
The Precise Breakpoint Mapping in Paracentric Inversion 10q22.2q23.3 by Comprehensive Cytogenomic Analysis, Multicolor Banding, and Single-Copy Chromosome Sequencing
 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. The Clinical Case
2.2. Chromosome Preparation and GTG-Banding Analysis
2.3. Creation of Microdissected DNA Probes
2.4. Multicolor Chromosome Banding (MCB)
2.5. aCGH
2.6. Sequencing of the Microdissected DNA Library Generated from the Single-Copy of Inv(10)(q?22.3q?23.3) Chromosome
2.7. Ethics
3. Results
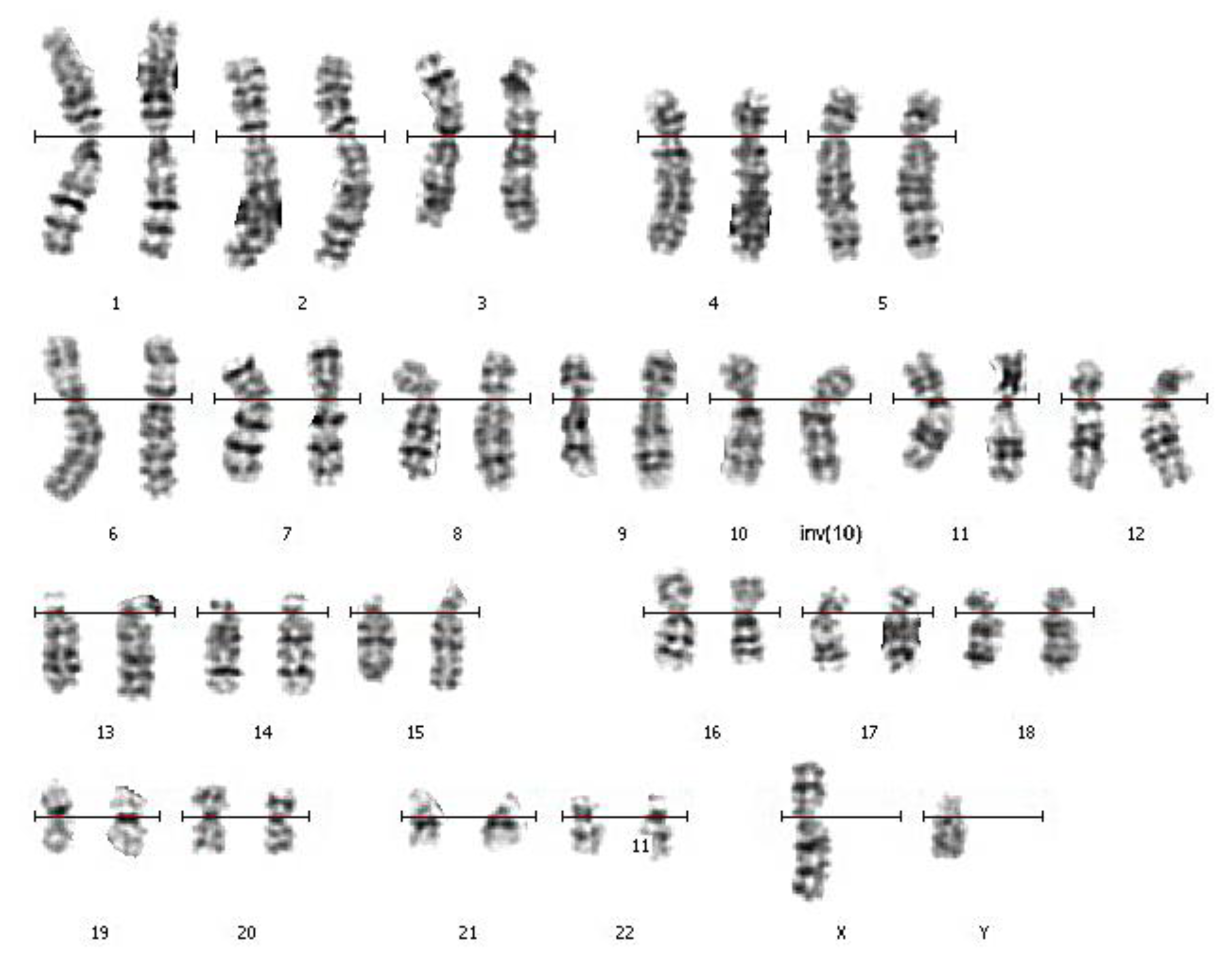
3.1. Routine Karyotyping
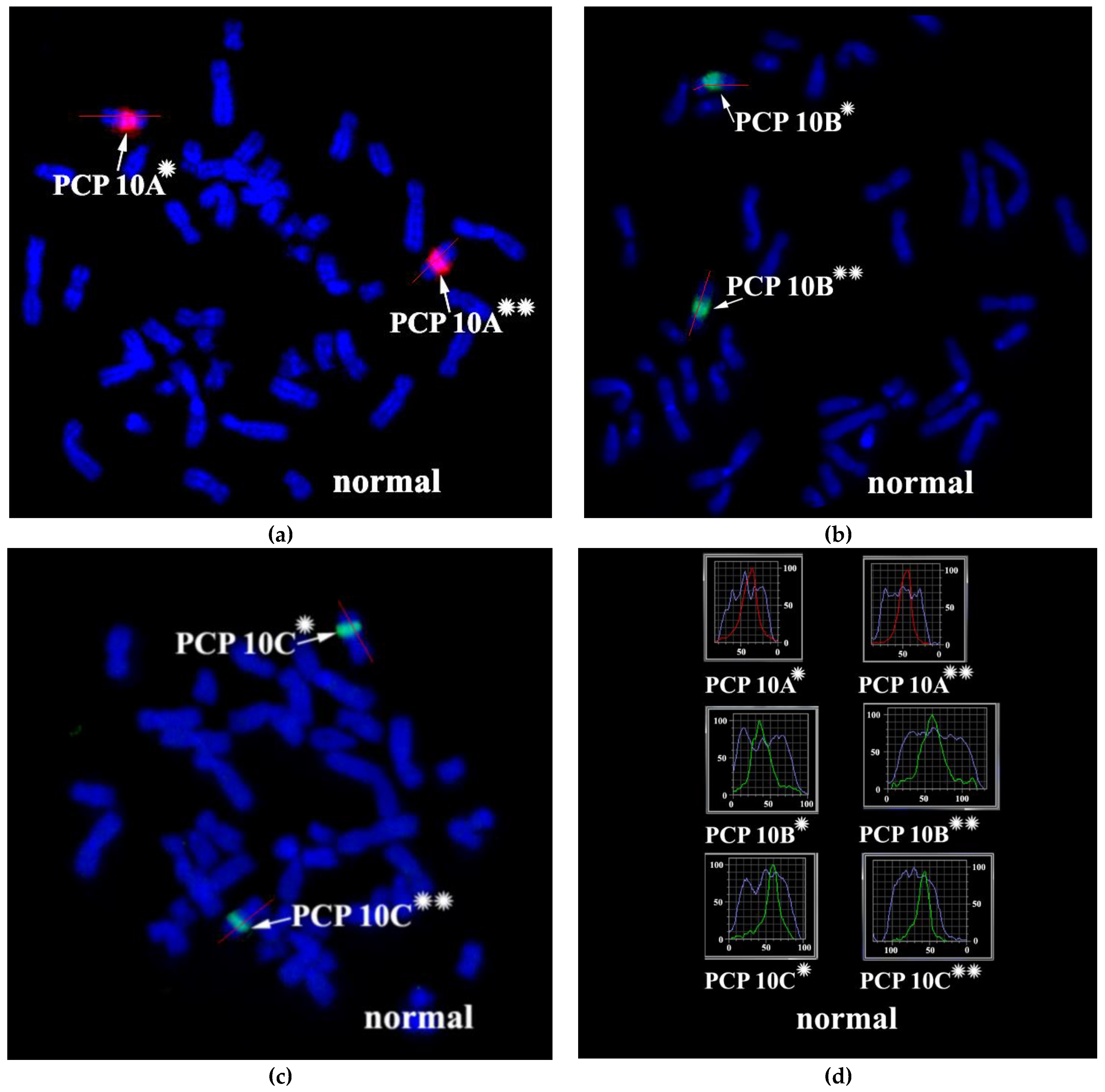
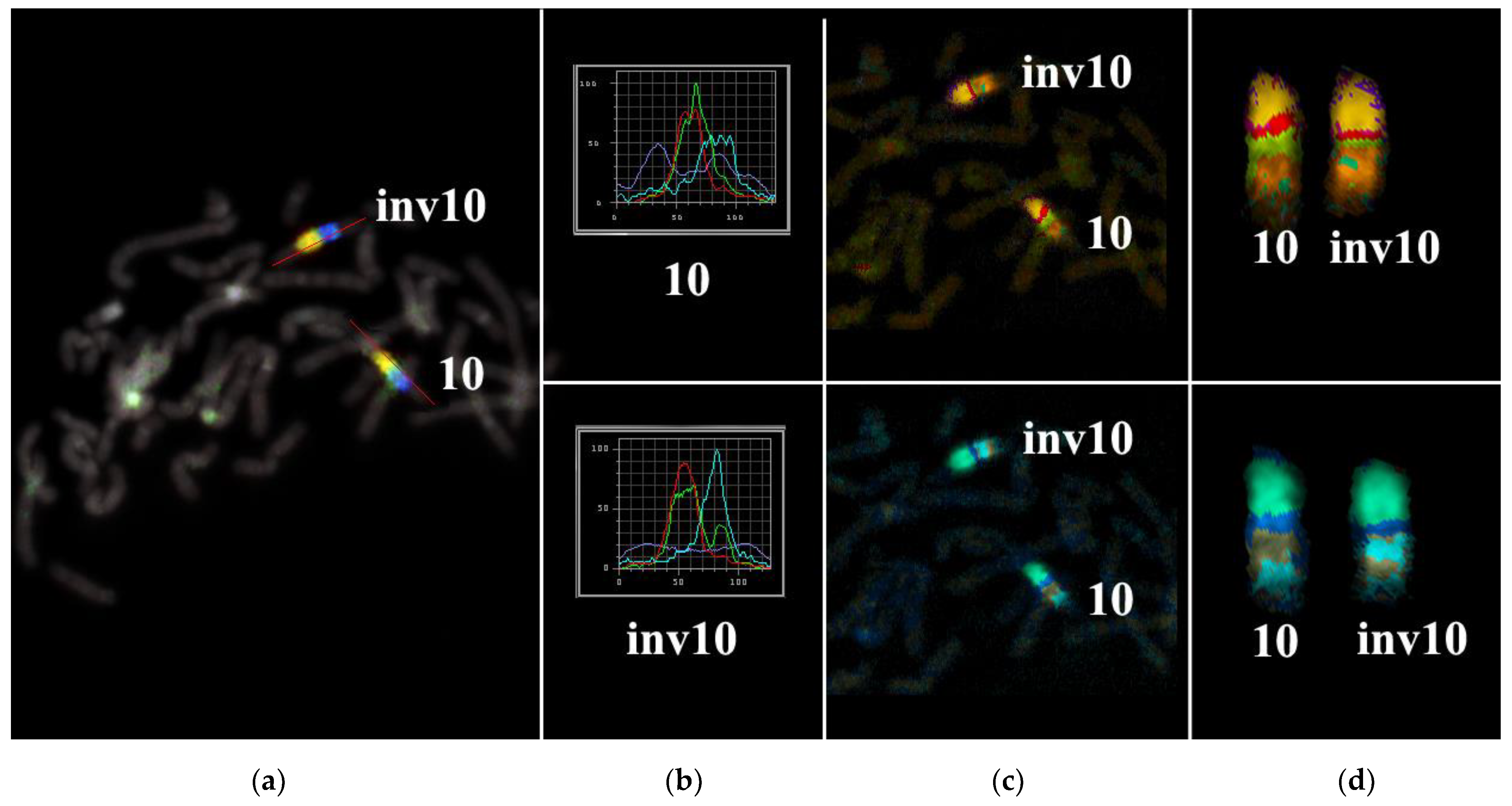
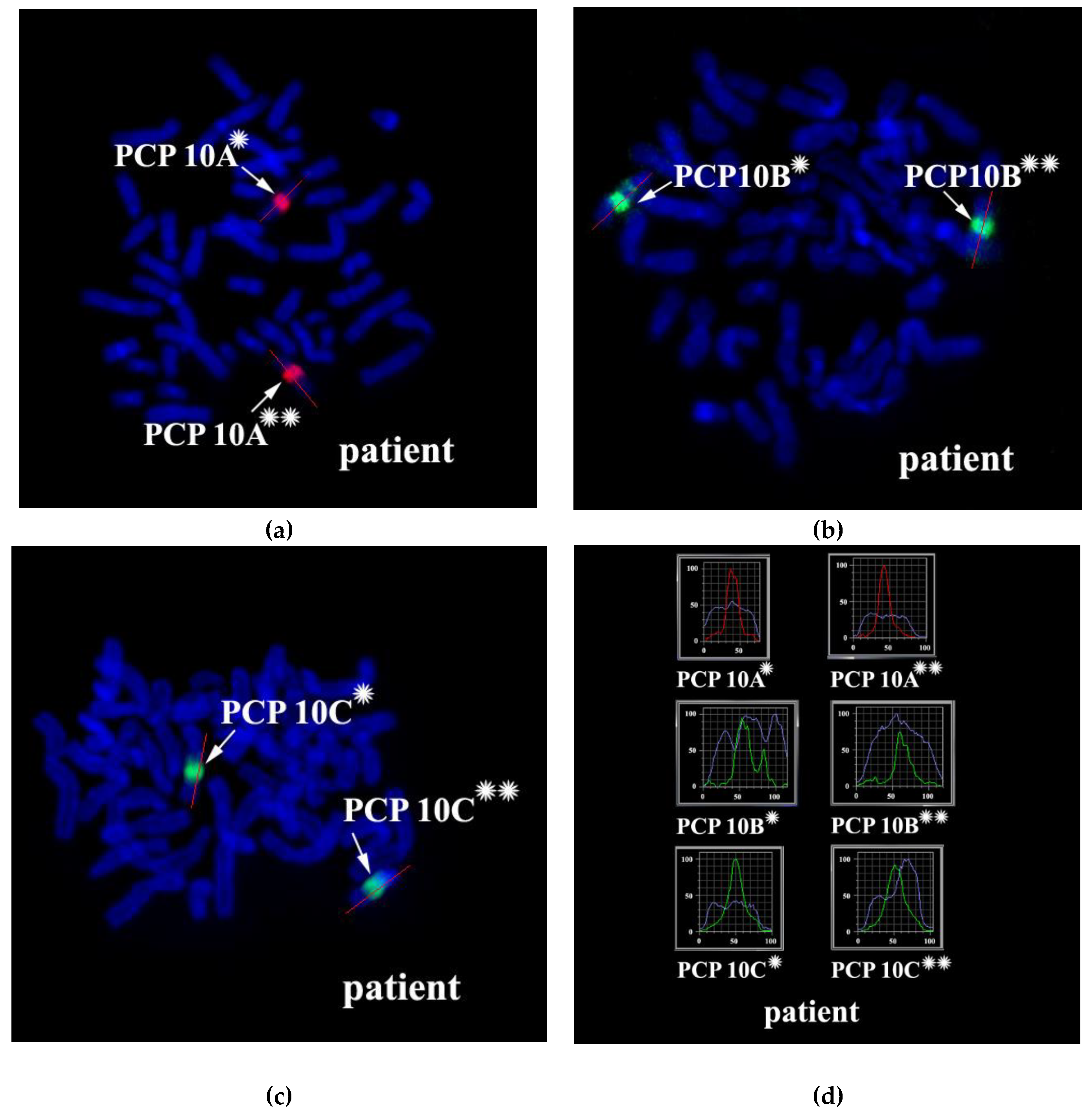
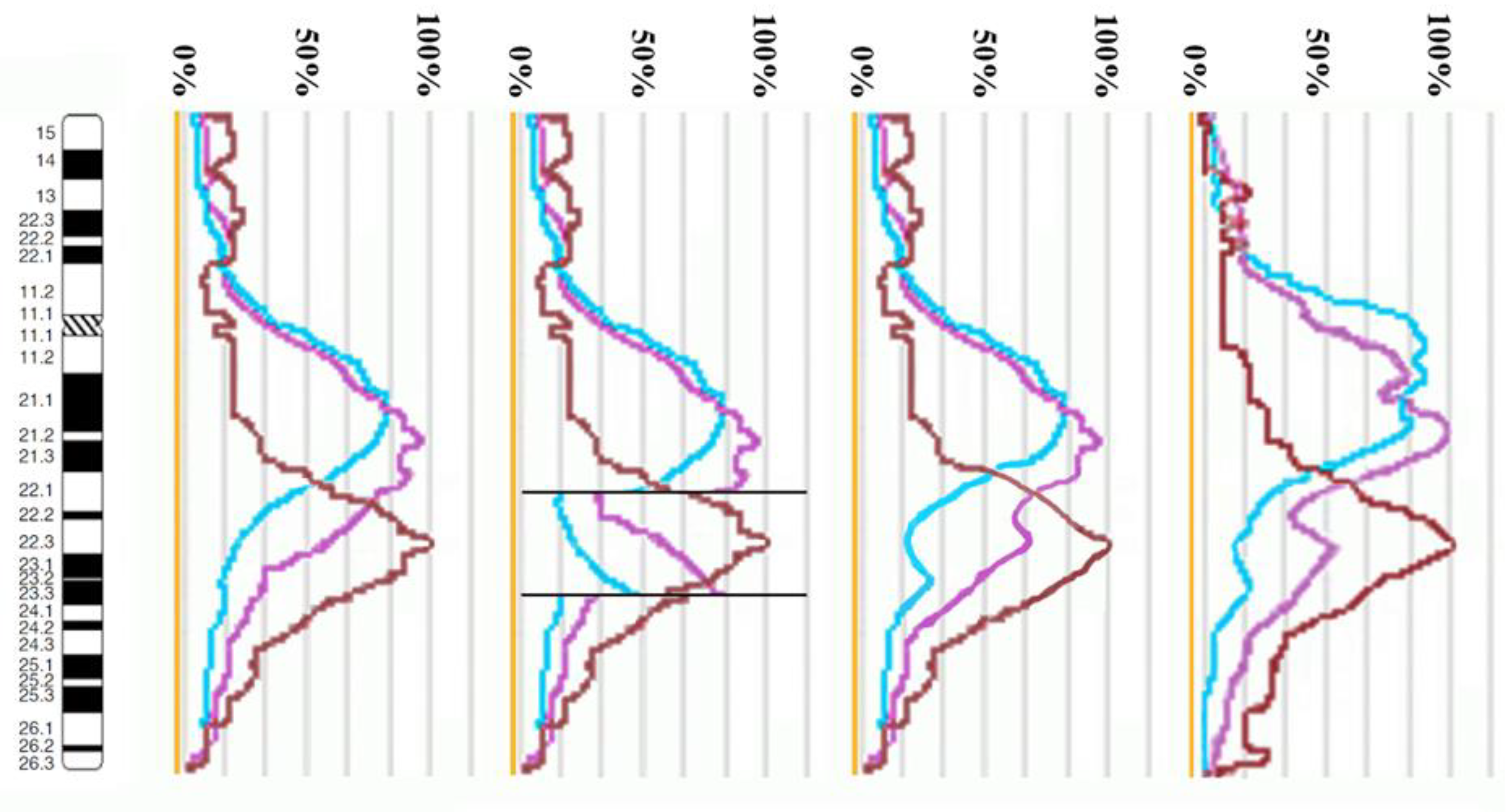
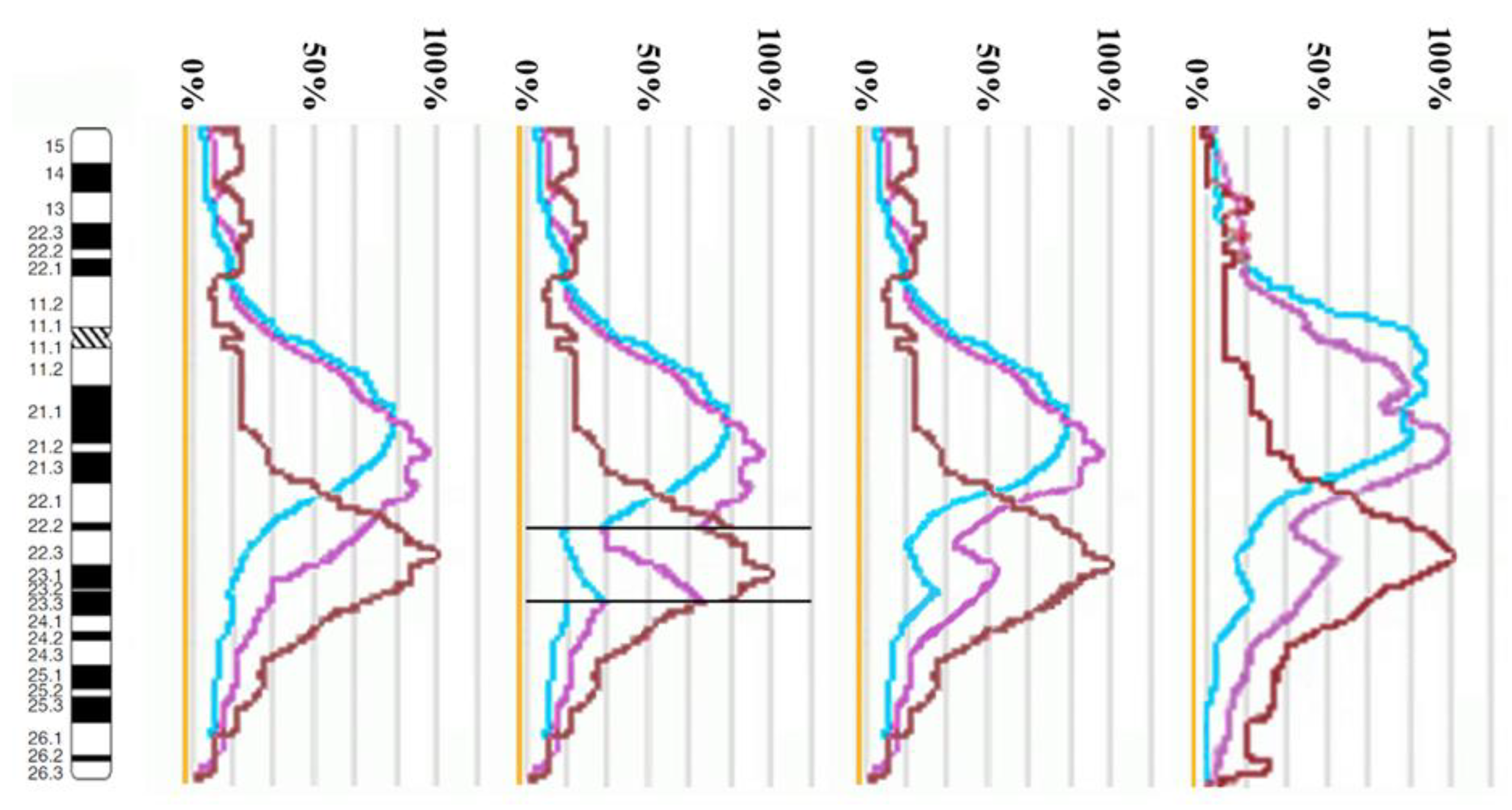
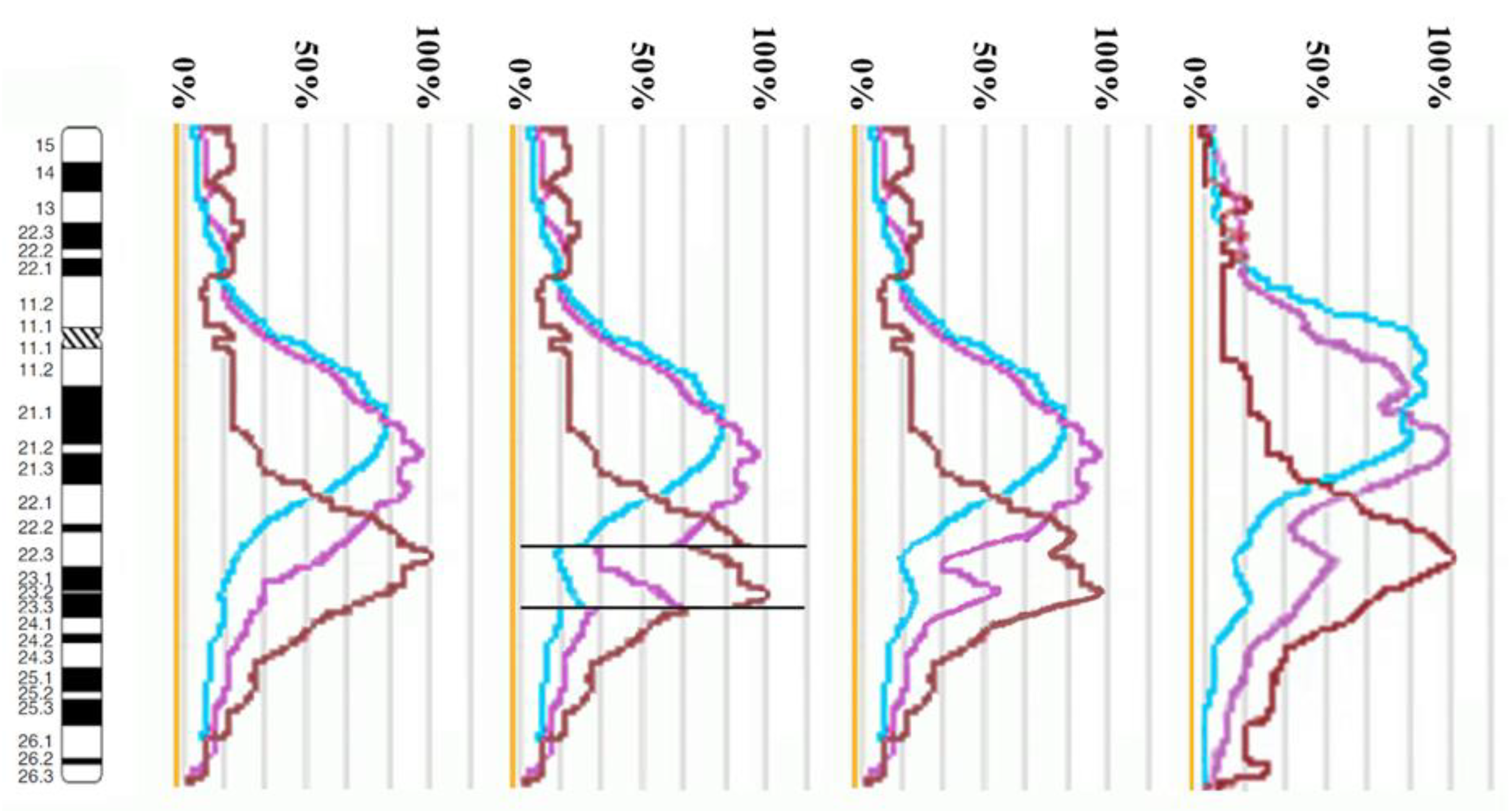
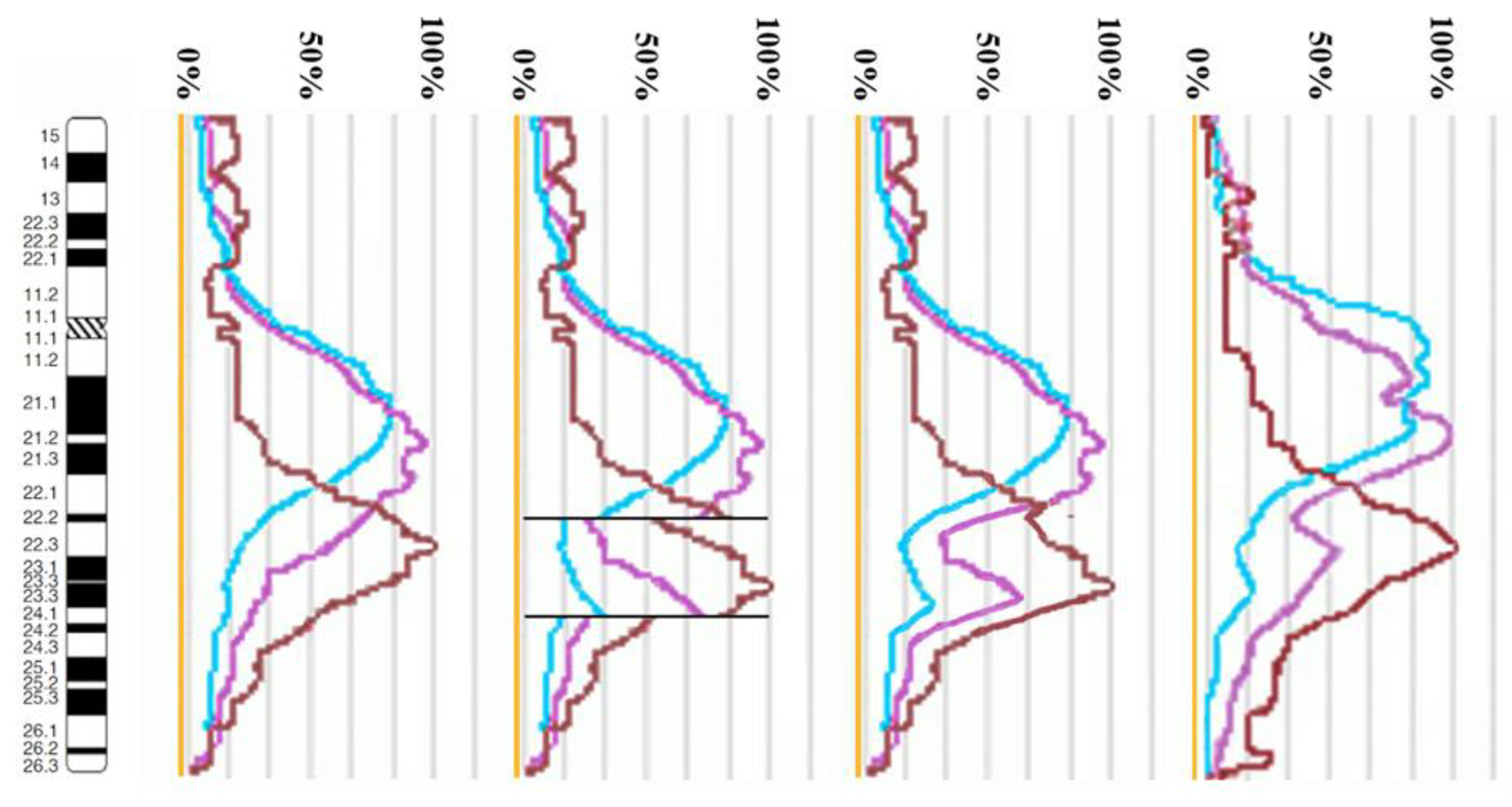
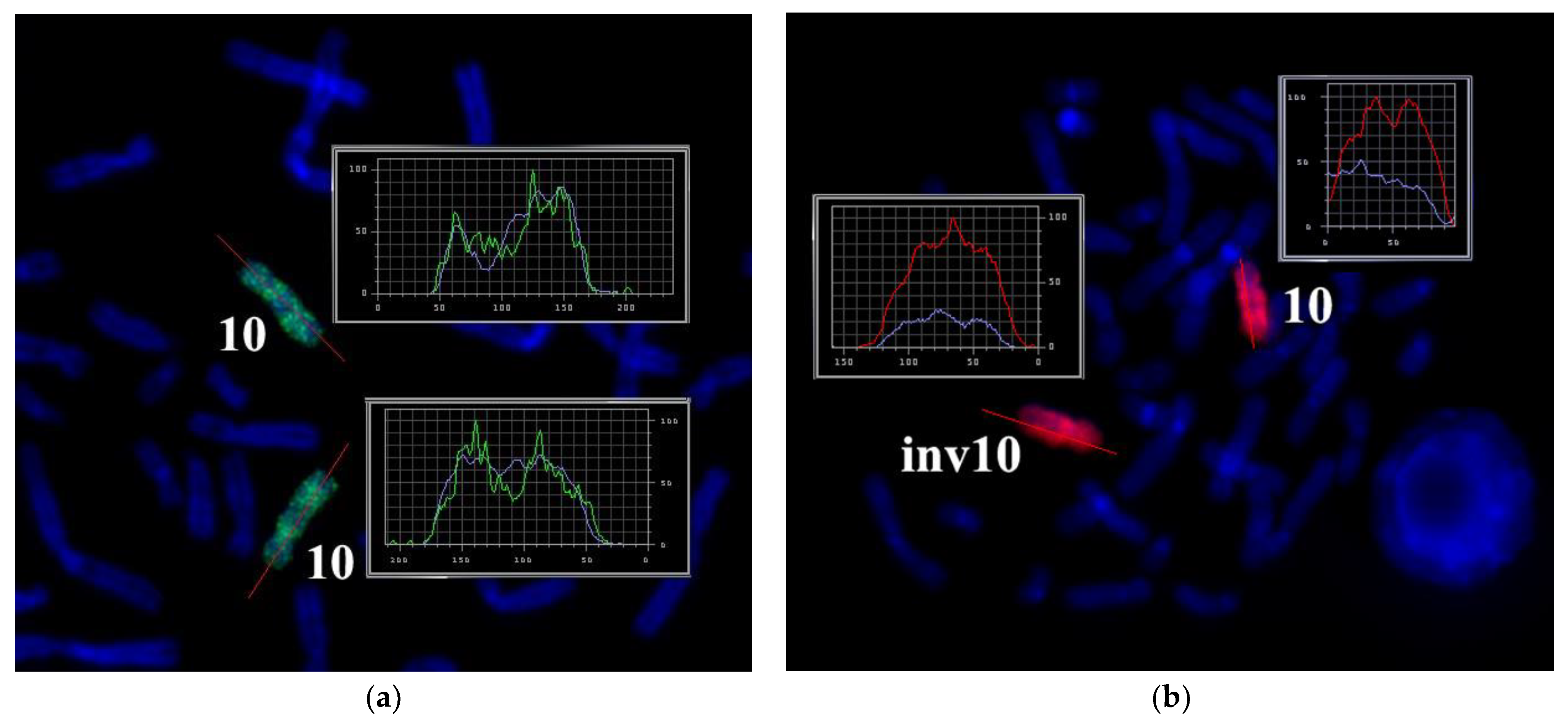
3.2. CISS Hybridization with PCPs and MCB Analysis
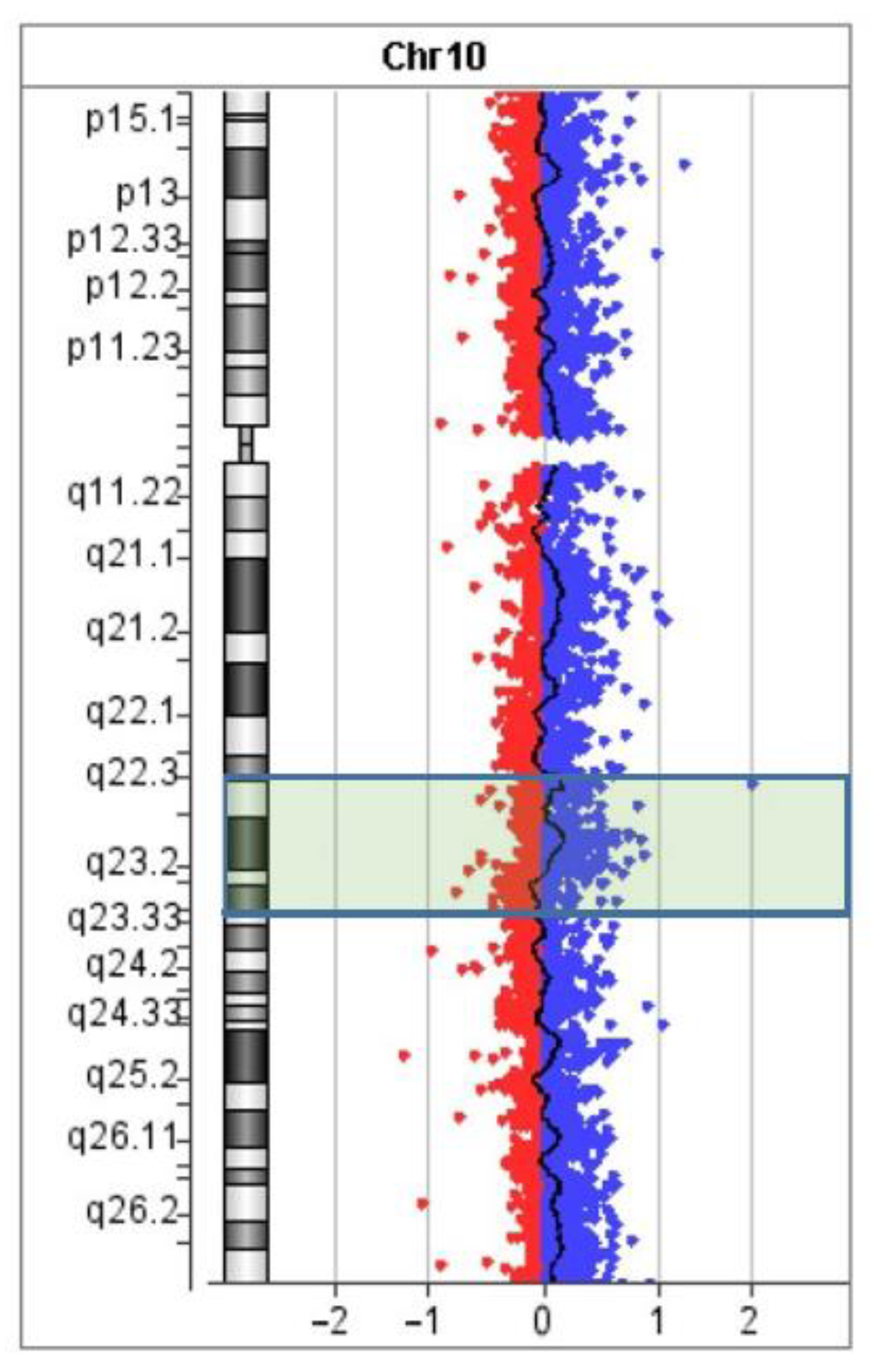
3.3. Looking for Small Microdeletions in the Breakpoint Regions of Inv(10)
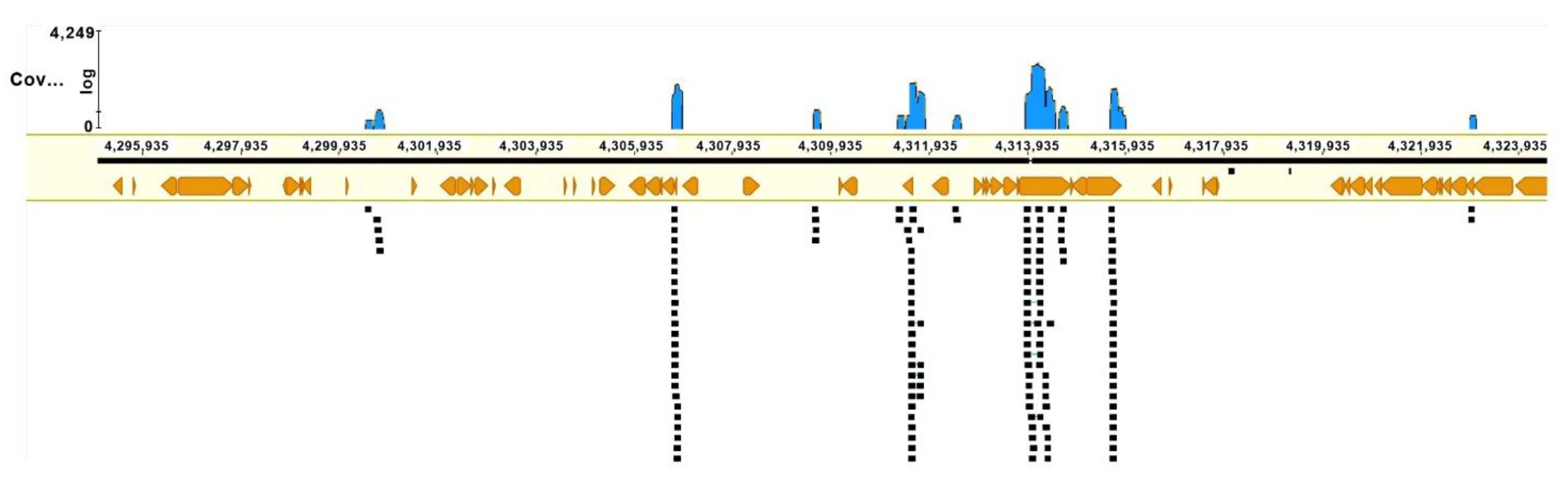
3.4. Microdissection and Single-Copy Chromosome Inv(10) Sequencing
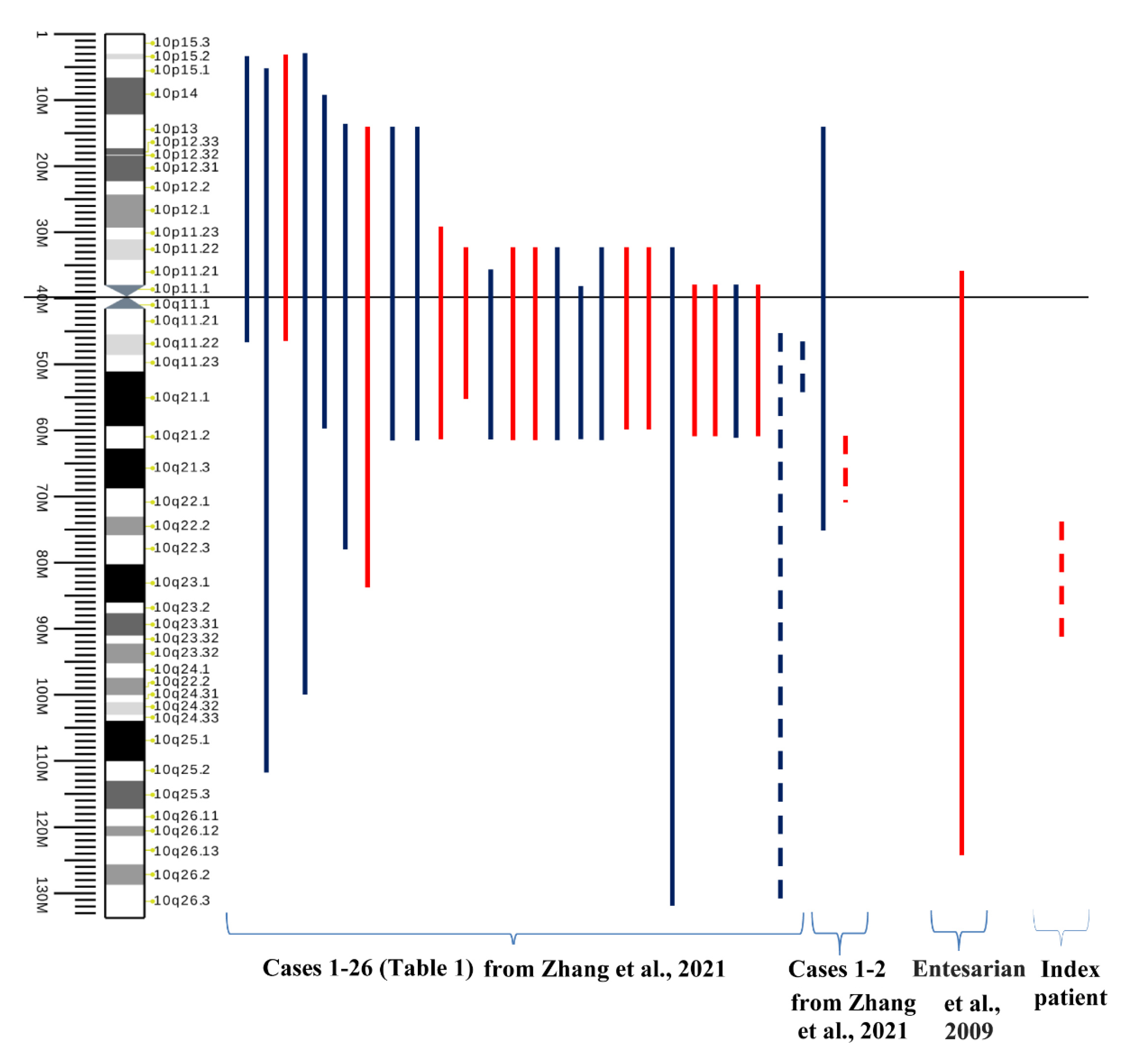
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muss, B.; Schwanitz, G. Characterization of Inversions as a Type of Structural Chromosome Aberration. Int. J. Hum. Genet. 2007, 7, 141–161. [Google Scholar] [CrossRef]
- Chien, C.-W.; Chao, A.-S.; Chang, Y.-L.; Chen, K.-J.; Peng, H.-H.; Lin, Y.-T.; Chao, A.; Chang, S.-D. Frequency and clinical significance of chromosomal inversions prenatally diagnosed by second trimester amniocentesis. Sci. Rep. 2022, 12, 2215. [Google Scholar] [CrossRef] [PubMed]
- Liehr, T.; Schreyer, I.; Kuechler, A.; Manolakos, E.; Singer, S.; Dufke, A.; Wilhelm, K.; Jančušková, T.; Čmejla, R.; Othman, M.A.K.; et al. Parental origin of deletions and duplications – about the necessity to check for cryptic inversions. Mol. Cytogenet. 2018, 11, 20. [Google Scholar] [CrossRef]
- Morel, F.; Laudier, B.; Guérif, F.; Couet, M.L.; Royère, D.; Roux, C.; Bresson, J.L.; Amice, V.; De Braekeleer, M.; Douet-Guilbert, N. Meiotic segregation analysis in spermatozoa of pericentric inversion carriers using fluorescence in-situ hybridization. Hum. Reprod. 2007, 22, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Morin, S.J.; Eccles, J.; Iturriaga, A.; Zimmerman, R.S. Translocations, inversions and other chromosome rearrangements. Fertil. Steril. 2017, 107, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-P.; Ko, T.-M.; Su, Y.-N.; Wang, L.-K.; Chern, S.-R.; Wu, P.-S.; Chen, Y.-N.; Chen, S.-W.; Ko, K.; Lee, C.-C.; et al. Prenatal diagnosis and molecular cytogenetic characterization of rec(10)dup(10p)inv(10)(p11.2q26.3) in a fetus associated with paternal pericentric inversion. Taiwan J. Obstet. Gynecol. 2016, 55, 733–737. [Google Scholar] [CrossRef]
- Nussbaum, R.L.; McInnes, R.R.; Willard, H.F.; Thompson, M.W.; Margaret, W.; Hamosh, A. Thompson & Thompson genetics in medicine, 7th ed.; Saunders/Elsevier: Philadelphia, PA, USA, 2007; ISBN 1416030808. [Google Scholar]
- Brugh, V.M.; Maduro, M.R.; Lamb, D.J. Genetic disorders and infertility. Urol. Clin. N. Am. 2003, 30, 143–152. [Google Scholar] [CrossRef]
- Stern, C.; Pertile, M.; Norris, H.; Hale, L.; Baker, H.W.G. Chromosome translocations in couples with in-vitro fertilization implantation failure. Hum. Reprod. 1999, 14, 2097–2101. [Google Scholar] [CrossRef]
- Gekas, J.; Thepot, F.; Turleau, C.; Siffroi, J.P.; Dadoune, J.P.; Briault, S.; Rio, M.; Bourouillou, G.; Carré-Pigeon, F.; Wasels, R.; et al. Chromosomal factors of infertility in candidate couples for ICSI: An equal risk of constitutional aberrations in women and men. Hum. Reprod. 2001, 16, 82–90. [Google Scholar] [CrossRef]
- Klepacka, D.; Young, D.; Brzeskiewicz, L.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Infertility patients with chromosome inversions are susceptible to an inter-chromosomal effect. Fertil. Steril. 2014, 102, e76. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization Press: Geneva, Switzerland, 2021. [Google Scholar]
- Ellis, P.M. Human Cytogenetics: A Practical Approach, Volume 1: Constitutional Analysis (second edition). Edited by D. E. Rooney and B. H. Czepulkowski. IRL Press at Oxford University Press. 1992. 274 pages. Paper £22.50. ISBN 0 19 963287 1. Spiralbound £30. ISBN 0 1. Genet. Res. 1993, 61, 152–153. [Google Scholar] [CrossRef]
- Verma, R.S.; Babu, A. Human Chromosomes: Manual of Basic Techniques; Pergamon Press: Elmsford, NY, USA, 1989. [Google Scholar]
- Karamysheva, T.B.; Matveeva, V.G.; Shorina, A.P.; Rubtsov, N.B. Clinical and Molecular Cytogenetic Analysis of a Rare Case of Mosaicism for Partial Trisomy 3p and Partial Trisomy 10q in Humans. Russ. J. Genet. 2001, 37, 811–816. [Google Scholar] [CrossRef]
- Rubtsov, N.; Senger, G.; Neumann, A.; Kelbova, C.; Junker, K.; Beensen, V.; Claussen, U.; Rubtsov, N.; Kuzcera, H. Interstitial deletion of chromosome 6q: Precise definition of the breakpoints by microdissection, DNA amplification, and reverse painting. Hum. Genet. 1996, 97, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, N.B.; Karamisheva, T.V.; Astakhova, N.M.; Liehr, T.; Claussen, U.; Zhdanova, N.S. Zoo-FISH with region-specific paints for mink chromosome 5q: Delineation of inter- and intrachromosomal rearrangements in human, pig, and fox. Cytogenet. Cell Genet. 2000, 268–270. [Google Scholar] [CrossRef]
- Lichter, P.; Cremer, T.; Borden, J.; Manuelidis, L.; Ward, D.C. Delineation of individual human chromosomes in metaphase and interphase cells by in situ suppression hybridization using recombinant DNA libraries. Hum. Genet. 1988, 80, 224–234. [Google Scholar] [CrossRef]
- McGowan-Jordan, J.; Hastings, R.J.; Moore, S. (Eds.) ISCN 2020; 2020; S.Karger AG: Basel, Switzerland, 2020; ISBN 978-3-318-06706-4. [Google Scholar]
- Liehr, T.; Weise, A.; Heller, A.; Starke, H.; Mrasek, K.; Kuechler, A.; Weier, H.-U.G.; Claussen, U. Multicolor chromosome banding (MCB) with YAC/BAC-based probes and region-specific microdissection DNA libraries. Cytogenet. Genome Res. 2002, 97, 43–50. [Google Scholar] [CrossRef]
- Chudoba, I.; Plesch, A.; Lörch, T.; Lemke, J.; Claussen, U.; Senger, G. High resolution multicolor-banding: A new technique for refined FISH analysis of human chromosomes. Cytogenet. Genome Res. 1999, 84, 156–160. [Google Scholar] [CrossRef]
- Liehr, T.; Heller, A.; Starke, H.; Rubtsov, N.; Trifonov, V.; Mrasek, K.; Weise, A.; Kuechler, A.; Claussen, U. Microdissection based high resolution multicolor banding for all 24 human chromosomes. Int. J. Mol. Med. 2002, 9, 335–339. [Google Scholar] [CrossRef]
- MacDonald, J.R.; Ziman, R.; Yuen, R.K.C.; Feuk, L.; Scherer, S.W. The Database of Genomic Variants: A curated collection of structural variation in the human genome. Nucleic Acids Res. 2014, 42, D986–D992. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Weise, A.; Heller, A.; Starke, H.; Mrasek, K.; Kuechler, A.; Pool-Zobel, B.L.; Claussen, U.; Liehr, T. Multitude multicolor chromosome banding (mMCB) – a comprehensive one-step multicolor FISH banding method. Cytogenet. Genome Res. 2003, 103, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, N.V.; Karamysheva, T.V.; Rubtsov, N.B. Clusters of Repetitive DNA Sequences in Chromosomes of Voles of the Subgenus Microtus (Microtus, Arvicolidae). Russ. J. Genet. 2019, 55, 1093–1102. [Google Scholar] [CrossRef]
- Sugaya, K. Chromosome instability caused by mutations in the genes involved in transcription and splicing. RNA Biol. 2019, 16, 1521–1525. [Google Scholar] [CrossRef]
- Puig, M.; Casillas, S.; Villatoro, S.; Cáceres, M. Human inversions and their functional consequences. Brief. Funct. Genom. 2015, 14, 369–379. [Google Scholar] [CrossRef]
- Zhang, F.; Khajavi, M.; Connolly, A.M.; Towne, C.F.; Batish, S.D.; Lupski, J.R. The DNA replication FoSTeS/MMBIR mechanism can generate genomic, genic and exonic complex rearrangements in humans. Nat. Genet. 2009, 41, 849–853. [Google Scholar] [CrossRef]
- Lam, H.Y.K.; Mu, X.J.; Stütz, A.M.; Tanzer, A.; Cayting, P.D.; Snyder, M.; Kim, P.M.; Korbel, J.O.; Gerstein, M.B. Nucleotide-resolution analysis of structural variants using BreakSeq and a breakpoint library. Nat. Biotechnol. 2010, 28, 47–55. [Google Scholar] [CrossRef]
- Kidd, J.M.; Graves, T.; Newman, T.L.; Fulton, R.; Hayden, H.S.; Malig, M.; Kallicki, J.; Kaul, R.; Wilson, R.K.; Eichler, E.E. A human genome structural variation sequencing resource reveals insights into mutational mechanisms. Cell 2010, 143, 837–847. [Google Scholar] [CrossRef]
- Pang, A.W.C.; Migita, O.; Macdonald, J.R.; Feuk, L.; Scherer, S.W. Mechanisms of Formation of Structural Variation in a Fully Sequenced Human Genome. Hum. Mutat. 2013, 34, 345–354. [Google Scholar] [CrossRef]
- Escaramís, G.; Docampo, E.; Rabionet, R. A decade of structural variants: Description, history and methods to detect structural variation. Brief. Funct. Genomics 2015, 14, 305–314. [Google Scholar] [CrossRef]
- Pettenati, M.J.; Rao, P.N.; Phelan, M.C.; Grass, F.; Rao, K.W.; Cosper, P.; Carroll, A.J.; Elder, F.; Smith, J.L.; Higgins, M.D.; et al. Paracentric inversions in humans: A review of 446 paracentric inversions with presentation of 120 new cases. Am. J. Med. Genet. 1995, 55, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Liehr, T.; Weise, A.; Mrasek, K.; Ziegler, M.; Padutsch, N.; Wilhelm, K.; Al-Rikabi, A. Recombinant Chromosomes Resulting From Parental Pericentric Inversions—Two New Cases and a Review of the Literature. Front. Genet. 2019, 10, 1165. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, I.N.; Karamysheva, T.V.; Elisaphenko, E.A.; Makunin, A.I.; Zhigalina, D.I.; Lopatkina, M.E.; Drozdov, G.V.; Cheremnykh, A.D.; Torkhova, N.B.; Seitova, G.N.; et al. Prenatal diagnosis of small supernumerary marker chromosome 10 by array-based comparative genomic hybridization and microdissected chromosome sequencing. Biomedicines 2021, 9, 1030. [Google Scholar] [CrossRef] [PubMed]
- Madan, K. Paracentric inversions: A review. Hum. Genet. 1995, 96, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, Q.; Liu, Y.; Jiang, Y.; Yang, X.; Liu, R.; Zhang, H. Fertility problems in males carrying an inversion of chromosome 10. Open Med. 2021, 16, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Entesarian, M.; Carlsson, B.; Mansouri, M.R.; Stattin, E.-L.; Holmberg, E.; Golovleva, I.; Stefansson, H.; Klar, J.; Dahl, N. A chromosome 10 variant with a 12 Mb inversion [inv(10)(q11.22q21.1)] identical by descent and frequent in the Swedish population. Am. J. Med. Genet. Part A 2009, 149A, 380–386. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
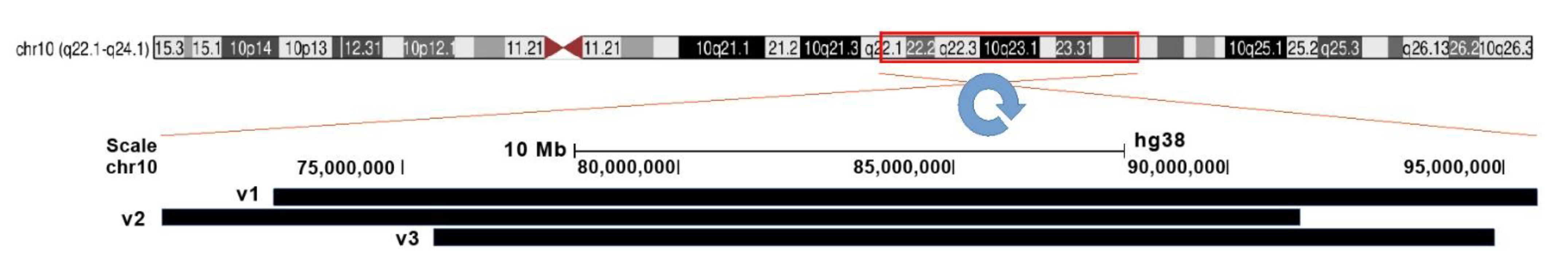
| No. | Start | End | Length | Bands | Repeat Family |
|---|---|---|---|---|---|
| 1 | 72675611 | 95619369 | 22943758 | 10q22.1–10q23.33 | L1P3 |
| 2 | 70631615 | 91322886 | 20691271 | 10q22.1–10q23.31 | SVA_D |
| 3 | 75577762 | 94828954 | 19251192 | 10q22.2–10q23.33 | L1P3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karamysheva, T.V.; Gayner, T.A.; Elisaphenko, E.A.; Trifonov, V.A.; Zakirova, E.G.; Orishchenko, K.E.; Prokhorovich, M.A.; Lopatkina, M.E.; Skryabin, N.A.; Lebedev, I.N.; et al. The Precise Breakpoint Mapping in Paracentric Inversion 10q22.2q23.3 by Comprehensive Cytogenomic Analysis, Multicolor Banding, and Single-Copy Chromosome Sequencing. Biomedicines 2022, 10, 3255. https://doi.org/10.3390/biomedicines10123255
Karamysheva TV, Gayner TA, Elisaphenko EA, Trifonov VA, Zakirova EG, Orishchenko KE, Prokhorovich MA, Lopatkina ME, Skryabin NA, Lebedev IN, et al. The Precise Breakpoint Mapping in Paracentric Inversion 10q22.2q23.3 by Comprehensive Cytogenomic Analysis, Multicolor Banding, and Single-Copy Chromosome Sequencing. Biomedicines. 2022; 10(12):3255. https://doi.org/10.3390/biomedicines10123255
Chicago/Turabian StyleKaramysheva, Tatyana V., Tatyana A. Gayner, Eugeny A. Elisaphenko, Vladimir A. Trifonov, Elvira G. Zakirova, Konstantin E. Orishchenko, Mariya A. Prokhorovich, Maria E. Lopatkina, Nikolay A. Skryabin, Igor N. Lebedev, and et al. 2022. "The Precise Breakpoint Mapping in Paracentric Inversion 10q22.2q23.3 by Comprehensive Cytogenomic Analysis, Multicolor Banding, and Single-Copy Chromosome Sequencing" Biomedicines 10, no. 12: 3255. https://doi.org/10.3390/biomedicines10123255
APA StyleKaramysheva, T. V., Gayner, T. A., Elisaphenko, E. A., Trifonov, V. A., Zakirova, E. G., Orishchenko, K. E., Prokhorovich, M. A., Lopatkina, M. E., Skryabin, N. A., Lebedev, I. N., & Rubtsov, N. B. (2022). The Precise Breakpoint Mapping in Paracentric Inversion 10q22.2q23.3 by Comprehensive Cytogenomic Analysis, Multicolor Banding, and Single-Copy Chromosome Sequencing. Biomedicines, 10(12), 3255. https://doi.org/10.3390/biomedicines10123255