
Markers of Bronchiolitis Obliterans Syndrome after Lung Transplant: Between Old Knowledge and Future Perspective

 ,
,  ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
Molecular and Immunological Mechanisms Involved in the Pathogenesis of BOS
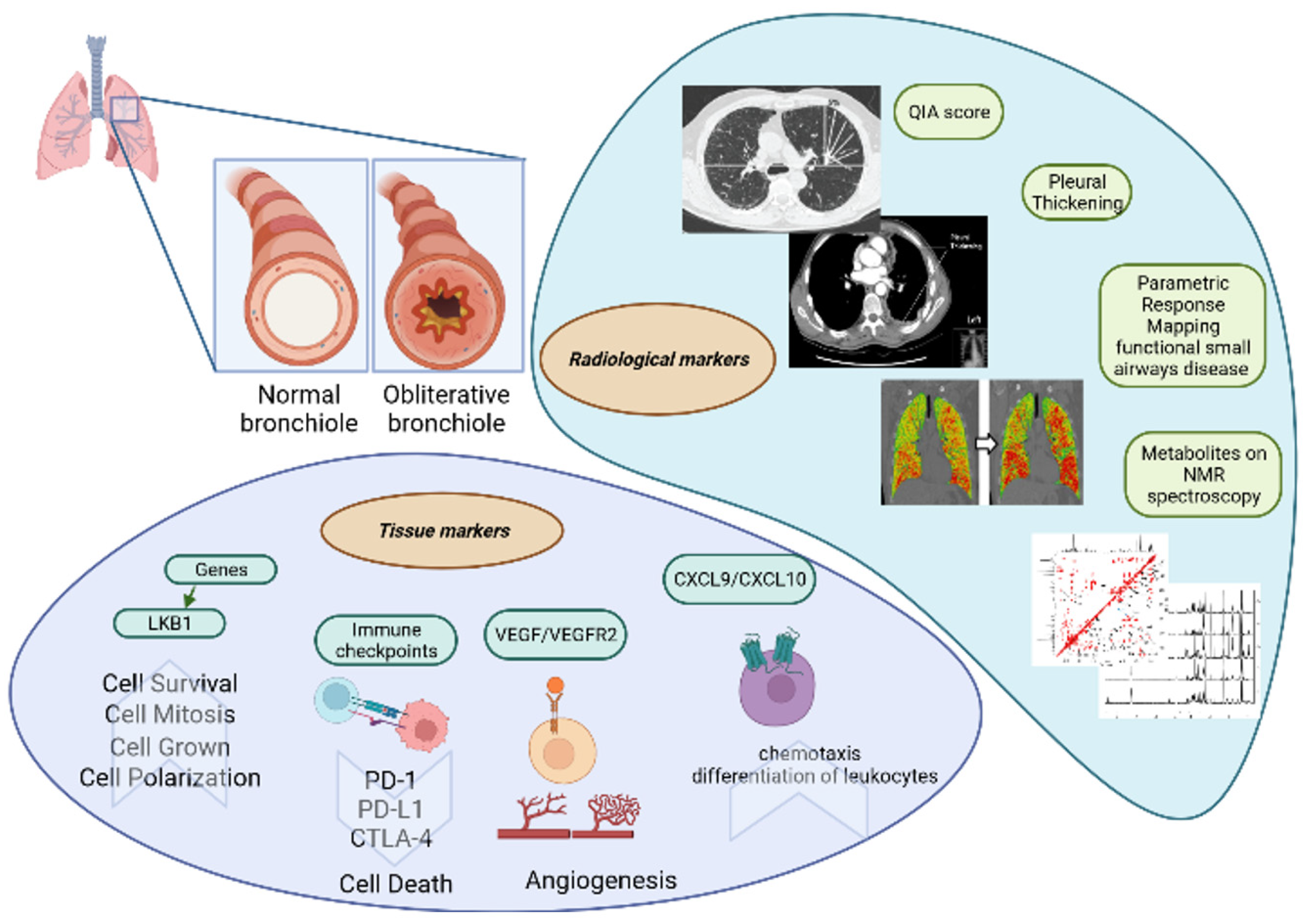
2. Tissue Markers
2.1. Liver Kinase B1 Gene
2.2. Checkpoint Molecules
2.3. VEGF/VEGFR2
2.4. CXCL9 and CXCL10
3. Exhaled Breath
3.1. Nitric Oxide
3.2. Exhaled Surfactant Protein A
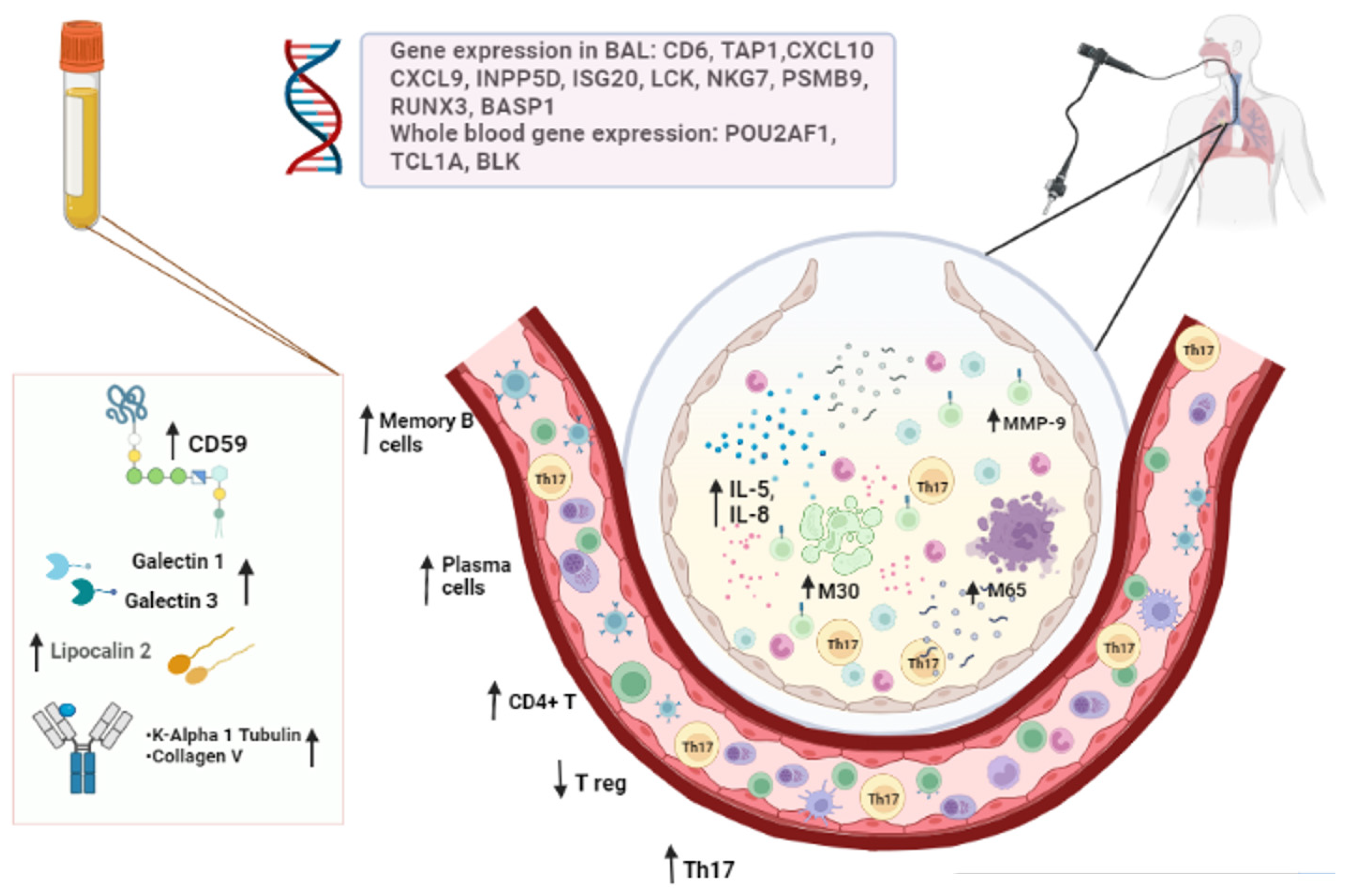
4. Circulating Biomarkers
4.1. Serum/Plasma Biomarkers
4.1.1. Lipocalin 2
4.1.2. Galectins 1 and 3
4.1.3. Soluble CD59
4.1.4. MMP-3
4.1.5. MMP-9
4.1.6. Self-Antigens (SAgs): K-Alpha 1 Tubulin and Collagen-V
4.2. Bronchoalveolar Lavage Fluid Biomarkers
4.2.1. Epithelial Cell Death Markers in BAL Fluid
4.2.2. Humoral and Cell Immunity Biomarkers in BAL Fluid
4.2.3. Cytokines and Chemokines in BAL Fluid
4.2.4. Gene Expression in BAL Fluid
4.2.5. NMR Spectroscopic Detection of Metabolites in BAL Fluid
5. Extracellular Vesicles
5.1. LKB1 from Tissue
5.2. miRNAs
6. Whole Blood and BAL Cell Subsets
6.1. B Cells
6.2. T Cells
6.3. Monocyte-Macrophage Lineage
6.4. Whole Blood Gene Expression Profiles
7. Cell Culture Supernatants
Matrix Metalloproteinase 9
8. Radiological Markers
8.1. HRCT Quantitative Image Analysis Score
8.2. Pleural Thickening Evaluated by Lung Ultrasound
8.3. Parametric Response Mapping of Functional Small Airway Disease by CT
9. Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chambers, D.C.; Cherikh, W.S.; Harhay, M.O.; Hayes, D.; Hsich, E.; Khush, K.K.; Meiser, B.; Potena, L.; Rossano, J.W.; Toll, A.E. The International Thoracic Organ Transplant Registry of the International Society for Heart and Lung Transplantation: Thirty-sixth adult lung and heart–lung transplantation Report—2019, Focus theme: Donor and recipient size match. J. Heart Lung Transplant. 2019, 38, 1042–1055. [Google Scholar] [CrossRef]
- Martinu, T.; Chen, D.-F.; Palmer, S.M. Acute rejection and humoral sensitization in lung transplant recipients. Proc. Am. Thorac. Soc. 2009, 6, 54–65. [Google Scholar] [CrossRef]
- Meyer, K.C.; Raghu, G.; Verleden, G.M.; Corris, P.A.; Aurora, P.; Wilson, K.C.; Brozek, J.; Glanville, A.R.; the ISHLT/ATS/ERS BOS Task Force Committee. An international ISHLT/ATS/ERS clinical practice guideline: Diagnosis and management of bronchiolitis obliterans syndrome. Eur. Respir. J. 2014, 44, 1479–1503. [Google Scholar] [CrossRef]
- Levy, L.; Huszti, E.; Renaud-Picard, B.; Berra, G.; Kawashima, M.; Takahagi, A.; Fuchs, E.; Ghany, R.; Moshkelgosha, S.; Keshavjee, S. Risk assessment of chronic lung allograft dysfunction phenotypes: Validation and proposed refinement of the 2019 International Society for Heart and Lung Transplantation classification system. J. Heart Lung Transplant. 2020, 39, 761–770. [Google Scholar] [CrossRef]
- Verleden, G.M.; Vos, R.; Verleden, S.E.; De Wever, W.; De Vleeschauwer, S.I.; Willems-Widyastuti, A.; Scheers, H.; Dupont, L.J.; Van Raemdonck, D.E.; Vanaudenaerde, B.M. Survival determinants in lung transplant patients with chronic allograft dysfunction. Transplantation 2011, 92, 703–708. [Google Scholar] [CrossRef]
- d’Alessandro, M.; Perillo, F.; Metella Refini, R.; Bergantini, L.; Bellisai, F.; Selvi, E.; Cameli, P.; Manganelli, S.; Conticini, E.; Cantarini, L.; et al. Efficacy of baricitinib in treating rheumatoid arthritis: Modulatory effects on fibrotic and inflammatory biomarkers in a real-life setting. Int. Immunopharmacol. 2020, 86, 106748. [Google Scholar] [CrossRef]
- Markopoulou, K.D.; Cool, C.D.; Elliot, T.L.; Lynch, D.A.; Newell, J.; Hale, V.A.; Brownz, K.K.; Schwarz, M.I.; Tuder, R.M. Obliterative bronchiolitis: Varying presentations and clinicopathological correlation. Eur. Respir. J. Eur. Respir. Soc. 2002, 19, 20–30. [Google Scholar] [CrossRef]
- Aguilar, P.R.; Michelson, A.P.; Isakow, W. Obliterative Bronchiolitis. Transplantation 2016, 100, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Todd, J.L.; Palmer, S.M. Bronchiolitis Obliterans Syndrome: The Final Frontier for Lung Transplantation. Chest 2011, 140, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Verleden, S.E.; von der Thusen, J.; Roux, A.; Brouwers, E.S.; Braubach, P.; Kuehnel, M.; Laenger, F.; Jonigk, D. When tissue is the issue: A histological review of chronic lung allograft dysfunction. Am. J. Transplant. 2020, 20, 2644–2651. [Google Scholar] [CrossRef] [PubMed]
- King, T.E. Bronchiolitis obliterans. Lung 1989, 167, 69–93. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; d’Alessandro, M.; Otranto, A.; Cavallaro, D.; Gangi, S.; Fossi, A.; Perillo, F.; Luzzi, L.; Zanfrini, E.; Paladini, P.; et al. Characterization of NKG2-A/-C, Kir and CD57 on NK Cells Stimulated with pp65 and IE-1 Antigens in Patients Awaiting Lung Transplant. Life 2022, 12, 1081. [Google Scholar] [CrossRef] [PubMed]
- Daga, S.; Fallerini, C.; Baldassarri, M.; Fava, F.; Valentino, F.; Doddato, G.; Benetti, E.; Furini, S.; Giliberti, A.; Tita, R.; et al. Employing a systematic approach to biobanking and analyzing clinical and genetic data for advancing COVID-19 research. Eur. J. Hum. Genet. 2021, 29, 745–759. [Google Scholar] [CrossRef]
- Hanif, Z.; Sufiyan, N.; Patel, M.; Akhtar, M.Z. Role of biobanks in transplantation. Ann. Med. Surg. 2018, 28, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Arjuna, A.; Olson, M.T.; Walia, R.; Bremner, R.M.; Smith, M.A.; Mohanakumar, T. An update on current treatment strategies for managing bronchiolitis obliterans syndrome after lung transplantation. Expert Rev. Respir. Med. 2021, 15, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; Bargagli, E.; d’Alessandro, M.; Refini, R.M.; Cameli, P.; Galasso, L.; Scapellato, C.; Montagnani, F.; Scolletta, S.; Franchi, F.; et al. Prognostic bioindicators in severe COVID-19 patients. Cytokine. 2021, 141, 155455. [Google Scholar] [CrossRef] [PubMed]
- Vietri, L.; Cameli, P.; Perruzza, M.; Cekorja, B.; Bergantini, L.; d’Alessandro, M.; Refini, R.M.; Pieroni, M.; Fossi, A.; Bennett, D.; et al. Pirfenidone in idiopathic pulmonary fibrosis: Real-life experience in the referral centre of Siena. Ther. Adv. Respir. Dis. 2020, 14, 1753466620906326. [Google Scholar] [CrossRef]
- d’Alessandro, M.; Bergantini, L.; Cameli, P.; Lanzarone, N.; Antonietta Mazzei, M.; Alonzi, V.; Sestini, P.; Bargagli, E. Serum KL-6 levels in pulmonary Langerhans’ cell histiocytosis. Eur. J. Clin. Investig. 2020, 50, e13242. [Google Scholar] [CrossRef]
- Vietri, L.; Bargagli, E.; Bennett, D.; Fossi, A.; Cameli, P.; Bergantini, L.; d’Alessandro, M.; Paladini, P.; Luzzi, L.; Gentili, F.; et al. Serum Amyloid A in lung transplantation. Sarcoidosis Vasc. Diffuse. Lung Dis. 2020, 37, 2–7. [Google Scholar]
- Vietri, L.; Bennett, D.; Cameli, P.; Bergantini, L.; Cillis, G.; Sestini, P.; Bargagli, E.; Rottoli, P. Serum amyloid A in patients with idiopathic pulmonary fibrosis. Respir. Investig. 2019, 57, 430–434. [Google Scholar] [CrossRef]
- d’Alessandro, M.; Carleo, A.; Cameli, P.; Bergantini, L.; Perrone, A.; Vietri, L.; Lanzarone, N.; Vagaggini, C.; Sestini, P.; Bargagli, E. BAL biomarkers’ panel for differential diagnosis of interstitial lung diseases. Clin. Exp. Med. 2020, 20, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.; Salvini, M.; Fui, A.; Cillis, G.; Cameli, P.; Mazzei, M.A.; Fossi, A.; Refini, R.M.; Rottoli, P. Calgranulin B and KL-6 in Bronchoalveolar Lavage of Patients with IPF and NSIP. Inflammation 2019, 42, 463–470. [Google Scholar] [CrossRef]
- Bargagli, E.; Olivieri, C.; Nikiforakis, N.; Cintorino, M.; Magi, B.; Perari, M.G.; Vagaggini, C.; Spina, D.; Prasse, A.; Rottoli, P. Analysis of macrophage migration inhibitory factor (MIF) in patients with idiopathic pulmonary fibrosis. Respir. Physiol. Neurobiol. 2009, 167, 261–267. [Google Scholar] [CrossRef]
- Rottoli, P.; Bargagli, E. Is bronchoalveolar lavage obsolete in the diagnosis of interstitial lung disease? Curr. Opin. Pulm. Med. 2003, 9, 418–425. [Google Scholar] [CrossRef]
- Hardison, M.T.; Galin, F.S.; Calderon, C.E.; Djekic, U.V.; Parker, S.B.; Wille, K.M.; Jackson, P.L.; Oster, R.A.; Young, K.R.; Blalock, J.E.; et al. The presence of a matrix-derived neutrophil chemoattractant in bronchiolitis obliterans syndrome after lung transplantation. J. Immunol. 2009, 182, 4423–4431. [Google Scholar] [CrossRef]
- Laohaburanakit, P.; Chan, A.; Allen, R.P. Bronchiolitis obliterans. Clin. Rev. Allergy Immunol. 2003, 25, 259–274. [Google Scholar] [CrossRef]
- Verleden, S.E.; Vasilescu, D.M.; Willems, S.; Ruttens, D.; Vos, R.; Vandermeulen, E.; Hostens, J.; McDonough, J.E.; Verbeken, E.K.; Verschakelen, J.; et al. The site and nature of airway obstruction after lung transplantation. Am. J. Respir. Crit. Care Med. 2014, 189, 292–300. [Google Scholar] [CrossRef]
- Kuehnel, M.; Maegel, L.; Vogel-Claussen, J.; Robertus, J.L.; Jonigk, D. Airway remodelling in the transplanted lung. Cell Tissue Res. 2017, 367, 663–675. [Google Scholar] [CrossRef]
- Hodge, G.; Hodge, S.; Holmes-Liew, C.-L.; Reynolds, P.N.; Holmes, M. Bronchiolitis obliterans syndrome is associated with increased peripheral blood natural killer and natural killer T-like granzymes, perforin, and T-helper-type 1 pro-inflammatory cytokines. J. Heart Lung Transplant. 2012, 31, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Hodge, S.; Hodge, G.; Ahern, J.; Liew, C.; Hopkins, P.; Chambers, D.C.; Reynolds, P.N.; Holmes, M. Increased levels of T cell granzyme b in bronchiolitis obliterans syndrome are not suppressed adequately by current immunosuppressive regimens. Clin. Exp. Immunol. 2009, 158, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.J.; Wilkes, D.S. The role of autoimmunity in obliterative bronchiolitis after lung transplantation. Am. J. Physiol. Lung Cell Mol. Physiol. 2013, 304, L307–L311. [Google Scholar] [CrossRef] [PubMed]
- Kleinewietfeld, M.; Hafler, D.A. The plasticity of human Treg and Th17 cells and its role in autoimmunity. Semin. Immunol. 2013, 25, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Verleden, S.E.; Ruttens, D.; Vandermeulen, E.; Bellon, H.; Van Raemdonck, D.E.; Dupont, L.J.; Vanaudenaerde, B.M.; Verleden, G.; Vos, R. Restrictive chronic lung allograft dysfunction: Where are we now? J. Heart Lung Transplant. 2015, 34, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, A.; Smith, M.A.; Phelan, D.; Sundaresan, S.; Trulock, E.P.; Lynch, J.P.; Cooper, J.D.; Patterson, G.A.; Mohanakumar, T. Development of ELISA-detected anti-HLA antibodies precedes the development of bronchiolitis obliterans syndrome and correlates with progressive decline in pulmonary function after lung transplantation. Transplantation 1999, 67, 1155–1161. [Google Scholar] [CrossRef]
- Kauke, T.; Kneidinger, N.; Martin, B.; Dick, A.; Schneider, C.; Schramm, R.; Meimarakis, G.; Preissler, G.; Eickelberg, O.; Von Dossow, V.; et al. Bronchiolitis obliterans syndrome due to donor-specific HLA-antibodies. Tissue Antigens 2015, 86, 178–185. [Google Scholar] [CrossRef]
- Borthwick, L.A.; Sunny, S.S.; Oliphant, V.; Perry, J.; Brodlie, M.; Johnson, G.E.; Ward, C.; Gould, K.; Corris, P.A.; De Soyza, A.; et al. Pseudomonas aeruginosa accentuates epithelial-to-mesenchymal transition in the airway. Eur. Respir. J. 2011, 37, 1237–1247. [Google Scholar] [CrossRef]
- Gregson, A.L.; Wang, X.; Weigt, S.S.; Palchevskiy, V.; Lynch, J.P.; Ross, D.J.; Kubak, B.M.; Saggar, R.; Fishbein, M.C.; Ardehali, A.; et al. Interaction between Pseudomonas and CXC chemokines increases risk of bronchiolitis obliterans syndrome and death in lung transplantation. Am. J. Respir. Crit. Care Med. 2013, 187, 518–526. [Google Scholar] [CrossRef]
- Rahman, M.; Ravichandran, R.; Bansal, S.; Sanborn, K.; Bowen, S.; Eschbacher, J.; Sureshbabu, A.; Fleming, T.; Bharat, A.; Walia, R.; et al. Novel role for tumor suppressor gene, liver kinase B1, in epithelial-mesenchymal transition leading to chronic lung allograft dysfunction. Am. J. Transplant. 2022, 22, 843–852. [Google Scholar] [CrossRef]
- Weigt, S.S.; Kim, G.-H.J.; Jones, H.D.; Ramsey, A.L.; Amubieya, O.; Abtin, F.; Pourzand, L.; Lee, J.; Shino, M.Y.; DerHovanessian, A.; et al. Quantitative Image Analysis at Chronic Lung Allograft Dysfunction Onset Predicts Mortality. Transplantation 2021, 106, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Arjuna, A.; Perincheri, S.; Poulson, C.; Bremner, R.M.; Smith, M.A.; Tokman, S.; Mohanakumar, T. Restrictive allograft syndrome vs bronchiolitis obliterans syndrome: Immunological and molecular characterization of circulating exosomes. J. Heart Lung Transplant. 2022, 41, 24–33. [Google Scholar] [CrossRef]
- Davidsen, J.R.; Laursen, C.B.; Højlund, M.; Lund, T.K.; Jeschke, K.N.; Iversen, M.; Kalhauge, A.; Bendstrup, E.; Carlsen, J.; Perch, M.; et al. Lung Ultrasound to Phenotype Chronic Lung Allograft Dysfunction in Lung Transplant Recipients. A Prospective Observational Study. J. Clin. Med. 2021, 10, 1078. [Google Scholar] [CrossRef] [PubMed]
- Veraar, C.; Kliman, J.; Benazzo, A.; Oberndorfer, F.; Laggner, M.; Hacker, P.; Raunegger, T.; Janik, S.; Jaksch, P.; Klepetko, W.; et al. Potential novel biomarkers for chronic lung allograft dysfunction and azithromycin responsive allograft dysfunction. Sci. Rep. 2021, 11, 6799. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; D’Alessandro, M.; De Vita, E.; Perillo, F.; Fossi, A.; Luzzi, L.; Paladini, P.; Perrone, A.; Rottoli, P.; Sestini, P.; et al. Regulatory and Effector Cell Disequilibrium in Patients with Acute Cellular Rejection and Chronic Lung Allograft Dysfunction after Lung Transplantation: Comparison of Peripheral and Alveolar Distribution. Cells 2021, 10, 780. [Google Scholar] [CrossRef]
- Itabashi, Y.; Ravichandran, R.; Bansal, S.; Bharat, A.; Hachem, R.; Bremner, R.; Smith, M.; Mohanakumar, T. Decline in Club Cell Secretory Proteins, Exosomes Induction and Immune Responses to Lung Self-antigens, Kα1 Tubulin and Collagen V, Leading to Chronic Rejection After Human Lung Transplantation. Transplantation 2021, 105, 1337–1346. [Google Scholar] [CrossRef]
- D’alessandro, M.; Bergantini, L.; Fossi, A.; De Vita, E.; Perillo, F.; Luzzi, L.; Paladini, P.; Sestini, P.; Rottoli, P.; Bargagli, E.; et al. The Role of Galectins in Chronic Lung Allograft Dysfunction. Lung 2021, 199, 281–288. [Google Scholar] [CrossRef]
- Righi, I.; Vaira, V.; Morlacchi, L.C.; Croci, G.A.; Rossetti, V.; Blasi, F.; Ferrero, S.; Nosotti, M.; Rosso, L.; Clerici, M. Immune Checkpoints Expression in Chronic Lung Allograft Rejection. Front. Immunol. 2021, 12, 714132. [Google Scholar] [CrossRef]
- Cameli, P.; Bargagli, E.; Fossi, A.; Bennett, D.; Voltolini, L.; Refini, R.M.; Gotti, G.; Rottoli, P. Exhaled nitric oxide and carbon monoxide in lung transplanted patients. Respir. Med. 2015, 109, 1224–1229. [Google Scholar] [CrossRef][Green Version]
- Durand, M.; Lacoste, P.; Danger, R.; Jacquemont, L.; Brosseau, C.; Durand, E.; Tilly, G.; Loy, J.; Foureau, A.; Royer, P.-J.; et al. High circulating CD4+CD25hiFOXP3+ T-cell sub-population early after lung transplantation is associated with development of bronchiolitis obliterans syndrome. J. Heart Lung Transplant. 2018, 37, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Danger, R.; Royer, P.-J.; Reboulleau, D.; Durand, E.; Loy, J.; Tissot, A.; Lacoste, P.; Roux, A.; Reynaud-Gaubert, M.; Gomez, C.; et al. Blood Gene Expression Predicts Bronchiolitis Obliterans Syndrome. Front. Immunol. 2017, 8, 1841. [Google Scholar] [CrossRef] [PubMed]
- Piloni, D.; Morosini, M.; Magni, S.; Balderacchi, A.; Scudeller, L.; Cova, E.; Oggionni, T.; Stella, G.; Tinelli, C.; Antonacci, F.; et al. Analysis of long term CD4+CD25highCD127- T-reg cells kinetics in peripheral blood of lung transplant recipients. BMC Pulm. Med. 2017, 17, 102. [Google Scholar] [CrossRef]
- Budding, K.; Rossato, M.; van de Graaf, E.A.; Kwakkel-van Erp, J.M.; Radstake TR, D.J.; Otten, H.G. Serum miRNAs as potential biomarkers for the bronchiolitis obliterans syndrome after lung transplantation. Transpl. Immunol. 2017, 42, 1–4. [Google Scholar] [CrossRef]
- Pain, M.; Royer, P.-J.; Loy, J.; Girardeau, A.; Tissot, A.; Lacoste, P.; Roux, A.; Reynaud-Gaubert, M.; Kessler, R.; Mussot, S.; et al. T Cells Promote Bronchial Epithelial Cell Secretion of Matrix Metalloproteinase-9 via a C-C Chemokine Receptor Type 2 Pathway: Implications for Chronic Lung Allograft Dysfunction. Am. J. Transplant. 2017, 17, 1502–1514. [Google Scholar] [CrossRef]
- Belloli, E.A.; Gu, T.; Wang, Y.; Vummidi, D.; Lyu, D.M.; Combs, M.P.; Chughtai, A.; Murray, S.; Galbán, C.J.; Lama, V.N. Radiographic Graft Surveillance in Lung Transplantation: Prognostic Role of Parametric Response Mapping. Am. J. Respir. Crit. Care Med. 2021, 204, 967–976. [Google Scholar] [CrossRef]
- Berastegui, C.; Gómez-Ollés, S.; Sánchez-Vidaurre, S.; Culebras, M.; Monforte, V.; López-Meseguer, M.; Bravo, C.; Ramon, M.-A.; Romero, L.; Sole, J.; et al. BALF cytokines in different phenotypes of chronic lung allograft dysfunction in lung transplant patients. Clin. Transplant. 2017, 31, e12898. [Google Scholar] [CrossRef] [PubMed]
- Ciaramelli, C.; Fumagalli, M.; Viglio, S.; Bardoni, A.M.; Piloni, D.; Meloni, F.; Iadarola, P.; Airoldi, C. 1H NMR To Evaluate the Metabolome of Bronchoalveolar Lavage Fluid (BALf) in Bronchiolitis Obliterans Syndrome (BOS): Toward the Development of a New Approach for Biomarker Identification. J. Proteome Res. 2017, 16, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Budding, K.; Van De Graaf, E.A.; Kardol-Hoefnagel, T.; Erp, J.M.K.-V.; Luijk, B.D.; Oudijk, E.-J.D.; Van Kessel, D.A.; Grutters, J.C.; Hack, C.E.; Otten, H.G. Soluble CD59 is a Novel Biomarker for the Prediction of Obstructive Chronic Lung Allograft Dysfunction After Lung Transplantation. Sci. Rep. 2016, 6, 26274. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yue, Z.; Yu, J.; Daguindau, E.; Kushekhar, K.; Zhang, Q.; Ogata, Y.; Gafken, P.R.; Inamoto, Y.; Gracon, A.; et al. Proteomic Characterization Reveals That MMP-3 Correlates with Bronchiolitis Obliterans Syndrome Following Allogeneic Hematopoietic Cell and Lung Transplantation. Am. J. Transplant. 2016, 16, 2342–2351. [Google Scholar] [CrossRef]
- Vandermeulen, E.; Verleden, S.E.; Bellon, H.; Ruttens, D.; Lammertyn, E.; Claes, S.; Vandooren, J.; Ugarte-Berzal, E.; Schols, D.; Emonds, M.-P.; et al. Humoral immunity in phenotypes of chronic lung allograft dysfunction: A broncho-alveolar lavage fluid analysis. Transpl. Immunol. 2016, 38, 27–32. [Google Scholar] [CrossRef]
- Ericson, P.A.; Mirgorodskaya, E.; Hammar, O.S.; Viklund, E.A.; Almstrand, A.-C.R.; Larsson, P.J.-W.; Riise, G.; Olin, A.-C. Low Levels of Exhaled Surfactant Protein A Associated with BOS After Lung Transplantation. Transplant. Direct. 2016, 2, e103. [Google Scholar] [CrossRef]
- Hayes, D.; Harhay, M.O.; Nicol, K.K.; Liyanage, N.P.M.; Keller, B.C.; Robinson, R.T. Lung T-Cell Profile Alterations are Associated with Bronchiolitis Obliterans Syndrome in Cystic Fibrosis Lung Transplant Recipients. Lung 2020, 198, 157–161. [Google Scholar] [CrossRef]
- Schreurs, I.; Meek, B.; Hijdra, D.; van Moorsel, C.; Luijk, H.; Erp, J.K.-V.; Oudijk, E.; van Kessel, D.; Grutters, J. Lung Transplantation Has a Strong Impact on the Distribution and Phenotype of Monocyte Subsets. Transplant. Proc. 2020, 52, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, I.; Meek, B.; Hijdra, D.; Rodriguez Gomez, M.; van Moorsel CH, M.; Luijk, H.D.; Kwakkel-van Erp, J.M.; Oudijk, E.J.; van Kessel, D.A.; Grutters, J.C. Lung Transplant Patients Show a Dissimilar Peripheral B-Cell Subset Ratio Compared With Healthy Controls. Exp. Clin. Transplant. 2020, 18, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Larsson-Callerfelt, A.-K.; Müller, C.; Andersson-Sjöland, A.; Thiman, L.; Larsson, H.; Hallgren, O.; Bjermer, L.; Eriksson, L.; Westergren-Thorsson, G. VEGF synthesis and VEGF receptor 2 expression in patients with bronchiolitis obliterans syndrome after lung transplantation. Respir. Med. 2020, 166, 105944. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Gunasekaran, M.; Ravichandran, R.; Fisher, C.E.; Limaye, A.P.; Hu, C.; McDyer, J.; Kaza, V.; Bharat, A.; Tokman, S.; et al. Circulating exosomes with lung self-antigens as a biomarker for chronic lung allograft dysfunction: A retrospective analysis. J. Heart Lung Transplant. 2020, 39, 1210–1219. [Google Scholar] [CrossRef]
- Hodge, G.; Hodge, S.; Liu, H.; Nguyen, P.; Holmes-Liew, C.-L.; Holmes, M. Bronchiolitis obliterans syndrome is associated with increased senescent lymphocytes in the small airways. J. Heart Lung Transplant. 2021, 40, 108–119. [Google Scholar] [CrossRef]
- Sacreas, A.; Yang, J.Y.C.; Vanaudenaerde, B.M.; Sigdel, T.K.; Liberto, J.M.; Damm, I.; Verleden, G.M.; Vos, R.; Verleden, S.E.; Sarwal, M.M. The common rejection module in chronic rejection post lung transplantation. PLoS ONE. 2018, 13, e0205107. [Google Scholar] [CrossRef]
- Levy, L.; Tigert, A.; Huszti, E.; Saito, T.; Mitsakakis, N.; Moshkelgosha, S.; Joe, B.; Boonstra, K.M.; Tikkanen, J.M.; Keshavjee, S.; et al. Epithelial cell death markers in bronchoalveolar lavage correlate with chronic lung allograft dysfunction subtypes and survival in lung transplant recipients—A single-center retrospective cohort study. Transpl. Int. 2019, 32, 965–973. [Google Scholar] [CrossRef]
- Brosseau, C.; Danger, R.; Durand, M.; Durand, E.; Foureau, A.; Lacoste, P.; Tissot, A.; Roux, A.; Reynaud-Gaubert, M.; Kessler, R.; et al. Blood CD9+ B cell, a biomarker of bronchiolitis obliterans syndrome after lung transplantation. Am. J. Transplant. 2019, 19, 3162–3175. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Li, H.-B. Recent progress on liver kinase B1 (LKB1): Expression, regulation, downstream signaling and cancer suppressive function. Int. J. Mol. Sci. 2014, 15, 16698–16718. [Google Scholar] [CrossRef]
- Saxena, M.; Balaji, S.A.; Deshpande, N.; Ranganathan, S.; Pillai, D.M.; Hindupur, S.K.; Rangarajan, A. AMP-activated protein kinase promotes epithelial-mesenchymal transition in cancer cells through Twist1 upregulation. J. Cell Sci. 2018, 131, jcs208314. [Google Scholar] [CrossRef]
- Bonanno, S.; Zulato, E.; Pavan, A.; Attili, I.; Pasello, G.; Conte, P.; Indraccolo, S. LKB1 and Tumor Metabolism: The Interplay of Immune and Angiogenic Microenvironment in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 1874. [Google Scholar] [CrossRef]
- Skoulidis, F.; Goldberg, M.E.; Greenawalt, D.M.; Hellmann, M.D.; Awad, M.M.; Gainor, J.F.; Schrock, A.B.; Hartmaier, R.J.; Trabucco, S.E.; Gay, L.; et al. STK11/LKB1 Mutations and PD-1 Inhibitor Resistance in KRAS-Mutant Lung Adenocarcinoma. Cancer Discov. 2018, 8, 822–835. [Google Scholar] [CrossRef]
- Skoulidis, F.; Byers, L.A.; Diao, L.; Papadimitrakopoulou, V.A.; Tong, P.; Izzo, J.; Behrens, C.; Kadara, H.; Parra, E.R.; Canales, J.R.; et al. Co-occurring genomic alterations define major subsets of KRAS-mutant lung adenocarcinoma with distinct biology, immune profiles, and therapeutic vulnerabilities. Cancer Discov. 2015, 5, 860–877. [Google Scholar] [CrossRef]
- Su, X.; Wang, Q.; Guo, W.; Pei, X.; Niu, Q.; Liu, M.; Liu, Y.; Chen, S.; Feng, S.; He, Y.; et al. Loss of Lkb1 impairs Treg function and stability to aggravate graft-versus-host disease after bone marrow transplantation. Cell Mol. Immunol. 2020, 17, 483–495. [Google Scholar] [CrossRef]
- Tiriveedhi, V.; Angaswamy, N.; Brand, D.; Weber, J.; Gelman, A.G.; Hachem, R.; Trulock, E.P.; Meyers, B.; Patterson, G.; Mohanakumar, T. A shift in the collagen V antigenic epitope leads to T helper phenotype switch and immune response to self-antigen leading to chronic lung allograft rejection. Clin. Exp. Immunol. 2012, 167, 158–168. [Google Scholar] [CrossRef]
- Shastri, N.; Yewdell, J.W. Editorial overview: Antigen processing and presentation: Where cellular immunity begins. Curr. Opin. Immunol. 2015, 34, v–vii. [Google Scholar] [CrossRef]
- Sacreas, A.; Taupin, J.-L.; Emonds, M.-P.; Daniëls, L.; Van Raemdonck, D.E.; Vos, R.; Verleden, G.M.; Vanaudenaerde, B.M.; Roux, A.; Verleden, S.E. Intragraft donor-specific anti-HLA antibodies in phenotypes of chronic lung allograft dysfunction. Eur. Respir. J. 2019, 54, 1900847. [Google Scholar] [CrossRef] [PubMed]
- Brunet, J.F.; Denizot, F.; Luciani, M.F.; Roux-Dosseto, M.; Suzan, M.; Mattei, M.G.; Golstein, P. A new member of the immunoglobulin superfamily--CTLA-4. Nature 1987, 328, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.H.; Pauken, K.E. The diverse functions of the PD1 inhibitory pathway. Nat. Rev. Immunol. 2018, 18, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhu, G.; Tamada, K.; Chen, L. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat. Med. 1999, 5, 1365–1369. [Google Scholar] [CrossRef]
- Xia, L.; Liu, Y.; Wang, Y. PD-1/PD-L1 Blockade Therapy in Advanced Non-Small-Cell Lung Cancer: Current Status and Future Directions. Oncologist 2019, 24, S31–S41. [Google Scholar] [CrossRef] [PubMed]
- Immune Checkpoint Inhibitors in Heart or Lung Transplantation: Early Results from a Registry Initiative—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/32265077/ (accessed on 11 April 2022).
- Shibuya, M. Vascular Endothelial Growth Factor (VEGF) and Its Receptor (VEGFR) Signaling in Angiogenesis: A Crucial Target for Anti- and Pro-Angiogenic Therapies. Genes Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Shijubo, N.; Kojima, H.; Nagata, M.; Ohchi, T.; Suzuki, A.; Abe, S.; Sato, N. Tumor angiogenesis of non-small cell lung cancer. Microsc. Res. Tech. 2003, 60, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed]
- Volm, M.; Koomägi, R.; Mattern, J.; Stammler, G. Angiogenic growth factors and their receptors in non-small cell lung carcinomas and their relationships to drug response in vitro. Anticancer Res. 1997, 17, 99–103. [Google Scholar]
- Volm, M.; Koomägi, R.; Mattern, J. Prognostic value of vascular endothelial growth factor and its receptor Flt-1 in squamous cell lung cancer. Int. J. Cancer 1997, 74, 64–68. [Google Scholar] [CrossRef]
- Vascular Endothelial Growth Factor in Human Lung Transplantation—CHEST. Available online: https://journal.chestnet.org/article/S0012-3692(15)37575-9/ppt (accessed on 11 April 2022).
- Kastelijn, E.A.; Rijkers, G.T.; Van Moorsel, C.H.; Zanen, P.; Erp, J.M.K.-V.; Van De Graaf, E.A.; Van Kessel, D.A.; Grutters, J.C.; Bosch, J.M.V.D. Systemic and exhaled cytokine and chemokine profiles are associated with the development of bronchiolitis obliterans syndrome. J. Heart Lung Transplant. 2010, 29, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, R.; Zhang, W.; Naseem, M.; Puccini, A.; Berger, M.D.; Soni, S.; McSkane, M.; Baba, H.; Lenz, H.-J. CXCL9, CXCL10, CXCL11/CXCR3 axis for immune activation—A target for novel cancer therapy. Cancer Treat. Rev. 2018, 63, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Khatri, P.; Roedder, S.; Kimura, N.; de Vusser, K.; Morgan, A.A.; Gong, Y.; Fischbein, M.P.; Robbins, R.C.; Naesens, M.; Butte, A.J.; et al. A Common Rejection Module (CRM) for Acute Rejection Across Multiple Organs Identifies Novel Therapeutics for Organ Transplantation. J. Exp. Med. 2013, 210, 2205–2221. Available online: https://rupress.org/jem/article/210/11/2205/41536/A-common-rejection-module-CRM-for-acute-rejection (accessed on 11 April 2022). [CrossRef]
- American Thoracic Society, European Respiratory Society. ATS/ERS recommendations for standardized procedures for the online and offline measurement of exhaled lower respiratory nitric oxide and nasal nitric oxide, 2005. Am. J. Respir. Crit. Care Med. 2005, 171, 912–930. [Google Scholar] [CrossRef]
- Tsoukias, N.M.; George, S.C. A two-compartment model of pulmonary nitric oxide exchange dynamics. J. Appl. Physiol. 1998, 85, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Veldhuizen, R.; Nag, K.; Orgeig, S.; Possmayer, F. The role of lipids in pulmonary surfactant. Biochim. Biophys. Acta 1998, 1408, 90–108. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.; Tornoe, I.; Nielsen, O.; Koch, C.; Steinhilber, W.; Holmskov, U. Expression and localization of lung surfactant protein A in human tissues. Am. J. Respir. Cell Mol. Biol. 2003, 29, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Barreira, E.R.; Precioso, A.R.; Bousso, A. Pulmonary surfactant in respiratory syncytial virus bronchiolitis: The role in pathogenesis and clinical implications. Pediatr. Pulmonol. 2011, 46, 415–420. [Google Scholar] [CrossRef]
- Meloni, F.; Salvini, R.; Bardoni, A.M.; Passadore, I.; Solari, N.; Vitulo, P.; Oggionni, T.; Viganò, M.; Pozzi, E.; Fietta, A.M. Bronchoalveolar lavage fluid proteome in bronchiolitis obliterans syndrome: Possible role for surfactant protein A in disease onset. J. Heart Lung Transplant. 2007, 26, 1135–1143. [Google Scholar] [CrossRef]
- Dekens, D.W.; Eisel, U.L.M.; Gouweleeuw, L.; Schoemaker, R.G.; De Deyn, P.P.; Naudé, P.J.W. Lipocalin 2 as a link between ageing, risk factor conditions and age-related brain diseases. Ageing Res. Rev. 2021, 70, 101414. [Google Scholar] [CrossRef]
- Redl, B.; Habeler, M. The diversity of lipocalin receptors. Biochimie 2022, 192, 22–29. [Google Scholar] [CrossRef]
- Lipocalin-2, Pro- or Anti-Apoptotic?—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/19160065/ (accessed on 11 April 2022).
- Treekitkarnmongkol, W.; Hassane, M.; Sinjab, A.; Chang, K.; Hara, K.; Rahal, Z.; Zhang, J.; Lu, W.; Sivakumar, S.; McDowell, T.L.; et al. Augmented Lipocalin-2 Is Associated with Chronic Obstructive Pulmonary Disease and Counteracts Lung Adenocarcinoma Development. Am. J. Respir. Crit. Care Med. 2021, 203, 90–101. [Google Scholar] [CrossRef]
- Vasta, G.R. Galectins in Host-Pathogen Interactions: Structural, Functional and Evolutionary Aspects. Adv. Exp. Med. Biol. 2020, 1204, 169–196. [Google Scholar]
- Ho, J.E.; Gao, W.; Levy, D.; Santhanakrishnan, R.; Araki, T.; Rosas, I.O.; Hatabu, H.; Latourelle, J.C.; Nishino, M.; Dupuis, J.; et al. Galectin-3 Is Associated with Restrictive Lung Disease and Interstitial Lung Abnormalities. Am. J. Respir. Crit. Care Med. 2016, 194, 77–83. [Google Scholar] [CrossRef]
- Shevchenko, O.; Tsirulnikova, O.; Sharapchenko, S.; Pashkov, I.; Bekov, M.; Shigaev, E.; Gichkun, O.; Velikiy, D.; Gautier, S. MiR-339 and galectin-3, diagnostic value in patients with airway obstruction after lung transplantation. Transpl. Int. 2021, 34, 1733–1739. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.A.; Harris, C.L.; Williams, A.S.; Mizuno, M.; Gallagher, S.; Smith, R.A.G.; Morgan, B.P. Generation of a recombinant, membrane-targeted form of the complement regulator CD59, activity in vitro and in vivo. J. Biol. Chem. 2003, 278, 48921–48927. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.M.; AAl-Binni, M.; Bani Hani, A.; Abu Abeeleh, M.; Abu-Humaidan, A.H.A. Complement Terminal Pathway Activation is Associated with Organ Failure in Sepsis Patients. J. Inflamm. Res. 2022, 15, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Craig, V.J.; Zhang, L.; Hagood, J.S.; Owen, C.A. Matrix metalloproteinases as therapeutic targets for idiopathic pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2015, 53, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, C.M.; Radisky, D.C.; Aschner, Y.; Downey, G.P. The importance of matrix metalloproteinase-3 in respiratory disorders. Expert Rev. Respir. Med. 2014, 8, 411–421. [Google Scholar] [CrossRef]
- Inamoto, Y.; Martin, P.J.; Onstad, L.E.; Cheng, G.S.; Williams, K.M.; Pusic, I.; Ho, V.T.; Arora, M.; Pidala, J.; Flowers, M.E.D.; et al. Relevance of Plasma Matrix Metalloproteinase-9 for Bronchiolitis Obliterans Syndrome after Allogeneic Hematopoietic Cell Transplantation. Transplant. Cell. Ther. 2021, 27, 759. [Google Scholar] [CrossRef]
- Nagase, H.; Woessner, J.F. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar] [CrossRef]
- Hendrix, A.Y.; Kheradmand, F. The Role of Matrix Metalloproteinases in Development, Repair, and Destruction of the Lungs. Prog. Mol. Biol. Transl. Sci. 2017, 148, 1–29. [Google Scholar]
- Kennedy, V.E.; Todd, J.L.; Palmer, S.M. Bronchoalveolar Lavage as a Tool to Predict, Diagnose and Understand Bronchiolitis Obliterans Syndrome. Am. J. Transplant. 2013, 13, 552–561. [Google Scholar] [CrossRef]
- McWilliams, T.; Zheng, L.; Orsida, B.; Levvey, B.; Walters, E.; Whitford, H.; Snell, G.; Williams, T. The profibrotic cytokine transforming growth factor beta (TGFβ) is elevated early in the development of bronchiolitis obliterans syndrome (BOS) in lung transplant recipients (LTR). J. Heart Lung Transplant. 2004, 23, S126. [Google Scholar] [CrossRef]
- Hachem, R.R.; Tiriveedhi, V.; Patterson, G.A.; Aloush, A.; Trulock, E.P.; Mohanakumar, T. Antibodies to K-α 1 tubulin and collagen V are associated with chronic rejection after lung transplantation. Am. J. Transplant. 2012, 12, 2164–2171. [Google Scholar] [CrossRef] [PubMed]
- Sumpter, T.L.; Wilkes, D.S. Role of autoimmunity in organ allograft rejection: A focus on immunity to type V collagen in the pathogenesis of lung transplant rejection. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 286, L1129–L1139. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Saini, D.; Weber, J.; Ramachandran, S.; Phelan, D.; Tiriveedhi, V.; Liu, M.; Steward, N.; Aloush, A.; Hachem, R.; Trulock, E.; et al. Alloimmunity-induced autoimmunity as a potential mechanism in the pathogenesis of chronic rejection of human lung allografts. J. Heart Lung Transplant. 2011, 30, 624–631. [Google Scholar] [CrossRef]
- Rao, U.; Sharma, M.; Mohanakumar, T.; Ahn, C.; Gao, A.; Kaza, V. Prevalence of antibodies to lung self-antigens (Kα1 tubulin and collagen V) and donor specific antibodies to HLA in lung transplant recipients and implications for lung transplant outcomes: Single center experience. Transpl. Immunol. 2019, 54, 65–72. [Google Scholar] [CrossRef]
- Tiriveedhi, V.; Gelman, A.E.; Mohanakumar, T. HIF-1α signaling by airway epithelial cell K-α1-tubulin: Role in fibrosis and chronic rejection of human lung allografts. Cell Immunol. 2012, 273, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Saenz, S.A.; Taylor, B.C.; Artis, D. Welcome to the neighborhood: Epithelial cell-derived cytokines license innate and adaptive immune responses at mucosal sites. Immunol. Rev. 2008, 226, 172–190. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, L.A.; Parker, S.M.; A Brougham, K.; Johnson, G.E.; Gorowiec, M.R.; Ward, C.; Lordan, J.L.; Corris, P.A.; Kirby, J.A.; Fisher, A.J. Epithelial to mesenchymal transition (EMT) and airway remodelling after human lung transplantation. Thorax 2009, 64, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Waddell, T.K.; Wagnetz, U.; Roberts, H.C.; Hwang, D.M.; Haroon, A.; Wagnetz, D.; Chaparro, C.; Singer, L.; Hutcheon, M.A.; et al. Restrictive allograft syndrome (RAS): A novel form of chronic lung allograft dysfunction. J. Heart Lung Transplant. 2011, 30, 735–742. [Google Scholar] [CrossRef]
- Verleden, G.M.; Raghu, G.; Meyer, K.C.; Glanville, A.R.; Corris, P. A new classification system for chronic lung allograft dysfunction. J. Heart Lung Transplant. 2014, 33, 127–133. [Google Scholar] [CrossRef]
- Colvin, R.B.; Smith, R.N. Antibody-mediated organ-allograft rejection. Nat. Rev. Immunol. 2005, 5, 807–817. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Song, W.-C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.D.; Wang, Z.; Chen, D.-F.; Reinsmoen, N.L.; Finlen-Copeland, C.A.; Davis, W.A.; Zaas, D.W.; Palmer, S.M. Implications for human leukocyte antigen antibodies after lung transplantation: A 10-year experience in 441 patients. Chest 2013, 144, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.; Le Lan, I.B.; Holifanjaniaina, S.; Thomas, K.A.; Hamid, A.M.; Picard, C.; Grenet, D.; De Miranda, S.; Douvry, B.; Beaumont-Azuar, L.; et al. Antibody-Mediated Rejection in Lung Transplantation: Clinical Outcomes and Donor-Specific Antibody Characteristics. Am. J. Transplant. 2016, 16, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
- Gregson, A.L.; Hoji, A.; Saggar, R.; Ross, D.J.; Kubak, B.M.; Jamieson, B.D.; Weigt, S.S.; Lynch, J.P.; Ardehali, A.; Belperio, J.A.; et al. Bronchoalveolar immunologic profile of acute human lung transplant allograft rejection. Transplantation 2008, 85, 1056–1059. [Google Scholar] [CrossRef]
- Zheng, L.; Orsida, B.; Whitford, H.; Levvey, B.; Ward, C.; Walters, E.; Williams, T.J.; Snell, G.I. Longitudinal comparisons of lymphocytes and subtypes between airway wall and bronchoalveolar lavage after human lung transplantation. Transplantation 2005, 80, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Vandermeulen, E.; Lammertyn, E.; Verleden, S.E.; Ruttens, D.; Bellon, H.; Ricciardi, M.; Somers, J.; Bracke, K.R.; Eynde, K.V.D.; Tousseyn, T.; et al. Immunological diversity in phenotypes of chronic lung allograft dysfunction: A comprehensive immunohistochemical analysis. Transpl. Int. 2017, 30, 134–143. [Google Scholar] [CrossRef]
- Bharat, A.; Narayanan, K.; Street, T.; Fields, R.C.; Steward, N.; Aloush, A.; Meyers, B.; Schuessler, R.; Trulock, E.P.; Patterson, G.A.; et al. Early posttransplant inflammation promotes the development of alloimmunity and chronic human lung allograft rejection. Transplantation 2007, 83, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Belperio, J.A.; Weigt, S.S.; Fishbein, M.C.; Lynch, J.P. Chronic lung allograft rejection: Mechanisms and therapy. Proc. Am. Thorac. Soc. 2009, 6, 108–121. [Google Scholar] [CrossRef]
- Sigdel, T.K.; Bestard, O.; Tran, T.Q.; Hsieh, S.-C.; Roedder, S.; Damm, I.; Vincenti, F.; Sarwal, M.M. A Computational Gene Expression Score for Predicting Immune Injury in Renal Allografts. PLoS ONE 2015, 10, e0138133. [Google Scholar] [CrossRef]
- Hong, J.-H.; Lee, W.-C.; Hsu, Y.-M.; Liang, H.-J.; Wan, C.-H.; Chien, C.-L.; Lin, C.-Y. Characterization of the biochemical effects of naphthalene on the mouse respiratory system using NMR-based metabolomics. J. Appl. Toxicol. 2014, 34, 1379–1388. [Google Scholar] [CrossRef]
- Fabiano, A.; Gazzolo, D.; Zimmermann, L.J.; Gavilanes, A.W.; Paolillo, P.; Fanos, V.; Caboni, P.; Barberini, L.; Noto, A.; Atzori, L. Metabolomic analysis of bronchoalveolar lavage fluid in preterm infants complicated by respiratory distress syndrome: Preliminary results. J. Matern. Fetal. Neonatal Med. 2011, 24 (Suppl. 2), 55–58. [Google Scholar] [CrossRef]
- Wolak, J.E.; Esther, C.R.; O’Connell, T.M. Metabolomic analysis of bronchoalveolar lavage fluid from cystic fibrosis patients. Biomarkers 2009, 14, 55–60. [Google Scholar] [CrossRef]
- Wen, C.; Li, F.; Zhang, L.; Duan, Y.; Guo, Q.; Wang, W.; He, S.; Li, J.; Yin, Y. Taurine is Involved in Energy Metabolism in Muscles, Adipose Tissue, and the Liver. Mol. Nutr. Food Res. 2019, 63, e1800536. [Google Scholar] [CrossRef]
- Chen, J.; Xue, X.; Cai, J.; Jia, L.; Sun, B.; Zhao, W. Protective effect of taurine on sepsis-induced lung injury via inhibiting the p38/MAPK signaling pathway. Mol. Med. Rep. 2021, 24, 653. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, J.; Wei, B.K.; Dong, G.; Wang, X. Protective Effect of Taurine on Paraquat-Induced Lung Epithelial Cell Injury. Adv. Exp. Med. Biol. 2019, 1155, 739–746. [Google Scholar] [PubMed]
- Di Carlo, S.; Rossi, E.; Politano, G.; Inghilleri, S.; Morbini, P.; Calabrese, F.; Benso, A.; Savino, A.; Cova, E.; Zampieri, D.; et al. Identification of miRNAs Potentially Involved in Bronchiolitis Obliterans Syndrome: A Computational Study. PLoS ONE 2016, 11, e0161771. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ramachandran, S.; Gunasekaran, M.; Zhou, F.; Trulock, E.; Kreisel, D.; Hachem, R.; Mohanakumar, T. MicroRNA-144 dysregulates the transforming growth factor-β signaling cascade and contributes to the development of bronchiolitis obliterans syndrome after human lung transplantation. J. Heart Lung Transplant. 2015, 34, 1154–1162. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bozzini, S.; Del Fante, C.; Morosini, M.; Berezhinskiy, H.O.; Auner, S.; Cattaneo, E.; Della Zoppa, M.; Pandolfi, L.; Cacciatore, R.; Perotti, C.; et al. Mechanisms of Action of Extracorporeal Photopheresis in the Control of Bronchiolitis Obliterans Syndrome (BOS): Involvement of Circulating miRNAs. Cells 2022, 11, 1117. [Google Scholar] [CrossRef]
- Mori, M.A.; Ludwig, R.G.; Garcia-Martin, R.; Brandão, B.B.; Kahn, C.R. Extracellular miRNAs: From Biomarkers to Mediators of Physiology and Disease. Cell Metab. 2019, 30, 656–673. [Google Scholar] [CrossRef]
- Yang, D.; Wang, J.-J.; Li, J.-S.; Xu, Q.-Y. miR-103 Functions as a Tumor Suppressor by Directly Targeting Programmed Cell Death 10 in NSCLC. Oncol. Res. 2018, 26, 519–528. [Google Scholar] [CrossRef]
- Bozzini, S.; Pandolfi, L.; Rossi, E.; Inghilleri, S.; Zorzetto, M.; Ferrario, G.; Di Carlo, S.; Politano, G.; De Silvestri, A.; Frangipane, V.; et al. miRNAs Potentially Involved in Post Lung Transplant-Obliterative Bronchiolitis: The Role of miR-21-5p. Cells 2021, 10, 688. [Google Scholar] [CrossRef] [PubMed]
- Melissa, L.T.; Jeremy, D.F.; Timothy, M.C. Pharmacotherapy of Lung Transplantation: An Overview. 2013. Available online: https://journals.sagepub.com/doi/abs/10.1177/0897190012466048 (accessed on 11 April 2022).
- Traitanon, O.; Mathew, J.M.; La Monica, G.; Xu, L.; Mas, V.; Gallon, L. Differential Effects of Tacrolimus versus Sirolimus on the Proliferation, Activation and Differentiation of Human B Cells. PLoS ONE 2015, 10, e0129658. [Google Scholar] [CrossRef] [PubMed]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Eibel, H.; Kraus, H.; Sic, H.; Kienzler, A.-K.; Rizzi, M. B cell Biology: An Overview. Curr. Allergy Asthma Rep. 2014, 14, 434. [Google Scholar] [CrossRef] [PubMed]
- Stegall, M.D.; Raghavaiah, S.; Gloor, J.M. The (re)emergence of B cells in organ transplantation. Curr. Opin. Organ Transplant. 2010, 15, 451–455. [Google Scholar] [CrossRef]
- Girmanova, E.; Hruba, P.; Viklicky, O. Circulating biomarkers of tolerance. Transplant. Rev. 2015, 29, 68–72. [Google Scholar] [CrossRef]
- Lung Transplant Patients Show a Dissimilar Peripheral B—Cell Subset Ratio Compared with Healthy Controls—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/32279657/ (accessed on 11 April 2022).
- Gauthier, J.M.; Li, W.; Hsiao, H.-M.; Takahashi, T.; Arefanian, S.; Krupnick, A.S.; Gelman, A.E.; Kreisel, D. Mechanisms of Graft Rejection and Immune Regulation after Lung Transplant. Ann. Am. Thorac. Soc. 2017, 14, S216–S219. [Google Scholar] [CrossRef]
- B Cell Development and Maturation | SpringerLink. Available online: https://link.springer.com/chapter/10.1007/978-981-15-3532-1_1 (accessed on 11 April 2022).
- Liu, Y.; Duan, W.-R.; Liu, S.; Liu, T.; Chang, Y.-J.; Fan, X.-M. Correlation of CD19+CD24hiCD38hi B cells in coronary artery disease with severity of atherosclerosis. Chin. Med. J. 2020, 133, 1257–1258. [Google Scholar] [CrossRef]
- Brosseau, C.; Colas, L.; Magnan, A.; Brouard, S. CD9 Tetraspanin: A New Pathway for the Regulation of Inflammation? Front. Immunol. 2018, 9, 2316. [Google Scholar] [CrossRef]
- Sun, J.; Wang, J.; Pefanis, E.; Chao, J.; Rothschild, G.; Tachibana, I.; Chen, J.K.; Ivanov, I.I.; Rabadan, R.; Takeda, Y.; et al. Transcriptomics Identify CD9 as a Marker of Murine IL-10-Competent Regulatory B Cells. Cell Rep. 2015, 13, 1110–1117. [Google Scholar] [CrossRef]
- Read, K.A.; Powell, M.D.; Sreekumar, B.K.; Oestreich, K.J. In Vitro Differentiation of Effector CD4+ T Helper Cell Subsets. Methods Mol. Biol. 2019, 1960, 75–84. [Google Scholar] [PubMed]
- Kurachi, M. CD8+ T cell exhaustion. Semin Immunopathol. 2019, 41, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Li, P.; Du, X.; Liu, Y.; Zhang, B.; Qi, F. Regulatory T cells induce transplant immune tolerance. Transpl. Immunol. 2021, 67, 101411. [Google Scholar] [CrossRef] [PubMed]
- Zou, W. Regulatory T cells, tumour immunity and immunotherapy. Nat. Rev. Immunol. 2006, 6, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.R. The Balance of Th17 versus Treg Cells in Autoimmunity. Int. J. Mol. Sci. 2018, 19, 730. [Google Scholar] [CrossRef]
- Hodge, G.; Hodge, S.; Li-Liew, C.; Chambers, D.; Hopkins, P.; Reynolds, P.N.; Holmes, M. Time post-lung transplant correlates with increasing peripheral blood T cell granzyme B and proinflammatory cytokines. Clin. Exp. Immunol. 2010, 161, 584–590. [Google Scholar] [CrossRef]
- Hodge, G.; Mukaro, V.; Reynolds, P.N.; Hodge, S. Role of increased CD8/CD28(null) T cells and alternative co-stimulatory molecules in chronic obstructive pulmonary disease. Clin. Exp. Immunol. 2011, 166, 94–102. [Google Scholar] [CrossRef]
- Bosch, T.P.P.V.D.; Caliskan, K.; Kraaij, M.D.; Constantinescu, A.A.; Manintveld, O.C.; Leenen, P.J.M.; von der Thüsen, J.H.; Groningen, M.C.C.-V.; Baan, C.C.; Rowshani, A.T. CD16+ Monocytes and Skewed Macrophage Polarization toward M2 Type Hallmark Heart Transplant Acute Cellular Rejection. Front. Immunol. 2017, 8, 346. Available online: https://www.frontiersin.org/article/10.3389/fimmu.2017.00346 (accessed on 11 April 2022).
- The Three Human Monocyte Subsets: Implications for Health and Disease | SpringerLink. Available online: https://link.springer.com/article/10.1007/s12026-012-8297-3 (accessed on 11 April 2022).
- 6-Sulfo LacNAc (Slan) as a Marker for Non-classical Monocytes—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/31572354/ (accessed on 11 April 2022).
- Chiu, S.; Bharat, A. Role of monocytes and macrophages in regulating immune response following lung transplantation. Curr. Opin. Organ Transplant. 2016, 21, 239–245. [Google Scholar] [CrossRef]
- Luo, Y.; Fujii, H.; Gerster, T.; Roeder, R.G. A novel B cell-derived coactivator potentiates the activation of immunoglobulin promoters by octamer-binding transcription factors. Cell 1992, 71, 231–241. [Google Scholar] [CrossRef]
- Dymecki, S.M.; Niederhuber, J.E.; Desiderio, S.V. Specific expression of a tyrosine kinase gene, blk, in B lymphoid cells. Science 1990, 247, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Laine, J.; Künstle, G.; Obata, T.; Sha, M.; Noguchi, M. The protooncogene TCL1 is an Akt kinase coactivator. Mol. Cell 2000, 6, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Brinas, F.; Danger, R.; Brouard, S. TCL1A, B Cell Regulation and Tolerance in Renal Transplantation. Cells 2021, 10, 1367. [Google Scholar] [CrossRef] [PubMed]
- White, E.S. Lung extracellular matrix and fibroblast function. Ann. Am. Thorac. Soc. 2015, 12 (Suppl. 1), S30–S33. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Luan, F.; Zhao, Y.; Hao, H.; Zhou, Y.; Han, W.; Fu, X. Epithelial-mesenchymal transition: An emerging target in tissue fibrosis. Exp. Biol. Med. 2016, 241, 1–13. [Google Scholar] [CrossRef]
- Downer, N.J.; Ali, N.J.; Au-Yong, I.T.H. Investigating pleural thickening. BMJ 2013, 346, e8376. [Google Scholar] [CrossRef] [PubMed]
- Morshid, A.; Moshksar, A.; Das, A.; Duarte, A.G.; Palacio, D.; Villanueva-Meyer, J. HRCT Diagnosis of Pleuroparenchymal fibroelastosis: Report of two cases. Radiol. Case Rep. 2021, 16, 1564–1569. [Google Scholar] [CrossRef]
- Hakim, A.; Cooke, K.R.; Pavletic, S.Z.; Khalid, M.; Williams, K.M.; Hashmi, S.K. Diagnosis and treatment of bronchiolitis obliterans syndrome accessible universally. Bone Marrow Transplant. 2019, 54, 383–392. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
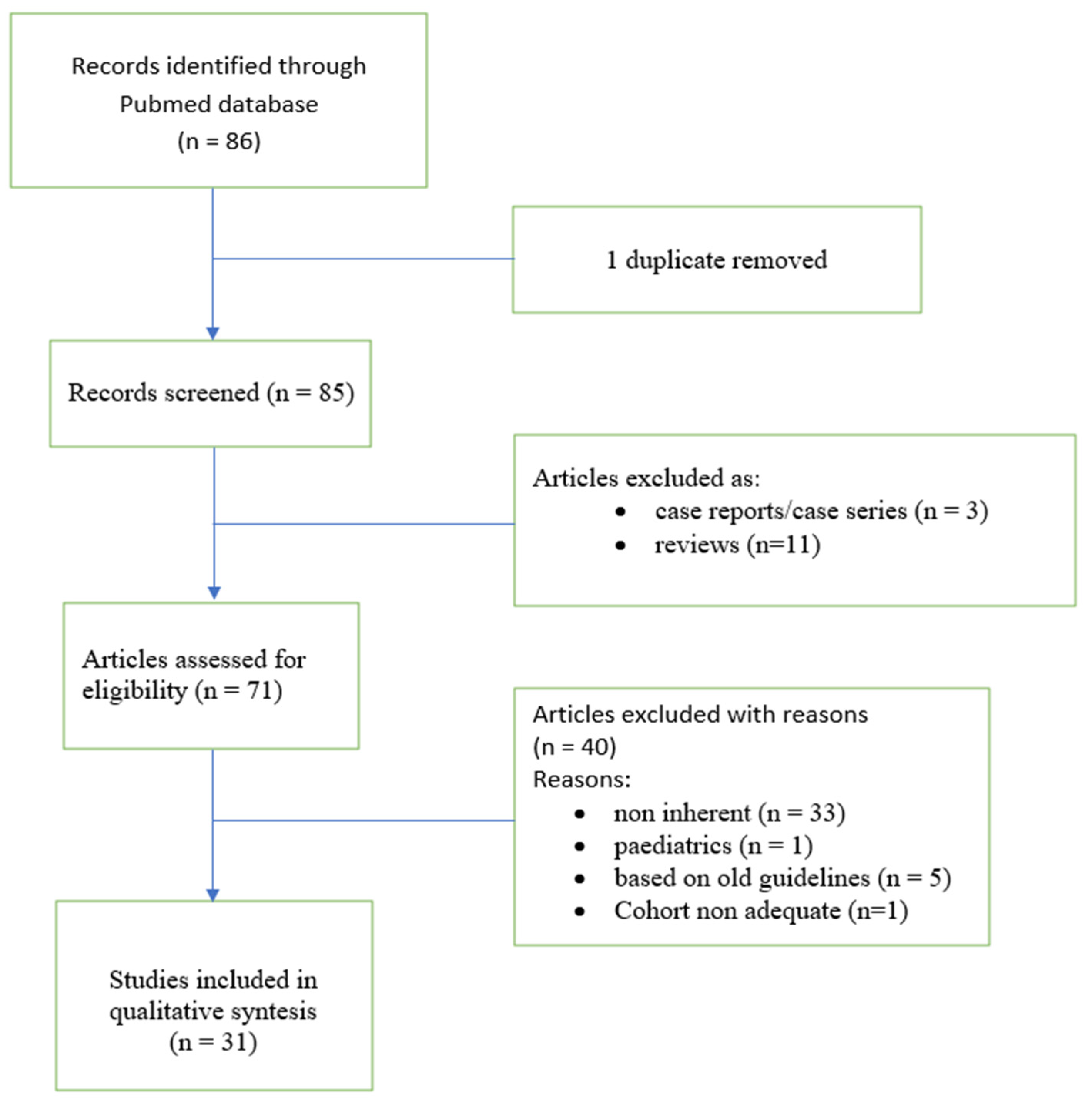
| Author (Year) | Study Type | Sample Size | Principal Results | Markers | Matrices |
|---|---|---|---|---|---|
| Rahman M, et al. Am J Transplant. 2021 [38] | Retrospective | 20 BOS and 20 stable | Exosomes released from lung tissue of BOS are associated with inactivated LKB1 that induces EMT | LKB1 gene | Tissue |
| Weigt SS, et al. Transplantation. 2021 [39] | Prospective | 15 BOS, 13 RAS, 4 mixed CLAD, 15 undefined CLAD phenotypes. | BOS had more air-trapping, lower quantitative image analysis (QIA) score than RAS | QIA score at HRCT | Imaging |
| Bansal S, et al. J Heart Lung Transplant 2022 [40] | Retrospective | 18 BOS, 13 RAS, 5 stable | Increased levels of circulating exosomes containing HLA-DR and HLA-DQ, NFkB, CIITA, 20S proteasome and PIGR can differentiate between the RAS and BOS | circulating exosomes | Plasma |
| Davidsen JR et al. J Clin Med 2021 [41] | Prospective | 19 BOS, 6 RAS | Pleural thickening more pronounced in RAS than BOS | Pleural thickening | Imaging |
| Veraar C, et al. Sci Rep. 2021 [42] | Prospective | 11 RAS, 30 BOS56 StableLT | RAS had higher Lipocalin-2 compared to BOS and Stable-LTx | Lipocalin-2 | Serum |
| Bergantini L, et al. Cells 2021 [43] | Retrospective | 6 BOS, 6 acute rejection, 4 stable | Decreased Treg cell BOS than acute and stable. Increase in Th1 and a decrease in Th17 in BAL of BOS patients than RAS and stable | Regulatory and effector cells | Peripheral blood and BAL |
| Itabashi Y, et al. Transplantation 2021 [44] | Retrospective | 18 BOS, 34 stable | BOSs have lower CCSP levels up to 9 months before BOS diagnosis | CCSP | BAL fluid |
| d’Alessandro M et al. Lung 2021 [45] | Retrospective | 10 BOS, 9 stable, 9 HC | Galectin-1 was higher in BOS than in stable LTx patients | Galectin1-3 | Serum |
| Righi I et al. Front.Immunol 2021 [46] | Retrospective | 4 RAS, 4 BOS, 6 HC | Higher T lymphocytes expressing PD-1, PD-L1 and CTLA4 immune checkpoint. | PD-1, CTLA-4, CD4, CD8, TIGIT. | Tissue |
| Cameli P et al. Respir Med 2015 [47] | Retrospective | 30 HCs,15 stable, 12 BOS | BOS patients had higher FeNO at 150 and 350 mL/s and CalvNO than non-BOS patients. | FeNO, CalvNO eCO | exhaled |
| Durand M et al. J Heart Lung Transplant 2018 [48] | Prospective | 50 stable, 32 BOS | An increase in Treg T-cells in BOS patients at 1 and 6 months after transplantation. | Treg-cells | Peripgeral blood |
| Danger R, et al. Front Immunol, 2018 [49] | Clinical trial | 62 stable, 43 prediction group, 32 BOS | POU, POU2AF1, TCL1A and BCLK were validated as predictive biomarkers of BOS more than 6 months before diagnosis | POU, POU2AF1, TCL1A and BCLK gene | Peripheral blood |
| Piloni D et al. BMC Pulm Med 2017 [50] | Retrospective | 137 LTx | T-reg cell counts progressively decreased according to the severity of CLAD | T reg cells. | Peripheral blood |
| Budding K, et al. TRanspl. Immun. 2017 [51] | Clinical trial | 10 BOS, 10 without BOS | MiR-21, miR-29a, miR-103 and miR-191 levels were significantly higher in BOS patients prior to clinical BOS diagnosis | miRnas | Serum |
| Pain M, et al. Am J Transplant 2017 [52] | Clinical trial | 49 stable, 29 BOS, 16 RAS | Plasma MMP-9 was associated with BOS and predicted the occurrence of CLAD, 12 months before the functional diagnosis | Matrix metalloproteins-9 | Culture supernatants and plasma |
| Belloli EA, et al. Am J Respir Crit Care Med 2017 [53] | Clinical trial | 52 LTx with spitometric decline | FEV1 decline has significantly higher PRMfSAD than their control | PRMfSAD | Imaging |
| Berastegui C. et al. Clin Transplant 2017 [54] | Cross-selectional study | 15 BOS, 7 RAS, 29 stable | BALF neutrophilia were higher in BOS than in stable. IL-5 presented significant differences between BOS and RAS | IL-5 | BAL Fluid |
| Ciaramelli C. et al. J. Proteom Res, 2017 [55] | Retrospective study and pilot study | 10 stable, 10 BOS 0p, 10 BOS | Suitability of the NMR approach in monitoring different pathological lung conditions | Metabolites | Imaging |
| Budding K, et al. Sci Rep. 2016 [56] | Case-control | 20 BOS, 69 non-BOS, 20 HC | BOS patients showed higher sCD59 diagnosis compared to non-BOS patients | sCD59 | Serum |
| Liu X, et al. Am J Transplant. 2017 [57] | Retrospective | 12 BOS, 16 infection, 15 CGVD without pulmonary involvement | Fibronectin 1 (FN1) and MMP-3 were associated with BOS development | FN1 and MMP-3 | Plasma |
| Vandermeulen E, et al. Transpl. Immunol. 2016 [58] | Retrospective | 15 BOS, 16 RAS, 14 stable | Increased levels of immunoglobulins and complement proteins are dominantly present in CLAD | Immunoglobulins C4d and C1q, MMPs | BAL Fluid, Tissue Peripheral blood |
| Ericson PA, et al. Transplant Direct. 2016 [59] | Case-control | 26 stable, 7 BOS, 33 HC | SP-A in exhaled particles and the SP-A/albumin ratio were lower in the BOS group compared to the BOS-free group | Surfactant protein A and the SP-A/albumin ratio | Exhaled |
| Hayes Jr D, et al. Lung. 2020 [60] | Case-control | 6 non-BOS, 10 BOS | A decline in CD4+ T-cell and increase in CD8+ T-cell in BOS when comparing baseline values and at 6 months follow-up | T-cell subsets | BAL Fluid |
| Schreurs I, et al. Transplant Proc. 2020 [61] | Case-control | 27 LTx, 17 BOS, 10 HC, 9 sarcoidosis patients | The absolute count of monocytes was decreased in BOS. The expression of both CD36 and CD163 was significantly increased in the LTx and BOS cohort | CD36 and CD163 | Peripheral blood |
| Schreurs I, et al. Exp Clin Transplant. 2020 [62] | Clinial trial | 17 BOS, 10 HC | Transitional and naïve B cells were decreased in BOS. | B-Cell subsets | Peripheral blood |
| Larsson-Callerfelt AK, et al. Respir Med 2020 [63] | Case-control | 14 LTx, 11 BOS | VEGF were lower 3 months after lung transplantation compared to non-transplanted subjects | VEGF, VEGF 2 receptor | Tissue |
| Sharma M, et al. J Heart Lung Transplant 2020 [64] | Retrospective | 21 with BOS, 10 stable | Circulating exosomes isolated from BOS demonstrated increased levels of lung SAgs (Kα1T and Col-V) 12 months prior to the diagnosis | Kα1T and Col-V | Plasma |
| Hodge G, et al. J Heart Lung Transplant. 2021 [65] | Case-control study | 10 BOS, 11 stable and 10 HC | Increased CD28null T and NKT-like cells were identified in BOS compared with that in the controls and stable transplant recipients | Granzyme B, IFN-γ, TNF-α, CD28 | Peripheral blood, Bal Fluid |
| Sacreas A, et al. PLoS One. 2018 [66] | Retrospective | 13 stable, 8 AR, 9 BOS, 10 RAS. | Transcriptional tissue analysis for CLAD distinguishes RAS from BOS. | CD6, TAP1, CXCL10, CXCL9, | BAL, Tissue |
| Levy L, et al. Transpl Int. 2019 [67] | Retrospective | 10 RAS, 16 BOS and 19 long-term CLAD-free controls. | M65 levels were significantly lower in BOS compared to RAS. Detection of BAL M65 may be used to differentiate CLAD subtypes | M65, fragments of cytokeratin-18 | BAL |
| Brosseau C, et al. Am J Transplant. 2019 [68] | Retrospective | 18 stable, 16 BOS | CD24hi CD38hi transitional B cells were increased in stable patients. These CD24hi CD38hi transitional B cells displayed significantly higher incidence of BOS | B cell profile | Peripheral blood |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavallaro, D.; Guerrieri, M.; Cattelan, S.; Fabbri, G.; Croce, S.; Armati, M.; Bennett, D.; Fossi, A.; Voltolini, L.; Luzzi, L.; et al. Markers of Bronchiolitis Obliterans Syndrome after Lung Transplant: Between Old Knowledge and Future Perspective. Biomedicines 2022, 10, 3277. https://doi.org/10.3390/biomedicines10123277
Cavallaro D, Guerrieri M, Cattelan S, Fabbri G, Croce S, Armati M, Bennett D, Fossi A, Voltolini L, Luzzi L, et al. Markers of Bronchiolitis Obliterans Syndrome after Lung Transplant: Between Old Knowledge and Future Perspective. Biomedicines. 2022; 10(12):3277. https://doi.org/10.3390/biomedicines10123277
Chicago/Turabian StyleCavallaro, Dalila, Marco Guerrieri, Stefano Cattelan, Gaia Fabbri, Sara Croce, Martina Armati, David Bennett, Antonella Fossi, Luca Voltolini, Luca Luzzi, and et al. 2022. "Markers of Bronchiolitis Obliterans Syndrome after Lung Transplant: Between Old Knowledge and Future Perspective" Biomedicines 10, no. 12: 3277. https://doi.org/10.3390/biomedicines10123277
APA StyleCavallaro, D., Guerrieri, M., Cattelan, S., Fabbri, G., Croce, S., Armati, M., Bennett, D., Fossi, A., Voltolini, L., Luzzi, L., Salvicchi, A., Paladini, P., Peris, A., d’Alessandro, M., Cameli, P., Bargagli, E., Tuscany Transplant Group, & Bergantini, L. (2022). Markers of Bronchiolitis Obliterans Syndrome after Lung Transplant: Between Old Knowledge and Future Perspective. Biomedicines, 10(12), 3277. https://doi.org/10.3390/biomedicines10123277