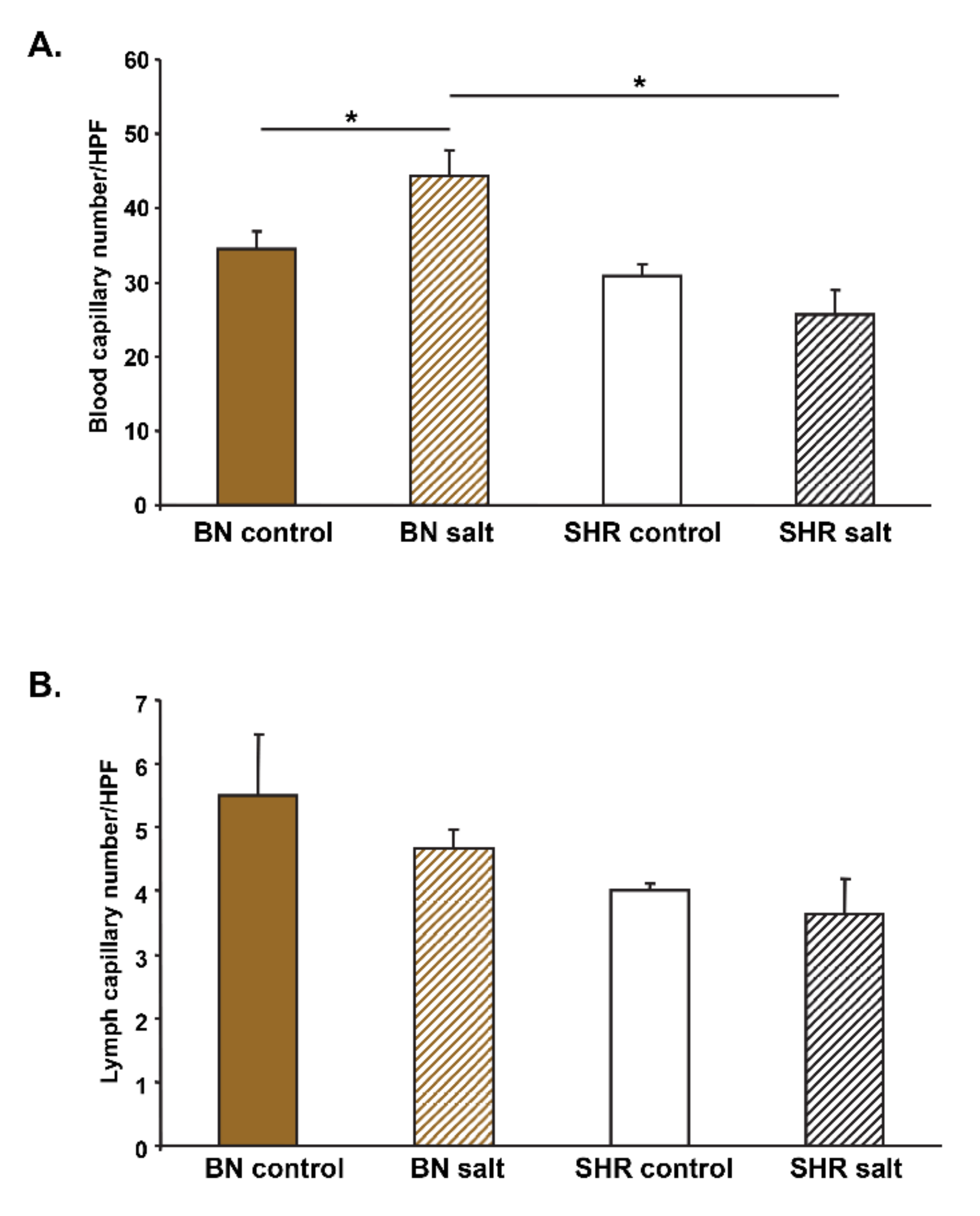
3.4. Skin Blood and Lymph Capillary Density
3.5. Gene Expression Profiles in the Skin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weinberger, M.H. Salt sensitivity of blood pressure in humans. Hypertension 1996, 27, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Kotchen, T.A.; Allen, W.; Cowley, A.W., Jr.; Frohlich, E.D. Salt in health and disease—A delicate balance. N. Engl. J. Med. 2016, 68, 1229–1237. [Google Scholar] [CrossRef]
- Barba, G.; Galletti, F.; Cappuccio, F.P.; Siani, A.; Venezia, A.; Versiero, M.; Della Valle, E.; Sorrentino, P.; Tarantino, G.; Farinaro, E.; et al. Incidence of hypertension in individuals with different blood pressure salt-sensitivity: Results of a 15-year follow-up study. J. Hypertens. 2007, 25, 1465–1471. [Google Scholar] [CrossRef] [PubMed]
- Titze, J.; Rittweger, J.; Dietsch, P.; Krause, H.; Schwind, K.H.; Engelke, K.; Lang, R.; Kirsch, K.A.; Luft, F.C.; Hilgers, K.F. Hypertension, sodium retention, calcium excretion and osteopenia in Dahl rats. J. Hypertens. 2004, 22, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Kopp, C.; Linz, P.; Dahlmann, A.; Hammon, M.; Jantsch, J.; Müller, D.N.; Schmieder, R.E.; Cavallaro, A.; Eckardt, K.U.; Uder, M.; et al. 23Na magnetic resonance imaging-determined tissue sodium in healthy subjects and hypertensive patients. Hypertension 2013, 61, 635–640. [Google Scholar] [CrossRef]
- Titze, J.; Dahlmann, A.; Lerchl, K.; Kopp, C.; Rakova, N.; Schröder, A.; Luft, F.C. Spooky sodium balance. Kidney Int. 2014, 85, 759–767. [Google Scholar] [CrossRef]
- Prasad, A.; Dunnill, G.S.; Mortimer, P.S.; MacGregor, G.A. Capillary rarefaction in the forearm skin in essential hypertension. J. Hypertens. 1995, 13, 265–268. [Google Scholar] [CrossRef]
- Antonios, T.F.; Singer, D.R.; Markandu, N.D.; Mortimer, P.S.; MacGregor, G.A. Rarefaction of skin capillaries in borderline essential hypertension suggests an early structural abnormality. Hypertension 1999, 34, 655–658. [Google Scholar] [CrossRef]
- Antonios, T.F.; Rattray, F.M.; Singer, D.R.; Markandu, N.D.; Mortimer, P.S.; MacGregor, G.A. Rarefaction of skin capillaries in normotensive offspring of individuals with essential hypertension. Heart 2003, 89, 175–178. [Google Scholar] [CrossRef]
- He, F.J.; Marciniak, M.; Markandu, N.D.; Antonios, T.F.; MacGregor, G.A. Effect of modest salt reduction on skin capillary rarefaction in white, black, and Asian individuals with mild hypertension. Hypertension 2010, 56, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Churchill, P.C.; Churchill, M.C.; Bidani, A.K.; Griffin, K.A.; Picken, M.; Pravenec, M.; Kren, V.; St Lezin, E.; Wang, J.M.; Wang, N.; et al. Genetic susceptibility to hypertension-induced renal damage in the rat. Evidence based on kidney-specific genome transfer. J. Clin. Investig. 1997, 100, 1373–1382. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liška, F.; Mancini, M.; Krupková, M.; Chylíková, B.; Křenová, D.; Šeda, O.; Šilhavý, J.; Mlejnek, P.; Landa, V.; Zídek, V.; et al. Plzf as a candidate gene predisposing the spontaneously hypertensive rat to hypertension, left ventricular hypertrophy, and interstitial fibrosis. Am. J. Hypertens. 2014, 27, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Liška, F.; Landa, V.; Zídek, V.; Mlejnek, P.; Šilhavý, J.; Šimáková, M.; Strnad, H.; Trnovská, J.; Škop, V.; Kazdová, L.; et al. Downregulation of Plzf gene ameliorates metabolic and cardiac traits in the spontaneously hypertensive rat. Hypertension 2017, 69, 1084–1091. [Google Scholar] [CrossRef]
- Lunova, M.; Kubovciak, J.; Smolková, B.; Uzhytchak, M.; Michalova, K.; Dejneka, A.; Strnad, P.; Lunov, O.; Jirsa, M. Expression of interferons lambda 3 and 4 induces identical response in human liver cell lines depending exclusively on canonical signaling. Int. J. Mol. Sci. 2021, 22, 2560. [Google Scholar] [CrossRef] [PubMed]
- Rossitto, G.; Mary, S.; Chen, J.Y.; Boder, P.; Chew, K.S.; Neves, K.B.; Alves, R.L.; Montezano, A.C.; Welsh, P.; Petrie, M.C.; et al. Tissue sodium excess is not hypertonic and reflects extracellular volume expansion. Nat. Commun. 2020, 11, 4222. [Google Scholar] [CrossRef] [PubMed]
- Titze, J.; Lang, R.; Ilies, C.; Schwind, K.H.; Kirsch, K.A.; Dietsch, P.; Luft, F.; Hilgers, K.F. Osmotically inactive skin Na+ storage in rats. Am. J. Physiol. Ren. Physiol. 2003, 285, F1108–F1117. [Google Scholar] [CrossRef]
- Titze, J.; Bauer, K.; Schafflhuber, M.; Dietsch, P.; Lang, R.; Schwind, K.H.; Luft, F.C.; Eckardt, K.-U.; Hilgers, K.F. Internal sodium balance in DOCA-salt rats: A body composition study. Am. J. Physiol. Ren. Physiol. 2005, 289, F793–F802. [Google Scholar] [CrossRef]
- Chrysant, S.G.; Walsh, G.M.; Kem, D.C.; Frohlich, E.D. Hemodynamic and metabolic evidence of salt sensitivity in spontaneously hypertensive rats. Kidney Int. 1979, 15, 33–37. [Google Scholar] [CrossRef]
- Flynn, F.W.; Culver, B.; Newton, S.V. Salt intake by normotensive and spontaneously hypertensive rats: Two-bottle and lick rate analyses. Physiol. Behav. 2003, 78, 689–696. [Google Scholar] [CrossRef]
- McConnell, S.D.; Henkin, R.I. Na,Cl preference in spontaneously hypertensive rats; age and blood pressure effects. Am. J. Physiol. 1973, 225, 624–627. [Google Scholar] [CrossRef]
- Di Nicolantonio, R.; Kren, V.; Zidek, V.; Pravenec, M. Salt preference of congenic strains derived from the spontaneously hypertensive rat. Physiol. Behav. 2004, 80, 617–622. [Google Scholar] [CrossRef]
- Hansen-Smith, F.M.; Morris, L.W.; Greene, A.S.; Lombard, J.H. Rapid microvessel rarefaction with elevated salt intake and reduced renal mass hypertension in rats. Circ. Res. 1996, 79, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Greene, A.S.; Tonellato, P.J.; Lui, J.; Lombard, J.H.; Cowley, A.W., Jr. Microvascular rarefaction and tissue vascular resistance in hypertension. Am. J. Physiol. 1989, 256 Pt 2, H126–H131. [Google Scholar] [CrossRef] [PubMed]
- Greene, A.S.; Lombard, J.H.; Cowley, A.W., Jr.; Hansen-Smith, F.M. Microvessel changes in hypertension measured by Griffonia simplicifolia I lectin. Hypertension 1990, 15 Pt 2, 779–783. [Google Scholar] [CrossRef]
- Triantafyllou, A.; Anyfanti, P.; Pyrpasopoulou, A.; Triantafyllou, G.; Aslanidis, S.; Douma, S. Capillary rarefaction as an index for the microvascular assessment of hypertensive patients. Curr. Hypertens. Rep. 2015, 17, 33. [Google Scholar] [CrossRef] [PubMed]
- Markiewski, M.M.; Daugherity, E.; Reese, B.; Karbowniczek, M. The role of complement in angiogenesis. Antibodies 2020, 9, 67. [Google Scholar] [CrossRef]
- Silverman, G.A.; Whisstock, J.C.; Bottomley, S.P.; Huntington, J.A.; Kaiserman, D.; Luke, C.J.; Pak, S.C.; Reichhart, J.-M.; Bird, P.I. Serpins flex their muscle: I. Putting the clamps on proteolysis in diverse biological systems. J. Biol. Chem. 2010, 285, 24299–24305. [Google Scholar] [CrossRef] [PubMed]
- Juráňová, J.; Franková, J.; Ulrichová, J. The role of keratinocytes in inflammation. J. Appl. Biomed. 2017, 15, 169–179. [Google Scholar] [CrossRef]
- Sivaprasad, U.; Kinker, K.G.; Ericksen, M.B.; Lindsey, M.; Gibson, A.M.; Bass, S.A.; Hershey, N.S.; Deng, J.; Medvedovic, M.; Hershey, G.K.K. SERPINB3/B4 contributes to early inflammation and barrier dysfunction in an experimental murine model of atopic dermatitis. J. Investig. Derm. 2015, 135, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Machnik, A.; Neuhofer, W.; Jantsch, J.; Dahlmann, A.; Tammela, T.; Machura, K.; Park, J.K.; Beck, F.X.; Müller, D.N.; Derer, W.; et al. Macrophages regulate salt-dependent volume and blood pressure by a vascular endothelial growth factor-C-dependent buffering mechanism. Nat. Med. 2009, 15, 545–552. [Google Scholar] [CrossRef]
- Machnik, A.; Dahlmann, A.; Kopp, C.; Goss, J.; Wagner, H.; van Rooijen, N.; Eckardt, K.U.; Müller, D.N.; Park, J.K.; Luft, F.C.; et al. Mononuclear phagocyte system depletion blocks interstitial tonicity-responsive enhancer binding protein/vascular endothelial growth factor C expression and induces salt-sensitive hypertension in rats. Hypertension 2010, 55, 755–761. [Google Scholar] [CrossRef]
- Wiig, H.; Schröder, A.; Neuhofer, W.; Jantsch, J.; Kopp, C.; Karlsen, T.V.; Boschmann, M.; Goss, J.; Bry, M.; Rakova, N.; et al. Immune cells control skin lymphatic electrolyte homeostasis and blood pressure. J. Clin. Investig. 2013, 123, 2803–2815. [Google Scholar] [CrossRef] [PubMed]
- Rendell, M.S.; Finnegan, M.F.; Healy, J.C.; Lind, A.; Milliken, B.K.; Finney, D.E.; Bonner, R.F. The relationship of laser-Doppler skin blood flow measurements to the cutaneous microvascular anatomy. Microvasc. Res. 1998, 55, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Pravenec, M.; Klír, P.; Kren, V.; Zicha, J.; Kunes, J. An analysis of spontaneous hypertension in spontaneously hypertensive rats by means of new recombinant inbred strains. J. Hypertens. 1989, 7, 217–221. [Google Scholar] [CrossRef]
- Kurtz, T.W.; DiCarlo, S.E.; Pravenec, M.; Schmidlin, O.; Tanaka, M.; Morris, R.C., Jr. An alternative hypothesis to the widely held view that renal excretion of sodium accounts for resistance to salt-induced hypertension. Kidney Int. 2016, 90, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, T.W.; Dicarlo, S.E.; Pravenec, M.; Morris, R.C., Jr. The pivotal role of renal vasodysfunction in salt sensitivity and the initiation of salt-induced hypertension. Curr. Opin. Nephrol. Hypertens. 2018, 27, 83–92. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | BN-Lx Control | BN-Lx Salt | SHR Control | SHR Salt | Source of Variation (p) | ||
|---|---|---|---|---|---|---|---|
| Strain | Salt | Strain × Salt Interaction | |||||
| Body WW (skin WW + Carcass WW) (g) | 228 ± 10 | 209 ± 4 | 308 ± 3 | 303 ± 2 | <0.001 | 0.067 | 0.253 |
| Body DW (g) (Carcass DW + Skin DW) | 76 ± 3 | 71 ± 3 | 109 ± 1 | 106 ± 2 | <0.001 | 0.102 | 0.664 |
| Carcass WW (g) | 186 ± 8 | 168 ± 3 | 235 ± 1 | 228 ± 2 | <0.001 | 0.032 | 0.246 |
| Carcass DW (g) | 58 ± 2 | 53 ± 2 | 77 ± 1 | 74 ± 2 | <0.001 | 0.050 | 0.571 |
| Carcass water (g) | 128 ± 6 | 115 ± 1 | 158 ± 1 | 155 ± 1 | <0.001 | 0.036 | 0.168 |
| Carcass relative water (mL/g WW) | 0.687 ± 0.003 | 0.685 ± 0.007 | 0.674 ± 0.002 | 0.678 ± 0.004 | 0.046 | 0.788 | 0.438 |
| Skin WW (g) | 43 ± 1 | 41 ± 1 | 72 ± 2 | 74 ± 1 | <0.001 | 0.964 | 0.428 |
| Skin DW (g) | 18 ± 2 | 18 ± 4 | 32 ± 4 | 32 ± 4 | <0.001 | 0.752 | 0.986 |
| Skin water (g) | 24 ± 1 | 23 ± 1 | 40 ± 2 | 41 ± 1 | <0.001 | 0.843 | 0.230 |
| Skin relative water (mL/g WW) | 0.571 ± 0.006 | 0.565 ± 0.009 | 0.552 ± 0.006 | 0.563 ± 0.009 | 0.204 | 0.770 | 0.310 |
| rTBW (mL/g WW) | 1.258 ± 0.009 | 1.250 ± 0.015 | 1.226 ± 0.005 | 1.241 ± 0.013 | 0.108 | 0.765 | 0.329 |
| Bone ash (g) | 7.6 ± 0.3 | 7.1 ± 0.1 | 8.6 ± 0.2 | 8.2 ± 0.1 | <0.001 | 0.059 | 0.982 |
| rTB Na+ (mmol/g DW) | 0.220 ± 0.005 | 0.213 ± 0.011 | 0.192 ± 0.005 | 0.239 ± 0.011 # | 0.938 | 0.045 | 0.013 |
| rTB K+ (mmol/g DW) | 0.293 ± 0.010 | 0.277 ± 0.018 | 0.246 ± 0.007 | 0.263 ± 0.010 | 0.030 | 0.974 | 0.202 |
| rTB Cl− (mmol/g DW) | 0.110 ± 0.027 | 0.072 ± 0.005 | 0.088 ± 0.011 | 0.123 ± 0.008 ‡ | 0.376 | 0.934 | 0.040 |
| rCarcass Na+ (mmol/g DW) | 0.093 ± 0.002 | 0.089 ± 0.006 | 0.077 ± 0.003 † | 0.090 ± 0.002 # | 0.070 | 0.220 | 0.036 |
| rCarcass K+ (mmol/g DW) | 0.197 ± 0.009 | 0.166 ± 0.009 | 0.158 ± 0.005 | 0.171 ± 0.004 | 0.026 | 0.726 | 0.091 |
| rCarcass Cl− (mmol/g DW) | 0.015 ± 0.001 | 0.013 ± 0.001 | 0.015 ± 0.001 | 0.021 ± 0.001 #‡ | 0.004 | 0.026 | 0.002 |
| rSkin Na+ (mmol/g DW) | 0.127 ± 0.002 | 0.124 ± 0.005 | 0.116 ± 0.002 | 0.149 ± 0.009 #‡ | 0.268 | 0.025 | 0.012 |
| rSkin K+ (mmol/g DW) | 0.096 ± 0.001 | 0.100 ± 0.005 | 0.088 ± 0.002 | 0.092 ± 0.006 | 0.078 | 0.422 | 0.950 |
| rSkin Cl− (mmol/g DW) | 0.070 ± 0.009 | 0.059 ± 0.004 | 0.073 ± 0.010 | 0.102 ± 0.007 ‡ | 0.370 | 0.978 | 0.037 |
| rBone Na+ (mmol/g carcass DW) | 0.068 ± 0.001 | 0.072 ± 0.001 | 0.064 ± 0.003 | 0.065 ± 0.001 | 0.173 | 0.775 | 0.156 |
| rBone K+ (mmol/g carcass DW) | 0.055 ± 0.004 | 0.070 ± 0.005 | 0.040 ± 0.003 | 0.038 ± 0.001 | 0.003 | 0.056 | 0.234 |
| rBone Cl− (mmol/g carcass DW) | 0.019 ± 0.001 | 0.020 ± 0.001 * | 0.018 ± 0.001 † | 0.019 ± 0.001 ‡ | <0.001 | 0.103 | 0.046 |
| Serum Na+ (mmol/L) | 141.9 ± 0.3 | 143.2 ± 0.6 | 143.2 ± 0.6 | 143.4 ± 0.8 | 0.245 | 0.265 | 0.381 |
| Serum K+ (mmol/L) | 5.2 ± 0.1 | 5.0 ± 0.1 | 5.2 ± 0.1 | 4.9 ± 0.1 ‡ | 0.524 | 0.009 | 0.294 |
| Serum Cl− (mmol/L) | 105.2 ± 0.8 | 106.6 ± 0.7 | 103.7 ± 0.7 | 105.2 ± 0.9 | 0.094 | 0.104 | 0.935 |
| Traits | BN-Lx Control | BN-Lx Salt | SHR Control | SHR Salt | Source of Variation (p) | ||
|---|---|---|---|---|---|---|---|
| Strain | Salt | Strain × Salt Interaction | |||||
| Na+-to-water ratio, mmol/mL | |||||||
| TB Na+/TB W | 0.051 ± 0.001 | 0.050 ± 0.001 | 0.049 ± 0.002 | 0.058 ± 0.001 #‡ | 0.032 | 0.005 | 0.002 |
| Carcass Na+/Carcass W | 0.042 ± 0.001 | 0.041 ± 0.001 | 0.037 ± 0.002 † | 0.043 ± 0.001 # | 0.229 | 0.132 | 0.017 |
| Skin Na+/Skin W | 0.095 ± 0.001 | 0.096 ± 0.001 | 0.094 ± 0.001 | 0.115 ± 0.004 #‡ | 0.003 | <0.001 | 0.001 |
| Bone Na+/TB W | 0.026 ± 0.001 | 0.028 ± 0.001 | 0.026 ± 0.002 | 0.024 ± 0.003 | 0.146 | 0.900 | 0.089 |
| K+-to-water ratio, mmol/mL | |||||||
| TB K+/TB W | 0.087 ± 0.002 | 0.081 ± 0.003 | 0.073 ± 0.008 † | 0.079 ± 0.004 | 0.005 | 0.924 | 0.018 |
| Carcass K+/Carcass W | 0.090 ± 0.003 | 0.082 ± 0.004 * | 0.074 ± 0.005 † | 0.081 ± 0.001 | 0.011 | 0.804 | 0.015 |
| Skin K+/Skin W | 0.073 ± 0.001 | 0.077 ± 0.002 * | 0.071 ± 0.001 | 0.070 ± 0.002 ‡ | 0.010 | 0.337 | 0.046 |
| Bone K+/TB W | 0.020 ± 0.001 | 0.027 ± 0.003 * | 0.016 ± 0.001 | 0.014 ± 0.001 ‡ | <0.001 | 0.068 | 0.019 |
| (Na++K+)-to-water ratio, mmol/mL | |||||||
| TB (Na++K+)/TB W | 0.138 ± 0.003 | 0.131 ± 0.004 | 0.124 ± 0.004 † | 0.137 ± 0.001 # | 0.293 | 0.397 | 0.016 |
| Carcass (Na++K+)/Carcass W | 0.132 ± 0.004 | 0.122 ± 0.005 | 0.114 ± 0.005 † | 0.124 ± 0.001 | 0.064 | 0.958 | 0.035 |
| Skin (Na++K+)/Skin W | 0.168 ± 0.002 | 0.172 ± 0.008 | 0.165 ± 0.002 | 0.186 ± 0.005 #‡ | 0.085 | 0.002 | 0.018 |
| Bone (Na++K+)/TB W | 0.046 ± 0.001 | 0.054 ± 0.003 * | 0.042 ± 0.002 | 0.039 ± 0.001 ‡ | 0.002 | 0.226 | 0.025 |
| Symbol | Name | logFC | Adjusted p Value |
|---|---|---|---|
| BP: Positive regulation of angiogenesis, p = 8.7 × 10−7 | |||
| Ereg | epiregulin | 3.39 | 0.0018 |
| Vegfd | vascular endothelial growth factor D | 1.60 | 0.0075 |
| Angpt4 | angiopoietin 4 | 2.86 | 0.0018 |
| Ccl11 | C-C motif chemokine ligand 11 | 2.06 | 0.0067 |
| Sfrp2 | secreted frizzled-related protein 2 | 3.80 | 0.00015 |
| F3 | coagulation factor III, tissue factor | 2.04 | 0.000027 |
| C6 | complement C6 | 2.05 | 0.0073 |
| BP: Positive regulation of endothelial cell proliferation, p = 1.30 × 10−4 | |||
| Vegfd | vascular endothelial growth factor D | 1.60 | 0.0075 |
| Ccl11 | C-C motif chemokine ligand 11 | 2.06 | 0.0067 |
| F3 | coagulation factor III, tissue factor | 2.04 | 0.000027 |
| Wnt2 | Wnt family member 2 | 2.30 | 0.0049 |
| BP: Complement activation, p = 1.90 × 10−5 | |||
| C6 | complement C6 | 2.05 | 0.0073 |
| Cfh | complement factor H | 1.84 | 0.025 |
| C7 | complement C7 | 0.0001 | 0.030 |
| BP: Cellular response to interferon-gamma, p = 4.34 × 10−4 | |||
| Ccl11 | C-C motif chemokine ligand 11 | 2.06 | 0.0067 |
| Mrc1 | mannose receptor, C type 1 | 1.59 | 0.029 |
| Ccl6 | chemokine (C-C motif) ligand 6 | 2.12 | 0.0056 |
| Cfh | complement factor H | 1.84 | 0.025 |
| BP: Cellular response to interleukin-1, p = 5.84 × 10−4 | |||
| Smpd3 | sphingomyelin phosphodiesterase 3 | 2.35 | 0.0049 |
| Ccl11 | C-C motif chemokine ligand 11 | 2.06 | 0.0067 |
| Fn1 | fibronectin 1 | 2.59 | 0.00051 |
| Ccl6 | chemokine (C-C motif) ligand 6 | 2.12 | 0.0056 |
| BP: Chemokine-mediated signaling pathway, p = 7.25 × 10−4 | |||
| Ccr1 | C-C motif chemokine receptor 1 | 2.19 | 0.0075 |
| Ccl11 | C-C motif chemokine ligand 11 | 2.06 | 0.0067 |
| Ccl6 | chemokine (C-C motif) ligand 6 | 2.12 | 0.0056 |
| BP: Chronic inflammatory response, p = 7.28 × 10−4 | |||
| Ccl11 | C-C motif chemokine ligand 11 | 2.06 | 0.0067 |
| Vnn1 | vanin 1 | 2.10 | 0.0018 |
| Symbol | Name | logFC | Adjusted p Value |
|---|---|---|---|
| BP: Keratinocyte differentiation, epidermis development, p = 1.69 × 10−7 | |||
| Lc1m | late cornified envelope 1M | 1.75 | 0.025 |
| Lce1f | late cornified envelope 1F | 1.62 | 0.036 |
| Krt1 | keratin 1 | 2.76 | 0.000003 |
| Krt10 | keratin 10 | 2.98 | 0.000013 |
| Lce1l | late cornified envelope 1L | 2.29 | 0.00015 |
| Krtdap | keratinocyte differentiation associated protein | 1.32 | 0.040 |
| BP: Negative regulation of peptidase activity, p = 2.85 × 10−4 | |||
| Serpinb3a | serine (or cysteine) peptidase inhibitor | 4.24 | 0.000027 |
| Spli | secretory leukocyte peptidase inhibitor | 1.02 | 0.036 |
| Slpil3 | antileukoproteinase-like 3 | 3.02 | 0.0015 |
| BP: Neutrophil chemotaxis, p = 2.77 × 10−6 | |||
| Il1b | interleukin 1, beta | 1.93 | 0.011 |
| Il36g | interleukin 36, gamma | 2.50 | 0.0024 |
| Il36rn | interleukin 36 receptor antagonist | 1.45 | 0.040 |
| Cxcr2 | C-X-C motif chemokine receptor 2 | 1.56 | 0.013 |
| Ccl22 | C-C motif chemokine ligand 22 | 1.87 | 0.0072 |
| Il36b | interleukin 36, beta | 1.81 | 0.031 |
| BP: Establishment of skin barrier, p = 1.83 × 10−4 | |||
| Alox12b | arachidonate 12-lipoxygenase, 12R type | 2.43 | 0.0028 |
| Krt1 | keratin 1 | 2.76 | 0.000003 |
| Flg2 | filaggrin family member 2 | 2.87 | 0.00002 |
| BP: Negative regulation of endopeptidase activity, p = 6.44 × 10−6 | |||
| Serpinb12 | serpin family B member 12 | 2.98 | 0.000004 |
| Stfa3 | stefin A3 | 2.99 | 2.89e-10 |
| Serpina12 | serpin family A member 12 | 2.12 | 0.0029 |
| Stfa2 | stefin A2 | 2.47 | 0.00017 |
| BP: Regulation of signaling receptor activity, p = 9.08 × 10−4 | |||
| Btc | betacellulin | 3.39 | 0.00058 |
| Il1b | interleukin 1, beta | 1.93 | 0.011 |
| Il36g | interleukin 36, gamma | 2.50 | 0.0024 |
| Il36rn | interleukin 36 receptor antagonist | 1.45 | 0.040 |
| Il18 | interleukin 18 | 1.96 | 0.013 |
| Ccl22 | C-C motif chemokine ligand 22 | 1.87 | 0.0072 |
| Il36b | interleukin 36, beta | 1.81 | 0.031 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šilhavý, J.; Mlejnek, P.; Šimáková, M.; Liška, F.; Kubovčiak, J.; Sticová, E.; Pravenec, M. Sodium Accumulation and Blood Capillary Rarefaction in the Skin Predispose Spontaneously Hypertensive Rats to Salt Sensitive Hypertension. Biomedicines 2022, 10, 376. https://doi.org/10.3390/biomedicines10020376
Šilhavý J, Mlejnek P, Šimáková M, Liška F, Kubovčiak J, Sticová E, Pravenec M. Sodium Accumulation and Blood Capillary Rarefaction in the Skin Predispose Spontaneously Hypertensive Rats to Salt Sensitive Hypertension. Biomedicines. 2022; 10(2):376. https://doi.org/10.3390/biomedicines10020376
Chicago/Turabian StyleŠilhavý, Jan, Petr Mlejnek, Miroslava Šimáková, František Liška, Jan Kubovčiak, Eva Sticová, and Michal Pravenec. 2022. "Sodium Accumulation and Blood Capillary Rarefaction in the Skin Predispose Spontaneously Hypertensive Rats to Salt Sensitive Hypertension" Biomedicines 10, no. 2: 376. https://doi.org/10.3390/biomedicines10020376
APA StyleŠilhavý, J., Mlejnek, P., Šimáková, M., Liška, F., Kubovčiak, J., Sticová, E., & Pravenec, M. (2022). Sodium Accumulation and Blood Capillary Rarefaction in the Skin Predispose Spontaneously Hypertensive Rats to Salt Sensitive Hypertension. Biomedicines, 10(2), 376. https://doi.org/10.3390/biomedicines10020376